
Abstract
To elucidate detailed functional mechanisms of key fate-determining transcription factors (eg, Nurr1, Pitx3, and Lmx1a) and their functional interplay for midbrain dopamine (mDA) neurons, we developed highly efficient gain-of-function system by transducing the neural progenitors (NPs) derived from embryonic stem cells (ESCs) with retroviral vectors, allowing the analysis of downstream molecular and cellular effects. Overexpression of each factors, Nurr1, Pitx3, and Lmx1a robustly promoted the dopaminergic differentiation of ESC-NP cells exposed to sonic hedgehog (SHH) and fibroblast growth factor 8 (FGF8). In addition, each of these factors directly interacts with potential binding sites within the tyrosine hydroxylase (TH) gene and activated its promoter activity. Interestingly, however, overexpression of Nurr1, but not of Pitx3 or Lmx1a, generated a significant number of nonneuronal TH-positive cells. In line with this, Pitx3 and Lmx1a, but not Nurr1, induced expression of the Ngn2 gene, which is critical for neurogenesis. We also observed that Pitx3 directly bound to its potential binding sites within the Ngn2 gene and the pan-neuronal marker β-tubulin III gene, suggesting that Pitx3 contributes to mDA neurogenesis by directly regulating these genes. Taken together, our data demonstrate that key mDA regulators (Nurr1, Pitx3, and Lmx1a) play overlapping as well as distinct roles during neurogenesis and neurotransmitter phenotype determination of mDA neurons.
Introduction
M
During early brain development, mDA neurons arise from neural progenitor (NP) cells generated by sequential developmental steps of regional specification and differentiation [9,10]. This development process is orchestrated by key signaling molecules [eg, sonic hedgehog (SHH), fibroblast growth factor 8 (FGF8), and Wnt signals] secreted from neighboring organizers and their downstream intrinsic transcription factors (eg, Otx2, Lmx1a/b, Foxa1/2, Ngn2, Pitx3, and Nurr1) [10 –24]. Importantly, some of these transcription factors have been successfully exploited for enhancing the generation of mDA neurons by their overexpression in stem cells. In particular, late transcription factors such as Nurr1 [25,26] and Pitx3 [27,28] as well as an early transcription factor Lmx1a [20,29] have been widely used to improve the yield of mDA neurons during in vitro differentiation of ESCs. Notably, several groups recently showed that it is possible to convert fibroblasts into functional mDA-like neurons by direct viral introduction of mDA-specific transcription factors (eg, Nurr1, Lmx1a, and/or FoxA2) along with general neurogenesis factors (eg, Brn2, Ascl1, and Myt1l) [30,31].
Thus, the identification and functional characterization of key mDA-specific transcription factors not only advanced our understanding of the developmental process of mDA neurons but also provided efficient in vitro methods to generate functional mDA neurons from stem cells or tissues via guided differentiation and direct conversion/transdifferentiation, respectively. However, we have still many obstacles to overcome before we can translate these potential methods into cell-based therapy or development of drug screening platforms [32 –35]. One important goal is to further understand the precise functional roles and analyze the overlapping and distinct roles of these key transcription factors in neurogenesis and DA phenotype determination. Toward this goal, we here analyzed and compared the functional effects when Nurr1, Pitx3, and Lmx1a genes are highly and universally overexpressed in ES cell-derived NPs. To our surprise, we found that overexpression of each of these three genes dramatically promoted the generation of TH-positive DA neuronal cells, demonstrating highly overlapping functions. However, they also exhibited significantly distinct functional effects in that overexpression of Nurr1, but not of Pitx3 or Lmx1a, generated a significant proportion of nonneuronal TH-positive cells. In line with this, Pitx3 and Lmx1a, but not Nurr1, increased mRNA expression of the neurogenesis-promoting Ngn2 gene. Finally, we found that Pitx3 directly interacts with potential promoter motifs and significantly increases mRNA expression of a pan-neuronal β-tubulin III gene, suggesting its multiple roles in neurogenesis and phenotype specification of mDA neurons.
Materials and Methods
Retroviral vectors construction, production, and titration
Human Nurr1, human Pitx3, and mouse Lmx1a cDNAs were amplified with primers for each gene using high fidelity Cloned Pfu DNA polymerase (STRATAGENE) and subcloned into the EcoRV site of the vector pUC19. Retroviral vectors expressing Nurr1, Pitx3, or Lmx1a were constructed by inserting the respective cDNA derived from pUC19 into the monocistronic retroviral vector pCL. The retroviral vectors were introduced into the retrovirus packaging cell line 293 GPG by transient transfection with Lipofectamine 2000 (Invitrogen). Forty-eight hours post-transfection, supernatants were harvested and kept at −80°C. Supernatants were collected every day for 2 weeks and used for transduction of cells. Cells were transduced with viruses in the presence of polybrene (2 μg/mL) for 2–3 h. Cells that were transduced with viruses were differentiated 2 days post-transduction in N2+AA media.
Maintenance and differentiation of mES cells
The mouse blastocyst-derived ES cell line J1 (kindly provided by Dr. En Li) was maintained as described previously [25,36]. To generate dopaminergic neuronal cells, we used the 5-stage in vitro differentiation procedure [37].
RT-PCR and real-time RT-PCR analysis
Total RNA from cells of in vitro differentiation was prepared using TriReagent (Sigma) followed by the treatment with DNase I (Ambion). Two micrograms of total RNA was reverse-transcribed into cDNA using oligo (dT) primers, according to the SuperScript Preamplification Kit (Life Technologies). The cDNA was then analyzed by polymerase chain reaction (PCR) using the following primers: GAPDH: 5′-TGACATCAAGAAGGTGGTGAAGC-3′, 5′-CCCTGTTGCTGTAGCCGTATTC-3′ (203 bp), En-1: 5′-TCAAGACTGACTCACAGCAACCCC-3′, 5′-CTTTGTCCTGAACCGTGGTGGTAG-3′, TH: 5′-TTGGCTGACCGCACATTTG-3′, 5′-ACGAGAGGCATAGTTCCTGAGC-3′, AADC: 5′-CCTACTGGCTGCTCGGACTAA-3′, 5′-GCGTACCAGGGACTCAAACTC-3′, DAT: 5′-CAGAGAGGTGGAGCTCATC-3′, 5′-GGCAGATCTTCCAGACACC-3′, c-Ret: 5′-TCCCAGAGTGAGTTACGAGACCTG-3′, 5′-GACAGCCCAAAGTCGGAAAT-3′.
