
Abstract
Vascular endothelial growth factor (VEGF) is reported to exhibit potent hematopoietic stem/progenitor cell (HSPC) mobilization activity. However, the detailed mechanisms of HSPC mobilization by VEGF have not been examined. In this study, we investigated the effect of VEGF on bone marrow (BM) cell and the BM environment by intravenous injection of VEGF-expressing adenovirus vector (Ad-VEGF) into mice. A colony assay using peripheral blood cells revealed that plasma elevation of VEGF leads to the mobilization of HSPCs into the circulation. Granulocyte colony-stimulating factor (G-CSF) is known to mobilize HSPCs by decreasing CXC chemokine ligand 12 (CXCL12) levels in the BM. However, we found almost no changes in the CXCL12 levels in the BM after Ad-VEGF injection, suggesting that VEGF can alter the BM microenvironment by different mechanisms from G-CSF. Furthermore, flow cytometric analysis and colony forming unit-fibroblast assay showed a reduction in the number of mesenchymal progenitor cells (MPCs), which have been reported to serve as niche cells to support HSPCs, in the BM of Ad-VEGF-injected mice. Adhesion of donor cells to the recipient BM after transplantation was also impaired in mice injected with Ad-VEGF, suggesting a decrease in the niche cell number. We also observed a dose-dependent chemoattractive effect of VEGF on primary BM stromal cells in vitro. These data suggest that VEGF alters the distribution of MPCs in the BM and can also mobilize MPCs to peripheral tissues. Taken together, our results imply that VEGF-elicited egress of HSPCs would be mediated, in part, by changing the number of MPCs in the BM.
Introduction
H
Not only is the vascular endothelial growth factor (VEGF) a well-known factor in angiogenesis, but it also plays an important role in the growth and differentiation of hematopoietic cells. Homozygous or heterozygous deletion of VEGF in mice leads to early embryonic lethality because of impaired vascular angiogenesis and hematopoiesis [11,12]. By conditional deletion of VEGF in hematopoietic cells, but not in stromal cells, Ferrara and colleagues clearly showed that VEGF is required for survival and repopulation of adult HSCs [13]. Furthermore, VEGF has been shown to be an essential factor for HSC niche formation through endochondral ossification [14]. These observations clearly demonstrate that VEGF exerts physiological actions on hematopoietic systems through both cell-autonomous and -nonautonomous mechanisms.
In addition to the functions described above, VEGF also has a potent HSPC mobilization capacity [15], although the mechanisms of VEGF-induced HSPC mobilization have not been addressed in detail. In the current study, we investigated the effect of VEGF on the BM cell mobilization and BM environment after the intravenous injection of VEGF-expressing adenovirus (Ad) vector (Ad-VEGF) into mice. The results showed that VEGF overexpression in mice could lead to a reduction of not only the HPSC number, but also the MPC number in the BM. We also observed an enhanced chemoattractive activity of BM stromal cells by VEGF. Our data suggest that the plasma elevation of VEGF in mice alters the distribution of MPCs in the BM, and this might cause HSPC egress from the BM.
