
Abstract
In a previous study we established colony assays suitable for studying murine adult (2–4 months) pancreatic progenitor cells plated in semisolid media containing methylcellulose and extracellular matrix proteins. Using these assays, we found robust in vitro progenitor cell activities (multilineage differentiation and self-renewal) from pancreatic cells of adult mice in the CD-1 outbred background. However, it was not clear whether progenitor cell activities can be detected from inbred mice, a preferred mouse model for various genetic studies. It was also not clear whether a single cell is sufficient to self-renew. Here, we show that fluorescent activated cell sorting pancreatic CD133+ but not CD133− cells from adult C57Bl/6 inbred mice are enriched for progenitor cells that self-renew and give rise to multilineage colonies in vitro. The number of cells in a colony is in proportion to its diameter. Around 60% of single handpicked 3-week-old colonies express trilineage markers, indicating most progenitors are tripotent for ductal, acinar, and endocrine lineage differentiation. Approximately 80% of primary (freshly sorted) colony-forming progenitor cells are capable of giving rise to secondary progenitors in vitro, indicating that a majority of the primary progenitors self-renew. A single cell is sufficient for self-renewal and a Wnt agonist, R-Spondin1, enhances the number of secondary progenitors from the primary progenitors. Together, our pancreatic colony assays allow quantitative analyses of progenitors at a single-cell level from inbred mice. These assays will be useful for elucidating in vitro mechanisms of pancreatic progenitor cell biology.
Introduction
C
The essence of an in vitro colony assay is that cells in a single-cell suspension are mixed in a semisolid medium that prevents cell migration. However, the medium is still soft enough to allow a single cell to proliferate and differentiate and form a colony of cells in a three-dimensional (3D) space. By analyzing the lineage composition of a resulting colony, the lineage potential of the originating single CFU can be deduced. For example, a colony composed of granulocytes (G), erythrocytes (E), macrophages (M), and megakaryocytes (M) would indicate that the initiating progenitor cell of that colony (ie, the CFU-GEMM) is multipotential for GEMM lineages. By dissociating and replating the cells collected from a single colony, the self-renewal capacity of the initiating primary progenitor cell can be determined. It is also a quantitative assay, in which the prevalence of CFUs in a given population of cells can be calculated by dividing the number of colonies formed with the total input cells. The use of semisolid media is essential to this quantitative aspect of the hematopoietic colony assay, which is achieved by inclusion of methylcellulose, a biologically inert material derived from wood fibers.
The pancreas is composed of three major cell lineages: acinar cells, ducts, and endocrine cells. Acinar cells secrete digestive enzymes. Ductal cells secrete mucin to fend off pathogens and transport digestive enzymes to the gut. Endocrine cells, including the insulin-secreting beta cells and the glucagon-expressing alpha cells, maintain glucose homeostasis.
CD133, also known as AC133 and prominin-1, was initially identified as being expressed by hematopoietic stem and progenitor cells [4]. It has since been used extensively as a stem cell marker for adult normal and cancerous tissues [5]. CD133 is not expressed in most postnatal and adult epithelia [4], but it is present in adult pancreatic ducts of mice and humans [6 –8]. Although CD133 is expressed on the cell surface of adult human pancreatic ductal trees [8 –10], some ductal cells express CD133 in the cytoplasm [10]. This has led to the speculation that adult human CD133+ duct cells may not be progenitor cells [8,10]. While it is still unclear whether human CD133+ cells contain progenitor cells, murine CD133+ cells isolated from embryonic [11,12], neonatal [7], or adult [13] pancreas displayed progenitor cell activities in vitro.
Until lately there has been no colony assay similar to the one used for bone marrow cells to study pancreatic progenitor cells. To fill this gap, we developed in vitro pancreatic colony assays that enable quantitative and functional analyses of adult murine pancreatic progenitor-like cells, which we named pancreatic colony-forming units (PCFUs) [13]. We find that adult PCFUs robustly self-renew and differentiate in vitro. However, these PCFUs were isolated from a CD-1 outbred genetic background, and it is not clear whether PCFUs from a more stringently inbred strain of mice may also possess progenitor cell activities. In this study, we therefore begin to test one of the most popular and widely used inbred mouse strain C57Bl/6 (B6). In addition, it was not clear in our prior studies whether a single cell is sufficient to self-renew in vitro.
Here, we show that adult PCFUs from B6 mice are enriched in CD133+ but not CD133− cells. Many (∼60%) of these PCFUs are multipotential and give rise to ductal-, acinar-, and endocrine-like cells in vitro. The majority (>80%) of single PCFUs also self-renew in vitro, as assessed by dissociation of individual primary colonies and replating the dissociated cells into secondary culture. Clonality of the colonies is assured by handpicking of single colonies visualized directly under a phase-contrast microscope and plating at low cell density. Thus, similar to hematopoietic colony assays, our pancreatic colony assays will be valuable in elucidating the cellular and molecular mechanisms of pancreatic progenitor cell formation and function.
Materials and Methods
Mice
C57BL/6 (2–4 months old; body weight ranged 20–24 g) inbred background (Charles River Laboratory, Wilmington, MA) and actin/EGFP [14] mice were used. All mice were maintained under specific pathogen-free conditions, and animal experiments were conducted according to the Institutional Animal Care and Use Committee at the City of Hope.
Preparation of single-cell suspension
Murine pancreata were dissected, cleared of fat tissue under a dissecting microscope, and rinsed three times in cold D-PBS containing 0.1% bovine serum albumin (BSA), and penicillin and streptomycin (PBS/BSA). Whole pancreata, in a dry petri dish placed on ice, were chopped using spring scissors for ∼2 min or until finely minced. The triturated tissue was transferred to a 15 mL conical tube, washed once, resuspended in PBS/BSA containing collagenase B (2–4 mg/mL per pancreas; Roche, Mannheim, Germany) and DNase I (2,000 U/mL per pancreas; Calbiochem, Darmstadt, Germany), and incubated (37°C, 20–30 min) to yield the single-cell suspension. To hasten the digestion, tissue was gently disrupted every 5–10 min using a 1-mL Gilson pipettman. Cells were then washed twice in cold PBS/BSA supplemented with 2,000 U/mL DNase I, which is used to prevent reaggregation of dissociated cells, followed by filtering through a 40-μm nylon mesh (BD Biosciences, San Jose, CA).