Relative expression of mRNAs was assessed by normalizing levels of cDNA to the signal from glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA. PCR were carried out in PCR Buffer (Promega) containing 1.4 nM of each primer and 1.25 Unit of Taq I DNA polymerase (Promega). Samples were amplified in a Continuous Fluorescence Detector (MJ RESEARCH, Waltham, MA) using DNA Engine Opticon software under the following conditions: denaturing step at 94°C, 30 s; annealing step at 55°C, 30 s; extension step at 72°C, 30 s for 50 cycles; and a final extension step at 72°C, 10 min.
Immunocytochemistry
Cells were fixed in 4% formaldehyde in PBS, mounted on glass slides, and stained with the following primary antibodies against mouse monoclonal anti-nestin (Hybridoma Bank), rabbit polyclonalanti-tubulin III (Covance), and rabbit anti-tyrosine hydroxylase (Pel-Freez). Appropriate Alexa 488- and Alexa 594-labeled secondary antibodies (Molecular Probes) and 4′,6-diamidino-2-phenylindole counterstain were used for visualization. For analyses of stained cells, slides were observed under an Axioskope 2 plus fluorescence microscope (Carl Zeiss).
Chromatin immunoprecipitation assay
Chromatin immunoprecipitation (ChIP) assays were performed according to the manufacturer's protocol (Upstate). Briefly, 1×106 neural precursor cells derived from mouse embryonic stem cells were plated in 60-mm plates and cultured for 2 days and then transduced with a retroviral vector expressing the HA-tagged Pitx3. Cells were cultured 2 more days and differentiated into dopaminergic neurons for 7 days, and then cross-linked with 1% formaldehyde for 10 min and harvested in the presence of protease inhibitor (EDTA-free Complete; Roche). These cells were then lysed and sonicated to generate 200–500 bp DNA fragments. One tenth of the lysates was used for input control. The remaining lysates were divided into half and treated with 1 μg of polyclonal anti-HA antibody (Upstate) or normal rabbit IgG as a negative control overnight at 4°C. After the addition of Salmon sperm DNA/Protein A agarose slurry to immunoprecipitate complexes, these were extensively washed (0.01% SDS, 1.1% Triton X-100, 1.2 mM EDTA, 16.7 mM Tris-HCl, pH8.1, 167 mM NaCl), and proteins were eluted (1% SDS, 0.1 M NaHCO3). The cross-linked protein-DNA complexes were reversed by the treatment with NaCl. The DNA was recovered by phenol extraction and suspended in 50 μL of DW. PCR was performed to detect specifically bound DNA using MasterAmp 2×PCR Premix IN buffer (Epicentre) using 1 μL of the suspended sample as a template at 94°C 30 s, 55°C 30 s, 72°C 30 s for 30 cycles with primer sets in 25 μL reaction volume. Primers for TH are 5′-CCTCCTACCTGGAAATAGCC-3, 5′-CCTTTTGGGGAGTTCAACTC-3′ (P-Site1); 5-CACGACATGAAGACAGGGGC-3′, 5′-CCTGGAGGGGACTTGAAGAC-3′ (P-Site2); 5′-GACACAATCTAGAGACACTTG-3′, 5′-GTCTGGCCATATCCAGTG-3′ (P-Site3); 5′-GAGGTAGCTGGGAGTTCTG-3′, 5′-GCTTTCTCTGCGATCTG-3′ (P-Site4); 5′-CACCCACATAGCAGCTCAC-3′, 5′-GCAGATCATATTGAGTAAGG-3′ (P-Site5); 5-GGCCACCCATTACAGACCAG-3′, 5′-GGGAGCTCCAGTGACAG-3′ (P-Site6), and 5′-GTGGTTCCCAGGGAGCTGAG-3′, 5′-GCTTTCAGATTATCCTTGGG-3′ (N-Site1); 5′-GTCTCTAAGCTCTTTGAGGTG-3′, 5′-CTTATTATTGAAGTAACAGC-3′ (N-Site2); 5′-GGAGTGCCCAGCGGCTTGCC-3′, 5-GATTGGTTCAGATGCTTGCTC-3′ (N-Site3). Primers for Tuj1 are 5′-GTCCTCCAGATAGTGCAGAG-3′, 5′-GACAGGGTTTCTCTGTGTAG-3′; 5′-GGCAGAATGTCTGAAGGAG-3′, 5′-GGAGACCCCTGATTTGTCC-3′; 5′-GCATATGGTGGCTCACAGCC-3′, 5′-CTCTGGCTGTCCTGGAACTC-3′. Primers for NeuroG2 are 5′-GTCCCACTGCTCTGTCCCAG-3, 5′-GCACTAATCGCTGAACCAGG-3′.
Luciferase assays
Neuroblastoma BE2C cell line was cultured in Dulbecco's modified Eagle's medium, supplemented with 10% fetal bovine serum, penicillin (100 IU/mL), streptomycin (100 μg/mL), 2.5 mM L-glutamine, 1% nonessential amino acids (all from Invitrogen), and cultured in humidified air containing 5% CO2. Cells were grown on 35-mm dishes and transfected upon reaching 70% confluence using the calcium phosphate method. A reporter construct was cotransfected with the expression vector, the TH promoter (2.6 kb)-luciferase plasmid, and carrier plasmid DNA. Forty-eight hours after transfection, the cells were lysed (0.1% SDS, 1.0% Triton X-100, 0.5% Na-Deoxycholate, 50 mM Tris-HCl, pH8.0, 150 mM NaCl) with Complete Mini protease inhibitor cocktail (Roche) and PMSF (Santa Cruz Biotechnology), the lysate was centrifuged, and the supernatant was used to measure the luciferase and β-galactosidase activity. All transfections were carried out in triplicate and repeated at least four times.
Cell counting and statistical analysis
Cell number was determined by counting the numbers of β-tubulin+ and TH+ cells per field at×100 magnification using the Axioskope 2 plus fluorescence microscope (Carl Zeiss). Five visual fields were randomly selected and counted for each sample. Numbers presented in figures represent the average percentage and SEM of TH+ cells over β-tubulin+ cells.
All statistical analyses were conducted using SAS v 9.1 (SAS Institute) for Windows 2000 Professional. The analysis was conducted on these means using a mixed models analysis of variance procedure (SAS, Proc mixed) to determine possible statistical differences between group means.
Results
Temporal overexpression of Nurr1, Pitx3, or Lmx1a at the NP stage during in vitro ESC differentiation robustly and comparably promotes the generation of mDA neurons
Previous studies from this and other laboratories extensively tested the functional effects of overexpression of Nurr1 [25,26,38], Pitx3 [27,28], and/or Lmx1a [20,29]. Using stably overexpressing lines, these investigations revealed the potential of these critical transcription factors for enhancing mDA neuronal differentiation. However, these studies are plagued by the intrinsic limitations associated with leaky and variable transgenic expression caused by random chromosomal integration. To overcome these limitations, we used retroviral transgene expression at the relevant differentiation stage, that is, NP stage cells derived from mouse ESCs (Fig. 1A). This approach advantages include tightly controlled temporal expression and extremely efficient transduction efficiency, overcoming the limitation associated with the analysis of limited number of stable cell lines. Indeed, double immunostaining for the endogenous NP marker Nestin and the retrovirally expressed GFP at 3 days after transduction revealed that the great majority of NP cells (>98%) was effectively transduced by retroviral vectors (Fig. 1B).