Materials and Methods
Ad vectors
Ad vectors were constructed by an improved in vitro ligation method [16,17]. The mouse VEGF165 cDNA and human G-CSF cDNA were obtained from pBLAST49-mVEGF and pORF9-hGCSFb, respectively (Invivogen). Each cDNA was cloned into a multicloning site of pHMCMV10 [18,19], which contains the cytomegalovirus (CMV) promoter/enhancer and intron A sequence flanked by the I-CeuI and PI-SceI sites, thereby resulting in pHMCMV10-VEGF and pHMCMV10-G-CSF. pHMCMV10-VEGF and pHMCMV10-G-CSF were digested with I-CeuI/PI-SceI and ligated into I-CeuI/PI-SceI-digested pAdHM41-K7 (C) [20], resulting in pAd-VEGF and pAd-G-CSF, respectively. To generate the virus, Ad vector plasmids were digested with PacI and purified by phenol–chloroform extraction and ethanol precipitation. Linearized DNAs were transfected into 293 cells with SuperFect (Qiagen) according to the manufacturer's instructions. The viruses were amplified in 293 cells. Before virus purification, the cell lysates were centrifuged to remove cell debris and were digested for 30 min at 37°C with 200 μg/mL DNase I and 200 μg/mL RNase A in the presence of 10 mM MgCl2. Viruses were purified by CsCl2 step gradient ultracentrifugation followed by CsCl2 linear gradient ultracentrifugation. The purified viruses were dialyzed against a solution containing 10 mM Tris-HCl (pH 7.5), 1 mM MgCl2, and 10% glycerol and were stored at −80°C. The control vector, Ad-Null, is similar in design, except that it contains no transgene in the expression cassette. The biological titers [infectious unit (ifu)] of Ad-VEGF, Ad-G-CSF, and Ad-Null were determined by using an Adeno-X Rapid Titer kit (Clontech).
Administration of Ad vectors in mice
C57BL/6j female mice aged 7–9 weeks were obtained from Nippon SLC, and all animals were maintained under specific pathogen-free conditions. Each Ad vector was intravenously injected into C57BL/6j mice at 1×109 ifu through the tail vein. All experiments were conducted according to the institutional ethics guidelines for animal experimentation of the National Institute of Biomedical Innovation.
Cell preparation
Blood and BM were harvested from mice using standard methods on day 5 after injection of Ad vector into mice, and the number of nucleated cells in these tissues was then counted using a Nuclecounter (Chemometec). To collect the nonhematopoietic cells from the femur and tibia, the bone fragments were minced with scissors, and were then incubated at 37°C with a type I collagenase (3 mg/mL; Worthington) in the Dulbecco's modified Eagle's medium (DMEM) with 10% fetal bovine serum (FBS) for 90 min [21]. The cells were filtered with a cell strainer to remove debris and bone fragments, and suspended in a staining buffer [phosphate buffer saline (PBS)/2% FBS]. These cell suspensions were kept on ice for further analysis.
Flow cytometry
The following antibodies (Abs), conjugated with fluorescein isothiocyanate (FITC), phycoerythrin (PE), allophycocyanin (APC), or PECy7, were used for flow cytometric analysis and cell sorting: biotinylated lineage cocktail [CD3 (145-2C11), B220 (RA3-6B2), Gr-1 (RB6-8C5), CD11b (M1/70), Ter119 (Ter-119)], c-Kit-APC (2B8), Sca-1-PECy7 (D7), Ter119-FITC (Ter-119), CD45-FITC (30-F11), CD11b-FITC (M1/70), Gr-1-PE (RB6-8C5), CD31-FITC (390), CD31-APC (390), CD51-PE (RMV-7), PDGFRa-APC (APA-5), Flt-1-PE (141522), Flk1-PE (Avas12a1), and Alcam-PE. For detection of biotinylated Abs, PerCP-Cy5.5- or FITC-conjugated streptavidin was used. Abs were purchased from e-Bioscience, BD Bioscience, Biolegend, and R&D Systems. Cells were incubated with primary Abs at 4°C for 30 min and washed twice with PBS/2% FBS. After staining, cells were analyzed and isolated by flow cytometry on an LSR II and FACSAria flow cytometer, respectively, using FACSDiva software (BD Bioscience).
Enzyme linked immunosorbent assay
Blood samples were collected through the inferior vena cava on day 5 after Ad vector injection, and transferred to polypropylene tubes containing heparin. Plasma was harvested by centrifugation. The BM supernatant was obtained by flushing a femur with 500 μL of PBS, followed by centrifugation at 500g for 5 min. The levels of VEGF and CXCL12 in the plasma and BM supernatant were measured using a commercial ELISA kit (R&D Systems) following the manufacturer's instructions.