Flow cytometry and cell sorting
Murine pancreatic single-cell suspensions were first incubated with anti-mouse CD16/32 (Clone 93; 10 μg/mL, 5 min, on ice; BioLegend, San Diego, CA) to reduce nonspecific binding. Biotin-conjugated anti-mouse CD133 (Clone 13A4, 5 μg/mL; eBioscience, San Diego, CA) or the control biotin-conjugated rat IgG1 isotype (Clone eBRG1, 5 μg/mL; eBioscience) antibody was added and cells were incubated (20 min, on ice), washed (twice), incubated with streptavidin-labeled allophycocyanin (APC) (2 μg/mL, 15 min, on ice; BioLegend), washed (twice), resuspended in PBS/BSA/DNase I containing DAPI (0.2 μg/mL), filtered through a 20-μm mesh, and kept on ice until flow cytometry analysis or sorting. Cell sorting was performed on a MoFlow™ MLS (Beckman Coulter). All analyses included an initial gating of forward and side scatters to exclude cell debris, followed by exclusion of doublets and dead cells.
Cell culture and in vitro colony assay
Cells were cultured in methylcellulose-based semisolid medium as described previously [15,16]. Briefly, cold culture mixture (1 mL) contained DMEM/F12 media, 1% 1,500 centipoise (high-viscosity) methylcellulose (Sinetsu Chemical, Tokyo, Japan), 5% Matrigel (BD Biosciences), 50% conditioned media from embryonic-stem-cell-derived pancreatic–like cells, 5% fetal calf serum, 10 mM nicotinamide (Sigma, St. Louis, MO), 10 ng/mL human recombinant activin-B, 0.1 nM exendin-4, and 1 ng/mL vascular endothelial growth factor-A (R&D Systems, Minneapolis, MN). Cells were plated in 24-well ultralow protein-binding plates and incubated in a humidified 5% CO2 atmosphere. Triplicate wells were routinely plated. Three weeks after culture, Ring colonies were numerated using an inverted microscope with a 10×objective lens. For microfluidic quantitative reverse transcription-polymerase chain reaction analysis, individual colonies were lifted from the methylcellulose medium by utilizing a 10-μL pipette under direct microscopic visualization. For replating experiments, individual colonies were handpicked, and single-cell suspensions were generated through incubation (37°C, 5 min) with 0.25% trypsin-EDTA, followed by gentle pipetting. Single cells were then washed and replated in a secondary Matrigel colony assay as described above. When indicated, murine recombinant R-Spondin1 (RSPO1; R&D Systems) was used at 750 ng/mL.
Conventional quantitative RT-PCR and microfluidic qRT-PCR analyses
Total RNA extraction, reverse transcription, and conventional qRT-PCR analyses were as described [17]. Taqman probes (Life Technologies, Grand Island, NY) and their catalog numbers are listed in Supplementary Table S1 (Supplementary Data are available online at
Whole-mount immunostaining
Colonies were manually picked and fixed in 4% PFA at 4°C overnight. Colonies were incubated with blocking buffer with 5% donkey serum and 0.1% triton at 4°C overnight. Primary antibodies used were as listed in the Supplementary Table S2 and were detected with donkey-raised secondary antibodies conjugated to Cy3, Cy5, or DyLight488 (Jackson Immunoresearch, West Grove, PA) at 1:2,000 dilution. Images were captured on a Zeiss LSM510 META NLO Axiovert 200M inverted microscope, and figures were prepared with LSM Image Browser software (Carl Zeiss, Oberkochen, Germany).
Transmission electron microscopy
Single colonies were collected, pooled, and fixed in 2% glutaraldehyde in 0.1 M Cacodylate buffer [Na(CH3)2AsO2·3H2O; pH7.2] at 4°C overnight. Colonies were washed three times with 0.1 M Cacodylate buffer (pH 7.2), postfixed with 1% OsO4 in 0.1 M Cacodylate buffer for 30 min, and washed three times with 0.1 M Cacodylate buffer. The samples were then dehydrated, embedded in Eponate, and polymerized at ∼64°C for 48 h. Ultrathin sections (∼70-nm thick) were cut using a Leica ultramicrotome with a diamond knife, picked onto 200-mesh EM grids, and were stained with 2% uranyl acetate in 70% ethanol for 1 min followed by Reynold's lead citrate staining for 1 min. Electron microscopy was carried out on a FEI Tecnai 12 transmission electron microscope equipped with a Gatan Ultrascan 2K CCD camera.
Statistical analysis
All values are shown as mean±standard deviation. P values were calculated using Student's two-tailed t-test. P<0.05 was considered significant.
Results
Dissociated adult pancreata of B6 mice form cystic “Ring” colonies in 3D culture
To obtain live single cells, a whole pancreas was minced and digested with collagenase B and DNase I to yield a mostly single-cell suspension. To obtain highly viable pancreatic cells, we found that it was necessary to clean the fat tissue from the pancreas during the dissection and kept the cells cold during the whole process except for the enzyme digestion. Preparations routinely yielded 94.5%±6.4% (range 82.5%–99.5%) live cells, as determined by gating on FSC, SSC, and DAPI negative staining (Fig. 1A, B), of which 97.7%±0.9% (range 96.6%–99.7%) were singletons (Fig. 1C). Single-cell suspensions were then mixed in the Matrigel-containing semisolid media and cultured. After 2–3 weeks, cystic colonies that we named “Ring” were formed (Fig. 2A), consistent to the finding from CD-1 strain [13]. The single pancreatic cell that gave rise to a Ring colony was termed as PCFU-Ring. Each Ring colony contained an average of 100–250 cells, depending on the size of the colony (Fig. 2B). The log diameter of a Ring colony was positively and linearly correlated with the number of cells in that colony (Fig. 2B). This may suggest that the size of a Ring colony was indicative of the proliferative potential of the originating PCFU-Ring. Alternatively, the different colony sizes may indicate a variable delay of the cells that enter replication or time needed for reprogramming to occur [18,19].