Efficient generation of tyrosine hydroxylase (TH)+ cells from ES cell-derived neural precursor (ES-NP) cells by retrovirus systems.
We next transduced NP cells (with or without SHH/FGF8 pretreatment) with retroviral vectors expressing Nurr1, Pitx3, or Lmx1a and analyzed the functional effects following neuronal differentiation for 2 weeks, based on the 5-stage in vitro differentiation procedure (13, 10–11) (Fig. 1A). Remarkably, retroviral overexpression of Nurr1, Pitx3, or Lmx1a in SHH/FGF8-pretreated NPs resulted in dramatically enhanced generation of TH+ cells [∼65.8%, 63.5%, and 60.3%, respectively, of the total 4′,6′-diamidino-2-phenylindole (DAPI)+ cells], compared to control empty vector-transduced cells (11.8%) (Fig. 1C–D). In contrast, when NP cells were transduced with these vectors without SHH/FGF8 pretreatment, only Nurr1 overexpression, but not Pitx3 or Lmx1a, significantly increased the generation of TH+ cells (∼29.2% of DAPI+ cells) compared to control-transduced cells (13.5%) (Fig. 1C). Together, our results show that Nurr1, Pitx3, and Lmx1a have similar potential to enhance DA neuronal differentiation only under appropriate cellular contexts that depend on pretreatment with SHH and FGF8.
When we re-analyzed the number of TH+ cells among β-tubulin III+ (Tuj1+) neuronal cells in SHH/FGF8-pretreated NPs following neuronal differentiation, the overexpression of Nurr1, Pitx3, and Lmx1a robustly increased its proportion up to ∼77.3%, 91.4%, and 92.2%, respectively, compared to control vector (26.7%) (Fig. 2A–B). Consistently, real-time RT-PCR analysis of differentiated cultures showed that TH mRNA expression was prominently induced by each transcription factor with the highest induction by Nurr1 (Fig. 2C). In addition, significantly increased expression of all mDA-specific markers tested in this study (ie, TH, AADC, VMAT2, and DAT genes) was observed (Fig. 2D). Our data show that each of these transcription factors (ie, Nurr1, Pitx3, and Lmx1a) is comparably able to increase expression of multiple mDA-specific genes following its overexpression during the SHH/FGF8-primed NP stage, indicating that they share overlapping/similar functions to enhance mDA differentiation.
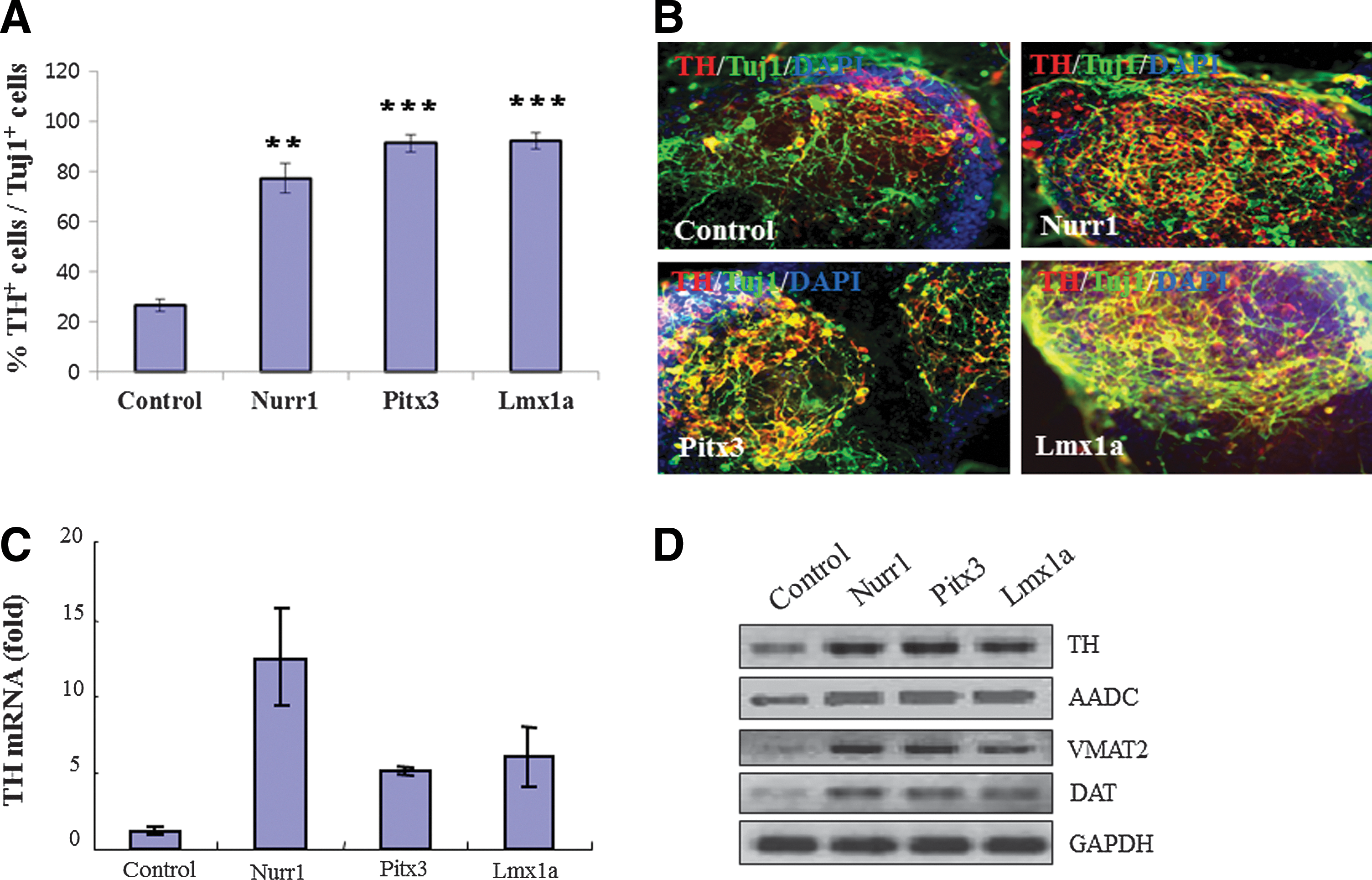
Overexpression of each individual transcription factor (Nurr1, Pitx3, and Lmx1a) promotes midbrain dopaminergic (mDA) neuronal differentiation.