Reverse transcription-polymerase chain reaction analysis
CD45-negative(−) Ter119− nonhematopoietic cells in the BM were sorted from mice injected with Ad-VEGF or Ad-Null, and total RNA was then extracted using ISOGEN (Nippon Gene). cDNA was synthesized from DNase I-treated total RNA with a Superscript VILO cDNA synthesis kit (Invitrogen), and quantitative real-time reverse transcription-polymerase chain reaction was performed using the Fast SYBR Green Master Mix with an ABI StepOne Plus system (Applied Biosystems). Relative quantification was performed against a standard curve and the values were normalized against the input determined for the housekeeping gene, glyceraldehyde 3-phosphate dehydrogenase. The sequences of the primers used in this study are listed in Table 1.
Gapdh, glyceraldehyde 3-phosphate dehydrogenase; cdh2, N-cadherin; Vcam-1, vascular cell adhesion molecule-1; Angpt1, angiopoietin-1; Thpo, thrombopoietin.
Colony assay
BM cells (2×104 cells) and peripheral blood cells (2×105 cells) were plated in the Methocult M3434 medium (StemCell Technologies, Inc.). Cultures were plated in duplicate and placed in a humidified chamber with 5% CO2 at 37°C for 10 days. The number of individual colonies was counted by microscopy. The colony number was normalized to the total number of the nucleated cells.
Colony forming unit-fibroblast assay
BM-derived CD45-Ter119- cells were added to the MesenCult MSC Basal Medium, including supplements (Stem Cell Technologies, Inc.), and then plated on a six-well plate at 1×105 cells per well. Cells were cultured for 14 days and stromal cell colonies (fibroblast-like colonies: >50 cells) derived from colony forming unit-fibroblasts (CFU-Fs) were stained with the Giemsa solution (Wako) after fixation with methanol. The colony number was counted by microscopy.
Cell migration assay
BM-derived stromal cells, including MPCs, were tested for migration toward VEGF using 8-μm pore-sized cell culture inserts (BD Falcon). Stromal cells (1×105 cells) resuspended in 200 μL of DMEM/2% FBS were added to the upper chamber, and 750 μL of DMEM/2% FBS containing recombinant mouse VEGF (10 or 100 ng/mL; Peprotech) was added to the bottom chamber. After 6 h of incubation at 37°C, the upper side of the filters was carefully washed with PBS, and cells remaining on the upper face of the filters were removed with a cotton wool swab. The filters were fixed with 100% methanol and stained with the Giemsa solution. Cells migrating into the lower compartment were counted manually in three random microscopic fields (×200).
Homing assay
Mice were administrated with Ad-Null or Ad-VEGF at 1×109 ifu. Five days later, BM cells (1×107 cells) derived from green fluorescent protein (GFP)-expressing transgenic mice [22] were intravenously transplanted into Ad-Null- or Ad-VEGF-injected mice. At 16 h after BM transplantation, the percentage of GFP-expressing donor cells in the BM was determined by flow cytometry.
Results
Effect of systemic VEGF overexpression on the distribution of myeloid cells and HSPCs in mice
To evaluate the effect of VEGF on the mobilization of hematopoietic cells, we generated a VEGF-expressing Ad vector, Ad-VEGF, because plasma VEGF levels were rapidly decreased with a t 1/2 of ∼25 min after intravenous injection of recombinant VEGF [23]. Single intravenous injection of Ad-VEGF (1×109 ifu) into mice led to a significant elevation of VEGF levels in plasma on day 5 compared with Ad-Null-injected mice (control mice) (Fig. 1a). On the other hand, unexpectedly, BM VEGF levels in the Ad-VEGF-injected mice were almost equivalent to those in the Ad-Null-injected mice (Fig. 1b). There were no signs of toxicity in mice treated with Ad-VEGF and Ad-Null at this dose (1×109 ifu). To investigate whether the hematopoietic cells could be mobilized from the BM into the circulation after injection of Ad-VEGF, we examined the number of total nucleated cells, myeloid cells (Gr-1+CD11b+ cells), and HSPCs [c-Kit+Sca-1+Lineage− (KSL) cells or CFU-granulocyte, erythroid, macrophage, megakaryocyte/CFU-Mix] in the peripheral blood. Compared with Ad-Null-injected mice, Ad-VEGF-injected mice showed an increased number of total nucleated cells and myeloid cells in the peripheral blood (Fig. 1c, d). We found that the number of multipotent hematopoietic progenitor cells, CFU-GEMM/CFU-Mix, in the blood of Ad-VEGF-injected mice was four times as great as that of Ad-Null-injected mice (Fig. 1e). Importantly, in Ad-VEGF-injected mice, the number of KSL cells in the blood was also increased (Fig. 1f). These results indicate that hematopoietic cells, including immature hematopoietic cells with colony-forming potentials, would be mobilized from the BM following systemic Ad-VEGF administration.