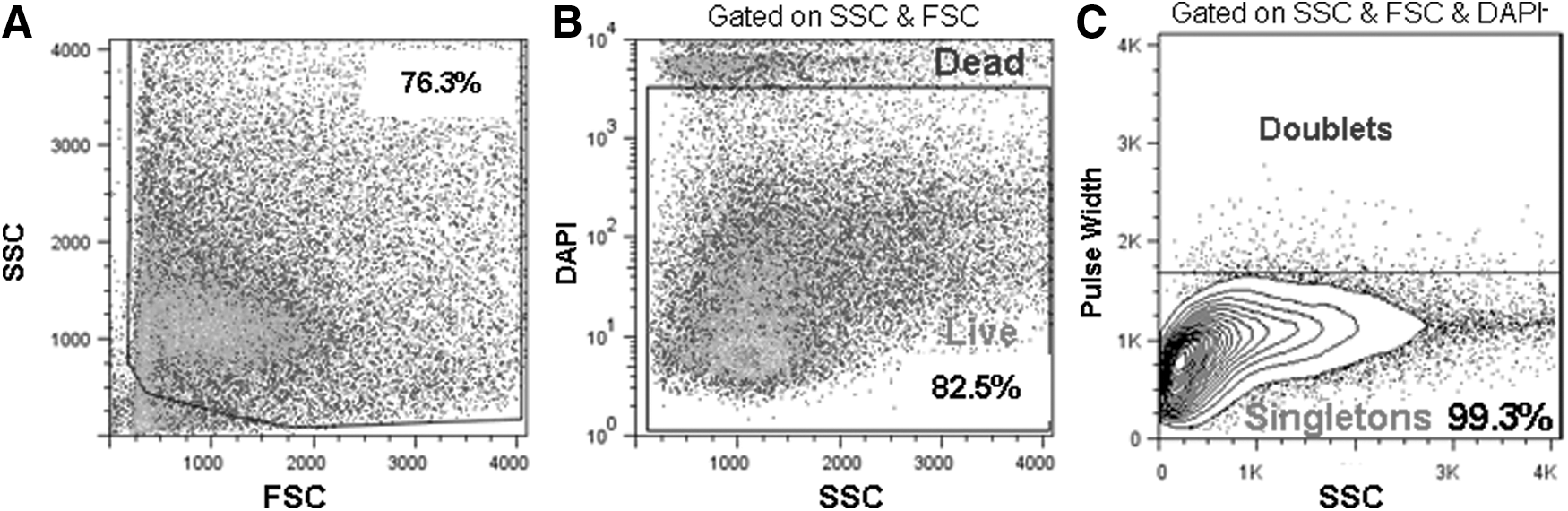
Dissociation of pancreata of adult B6 mice into viable single-cell suspension. Representative flow cytometry analyses of dissociated pancreatic cells from adult (2–4 months old) B6 mice.
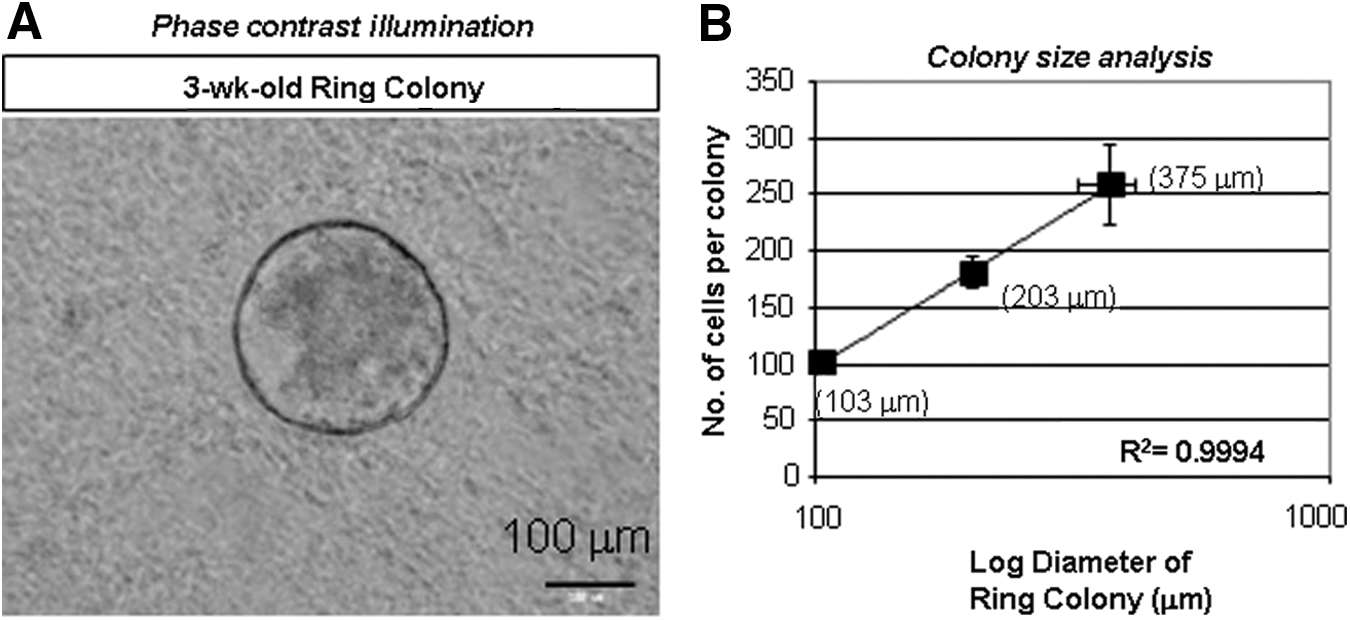
Dissociated adult B6 pancreatic cells give rise to Ring colonies after 3 weeks in culture.
Freshly sorted CD133+ cells express higher levels of ductal cell markers and lower levels of endocrine and acinar cell markers
Adult (2–4 months old) B6 pancreatic CD133+ cells represented 13.1%±4.3% (range 6.5%–17.3%, n=7) of the total population (Fig. 3A), which corresponded to the estimated proportion of ductal cells in the pancreas [20]. Sorted CD133+ cells were enriched for ductal transcripts encoding CD133, CK7, Mucin1, and Sox9 (Fig. 3B), compared with presorted cells. In contrast, CD133− cells were enriched for other lineage-specific markers, such as acinar (Amylase2A) or endocrine (Glucagon, Insulin1, and Insulin2) (Fig. 3C). These results were in agreement with previous studies, including ours, suggesting a ductal identity of CD133+ cells [6 –8,13].