Nurr1, Pitx3, and Lmx1a directly bind to the TH gene promoter and activate its mRNA expression
We next addressed whether these transcription factors directly activate TH gene transcription via binding to its promoter. Toward this goal, we performed ChIP–PCR analysis. When we analyzed the mouse and rat TH gene promoter regions ranging from 70 kb upstream (−70 kb) to 9 kb downstream (+9kb) of transcriptional start site using the Nurr1 binding motif (5′-AAAGGTCA-3′) [39] with the ECR browser program [40], we identified three conserved sites, residing at +831 to +838 (TGACCTTT), at −17542 to −17535 (AAAGGTCA), and at −45882 to −45844 (TGACCTTT) (Fig. 3A–C). We designed the primers to amplify those regions that contained three fully conserved Nurr1 binding sites in TH gene of mouse genomic sequence (Fig. 3A). We also compared the same mouse and rat TH promoter regions using the Pitx3 binding motif 5′-S(C or G)N(G or A or T or C)TAATCCM(A or C)-3′ [41,42] with ECR browser program [40]. We identified one conserved site at −41584 to −41593 (GTTAATCCC) and five consensus sites at +1151 to +1160 (CCTAATCCC), at −350 to −358 (TGGGATTACC), at −9891 to −9899 (GGGATTACC), at −13266 to −13274 (CCTAATCCC), and at −48341 to −48349 (CCTAATCCA) (Fig. 3D–F). We designed the primers to amplify those regions that contained one fully conserved and five-consensus Pitx3 binding sites in TH gene of mouse genomic sequence (Fig. 3D). In addition, we compared the same mouse and rat TH promoter regions ranging from 10 kb upstream (−10 kb) of the transcriptional start site using the Lmx1a homeodomain binding motif 5′-TAAT-3′ [12,43] with ECR browser program [40]. We identified eight consensus sites (Fig. 3G–I) and designed primers to amplify those regions of the mouse TH genomic sequence. We next differentiated ESCs into NP cells and transduced them with retrovirus expressing FLAG-tagged Nurr1 or HA-tagged Pitx3 or HA-tagged Lmxla. Following neuronal differentiation, cross-linked chromatin complexes were immunoprecipitated using either αFLAG or αHA antibodies to detect each tagged transcription factor. Immunoprecipitated DNA fragments were subjected to PCR analysis, using IgG-immunoprecipitated DNA and -input DNA as negative and positive control, respectively. As shown in Fig. 3B, all three Nurr1 conserved sites residing in the TH promoter region were found to interact with FLAG-tagged Nurr1, indicating that they are genuine Nurr1-binding sites. In the case of the potential Pitx3 sites, the conserved site (site 5) as well as four sites (region 1, 2, 3 and 6) were found to interact with HA-tagged Pitx3 (Fig. 3E). The potential site 4 was not amplified by ChIP analysis despite the fact that it contains the consensus Pitx3-binding motif CCTAATCCC (Fig. 3E). Looking at the putative Lmx1a sites, we found that Lmx1a bound to sites 1, 34, 56, and 78, but not site 2, indicating the specificity of Lmx1a binding (Fig. 3H). To further test the regulation of the TH gene promoter by these factors, we next assayed the transcriptional activity of the TH reporter construct (containing the 2.6 kb upstream region fused to the reporter luciferase gene) by transient overexpression of each factor in human neuroblastoma SK-N-BE(C) cells. As shown in Fig. 3J, overexpression of Nurr1, Pitx3, or Lmx1a robustly induced the TH promoter activity. In contrast, none of them induced the promoter activity of the dopamine β-hydroxylase (DBH) gene, which is not expressed in mDA neurons and lacks these factors binding motifs (Fig. 3J). Taken together, our results strongly suggest that Nurr1, Pitx3, and Lmx1a directly bind to the promoter motifs of the TH gene and activate its transcription in a highly overlapping and concerted manner.
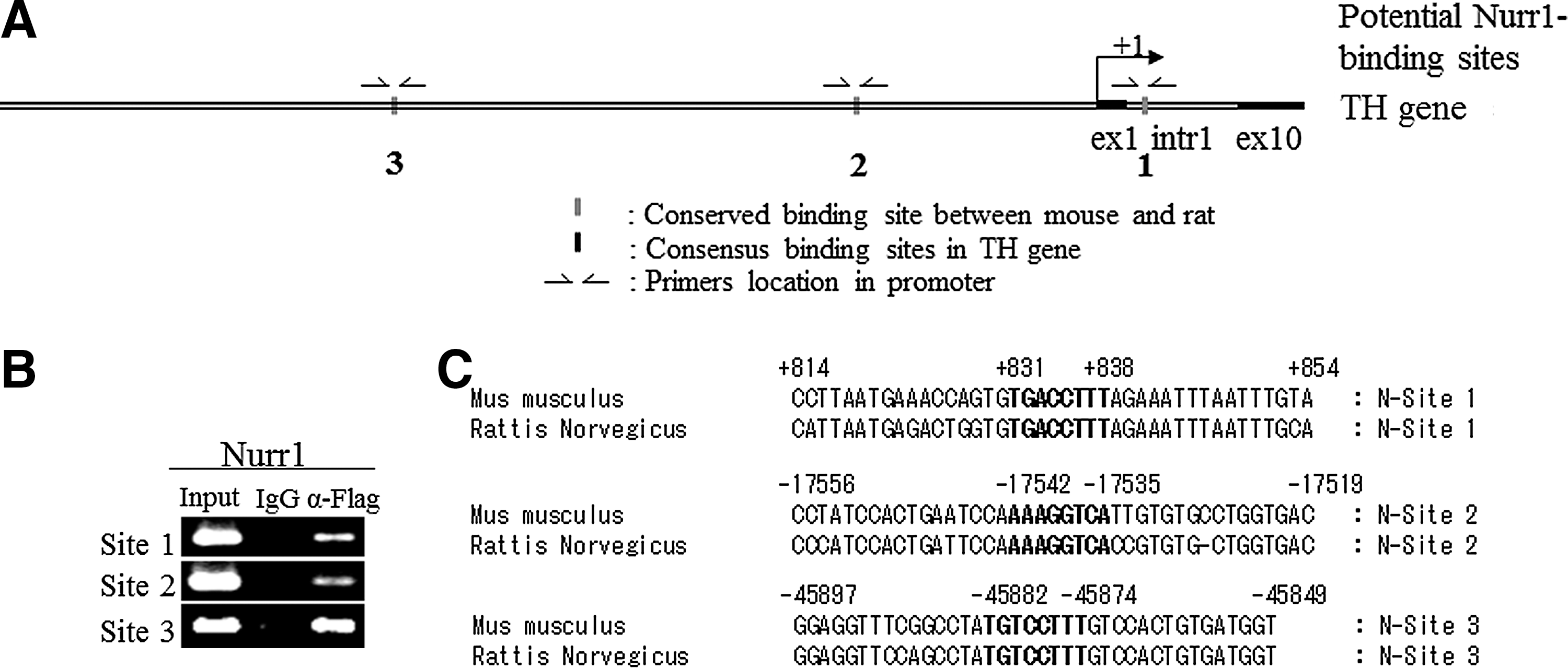
Nurr1 and Pitx3 directly bind to tyrosine hydroxylase (TH) gene from in vitro differentiated cells from Nurr1- and Pitx3- individually transduced NP cells.