Effect of vascular endothelial growth factor (VEGF) on the number of myeloid cells and hematopoietic stem/progenitor cells (HSPCs) in peripheral blood.
An increased number of mobilized cells in VEGF-treated mice were reported previously [15], but little is known about the effect of VEGF on BM cells during the mobilization period. Thus, we next investigated the number of total BM cells, myeloid cells, and HSPCs. In contrast to the peripheral blood, the number of total hematopoietic cells, myeloid cells, and CFU-Mix was significantly decreased (Fig. 2a–c). It is of note that the VEGF overexpression in mice resulted in the reduction in both the frequency and the absolute number of KSL cells in BM (Fig. 2d). Thus, these data suggest that VEGF exerts a physiological effect on the various types of cells within the BM.
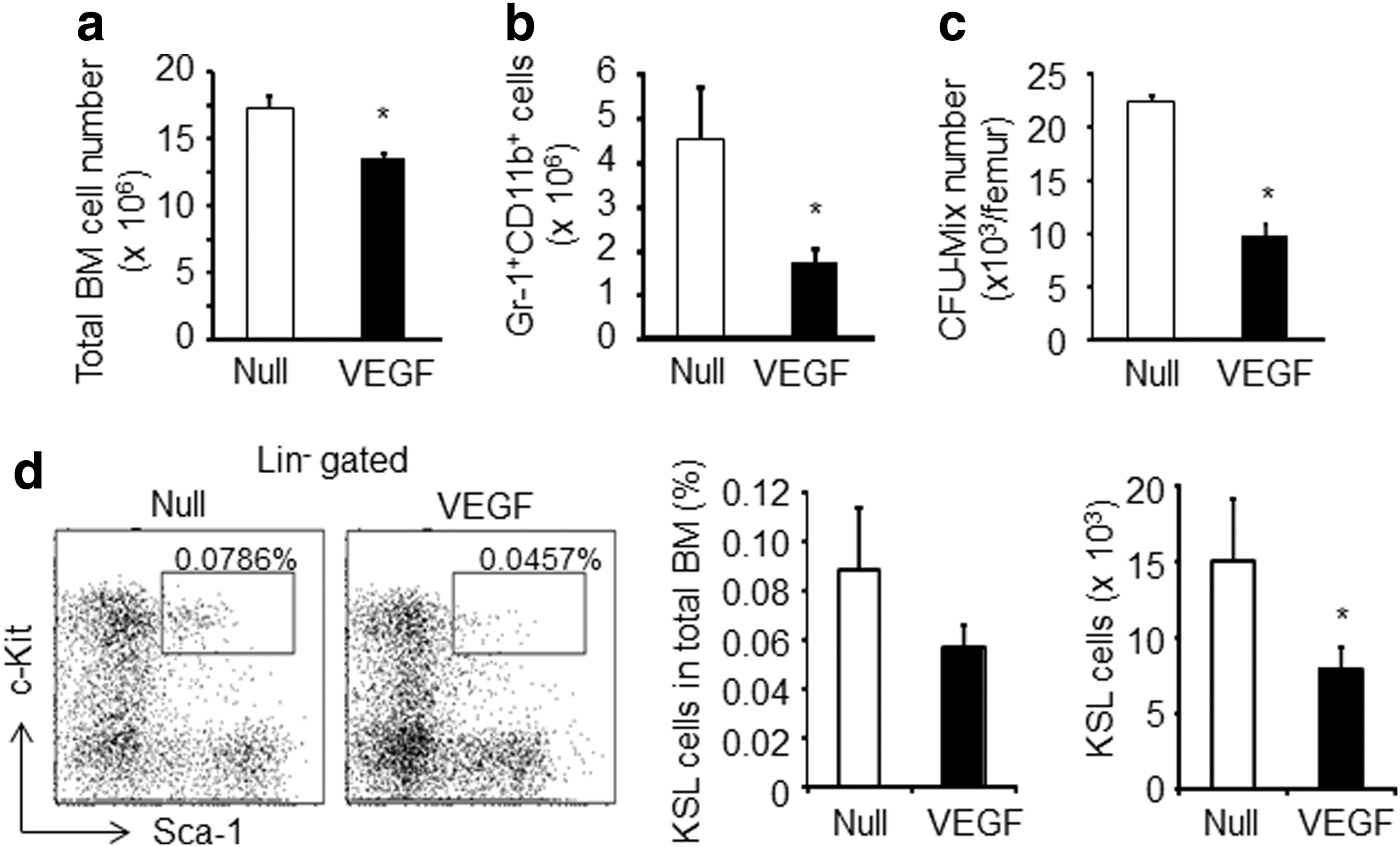
Plasma elevation of VEGF leads to a decrease in the myeloid cells and HSPCs in the BM.
Unchanged level of CXCL12 after VEGF overexpression
To examine the mechanisms of BM cell mobilization by VEGF treatment, we analyzed the expression levels of genes associated with HSC maintenance in the BM [N-cadherin (cdh2), vascular cell adhesion molecule-1 (Vcam-1), angiopoietin-1 (Angpt1), and thrombopoietin (Thpo)]. The expression levels of these genes in BM nonhematopoietic cells were modestly downregulated after Ad-VEGF injection (Fig. 3a). We next measured the CXCL12 levels in Ad-VEGF-injected mice. Chemokine CXCL12 is an indispensable factor for the maintenance and retention of HSPCs in the BM [5,24]. Previous