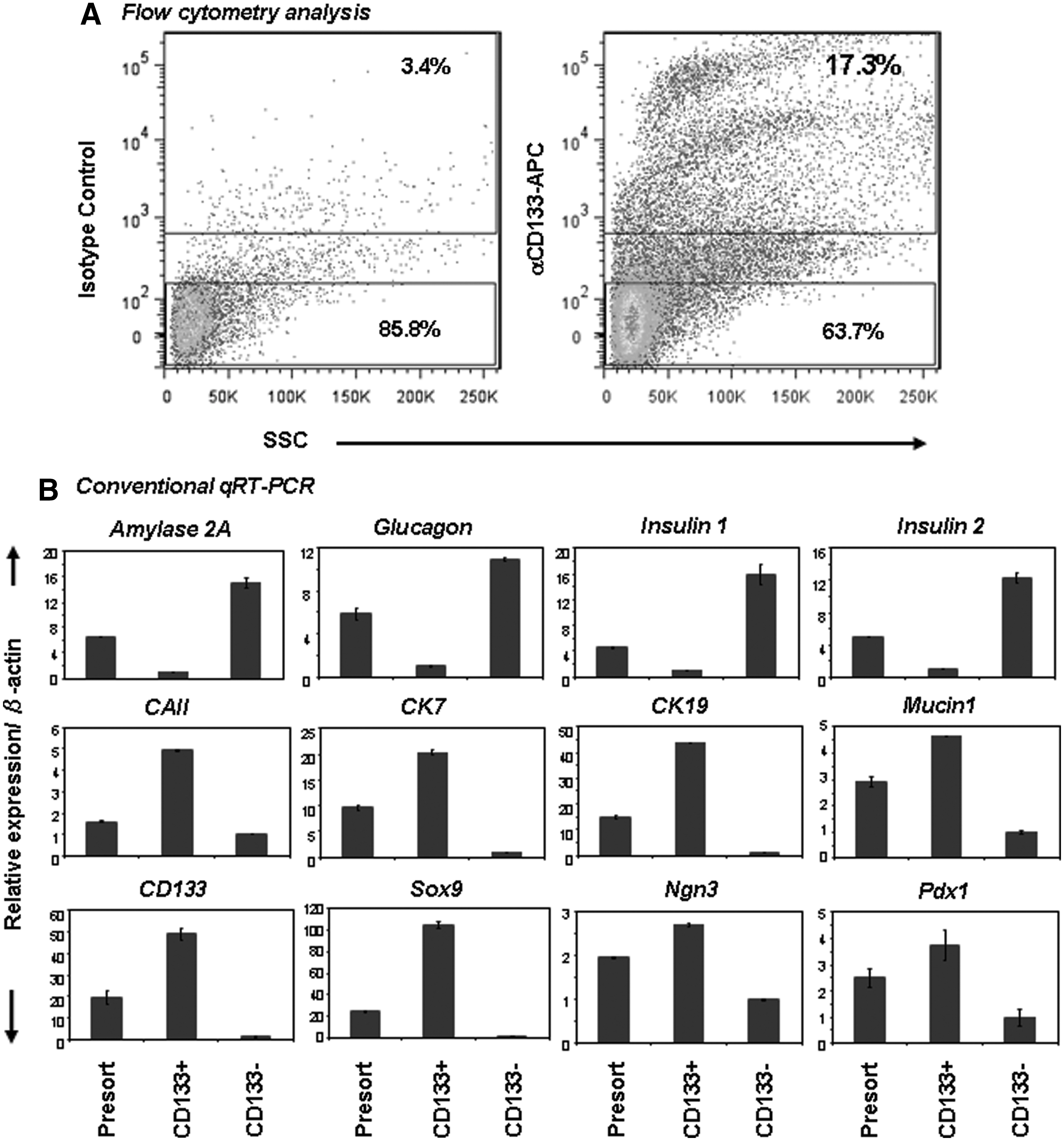
Freshly sorted adult B6 CD133+ cells express ductal cell markers. Cells were stained with anti-CD133 antibody and sorted by a fluorescence-activated cell sorter.
Pancreatic CD133+ cells are enriched for ring colony-forming progenitor cells
Dissociated adult B6 murine pancreata were then sorted based on CD133 expression, plated, and examined for colony formation. After 3 weeks, Ring colonies were found only in wells seeded with CD133+ but not with CD133− cells (Fig. 4A), consistent to our earlier preliminary finding [13]. Colony-forming efficiency (number of colony divided by total input cells) was 4.3%±2.0% (ranged 2.9%–6.9%, n=5) from CD133+ cells, which was 5.1±0.8-fold enriched for PCFUs-Ring compared with presorted cells (Fig. 4B). Colony-forming efficiency of the presorted cells is ∼1% from B6 pancreas, consistent to the results from CD-1 mice [13].