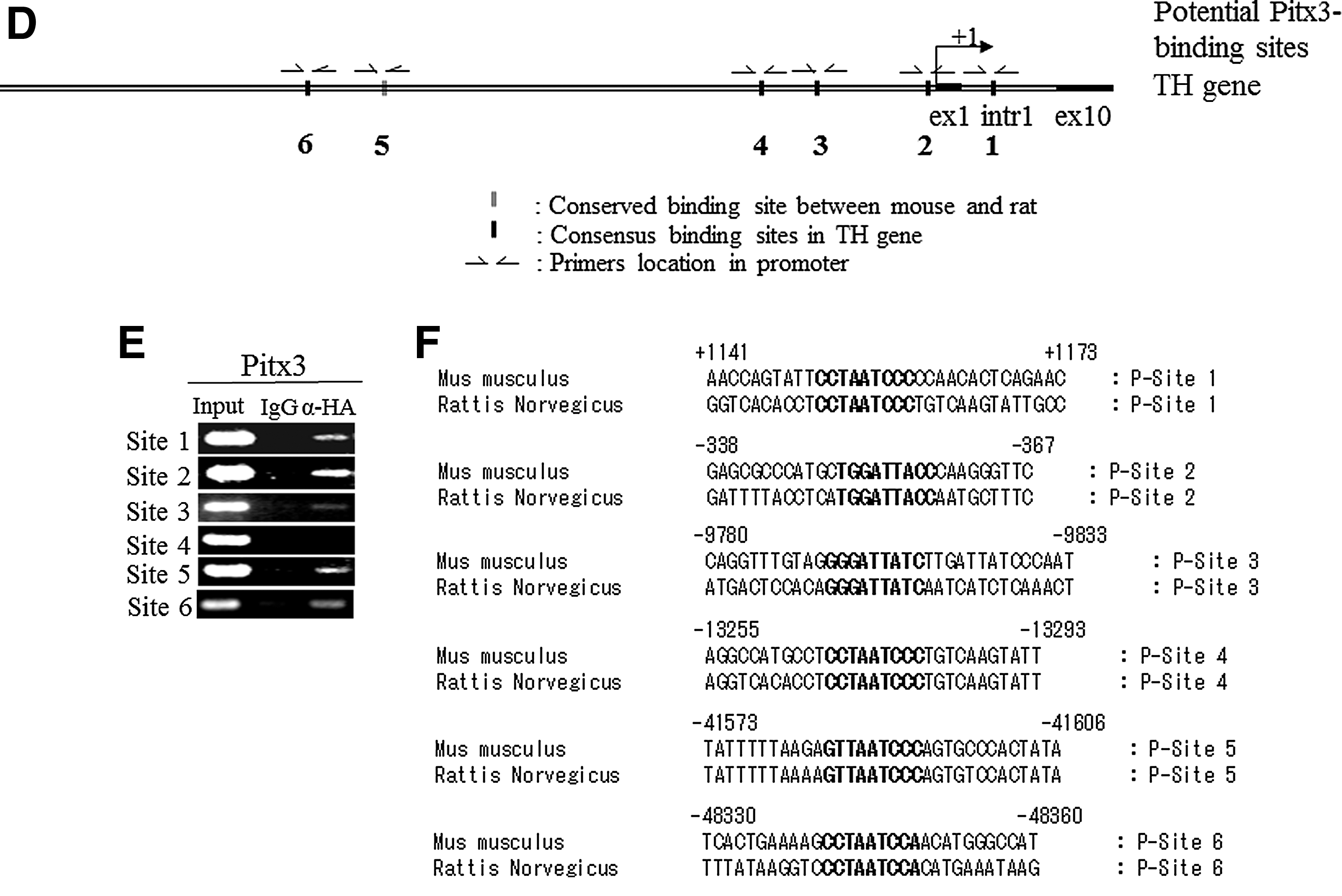
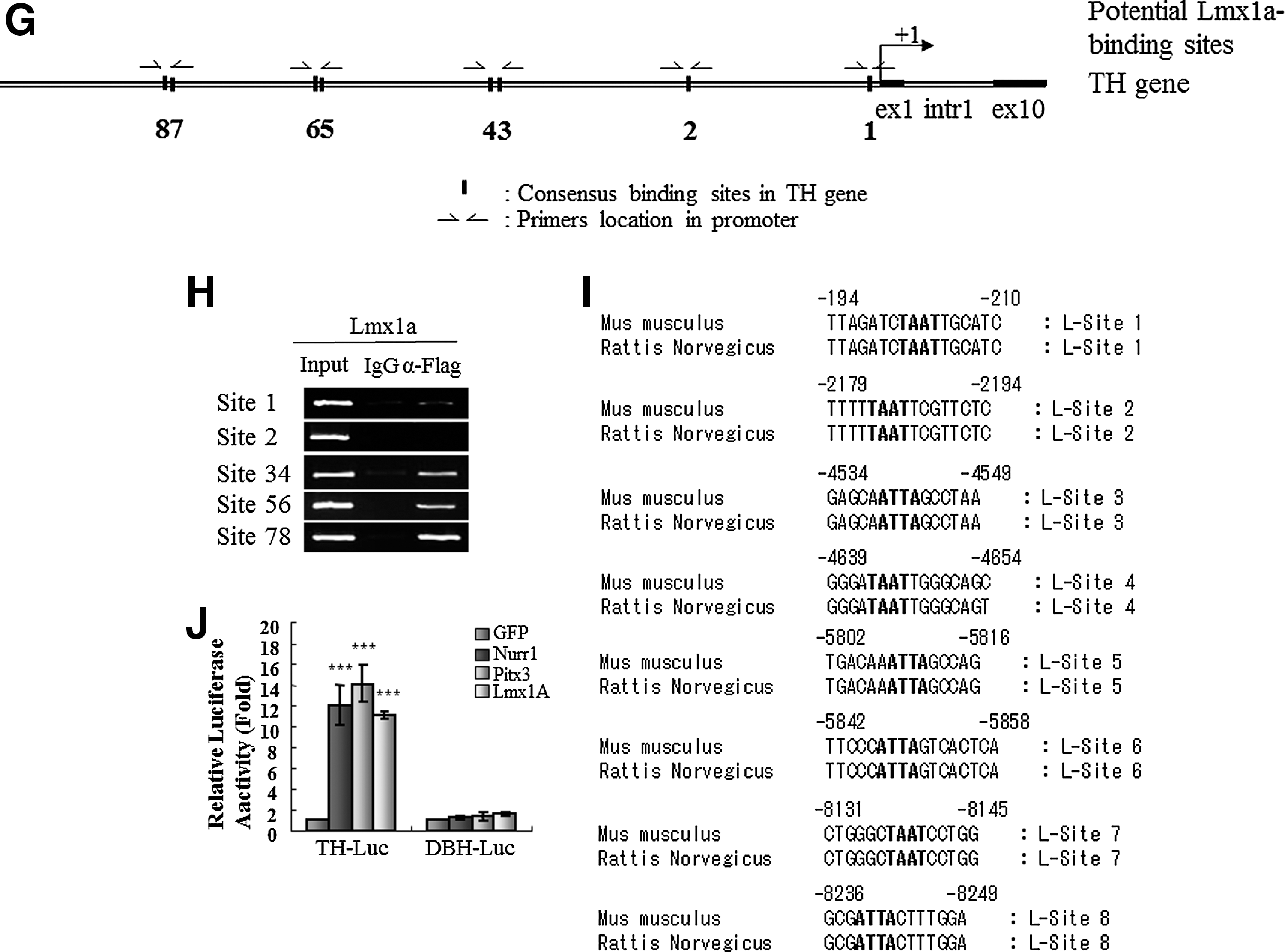
Distinct functional role(s) of Nurr1, Pitx3, and Lmx1a on mDA neuronal differentiation
Notably, the overexpression of Nurr1 increased TH+/Tuj1+ neuronal cells to a lesser degree (∼77%) than Pitx3 and Lmx1a (∼92%) (Fig. 2A), whereas they all similarly increased the proportion of TH+ cells (Fig. 1C). This discrepancy suggests the possibility that Nurr1 overexpression generated a significant proportion of TH+/Tuj1− cells. Indeed, our stereological cell counting revealed that ∼19% of TH+ cells are Tuj1− in Nurr1-overexpressing cells while only 1.5% to 2.0% of TH+ cells are Tuj1− in control vector-, Pitx3- or Lmx1a-transduced cell (Fig. 4A–B). These surprising findings prompted us to hypothesize that Nurr1 primarily controls the DA phenotype specification while Pitx3 and Lmx1a regulate both the DA phenotype and general neurogenesis during in vitro differentiation of ES cells.

Pitx3 and Lmx1a genes rather than Nurr1 specify dopaminergic phenotype during in vitro differentiation of transduced NP cells.
Based on our recent identification of the proneural gene Ngn2 as a potential target of Pitx3 [44], we hypothesized that differential regulation of Ngn2 by Pitx3, Nurr1, and/or Lmx1a may underlie the above distinct functional roles by these transcription factors. To test