
Abstract
Hepatic differentiation of embryonic stem cells (ESCs) usually results in a single cell lineage, and the formation of liver tissues remains difficult. Here, we examine the role of epithelial-mesenchymal transition (EMT) that is regulated by epithelial cadherin (E-cad) expression in hepatic tissue formation from ESCs. E-cad was transfected into mouse ESCs to enable a stable expression of E-cad. Hepatic differentiation of ESCs was then induced by hepatic growth factors. Wnt/β-catenin signaling and EMT speed were examined to determine the differentiation process. Hepatic and angiogenesis markers, as well as differentiated cell-adhesive force were also examined to identify the hepatic tissue differentiation. In our results, E-cad expression gradually decreased in normal ESC (N-ESC) differentiation, but remained stable in the E-cad transfected ESC (EC-ESC) group. In EC-ESC differentiation, expressions of cytoplastic β-catenin and EMT were much lower and significantly prolonged. Angiogenesis markers vascular endothelial growth factor receptor-1 (VEGFR-1) and CD31/PECAM-1 were expressed only on day 5–13 in N-ESC differentiation, whereas VEGFR-1 and CD31/PECAM-1 were expressed prolonged on day 5–17 in the EC-ESC group and were coincident with the expression of hepatic markers. Finally, EC-ESC differentiation maintained multilayer-growth patterns, and abundant vascular network structures appeared and migrated in albumin-positive cell areas. The cellular adhesion forces between embryonic body cells in EC-ESC differentiation during day 13–17 were similar to those of mouse liver tissue. In conclusion, accelerated EMT due to the decreased E-cad expression may partially contribute to the failure of hepatic tissue formation in N-ESC differentiation. E-cad can act in synergy with hepatic growth factors and facilitate the early-stage formation of hepatic tissues through down-regulating Wnt/β-catenin signaling and delaying EMT. This work provides a new insight into hepatic tissue differentiation that is mediated by E-cad from ESC.
Introduction
W
During hepatic differentiation of ESCs in vitro, ESCs differentiate from clone into scatter [3]. In the early stage, ESC differentiates into an embryonic body (EB), which is composed of endodermal, mesodermal, and ectodermal germ layers and has a three-dimensional (3D) structure. However, in the late stage, EBs rapidly differentiate into loose monolayer cells and lose tight 3D structures [4]. Simultaneously, the cellular adhesive molecular-epithelial cadherin (E-cad) expression decreased in ESC differentiation and the cell adhesion became weak [5]. These morphologic changes and decreased E-cad expression suggested that ESC underwent an accelerated epithelial-mesenchymal transition (EMT) process, which may result in the difficulty of in vitro liver-tissue formation. EMT was identified as an early and essential step in embryo tissue development and ESC differentiation, which makes epiblast cells facilitate their ingression within the primitive streak through losing epithelial features and acquisition of mesenchymal characteristics [6,7]. During EMT, embryo epiblasts give rise to definitive endodermal and mesodermal cells and then form various kinds of tissue [6,7]. While a moderate level of EMT is helpful for tissue development, an accelerated EMT could lead to inhibited tissue formation, because EMT may produce too many nonpolarized mesenchymal cells, and EMT is usually associated with a lack of intercellular junctions. Thus, accelerated EMT usually leads to failure of formation of the regular and tight tissue structures, for example, in the tumorigenesis [8]. Therefore, accelerated EMT in ESC differentiation in vitro than that in embryogenesis in vivo may inhibit the tissue formation from ESCs.
A defining characteristic of EMT is the loss of cell–cell contacts that are mediated by E-cad and the acquisition of a mesenchymal phenotype [9]. As a calcium-dependent cell adhesion molecule, E-cad plays a key role in regulating the structural development of epithelial tissues, including liver tissue, by mediating epithelial cell adhesions [10 –12]. In addition, as a transmembrane constituent of intercellular adherens junctions, E-cad also binds to membrane β-catenin, decreases the cytoplasmic dissociative β-catenin, and down-regulates Wnt/β-catenin signaling [13,14], a pathway inducing EMT in embryos and stem cell differentiation [15,16]. Therefore, E-cad may decelerate EMT in ESC differentiation to be an appropriate speed and then help regulate tissue development by regulating cell adhesion and Wnt/β-catenin pathway [17,18].
To date, it is still unclear as to whether E-cad could slow down EMT and help regulate hepatic tissue development during ESC differentiation. In addition, the role of Wnt/β-catenin and EMT pathway in ESCs differentiation regulated by E-cad remains to be clarified. Hepatic cells and biliary epithelial cells (BECs) have been simultaneously induced by us from ESCs. However, both hepatic cells and BECs differentiated from ESCs were still in the single-cell state, and it is difficult to induce the formation of hepatic tissues [19,20]. Since E-cad may regulate EMT speed in ESC differentiation and help promote tissue development by regulating cell adhesion and the Wnt/β-catenin pathway, we hypothesize that E-cad can regulate the EMT and mediate hepatic tissue differentiation from ESC.
To address this hypothesis, exogenous E-cad was transfected into ESCs to ensure its relatively stable expression during hepatic differentiation. Then, E-cad transfected ESC (EC-ESC) were induced to hepatic differentiation, as previously described [19,20]. During this process, we found that E-cad transfection could slow down the accelerated EMT and promote tissue formation during in vitro hepatic differentiation from ESCs.
Materials and Methods
Culture and hepatic differentiation of ESCs
ESCs of BALB/c mice and 129 mice were established and preserved by Professor Huang Bing of the Ophthalmic Center of Sun Yat-Sen University. Basic culture agents include Dulbecco's-modified Eagle's medium (Gibco, Bra), 20% fetal bovine serum (Hyclone), 0.1 M 2-mercaptoethanol, 25 mM HEPES, 100 U/mL penicillin, 100 μg/mL streptomycin, and 1,000 U/mL mouse leukemia inhibitory factor (Gibco, Bra). After suspension culturing for 5 days, ESCs developed into EBs, and EBs were then plated onto a culture dish that was coated with 10 μg/cm2 type I collagen. In subsequent experiments, several growth factors were added into culture medium for hepatic differentiation: 100 ng/mL acidic fibroblast growth factor (aFGF), 20 ng/mL transforming growth factor-α (TGFα), 20 ng/mL hepatocyte growth factor (HGF), 20 ng/mL epidermal growth factor (EGF), 10 ng/mL oncostatin M (OSM; Sigma), 10−7 M dexamethasone (DEX), 5 mg/mL insulin, 5 mg/mL transferrin, and 5 mg/mL selenious acid (Gibco BRL). These growth factors were added to the culture medium at different time points: aFGF was added to EBs from day 7 to 11, TGFα was added from day 7 to 19 as early-stage factors; EGF and HGF were added from day 11 to 19 as middle-stage factors; and OSM, DEX, insulin, transferrin, and selenious acid were added from day 15 to 19 as late-stage factors [19,20].