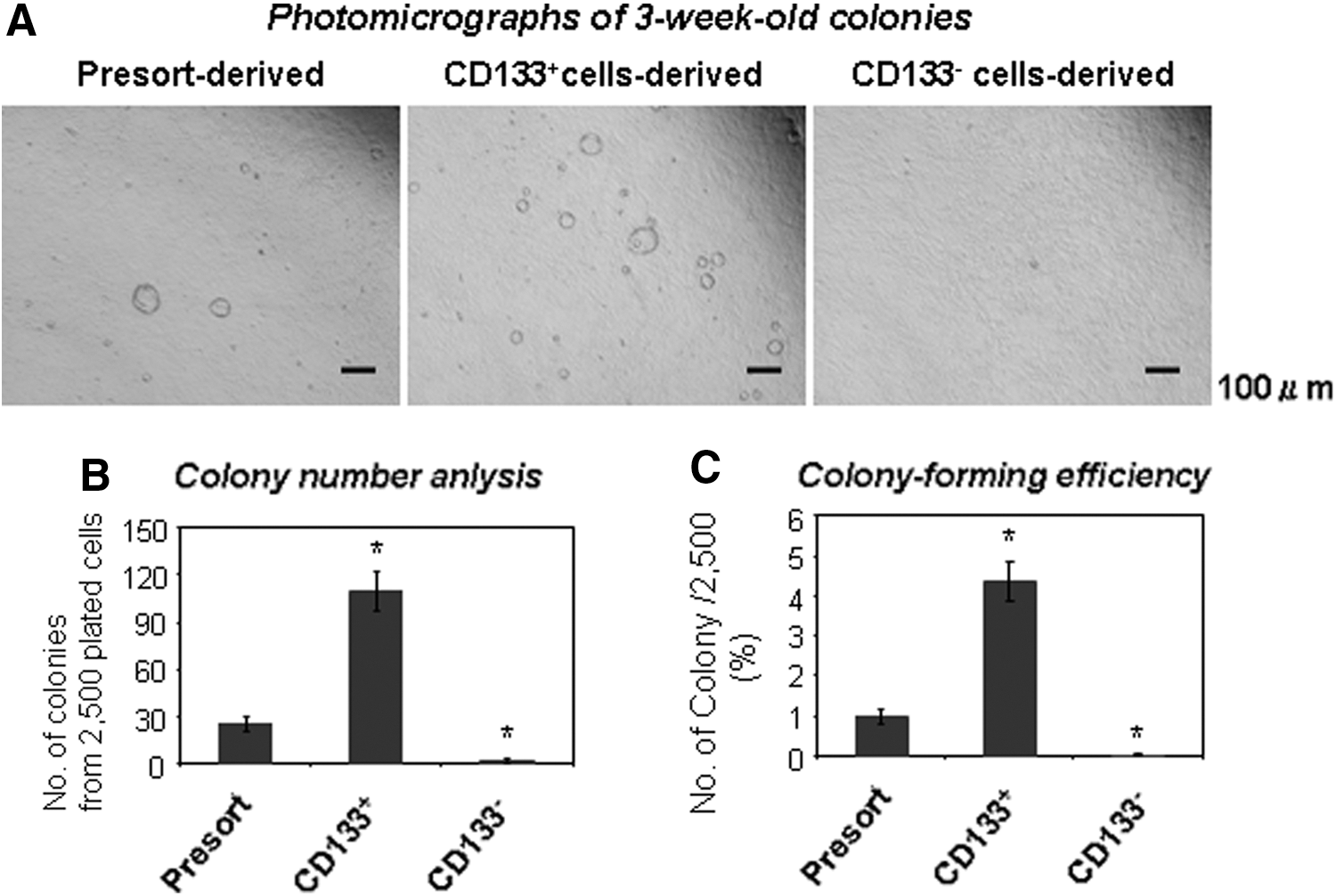
Adult B6 CD133+ but not CD133− cells are enriched for PCFUs-Ring.
Optimal plating density is 5,000 CD133+ cells/24-well/0.5 mL
To determine appropriate seeding cell density, pancreatic CD133+ cells from chicken actin/EGFP transgenic mice and from wild-type litter mates were sorted, and mixed at a 1:1 ratio of graded cell doses. The actin/EGFP transgenic mice are in C57BLKa-Thy1.1 inbred background [21] that is closely related to B6. A colony composed of a mixture of EGFP- and non-EGFP-expressing cells (Fig. 5A; right panel) would indicate that the originating cells may come from at least two seeded PCFUs-Ring positioned close together. We found that such mixed colonies were less common as plating cell density decreased (Fig. 5B). At cell densities equal to 10 CD133+ cells/μL (or 5,000 CD133+ cells/well/0.5 mL) (Fig. 5B; 2.5K+2.5K), <1% of the total colonies were mixed, suggesting that the majority of colonies formed under this condition were clonally derived. To ensure the clonality of the colonies in subsequent studies, we therefore plated only 2,500 CD133+ cells per well, which will result in ∼100 colonies per well (Fig. 4B).
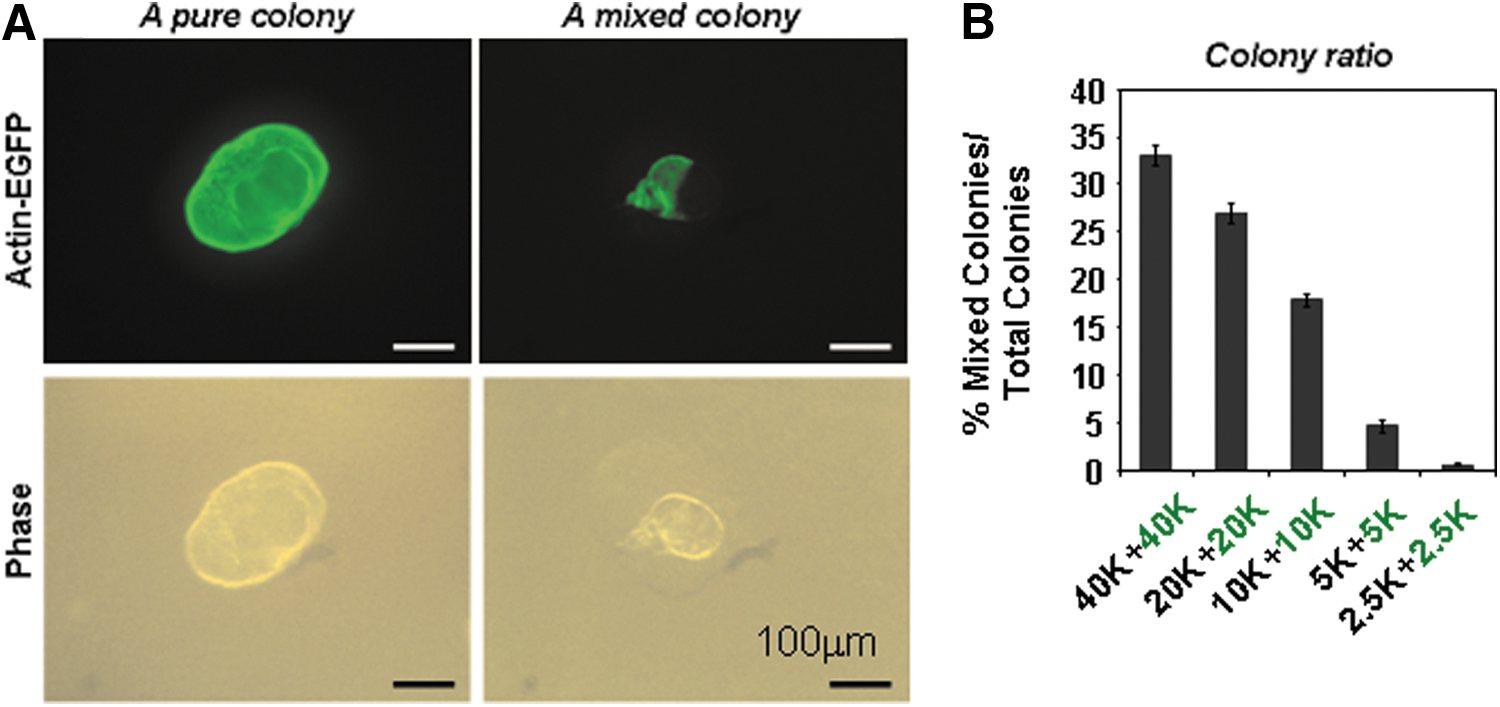
Minimal clonal density requirements. A plating density of <5,000 CD133+ cells/0.5 mL/well was required for clonal formation. CD133+ cells were sorted from actin-EGFP mice or wild-type littermates (C57BLKa-Thy1.1 inbred background) and plated in colony assay for 3 weeks.