studies showed that the BM CXCL12 levels were reduced by the injection of mobilization-inducing factors, such as G-CSF and stem cell factor (SCF) [10,25]. We also found that the CXCL12 levels were markedly reduced in the BM, but not the plasma, of Ad-G-CSF-injected mice (Fig. 3b). However, there was almost no difference in the BM CXCL12 levels between Ad-VEGF-injected mice and Ad-Null-injected mice (Fig. 3b). Therefore, these data indicate that VEGF would alter the BM microenvironment, probably by a different mechanism from other mobilization factors.

Expression of HSPC maintenance factor genes after Ad-VEGF administration.
Reduction of MPCs in the BM after Ad-VEGF injection
Recent studies have demonstrated that MPCs play a pivotal role in HSPC maintenance in the BM [4,6 –8]. Therefore, we examined the disposition of MPCs in the BM after Ad-VEGF administration. Flow cytometric analysis of the enzymatically dissociated BM cells revealed that Ad-VEGF overexpression led to a significant reduction of CD45-Ter119-CD31-Alcam−Sca-1+ cells, which are reported to be MPCs [21,26] (Fig. 4a). In addition, the percentage of other MPC populations, such as CD45−Ter119−PDGFRα+Sca-1+ cells [27] and CD45−Lineage-CD31−CD51+Sca-1+ cells [28], in the BM of Ad-VEGF-injected mice was also lower than those of Ad-Null-injected mice (Fig. 4b, c). These data clearly showed the decreased number of phenotypically identified MPCs in the BM after injection of Ad-VEGF.

The number of mesenchymal progenitor cells (MPCs) in the BM is decreased following Ad-VEGF injection.