this, we examined Ngn2 mRNA expression by qPCR analysis in differentiated neuronal cells of Nurr1-, Pitx3-, and Lmx1a-transduced cells. As shown in Fig. 5A–B, overexpression of Pitx3 and Lmx1a significantly increased Ngn2 mRNA expression, compared to control. In contrast, Nurr1 overexpression did not affect Ngn2 gene expression, demonstrating that Nurr1 vs. Pitx3 and Lmx1a differentially regulate it. When we compared the Ngn2 gene promoter region from 20 kb upstream (−20 kb) to 3 kb downstream (+3 kb) of the transcriptional start site with the consensus Pitx3 binding motif (5′-SNTAATCCM-3′) using the ECR browser program, we identified a highly conserved binding site, residing at −6230 to −6222 bp (5′-GGGATTAAC-3′) among mouse, human, monkey, dog, chicken, rat, and frog sequences (Fig. 5C–D). We next amplified this highly conserved Pitx3 binding site from the mouse Ngn2 gene and used it for ChIP-PCR analysis, following the transduction of NP cells with retroviral vector expressing HA-tagged Pitx3. Interestingly, this Pitx3 motif was specifically immunoprecipitated (Fig. 5E), strongly suggesting that Pitx3 directly binds to the conserved site of the Ngn2 gene promoter and induces its transcription.
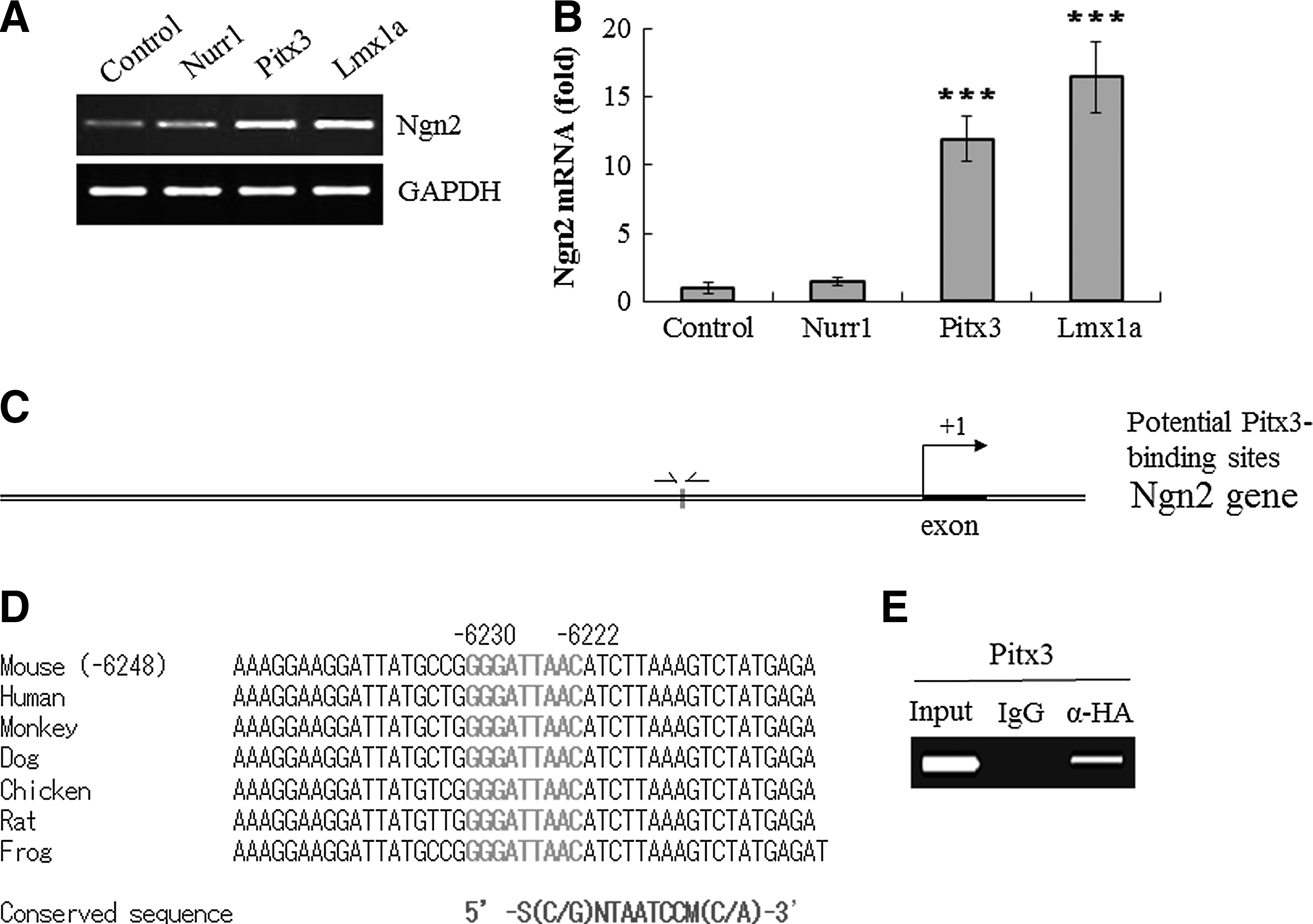
Pitx3 induces pro-neural marker neurogenin 2 (Ngn2) mRNA expression and also directly binds to Ngn2 gene.