Many PCFUs-Ring are multipotential and give rise to multilineage colonies
To determine lineage composition of individual Ring colony, colonies were directly visualized under the phase-contrast illumination, handpicked one by one, and analyzed by microfluidic qRT-PCR (Fig. 6A, B). Microfluidic qRT-PCR is a relatively new technology that allows reaction volumes in the nanoliter range, thus enabling detection of gene expression from a small number of cells, such as a single colony [22]. Individual Ring colonies (n=9) were handpicked at week 3 and analyzed by a panel of TaqMan probes for housekeeping (β-actin), ducts (CD133, Sox9, Mucin1, Carbonic anhydrase II, and CK7), endocrine (Insulin 2 and Glucagon), and acinar cells (Amylase 2A and carboxypeptidase A1). We found that all of the colonies examined expressed duct markers, suggesting that all PCFUs-Ring have potential for ductal lineage differentiation. In contrast, five out of nine colonies expressed both acinar and endocrine genes (colonies No. 1, 3, 7, 8, and 9), suggesting that only a subpopulation of PCFUs-Ring was tripotent or that the probability of making a successful transition to the endocrine/acinar lineages is limited due to the particular extracellular matrix proteins used [13]. Transmission electron microscopy of 3-week-old Ring colonies showed cells with microvilli and multilobed nucleus—hallmarks of ductal cells (Fig. 6C). Intracellular granules that resemble zymogen of acinar cells (Fig. 6D) or insulin of beta cells (Fig. 6E) were detected. Whole-mounted immunostaining of handpicked colonies further confirmed protein expression of C-peptide (a surrogate marker for insulin) (Fig. 6F), glucagon (Fig. 6G), or amylase (Fig. 6H). Finally, protein markers that represent ductal (CK19), acinar (Amylase), and endocrine (Glucagon) lineages were found expressed in one single colony (Fig. 6I), unequivocally demonstrating the trilineage potential of the originating PCFU-Ring. Taken together, these results demonstrate that many adult PCFUs-Ring from adult B6 mice are capable of multilineage differentiation in vitro.
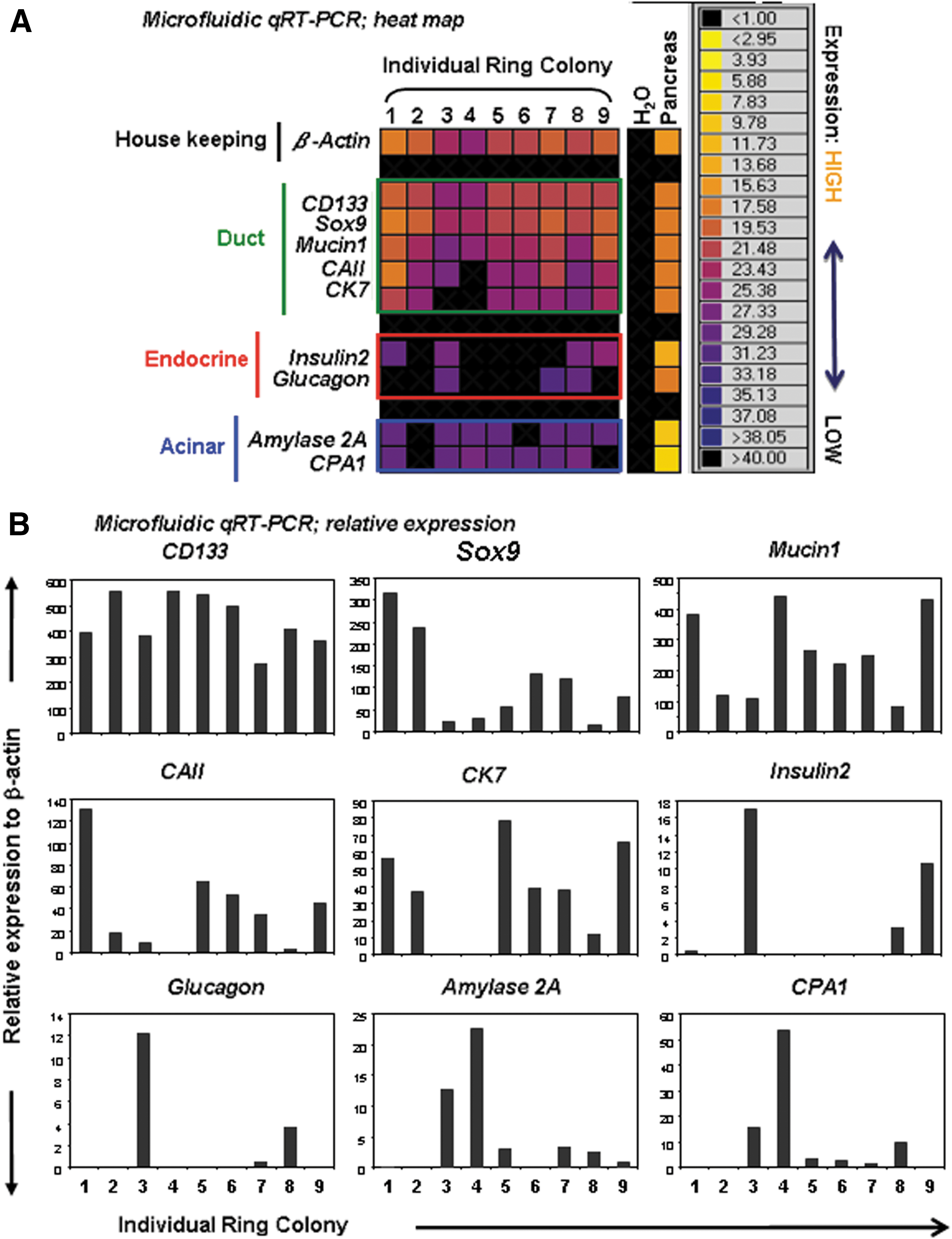
Lineage composition analysis of individually handpicked B6 Ring colonies.