Transfection of E-cad gene to ESCs
Eukaryotic cells pCAG-MCS (Yrbio) that contain the CAG promoter were selected. FuGENE HD (Roche) was used for plasmid transfection. The PCR primers were designed according to the full cDNA sequences of mice E-cad in GeneBank: (5′-GGAAGCTTATGGGAGCCCGGTGC-3′, 5′-GCGGATCCCTAGTCGTCCTCACCA-3′). The whole RNA of liver tissue of BALB/c was purified, then reverse transcribed into cDNA and the target gene was subjected to PCR amplification. The products were subcloned into a vector of pCAG-MCS to obtain the recombinant pCAG-E-cad plasmid; pCAG-E-cad was then transfected into ESCs with FuGENE HD, as specified by the supplier. Resistant clones were obtained from stable transfection with 2 weeks of G418 (300 μg/mL) selection. The successfully transfected ESCs were designated as “EC-ESCs.”
Real-time PCR
Total RNA was isolated from EBs cells using TRIzol® Reagent (Invitrogen), and cDNA was synthesized using a High-Capacity RNA-to-cDNA Kit (Applied Biosystems). DNA templates were amplified using Fast SYBR® Green Master Mix (Applied Biosystems) on the 7900HT Fast Real-Time PCR System (Applied Biosystems). β-actin was used as an internal control. Experiments were performed in triplicate for each data point. The primer synthesis was provided by SANGON. Sequences of the primers are listed in Supplementary Table S1 (Supplementary Data are available online at
Western blot experiment
EBs cells were lysed using lysis buffer and mixed. The supernatant liquid was collected after centrifugation at 12,000 rpm for 10 min under 4°C, and all of the cellular protein was extracted. Cytosol protein was collected using a Cytosol Protein Extraction Kit (Beyotime). Electrophoretic separation of 10 μg protein per channel was conducted using 8% polyacrylamide gel. The protein was transferred onto a polyvinylidene fluoride (PVDF) membrane by electroblotting. An anti-blocking reaction (antibody concentration of 1:200–1:1,000) after blocking with PBS-5%, nonfat dry milk-0.1% Tween-20; secondary antibody IgG-HRP was used to combine with the primary antibody. The enhanced light-emitting agent and oxidizer were then mixed, and PVDF membrane was incubated for 3 min and then exposed under X-rays for 1–5 min to visualize the target protein. The antibodies used included rabbit anti-mouse E-cad, β-catenin, β-actin, OCT-4, Snail, Vimentin, Cytokeratin-19 (CK19), vascular endothelial growth factor receptor-1 (VEGFR-1), goat anti-rabbit IgG-HRP, rabbit anti-goat IgG-HRP (Cell Signaling Technology), goat anti-mouse albumin (ALB), Brachyury, tyrosine aminotransferase (TAT), cholesterol 7α-hydroxylase (CYP7a1), γ-glutamyltransferase (GGT), and platelet endothelial cell adhesion molecule-1 (PECAM-1/CD31) (Santa Cruz Biotech).
Immunocytochemistry
The EBs started differentiation on the 24-well plate on day 5. Immunocytochemistry (ICC) tests were undertaken on days 5, 9, 13, and 17 using the indirect immunofluorescence detection method. The antibodies used were as follows: rabbit anti-mouse E-cad, ALB, CK19, VEGFR-1 (DAKO), goat anti-mouse CD31/PECAM-1 (Santa Cruz Biotech), goat anti-rabbit IgG, rabbit anti-goat IgG, and immunofluorescence kit (BOSTER). A fluorescent inverted phase-contrast microscope (Olympus IX70-S8F2) was used.
Adhesive force between differentiating cells measured by micropipette suction systems
The microtubule system is based on the balance between negative pressure when a microvacuum tube is used on the cells and cell adhesion force, including a glass micropipette (inner diameter from 2.12 to 3.03 μm), an inverted microscope (Axiovert 35; Zeiss Corp.), a micromanipulator (MR5170; Eppendorf Corp.), a negative pressure governor, video recording systems (VTG For A Corp.), and image processing devices (Vidas21; Kontron Corp.). The changes in adhesion force between the differentiated cells were measured on days 5, 9, 13, and 17. Mice fetal liver tissue was used as the positive control.
Statistical analysis
Statistical analyses were performed using SPSS 17.0 software (SPSS, Inc.). The mean levels of genes expression and cell adhesive force (AF) values in each group were expressed as means±standard error. P values<0.05 were considered statistically significant. The Bonferroni correction was used with the t-test.
Results
E-cad expression is associated with down-regulation of Wnt/β-catenin signaling during in vitro ESC differentiation
To overexpress E-cad in ESCs in vitro, we use two ESCs from BALB/c and 129 mice. The qPCR and western blot results showed a significantly higher expression of E-cad in both EC-ESC than normal ESC (N-ESC) (Supplementary Fig. S1). Since more significant increases in E-cad were observed in BALB/c EC-ESCs, we used BALB/c EC-ESCs and N-ESCs for the following experiments. As an initial step, to determine the role of E-cad in EMT with regard to ESC differentiation, we detected dynamic expression levels of E-cad in differentiated cells. In N-ESC differentiation, E-cad was strongly expressed from day 1 to 5 and then gradually decreased to no expression on day 17. In EC-ESC differentiation, E-cad expression was elevated in comparison to the N-ESC group and was still observable on day 17 (Fig. 1A, B).