Pitx3 induces transcription of the pan-neuronal marker β-tubulin III via direct binding to its promoter
We next addressed if Nurr1, Pitx3, and Lmx1a differentially regulate the expression of the β-tubulin III gene because it is a prototype pan-neuronal gene whose expression is essential for general neuronal property. Toward this end, we first examined the percentage of Tuj1+ cells among the total DAPI+ cells following neuronal differentiation of control-, Nurr1-, Pitx3-, and Lmx1a-transduced cells. The proportion of Tuj1+ cells was significantly increased (by ∼10%) in Pitx3-transduced cells compared to control and other groups (Fig. 6A). In agreement with this observation, real-time RT-PCR analysis showed that overexpression of Pitx3, but not of Nurr1 or Lmx1a, significantly increased mRNA expression of Tuj1, compared to control (Fig. 6B). In addition, using the ECR browser program, we searched for conserved potential Pitx3 binding sites between the mouse and rat β-tubulin III (Tuj1) gene promoter regions ranging from 2.5 kb upstream (−2.5 kb) to 9 kb downstream (+9 kb) of the transcriptional start site using the Pitx3 binding motif 5′-SNTAATCCM-3′. We identified three putative binding sites, residing at +6294 to +6302 (TTTAATCCC), +7632 to +7640 (AATAATCCC), and +7940 to +7948 (TTTAATCCC) of the mouse gene sequence (Fig. 6C–D), although there was no conserved site. Using the three putative motifs (P-site 1 at +6149 to +6403, P-site 2 at +7470 to +7738, and P-site 3 at +7811 to +8050), our ChIP-qPCR analysis showed that all three sites are specifically immunoprecipitated by αHA antibody (Fig. 6E). These results indicate that Pitx3 directly binds to its cognate sites and induce the transcription of the Tuj1 gene, directly contributing to pan-neuronal phenotypes as well as to mDA-specific phenotypes during in vitro differentiation of ES cells.
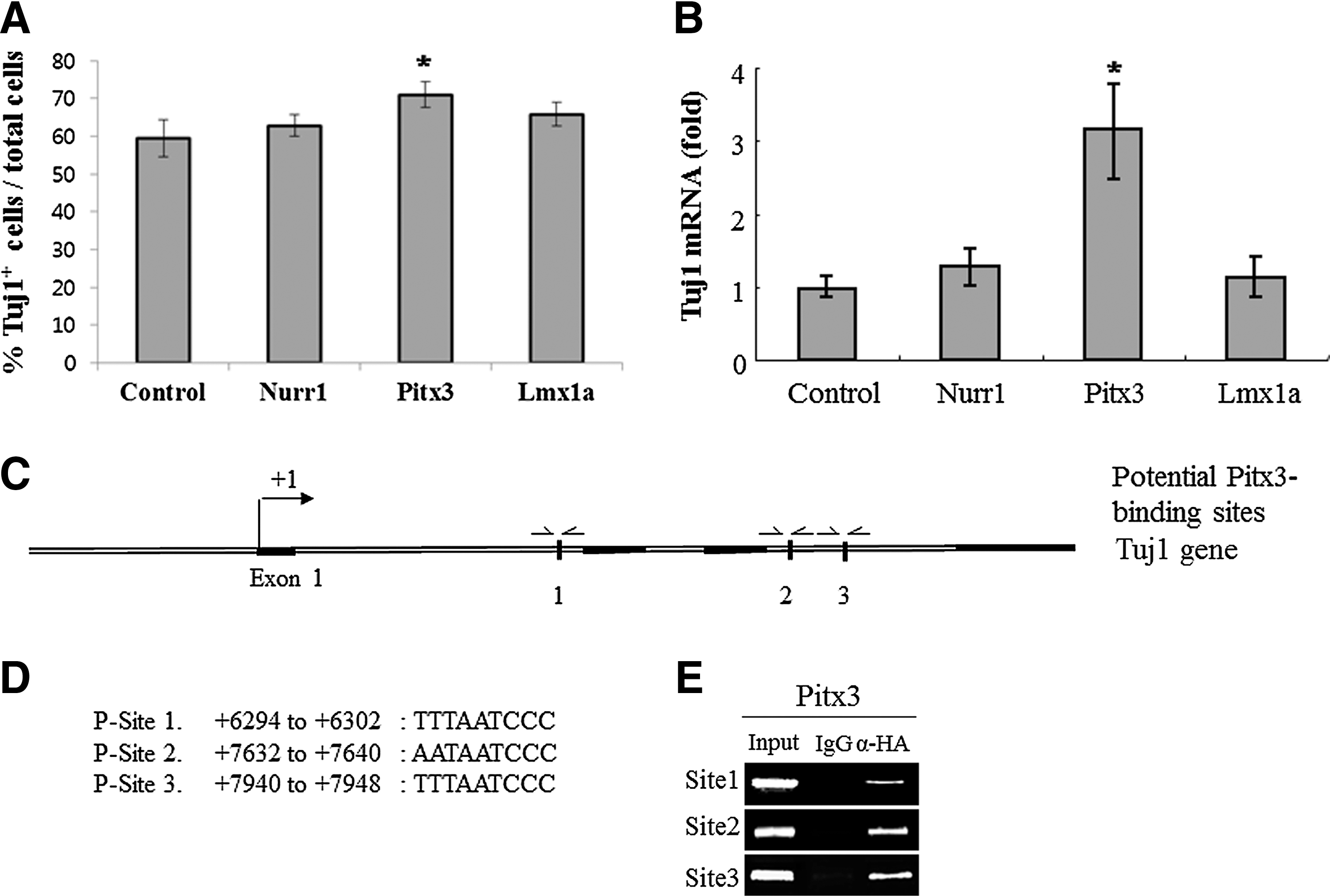
Pitx3 significantly increases β-tubulin III+ (Tuj1+) cells and mRNA expression and also directly binds to Tuj1 gene.