Single PCFU-Ring self-replicate in vitro
In our previous studies using CD-1 mice, Ring colonies were serially dissociated and replated over a total of 5 generations in culture, and more than 10,000-fold expansion of PCFUs-Ring was observed [13]. Because total colonies from each generation were collected and replated, it was not clear whether each single PCFU-Ring may possess self-renewal ability. To address this, individually handpicked primary B6 Ring colonies (n=5) were dissociated into single-cell suspensions and replated into a respective secondary well for colony formation (Fig. 7A). Four out of the five colonies examined formed secondary colonies (Fig. 7B), but the ability of primary PCFUs-Ring to self-replicate varied, as evidenced by a wide range in the number of secondary progenitor cells generated (Fig. 7B). Because of the low plating density, each single colony is assumed to be derived from a single PCFU-Ring in our culture condition (Fig. 5). Together, these findings suggest that the freshly isolated PCFUs-Ring is heterogeneous in their potential to self-replicate under our typical culture condition. Wnt signaling molecules increase self-renewal of various adult stem cells [23 –25]. RSPO1 is a recently discovered Wnt ligand [26]. It was also the reagent that caused the exponential expansion of PCFUs-Ring from CD-1 mice [13]. We therefore tested the effects of exogenous recombinant RSPO1 on B6 cells. All 10 micromanipulated colonies stimulated by 750 ng/mL RSPO1 formed secondary colonies, while only 8 out of 10 colonies from the control culture formed secondary colonies (Fig. 7C). The average number of secondary colonies was increased by RSPO1 by 8.6-fold (Fig. 7C; 179.9±72.5 RSPO1-treated group versus 20.8±20.7 control group). Together, these results demonstrate that ∼80% of the freshly sorted PCFUs-Ring showed the self-renewal ability in the “base” media, and exogenous RSPO1 enhanced that self-renewal ability.
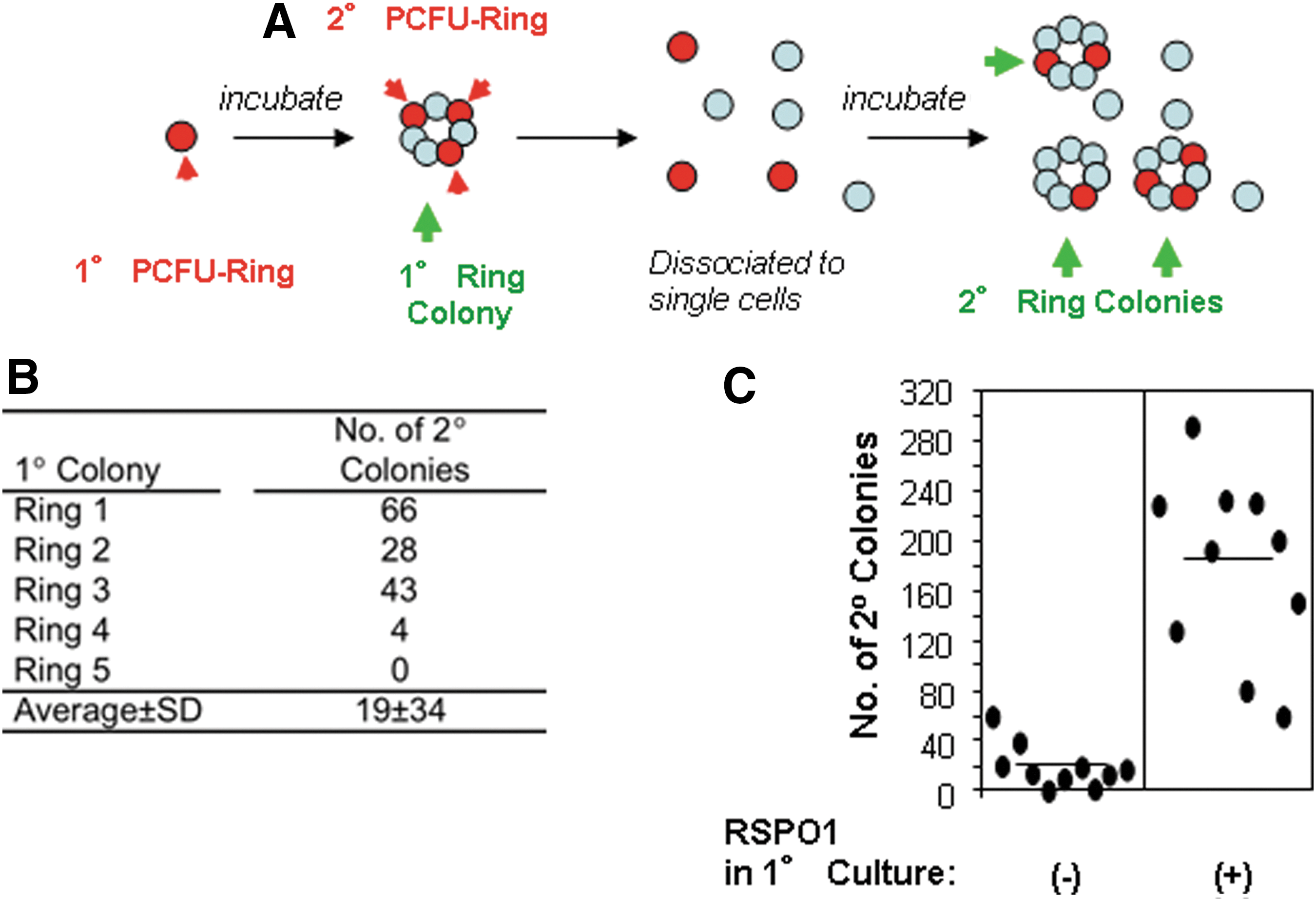
Single primary PCFU-Ring self-renews and generates secondary PCFUs-Ring in vitro.