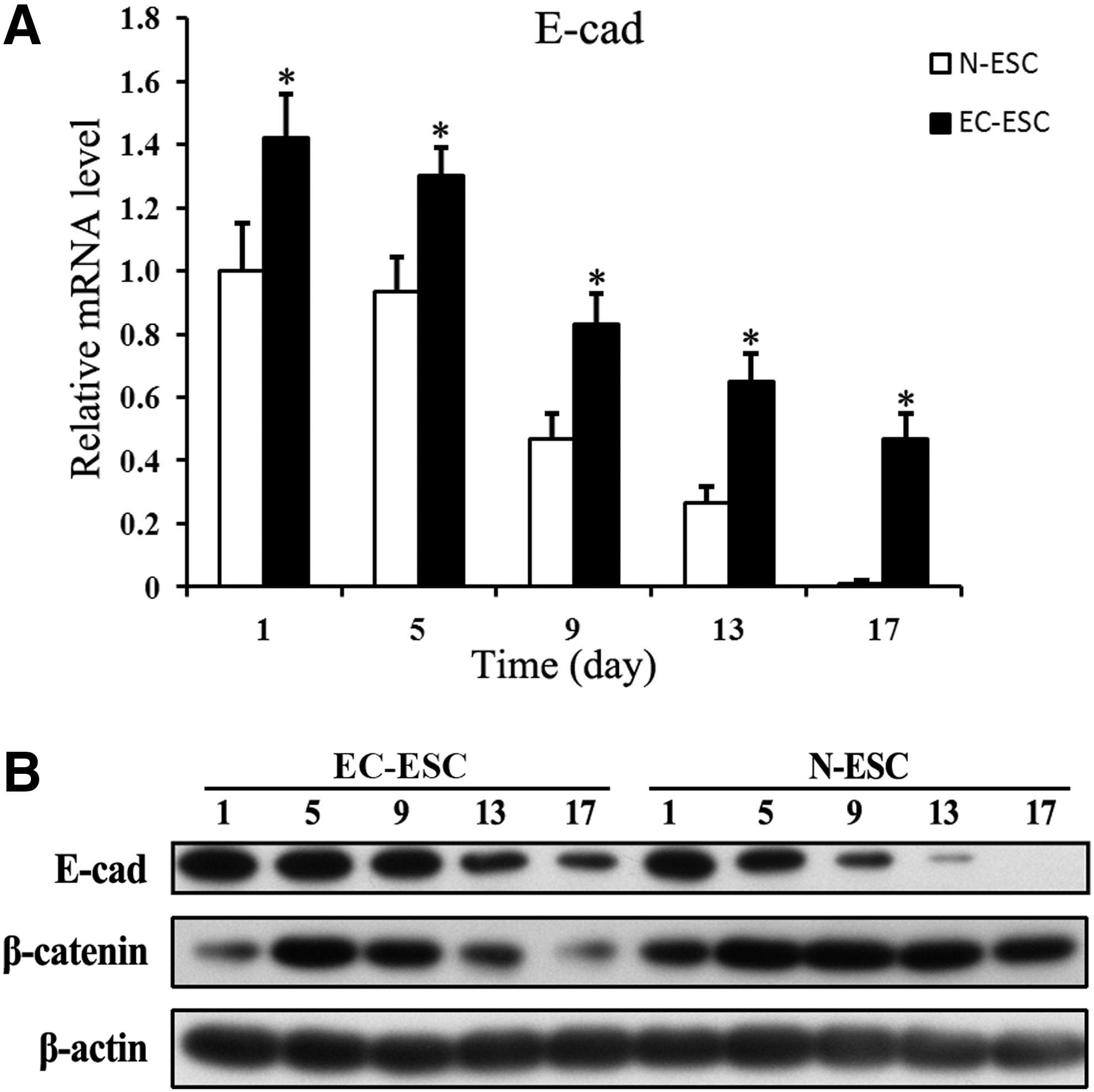
E-cad stable expression down-regulates Wnt/β-catenin pathway. Cells were harvested from N-ESC or EC-ESC at the indicated time points.
We then examined the effect of E-cad transfection on the Wnt/β-catenin pathway. Wnt/β-cateinin signaling plays an important role in inducing EMT in embryogenesis and ESC differentiation [16]. The increased E-cad expression may sequester more β-catenin at cell junctions and result in less cytoplastic β-catenin, leading to the down-regulation of Wnt/β-catenin signaling. In our result, in N-ESC differentiation, cytoplastic β-catenin expression increased from day 1 to 9 and then decreased from day 9 to 17. However, in the EC-ESC group, cytoplastic β-catenin expression increased from day 1 to 5 and then decreased. Thus, expression levels of cytoplastic β-catenin in EC-ESC differentiation were much lower than those in N-ESC (Fig. 1B). These data suggested that stable expression of E-cad is associated with inhibition of cytoplastic β-catenin and down-regulation of Wnt/β-catenin signaling, which may, therefore, contribute toward decelerating EMT in ESC differentiation.
E-cad delayed EMT in ESC differentiation process
We assessed whether E-cad could regulate EMT in ESC differentiation. EC-ESC differentiation and EMT process were first studied by detecting the expression of Oct-4, an important marker for pluripotency and an undifferentiation state of ESC [17]. In N-ESC differentiation, the expression of Oct-4 was observed from day 1 to 9. In the EC-ESC group, the expression of Oct-4 was observed from day 1 to 13, and the expression level of Oct-4 in EC-ESC was much higher than that in the N-ESC group (Fig. 2A). Western blot also showed a higher expression of Oct-4 in EC-ESC on day 13 (Supplementary Fig. S2). These data suggested that E-cad might contribute to the maintenance of the pluripotency of ESCs and the inhibition of EMT.
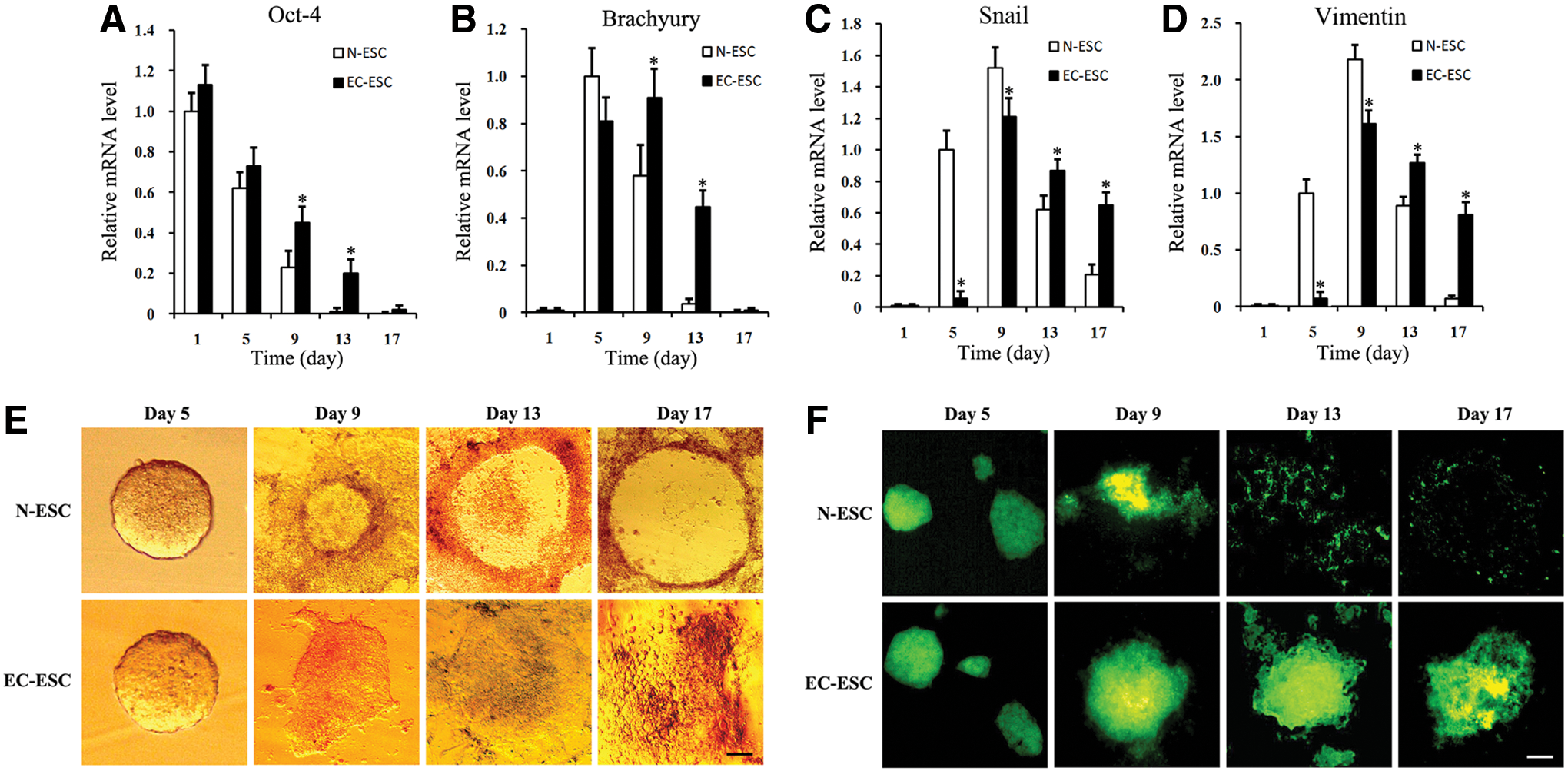
E-cad stable expression delays EMT and ESC differentiation in vitro. Cells were harvested from N-ESC or EC-ESC at the indicated time points.