Next, to investigate whether functional MPCs in the BM were reduced in Ad-VEGF-injected mice, we performed a CFU-F assay and homing assay. Consistent with the above data, we observed decreased CFU-F numbers in the BM in Ad-VEGF-injected mice (Fig. 4d). For homing studies, Ad-Null- or Ad-VEGF-injected mice were used as the recipient mice. Donor BM cells derived from GFP transgenic mice were intravenously injected into nonirradiated recipient mice, and the frequency of GFP-expressing cells in the recipient BM was then estimated by flow cytometry. The results showed that the homing activity of GFP-expressing cells was partially inhibited in Ad-VEGF-treated recipient mice (Fig. 4e). Thus, the decreased homing efficiency of donor HSPCs in Ad-VEGF-injected mice suggests the decreased number of niche cells in the BM. Taken together, our findings indicate that overexpression of VEGF in mice leads to a reduction of phenotypic and functional MPCs in the BM.
VEGF stimulates the migration of MPCs
We next examined the mechanisms of the reduction of MPCs in the BM after VEGF overexpression. In vitro-expanded primary mouse BM stromal cells (mBMSCs), including MPCs, showed slight expression of Flt-1 (VEGFR1), but not Flk-1 (VEGFR2), on the cellular surface (Fig. 5a). We speculated that MPCs might egress from the BM in response to the plasma level of VEGF, because there was almost no change in the BM VEGF levels in Ad-VEGF-injected mice (Fig. 1b). We performed an in vitro migration assay and found a dose-dependent chemoattractive effect of VEGF on mBMSCs (Fig. 5b), suggesting the possibility that a decreased number of BM MPCs in Ad-VEGF-injected mice would result from the mobilization of MPCs to the peripheral tissue in response to an elevation of plasma VEGF.

VEGF enhances the migration capacity of MPCs. BM-derived stromal cells were collected and propagated in a tissue culture dish.
Discussion
Recent studies have clearly reported that the HSPC numbers in the BM are significantly decreased by conditional deletion of MPCs, including nestin-expressing stromal cells [4] and CXCL12-abundant reticular cells [5]. It is of note that deletion of MPCs led to the increased number of HSPCs in the spleen, demonstrating the mobilization of HSPCs from BM to peripheral tissues [4]. Therefore, maintenance and retention of HSPCs in the BM would considerably be dependent on the number of MPCs [4,5]. In the present study, we examined the effect of VEGF on the disposition of BM HSPCs and MPCs in mice. Our main finding was that VEGF overexpression in mice resulted in a reduction of not only HSPCs but also MPCs in the BM. We also found that VEGF could promote the migration of mBMSCs in vitro. The data described here suggest that, as in the case of HSPCs, MPCs would also be mobilized to the peripheral tissues in response to an elevation of plasma VEGF levels, and a reduced number of BM MPCs by VEGF would lead to HSPC egress from the BM, because MPCs would function as niche cells in the BM.
It is well known that BM CXCL12 levels are downregulated following G-CSF administration and thereby induce an egress of HSPCs [25,29]. Christopher et al. previously showed the reduced BM CXCL12 levels after administration of other mobilization factors, such as SCF and Flt3-ligand [10]. In addition to their mobilization-inducing effects, these factors also impact the number of stem and progenitor cells in the BM. For instance, it has been reported that the number of HSPCs and MPCs in the BM was significantly increased after G-CSF administration [30,31]. Unlike in the case of G-CSF and other mobilization factors, however, VEGF had almost no effect on BM CXCL12 levels (Fig. 3b). Furthermore, systemic VEGF expression resulted in a significant reduction in the number of HSPCs (KSL cells and CFU-Mix) in the BM (Fig. 2). The number of MPCs in the BM was also reduced in Ad-VEGF-injected mice (Fig. 4). Therefore, these data strongly indicate that VEGF would induce HSPC mobilization by altering the BM environment through different mechanisms from G-CSF. Notably, a recent study showed that HSPCs could be mobilized from the BM into the circulation by administration of a prostagrandin E2 (PGE2) inhibitor, and this effect was independent of CXCL12-CXCR4 signaling [32]. A nucleotide sugar, uridine diphosphate (UDP)-glucose, has also been shown to mobilize subsets of HSPCs functionally distinct from those mobilized by G-CSF, suggesting that UDP-glucose-induced HSPC mobilization would be mediated, at least in part, by different mechanisms from G-CSF [33]. Thus, it would be of interest to examine whether VEGF could influence the levels of BM PGE2 and/or plasma UDP glucose.