Discussion
In this study we showed that ∼60% of single PCFUs-Ring have a duct potential plus at least some endocrine or acinar cell lineage potentials, and 80%–100% can initiate secondary colonies upon replating. These results suggest that many PCFUs-Ring are multipotential progenitors, and they can self-renew and form secondary colonies. The frequency of the PCFUs-Ring is ∼1% of the total pancreatic cells from adult B6 mice. These results are consistent to our earlier studies conducted on CD-1 outbred mice [13] and demonstrate that adult pancreatic cells contain a rare population of cell that possesses multilineage differentiation and self-renewal activities in vitro. However, it remains to be determined whether a single pancreatic progenitor cell may self-renew while maintaining ability for multilineage differentiation, which are the ultimate defining properties of stem cells. To test this, one may employ a retroviral vector to label single PCFUs before transplantation, and then follow their cell fate long-term in vivo. Such strategy has been used in identifying long-term reconstituting hematopoietic stem cells in the past [27].
One unique characteristic of our pancreatic colony assay is the mixing of methylcellulose with extracellular matrix proteins, such as Matrigel used in the current study. Matrigel contains various extracellular matrix proteins that are important for cellular function [28]. Methylcellulose but not Matrigel is included in the traditional hematopoietic colony assays. Other existing culture assays for epithelial cells employ Matrigel but not methylcellulose. In those assays, epithelial cells of gut [29], liver [30], or pancreas [31,32] are mixed in high concentrations (>60% vol/vol) of Matrigel, which is required to render a 3D culture environment. Subsequently, liquid media are layered over on top of the Matrigel. The liquid media are required to be changed every 2–3 days to maintain the health of the cells. In contrast, our Matrigel and methylcellulose semisolid media contain only 5% (vol/vol) Matrigel and still maintain a 3D milieu. This will enable dose–response studies of various ECM proteins on PCFUs in future studies. As in hematopoietic colony assay, no liquid media are involved, and thus no media change is required for at least 3 weeks in our pancreatic colony assays. Thus, our assay is easier to perform and maintain.
The second unique feature of our pancreatic colony assay is the ease with which self-renewal properties can be assessed. Reports on in vitro self-renewal of pancreatic progenitor cells have been scarce. This could be due to technical difficulties of the existing assay systems. One commonly used culture system to address self-renewal is the “pancreatosphere” assay [33 –35], which was originally adapted from the “neurosphere” assay [36,37]. In this assay, progenitor cells are cultured in liquid media and they form a sphere of cells. When collected, dissociated, and replated into another liquid culture, secondary spheres form. However, due to the fact that cells move freely in liquid media, the clonality of the spheres cannot be ensured. To ascertain clonality, limiting dilution or single-cell deposition into large number of 96-wells is required, but it can be technically challenging and time and labor consuming. Other reported self-renewal assay for pancreatic cells uses 2D attachment culture in liquid media and clonality ensured by limiting dilution [38,39], which again is a labor and time intensive exercise. In contrast, our assay allows the functional analysis of thousands of plated cells per experiment. In addition, the clonality of colonies in our assay can be ensured by low plating cell density (Fig. 5) or micromanipulation of single colonies (Fig. 7) or single cells [13].
There have been intense debates about whether the adult pancreas in vivo contains “facultative” ductal progenitor cells [40]. Using strategies such as Cre-lox lineage tracing [41] to label adult ductal cells with CAII [42], HNF1b [43], or Sox9 [44,45] or other techniques, both positive [42,46] and negative [43,44,47,48] results have been obtained. The word “facultative” is referring to the occasional trans-differentiation property of the mature ductal cells to a progenitor cell phenotype [49]. However, based on our current and previous data [13] we now propose an alternative hypothesis: adult ductal cells are heterogeneous and only a small subset of the ductal cells have either (1) progenitor cell characteristics or (2) remain sufficiently plastic to be reprogrammed or trans-differentiated. If this hypothesis is correct, then labeling cells using a pan-duct marker, such as HNF1b and Sox9, will miss the rare cells' activities in vivo. On the other hand, we cautiously note that our current and previous studies [13] only demonstrate the in vitro but not in vivo actions of adult pancreatic progenitor cells. It is possible that self-renewal and multilineage differentiation of adult PCFUs may represent an in vitro phenomenon because the cells have been displaced out of their natural environment in vivo. More studies are required to interrogate this hypothesis.
Due to the popularity of B6 mice, many reagents, tools, and data bases are created and suited for studies in this inbred strain of mice. For example, many genetic mouse models are maintained in the B6 background, making it possible to study the effects of specific genes on PCFU biology in normal physiology and disease models in future studies. In addition, transplantation studies can be done in syngeneic mice, thus avoiding the use of costly immune-deficient mice as recipients. With the ability to fluorescent activated cell sorting and enrich PCFUs-Ring, we envision that specific markers will be identified in ductal cells that have progenitor-like properties compared with those that do not. This will also facilitate future in vivo analysis for the rare cells.
Footnotes
Acknowledgments
The authors thank Lucy Brown, Alexander Spalla, and Pavinder Kaur from the Analytical Cytometry Core of City of Hope for assistance in flow sorting. This work is supported in part by National Institute of Health (NIH) grant R01DK081587 to H.T.K.; U01DK089533 to A.D.R.; R00 HL087936, Children Leukemia Research Association, ThinkCure! Foundation, Margaret E. Early Medical Research Trust, and the Tim Nesvig Lymphoma Research Fund to C.-C.C.; and Office of Naval Research ONR-N00014-02-1 0958 and National Science Foundation NSF-DBI-9970143 to electron microscopy core facility at City of Hope.
Author Disclosure Statement
The authors have nothing to disclose.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.