Accumulating evidence has suggested that Brachyury, Snail, and Vimentin may facilitate the EMT, and the expression of Brachyury, Snail, and Vimentin represents gastrulation and mesoderm appearing in ESC differentiation [21 –23]. To further confirm that E-cad overexpression and down-regulation of Wnt/β-catenin are associated with the EMT in ESC differentiation, we also examined the expression of Brachyury, Snail, and Vimentin. We found that Brachyury was expressed during day 5–9 in N-ESC differentiation, and the expression of Brachyury was observed during day 5–13 in the EC-ESC group. This suggests that E-cad transfection prolonged the expression of Brachyury. In addition, the expression level of Brachyury on day 5 was much lower in EC-ESC differentiation than that in the N-ESC group. Similar prolonged expression of Snail and Vimentin such as Brachyury was also observed (Fig. 2B–D). We also detected these three proteins (Brachyury, Snail, and Vimentin) by western blot. It showed that the expression of Brachyury, Snail, and Vimentin is higher and stable in EC-ESC than in N-ESC. These results were in accordance with qPCR (Supplementary Fig. S2). The prolonged expression of Brachyury, Snail, and Vimentin suggested that EMT and the in vitro ESC differentiation process have been delayed by E-cad expression.
Our morphological observation also suggested that E-cad could slow down the EMT process in the differentiation of ESCs (Fig. 2E, F). After attaching the culture dish to differentiation, E-cad transfected EBs (EC-EBs) showed strong adhesion during differentiation and still maintained 3D structures on day 17, while normal EBs gradually spread from the multilayer structure into the monolayer (Fig. 2E). E-cad protein expression was also much higher in 3D structures of EC-EBs than that in N-ESCs group on days 9, 13, and 17 (Fig. 2F). These data further suggested that E-cad delayed EMT in ESC differentiation.
E-cad enhances the expression of liver-specific genes and proteins
Since we have observed that E-cad might delay EMT in ESC differentiation, we further examined whether E-cad can enhance hepatic differentiation. We detected the expression of a group of hepatic genes, including ALB, TAT, CYP7a1, GGT, and CK19 by Real-time PCR [24 –26]. In EC-ESC differentiation, the expression of these hepatic genes is much higher or earlier than that in the N-ESC group (Fig. 3A–E). In western Blot result, the dynamic expression of these hepatic proteins is consistent with their mRNA expression in ESC differentiation (Fig. 3F). These data collectively suggest that E-cad may facilitate the expression of hepatic markers with the incorporation of additional hepatic growth factors.
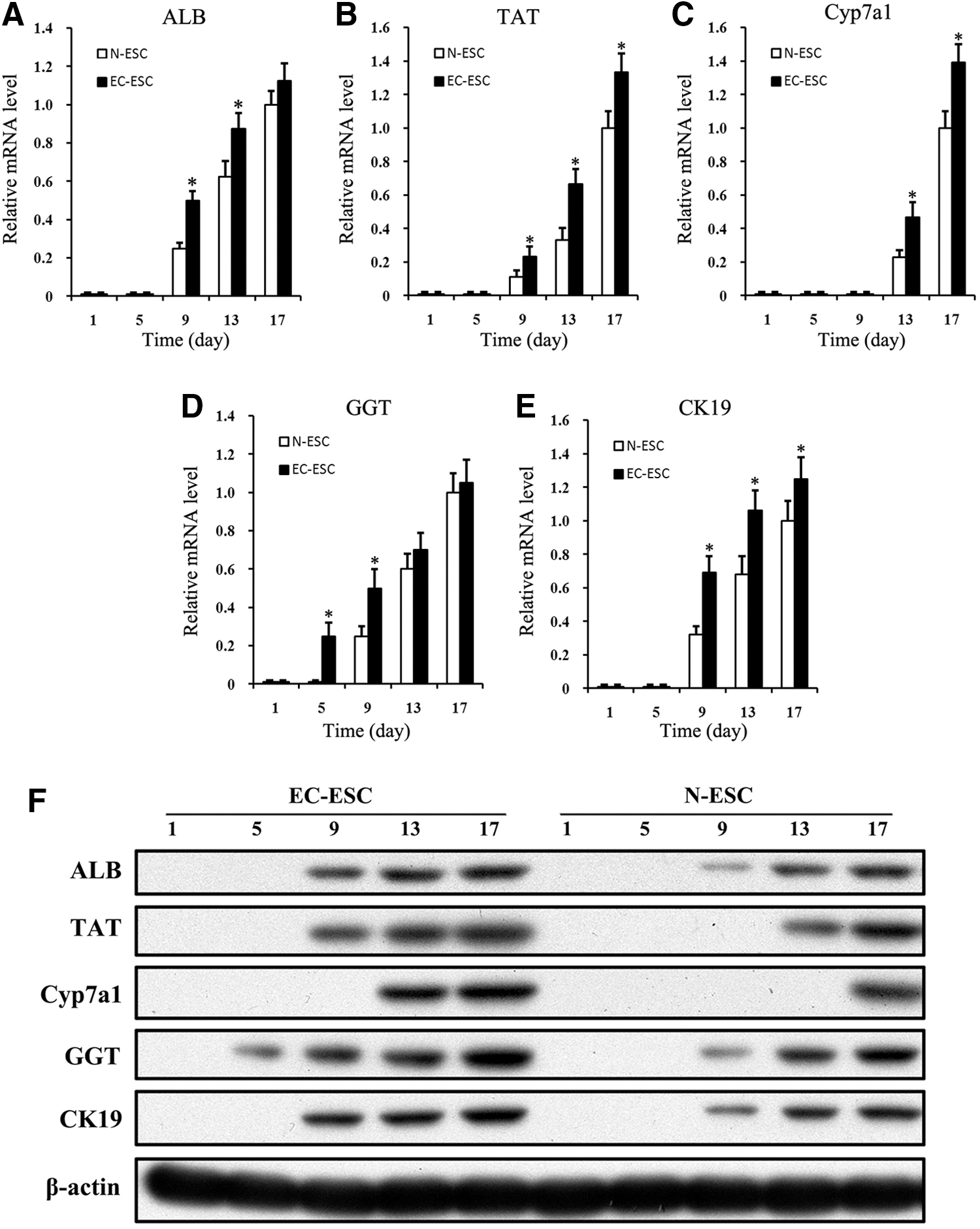
Exogenous E-cad enhances the expression of liver-specific genes and proteins. Cells were harvested from N-ESC or EC-ESC at the indicated time points.