The expression levels of HSC maintenance genes (Cdh2, Vcam-1, Angpt1, and Thpo) in BM nonhematopoietic cells were decreased in Ad-VEGF-injected mice (Fig. 3a). This would be due to the reduction in the number of MPCs in the BM after Ad-VEGF injection (Fig. 4). However, we have no idea why BM CXCL12 levels were not changed in Ad-VEGF-injected mice, because MPCs abundantly produce CXCL12 [7,8]. A detailed investigation would be required to clarify the regulation of CXCL12 expression in niche cells, including MPCs, endosteal osteoblasts, and endothelial cells.
We observed enhanced in vitro migration activities of mBMSCs by VEGF, suggesting the possibility that MPCs in the BM would be mobilized to the peripheral tissue in response to the plasma VEGF concentration. However, at present, we did not detect the CFU-F in the blood in Ad-VEGF-injected mice (data not shown). MPCs are known to be rare cells even in the BM, representing ∼1 in 10,000–100,000 total nucleated cells [34], and it is therefore possible that the frequency of MPCs in the blood was too low to detect under our experimental conditions. Alternatively, it is also possible that VEGF overexpression in mice might lead to the homing of MPCs to organs, such as the liver, because transgene expression in the liver was extremely high following systemic Ad vector injection [35]. Therefore, it might be necessary to investigate whether or not the frequency and the number of MPCs are changed in tissues or organs other than the peripheral blood.
Recently, Liu et al. showed that MPCs could be mobilized to the peripheral tissue when rats were exposed to hypoxic conditions, and this hypoxia-induced MPC mobilization was caused by the elevation of plasma CXCL12 levels and BM VEGF levels [36]. Under our conditions, however, plasma CXCL12 levels and the BM VEGF levels in Ad-VEGF-injected mice were almost equal to those in Ad-Null-injected mice (Figs. 1b and 3b), suggesting that the mechanisms of decreased number of BM MPCs in Ad-VEGF-injected mice would be different from those of hypoxia-induced MPC mobilization.
Consistent with previous reports [15], we confirmed the HSPC mobilization from BM into the circulation by VEGF overexpression using an Ad vector system (Fig. 1). On the other hand, a previous report showed that administration of a recombinant VEGF protein into mice failed to induce the HSPC mobilization [37]. In our Ad vector systems, plasma VEGF levels were maintained at 400–600 ng/mL on day 3–5 after single intravenous injection. Although we do not know the VEGF levels in the plasma under their experimental protocols, plasma VEGF levels might not be sufficient for HSPC egress from the BM, because exogenous VEGF levels in the plasma were rapidly decreased after administration of a recombinant VEGF protein [23]. Therefore, this difference would be partly due to the difference in the plasma VEGF levels, and we concluded that an Ad vector system would be an appropriate one to estimate the in vivo physiological action of VEGF.
In summary, our results showed that plasma VEGF levels could regulate the distribution of BM HSPCs and MPCs, probably by a mechanism distinct from that of other mobilization factors, and we suggest that a reduction in the number of MPCs in the BM would be one of the mechanisms involved in VEGF-induced HSPC mobilization. Although further investigation of the BM environment will be needed to uncover the VEGF-mediated HSPC mobilization, our findings obtained in this study provide a novel insight into the mechanisms of HSPC mobilization and would be helpful in the development of new clinical mobilizing agents.
Footnotes
Acknowledgments
This work was supported by grants from the Ministry of Health, Labour, and Welfare of Japan, and by the Sasakawa Scientific Research Grant from the Japan Science Society.
Author Disclosure Statement
The authors have no financial conflict of interests.