Discussion
Due to the critical importance of mDA neurons for diverse brain functions in health and disease, numerous scientists have investigated the regulatory mechanisms underlying their development and phenotype determination. In particular, there have been impressive progresses in identification and functional characterization of key transcription factors and in our understanding of how key signaling molecules and transcription factors orchestrate the development of mDA neurons in the mouse brain [9,45,46]. Despite this progress, precise functional roles and interplays between these transcription factors have yet to be clearly defined. In this study, we focused our attention on three key transcription factors, Nurr1, Pitx3 and Lmx1a, and investigated their functional roles during in vitro differentiation of ESCs into mDA neurons. Toward this goal, we established a highly efficient gain-of-function analysis by transducing ESC-derived neural precursors (ESC-NPs) with retroviral vectors expressing each of these factors. Compared to previous gain-of-function studies in which individual clones overexpressing each transcription factor were used, our approach offers two advantages. First, by allowing precise temporal control of exogenous gene expression, it likely recapitulates the in vivo effect from each transcription factor more accurately than analyzing stable ESC clones expressing each exogenous transcription factor from pluripotent stages. Second, retroviral overexpression of a transcription factor in a large number of cells simultaneously and efficiently (>98% of ESC-NPs are transduced by retroviral vector; Fig. 1) is likely more accurate and offers an unbiased study of functional effects than characterizing a handful of stable clones.
When each Nurr1, Pitx3, or Lmx1a was overexpressed at the ESC-NP stage without prior exposure to SHH and FGF8, Nurr1, but not Pitx3 or Lmx1a, was able to significantly increase the number of TH+ cells among Tuj1+ cells. In contrast, when ESC-NPs were exposed to SHH and FGF8 for 2 days before retroviral expression, each transcription factor (Nurr1, Pitx3, and Lmx1a) robustly (>six-fold) enhanced the generation of TH+ cell number compared to control empty vector. In line with these results, real-time RT-PCR analysis showed that the level of TH mRNA was robustly increased in each case. Taken together, our results show that Nurr1, Pitx3, and Lmx1a are similarly capable of enhancing the generation of TH+ cells when NP cells are appropriately primed by prior exposure to SHH and FGF8 emphasizing the importance of a proper temporal combination of signaling molecules and transcription factors for optimal generation of TH+ neurons.
To address whether Nurr1, Pitx3, and/or Lmx1a activate TH gene transcription by directly binding to the promoter motif(s), we scanned the whole mouse TH gene with an ECR browser program to identify putative binding motifs for Nurr1, Pitx3, or Lmx1a based on their respective putative-specific squences: 5′-AAAGGTCA-3′, 5′-S(C or G)NTAATCCM(C or A)-3′, and 5′-TAAT-3′, followed by ChIP analysis. We found that all three potential Nurr1-binding motifs, highly conserved in both mouse and rat TH gene promoters, were directly bound by overexpressed Nurr1 (Fig. 3), which is consistent with our previous study [39]. Of the six putative Pitx3-binding sites that are conserved between the mouse and rat TH promoters, we found that overexpressed Pitx3 binds five of them. In addition, overexpressed Lmx1a specifically bound seven sites out of eight putative binding sites. These results suggest that all three transcription factors directly and coordinately activate TH gene transcription via interaction with promoter motifs. This is supported by our co-transfection assays showing that each of these transcription factors robustly activates the reporter expression of the TH promoter–reporter construct, but not that of the DBH promoter reporter. Our findings of direct activation of the TH promoter by these transcription factors are consistent with our previous study [39] where we identified Nurr1 binding motifs and similar TH promoter activation. This is in contrast with previous studies from this and other laboratories showing that Pitx3 overexpression is not sufficient to enhance the TH gene transcription [27,28]. These differences can be explained by the expression of transcription factors at different stage of ESCs differentiation, again emphasizing the potential importance of temporal control of exogenous gene expression for its optimal effects.
A great majority (>98%) of TH+ neurons generated by overexpression of Pitx3 or Lmx1a were also Tuj1+. In contrast, a significant proportion (∼20%) of TH+ neurons generated by overexpression of Nurr1 were found in the Tuj1− population, suggesting that these transcription factors have significantly distinct functional role(s). Since the proneural marker Ngn2 is known to regulate neuronal differentiation of mDA progenitors [20,22], we speculated that Nurr1, Pitx3, and/or Lmx1a differentially regulate Ngn2 gene expression. Indeed, using the consensus binding motif, 5′-GGGATTAAC-3′, we identified a site in the upstream promoter of the Ngn2 gene that is highly conserved in mouse, human, monkey, dog, chicken, rat, and frog Ngn2 genes. Our ChIP analysis demonstrated that Pitx3 directly binds this site, suggesting that it plays a direct role in Ngn2 gene transcription. Furthermore, Pitx3 also binds to potential binding motifs of the pan-neuronal β-tubulin III gene promoter region significantly increasing its mRNA expression. Whether Lmx1a also directly activates Ngn2 gene expression is unknown and awaits further investigation.
Taken together, our results demonstrate that three key transcription factors, Nurr1, Pitx3, and Lmx1a, importantly regulate mDA neuron development and phenotype specification via overlapping as well as distinct mechanisms. In particular, our findings support the notion that Pitx3 plays multiple roles during neurogenesis and dopamine neuronal phenotype during ESC differentiation into mDA neurons, at least in part via direct binding to gene promoters and expression of TH, Ngn2, and β-tubulin III genes. These data will help further mechanistic understanding as well as inform optimal design of guided differentiation of stem cells and/or direct conversion of somatic cells into mDA neurons by a combined use of key transcription factors.
Footnotes
Acknowledgments
Supported by the National Institutes of Health Grants NS070577 and MH048866, and the Korea Ministry of Science, ICT and Future Planning Grants 2012M3A9C6050131 and the Korea Ministry of Health and Welfare Grant A120392.
Author Disclosure Statement
No competing financial interests exist.