We also performed ICC staining of ALB and CK19 on day 17. In the EC-ESC group, ALB- and CK19-positive cells showed compact cuboidal morphology, significantly stronger adhesion, and maintained better phenotypes of multidimensional growth (Fig. 4A, B). In contrast, ALB- and CK19-positive cells in the N-ESC group displayed migratory phenotypes and maintained phenotypes of monolayer growth (Fig. 4A, B). E-cad also expressed higher in the cytomembrane of multilayer cells in the EC-ESC group than that in the N-ESC group (Fig. 4C). ALB- and CK19-positive cells in EC-ESC differentiation showed more pronounced hepatic morphogenesis and maturation than that in the N-ESC group, because compact cuboidal hepatic morphology conventionally represents more differentiated cells [27].
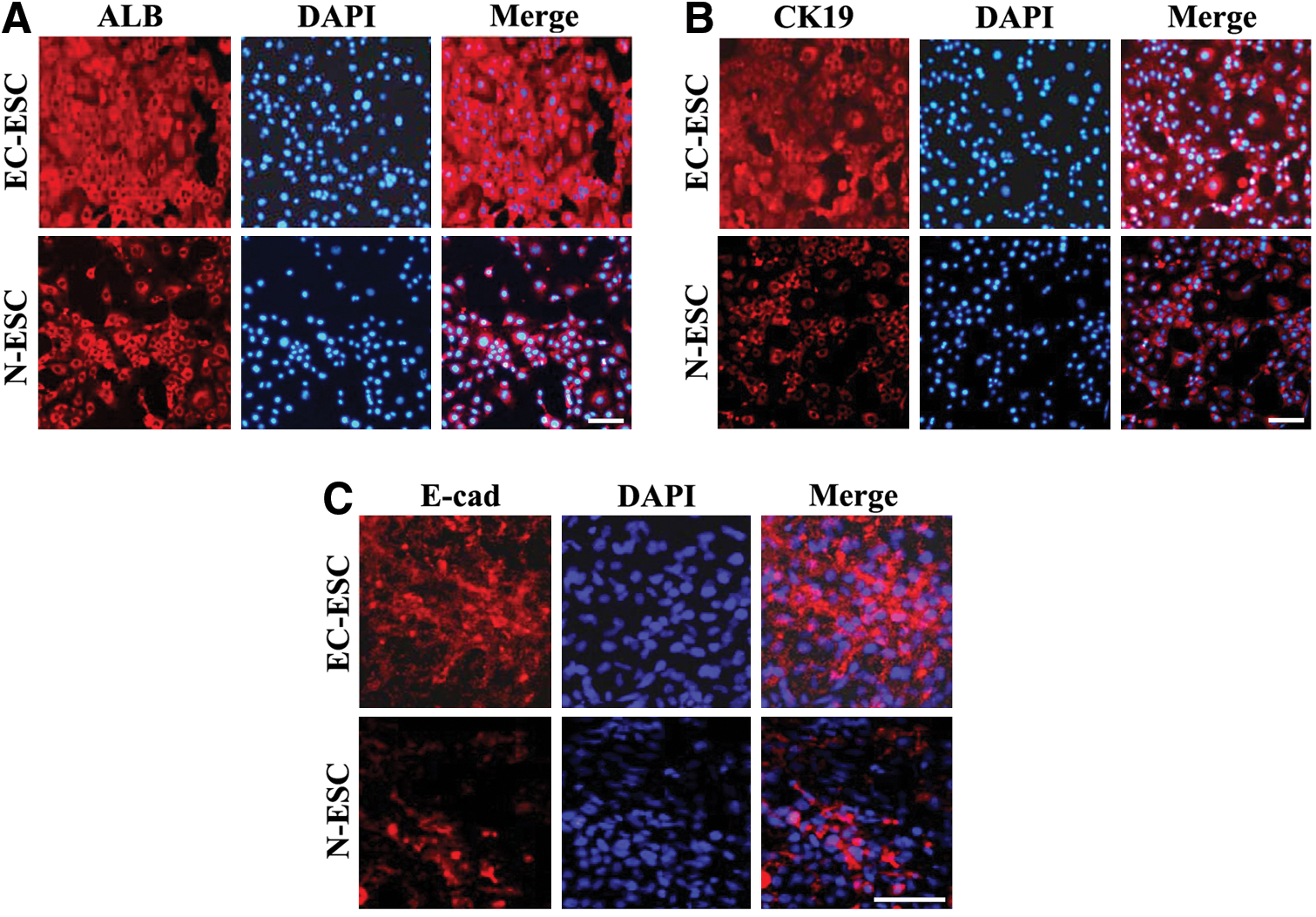
E-cad may facilitate a more native cytoskeletal organization and phenotype of hepatic cells. EBs from N-ESC or EC-ESC on day 17 were detected by immunofluorescence staining of ALB
E-cad facilitates the expression and function of vasculogenesis genes
In addition to parenchymal liver cells, endothelial cells and vascular structure are important indicators of the development of liver tissue [28]. Given the facts that E-cad could delay EMT and induce more differentiated hepatic cells, we detected the expression of VEGFR-1 and CD31/PECAM-1. The rationale behind the examination of VEGFR-1 and CD31/PECAM-1 is that they are important markers for endothelial cells and angiogenesis [29,30]. The expression of Brachyury was also examined, as it is a gastrulation marker that may suggest angiogenesis.
Our RT-PCR and western blotting assays suggested that, in N-ESC differentiation, mRNA of VEGFR-1 expressed on day 9–13 (Fig. 5A), CD31/PECAM-1 expressed on day 5–13 (Fig. 5B), and Brachyury expressed on day 5–9 (Fig. 2B). In the EC-ESC group, VEGFR-1, CD31/PECAM-1, and Brachyury expression extended to day 9–17, 5–17, and 5–17, respectively. Furthermore, their expression levels on day 9, 5 were lower than those in the N-ESC group. Thus, the expression of these genes was delayed and prolonged in EC-ESC differentiation than that in the N-ESC group. These data collectively suggested that the slowing down of EMT prolonged endothelial differentiation to overlap with the later stage of hepatic differentiation from ESCs and may help promote liver tissue formation in vitro.
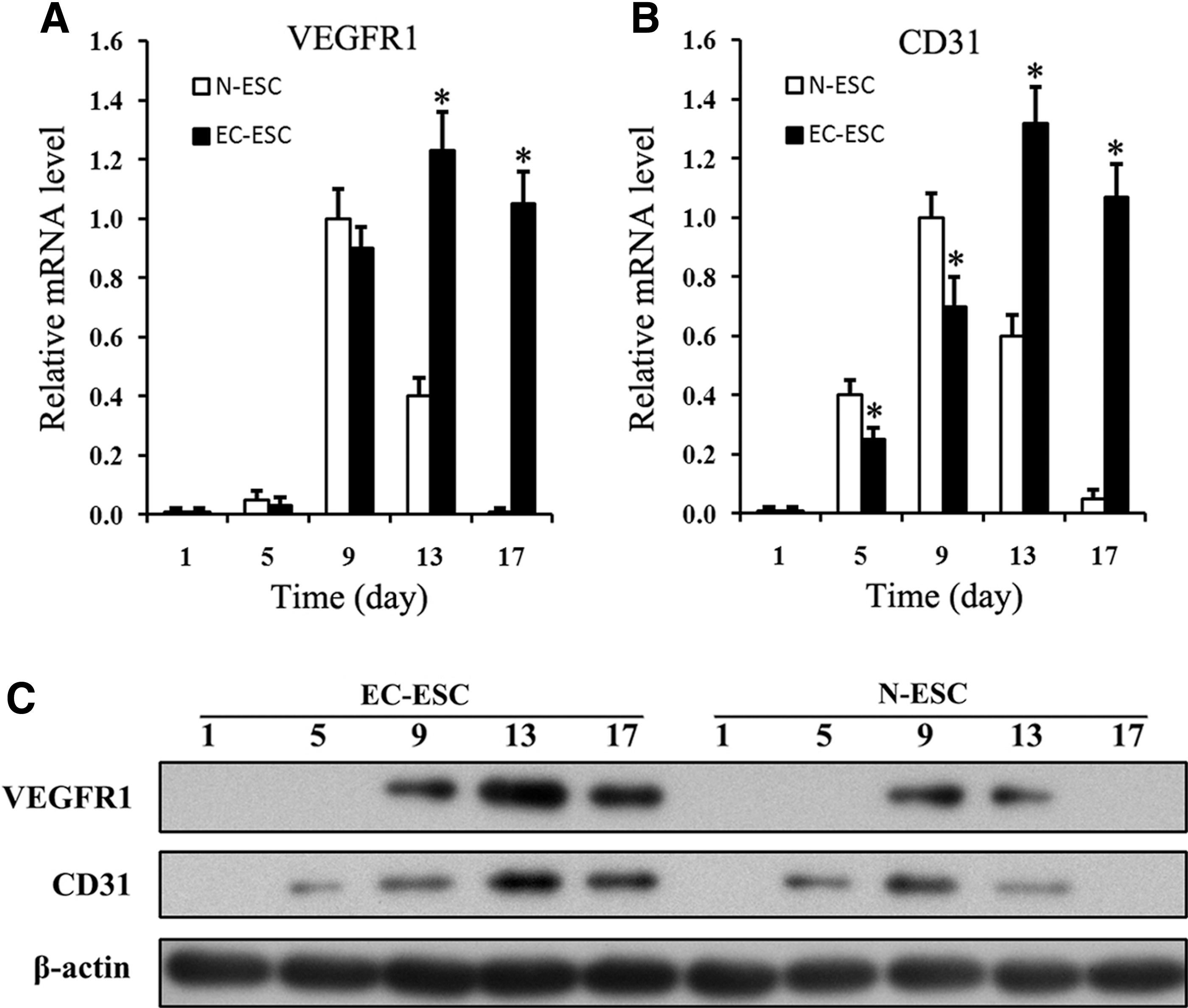
E-cad helps maintain the expression and function of vasculogenesis genes. Cells were harvested from N-ESC or EC-ESC at the indicated time points.
E-Cad enhances the adhesion in differentiated cells and facilitates the liver tissue-like structure formation during in vitro hepatic differentiation of ESC
Since cell adhesion and junction are the main mechanisms for in vivo tissue development, an examination of AFs in differentiated cells is important for the determination of tissue structure formation [31]. We hypothesized that E-cad transfection may enhance the adhesion in differentiated cells and, therefore, induce greater AFs in EBs in EC-ESC. To address this, we examined the AFs between EBs cells in N-ESC and EC-ESC differentiation using a micropipette suction system (See “Materials and Methods” section for details). We found that AF in EBs gradually decreased from day 5 to 17 in both groups. However, AF in the EC-ESC group was significantly stronger and decreased more slowly than that in the N-ESC group on day 9–17 (P<0.05). During day 13–17, cellular AF in the EC-ESC group was similar to that of mouse liver tissue (P>0.05) (Fig. 6A). These data provide an AF basis, suggesting that EC-ESC differentiation shows a liver tissue characteristic in terms of cellular AFs in differentiated cells.
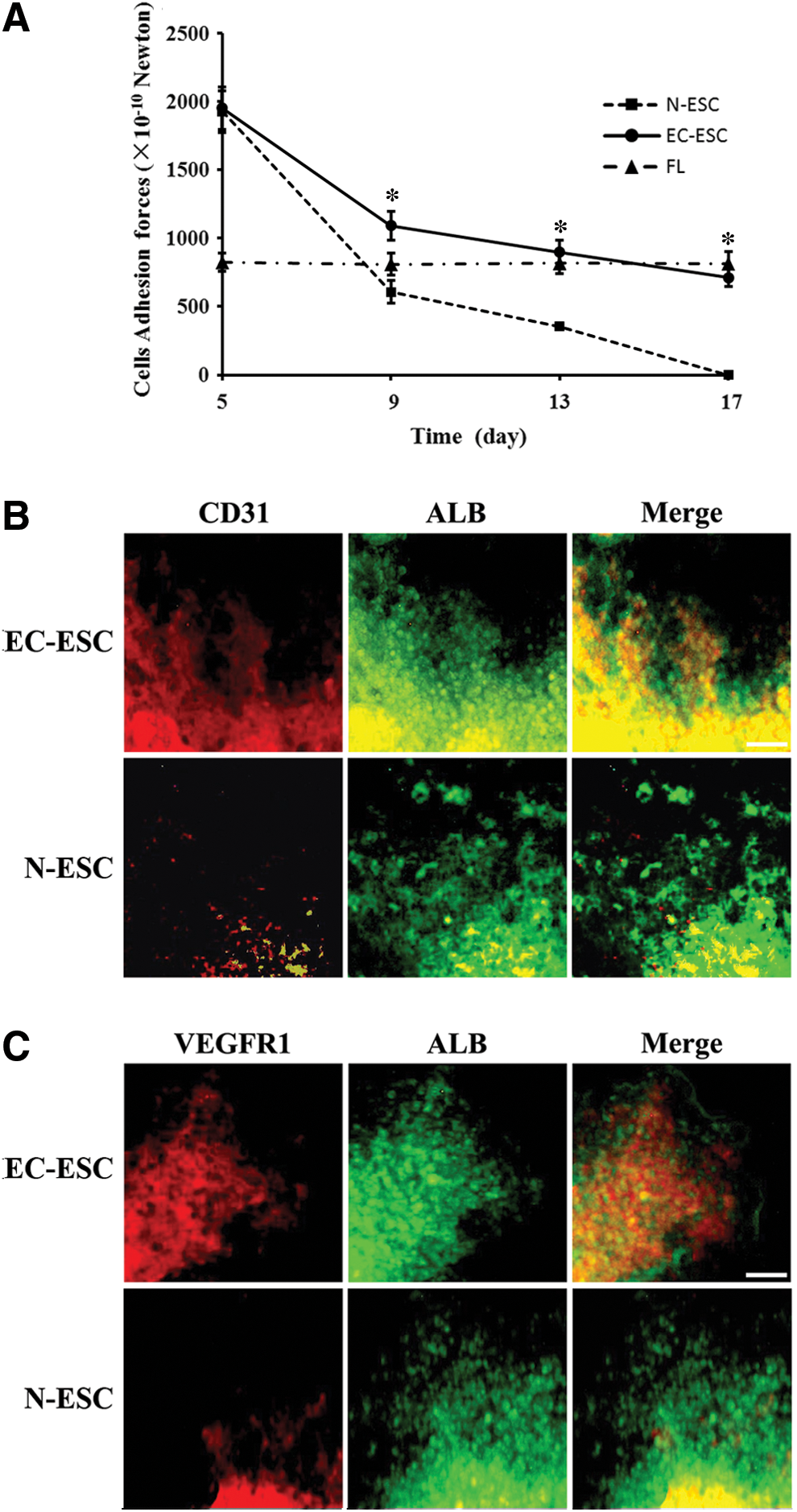
EC-ESC differentiation into liver tissue-like structures.
Finally, to further determine whether liver tissue-like structures have been developed from an EC-ESC culture system, we examined whether hepatic cells co-existed with vasculogenesis. We observed that some vascular network structures developed on differentiation at day 17 and endothelial cell markers CD31/PECAM-1 and VEGFR-1 were expressed simultaneously (Fig. 6B, C). Meanwhile, the vascular network structure contained a number of ALB-positive hepatocyte-like cells, which were distributed among the structures and presented multidimensional growth (Fig. 6B, C). The CD31/PECAM-1 and VEGFR-1-positive cells were seen to be migrating in the ALB-positive areas and made contact with the ALB-positive cells, Furthermore, biliary epithelial structures (CK19-positive) were surrounding ALB-positive cells (Supplementary Fig. S3). Taken together, this 3D structure was similar to the situation of liver organogenesis in the developing embryo [32]. The 3D structures composed of hepatocyte-like cells, endothelial cells, and a vascular network suggest the initial morphologic and functional features of liver tissue-like structures.
Discussion
In this study, we investigated the EMT regulated by E-cad expression and the role of EMT during hepatic differentiation from ESC. We observed that, in N-ESC differentiation without transfection of E-cad, E-cad expression obviously decreased during differentiation, and EBs dispersed to single layers. Thus, the EMT speed during in vitro ESC differentiation without E-cad transfection might be accelerated. However, in EC-ESC differentiation, we found that E-cad could slow down the EMT during hepatic differentiation from mouse ESCs, and such delayed EMT promoted the preservation of 3D structures of EC-EBs till day 17 and provided a histological basis for liver tissue formation. EMT is an important course of tissue development in embryogenesis and ESC differentiation [7,33]. It was postulated that, in ESC differentiation, E-cad expression should be decreased in order to facilitate the migratory phenotype of differentiated cells through EMT [34]. However, it has been previously shown that the accelerated EMT may somehow hinder the tissue development [8]. For example, accelerated EMT in tumorigenesis usually leads to a loss of regular or functional tissue structure [10], and the reason behind this might be that the loss of E-cad weakens cellular contacts and promotes β-catenin release. Our results suggest that the expression of E-cad can control the speed of the EMT and make it much closer to natural embryogenesis in vivo [35], which then facilitates the hepatic tissue differentiation from ESC. Thus, our data present a new line of evidence, suggesting that EMT should be carefully orchestrated for a successful hepatic tissue differentiation from ESCs.
Our results in this work showed that E-cad combined with hepatic growth factors could promote hepatocyte differentiation from ESCs. In this study, E-cad may have three kinds of mechanisms that promote hepatocte differentiation from ESC. First, E-cad enables differentiated cells to acquire more hepatic phenotypes by establishing correct cell polarity and by facilitating cell adhesion, regulating cytoskeleton, and maintaining cell polarization in epithelia [36]. Indeed, our adhesion forces measurement suggested that E-cad enhanced the adhesion between EB cells. Second, E-cad down-regulated Wnt/β-catenin pathway by sequestering β-catenin at the membrane and then induced hepatic differentiation of ESC [37,38]. Third, differentiations of hepatocytes, such as the differentiation of other epithelial cells, are usually not inhibited by the slowing of EMT in EC-ESC differentiation. In addition, we also found that the hepatocyte-like and BEC-like cells maintained multidimensional growth and showed a situation such as liver tissue in the EC-ESC group.
Another important finding in this study is that hepatic tissue differentiated from ESCs also expressed endothelial cell and angiogenesis markers in the late stage of differentiation. In addition to hepatic parenchymal cells, endothelial cells and vascular structure are basic indicators for the development of liver tissue [28]. When exogenous hepatic growth factors were added, endothelial cells were not detectable from day 13 to 17 in the N-ESC group, while E-cad helped preserve the endothelial cells in EC-ESC differentiation till day 17. As three germ layer structures, EBs owned endothelial cells in the early stage. However, here, we found that, after exogenous hepatic growth factors had been added, the percentages of endothelial cells among total EB cells gradually decreased, and EBs cells were rapidly directed to the differentiation into hepatic cells. In the late stage of differentiation, there may not be enough endothelial cells to be detectable. However, in the EC-ESC group, the EB differentiation process and EMT speed were significantly slowed down but not simply inhibited. This enabled the endothelial cells in EBs to be preserved for a much longer time. The slowing down of EMT prolongs endothelial differentiation to overlap with the later stage of hepatic differentiation from ESCs and may help promote liver tissue formation in vitro (Fig. 7).
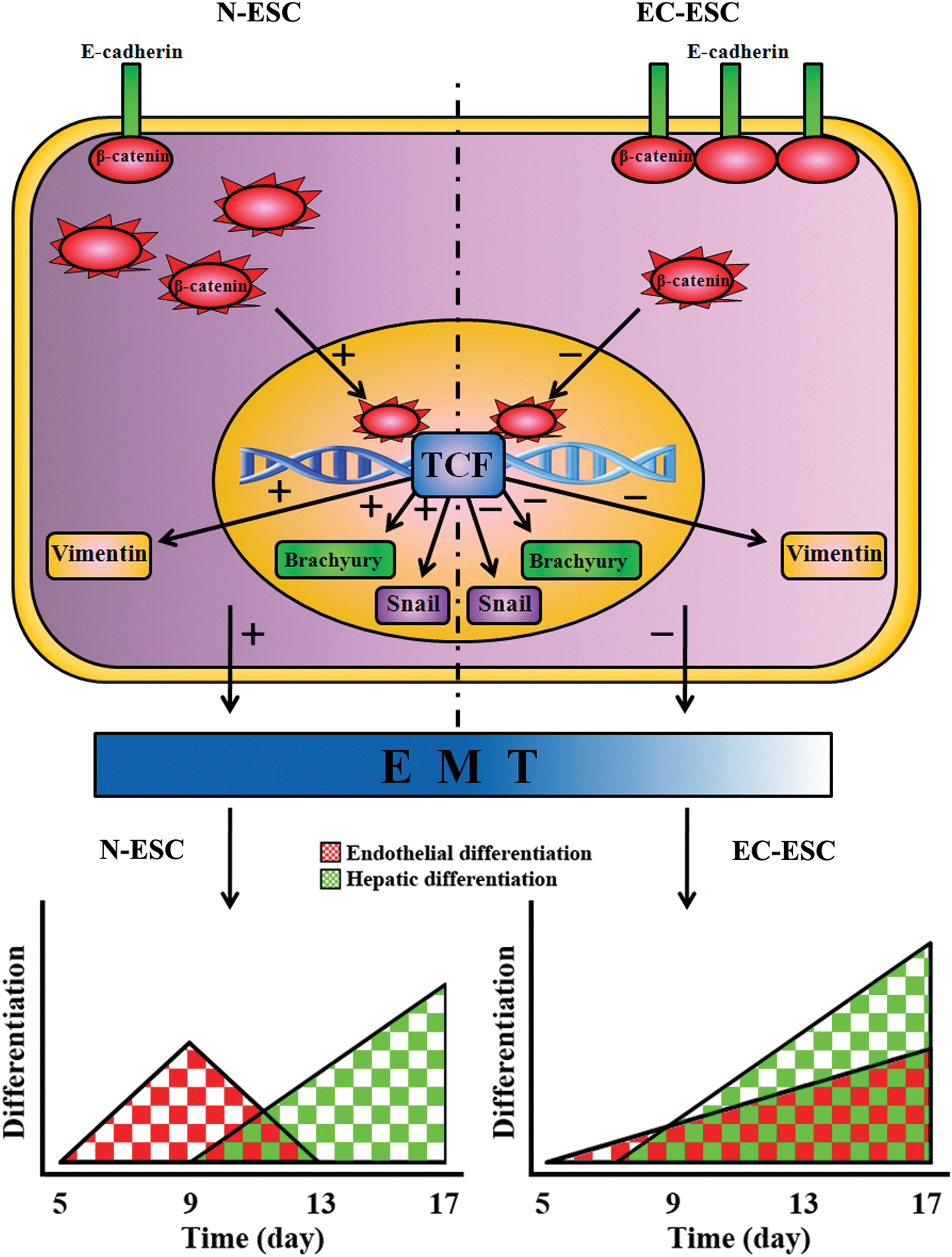
E-cad may regulate liver structure formation by slowing down and prolonging the accelerated differentiation and EMT progress in ESC differentiation. E-cad sequesters β-catenin at cell junctions and results in reducing the cytoplastic β-catenin. Cytoplastic β-catenin can translocate to the nucleus and function as an activator for T-cell factor (TCF), which activates the expression of a wide spectrum of EMT-responsive proteins. Then, the reducing cytoplastic β-catenin can slow down EMT. The slowing down of EMT then prolongs endothelial differentiation to overlap with the late stage of hepatic differentiation of ESCs and then helps form liver tissue in vitro. Arrows with “+” or “−” symbols indicate “increased” or “decreased,” respectively. Color images available online at
Morphological angiogenesis and regular cellular junction may provide direct evidence, indicating successful tissue development [39,40]. During in vivo angiogenesis development, adjacent endothelial cells adhered to each other to compose the blood vessel. They expressed VEGFR, CD31/PECAM-1, and their adhesion was induced by CD31/PECAM-1 and VE-cadherin. Others have identified some cell adhesion molecules such as CD31/PECAM-1, which may be required for microvessel growth [41]. In addition, ALB-positive cells were distributed among the vascular network, which was similar to the situation of liver organogenesis in the developing embryo [32]. Moreover, cellular AF in these multilayer cells in EC-ESCs group was significantly stronger than that in N-ESC, and similar to that in mouse liver tissue. Therefore, we may draw an inference that positive hepatic tissue appeared in our differentiation system.
In this study, we presented a novel system for liver tissue-like structure developed from murine ESCs. First, the EMT and Wnt/β-catenin pathway in the ESC differentiation process was regulated by E-cad and then, the 3D tissue structures of EBs could be maintained till day 17, which provides a histological basis for liver organogenesis. Second, essential cellular components, including hepatocytes, BECs, and endothelial cells, were simultaneously induced from ESCs in our differentiation system for liver tissue formation. The slowing down of β-catenin signaling and the EMT process prolongs endothelial differentiation to overlap with hepatic differentiation of ESCs and helps promote liver tissue formation in vitro. Third, positive angiogenesis was morphologically observed. These essential parenchymal cells and vascular network structures contacted each other, which was similar to the situation of in vivo liver organogenesis [32]. Lastly, the cellular AFs in this kind of liver tissue-like structures were found to be similar to those in mouse liver tissue. These data, therefore, collectively suggested that E-cad might regulate the liver structure formation by slowing down and prolonging the accelerated differentiation and EMT process in ESC differentiation (Fig. 7).
In summary, we examined the effects of E-cad on the ESC differentiation process and liver tissue formation by comparing N-ESC with EC-ESC. Although hepatic cells down-regulate the expression of E-cad in order to invade the surrounding stroma as migrating cords of hepatoblasts after primary liver bud formation in vivo [42], a certain amount of E-cad expression for the early stage of liver tissue formation is, indeed, necessary. E-cad can promote liver tissue formation during hepatic differentiation from ESCs through prolonging endothelial differentiation to overlap with hepatic differentiation by down-regulating the EMT and Wnt/β-catenin pathway. Furthermore, the phenomenon we found here that E-cad controls the ESC wild differentiation may provide a new insight for understanding the problem of tumorigenicity in stem cell transplantation in clinical practices. However, the identification of the liver tissue structure during differentiation in vitro was still difficult, and accurate identification methods were still lacking. Moreover, further studies are also needed to determine the precise expression levels of E-cad that are appropriate to tissue development at various differentiation stages of ESCs in vitro.
Footnotes
Acknowledgments
This study was supported by National Natural Science Foundation of China (30700398, 81172831, 81371693), national 863 plans projects of China (2007AA02Z117), Natural Science Foundation of Guangdong Province of China (06021354, 10151008901000200, and S2013010015935), scientific and technological projects of Guangdong Province of China (2009B030801126, 2012B031800257), the fundamental research funds for the central universities of China, and the young teacher culturing projects of Sun Yat-Sen University (09ykpy28).
Author Disclosure Statement
This work has no conflict of interest, including financial and other relationships.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.