
Abstract
The CXCR4/SDF1 axis participates in various cellular processes, including cell migration, which is essential for skeletal muscle repair. Although increasing evidence has confirmed the role of CXCR4/SDF1 in embryonic muscle development, the function of this pathway during adult myogenesis remains to be fully elucidated. In addition, a role for CXCR4 signaling in muscle maintenance and repair has only recently emerged. Here, we have demonstrated that CXCR4 and stromal cell-derived factor-1 (SDF1) are up-regulated in injured muscle, suggesting their involvement in the repair process. In addition, we found that notexin-damaged muscles showed delayed muscle regeneration on treatment with CXCR4 agonist (AMD3100). Accordingly, small-interfering RNA-mediated silencing of SDF1 or CXCR4 in injured muscles impaired muscle regeneration, whereas the addition of SDF1 ligand accelerated repair. Furthermore, we identified that CXCR4/SDF1-regulated muscle repair was dependent on matrix metalloproteinase-10 (MMP-10) activity. Thus, our findings support a model in which MMP-10 activity modulates CXCR4/SDF1 signaling, which is essential for efficient skeletal muscle regeneration.
Introduction
S
Chemokines constitute a family of structurally related low-molecular-weight cytokines that play fundamental roles in cellular trafficking and migration [6]. Stromal cell-derived factor-1 (SDF1) belongs to the CXC chemokine subfamily and was identified as a pre-B cell stimulatory factor [7]. SDF1 is known to be a chemoattractant for stem/progenitor cells [8,9] and mainly functions as a ligand for the G protein-coupled receptor, CXCR4 [10]. Notably, SDF1-mediated CXCR4 activation is crucial in the regulation of various biological processes involving cell motility, chemotactic response, cell adhesion, gene transcription, cell proliferation, and survival [11 –15]. In addition, SDF1 seems to play a beneficial role in tissue repair after myocardial infarction [16 –18], vascular disease [19,20], and fracture healing [21]; however, it may also contribute to cancer progression [22,23]. The discovery of SDF1 and CXCR4 expression in skeletal muscle has led to the study of their potential roles in embryonic myogenesis [24]. In fact, muscle progenitor cells, which express CXCR4, have been shown to migrate toward SDF1 during the generation of new myofibers in limb muscles [25]. Accordingly, null mice for CXCR4 have a reduced number of satellite cells, leading to muscular deficiencies in the embryo [26]. However, the function of the CXCR4/SDF1 axis during adult myogenesis remains to be fully investigated [27 –30], and its role in muscle repair has only recently begun to be characterized [31].
Myogenic cells produce and secrete MMPs, a family of zinc endopeptidases that degrade components of the extracellular matrix (ECM) and participate in diverse cellular processes [32 –34]. In addition, the CXCR4/SDF1 signaling axis and MMPs have been shown to participate in some of the same cellular processes, including migration of muscle-derived cells during muscle repair [5,35,36]. This might suggest that these two systems functionally interact. In this regard, MMP-10 (stromelysin-2), which belongs to the stromelysin family of MMPs, might be of primary interest. Although MMP-10 has been shown to be involved in various pathologies, including cancer and vascular disease [37 –39], we recently described its pivotal role in muscle maintenance and repair. In fact, MMP-10 deficiency delays muscle repair after injury and deteriorates the dystrophic phenotype of mdx mice. Furthermore, the administration of recombinant human MMP-10 (rhMMP-10) improves muscular regeneration after damage [40].
In the present study, we demonstrated that the CXCR4/SDF1 axis is required for efficient muscle repair. Accordingly, we observed that the CXCR4 agonist, AMD3100, delayed skeletal muscle regeneration. In addition, small-interfering RNA (siRNA)-mediated silencing of SDF1 or CXCR4 in injured muscle impaired muscle regeneration, whereas the addition of SDF1 ligand accelerated the repair process. In addition, our in vitro data revealed that the CXCR4/SDF1 axis functions mechanistically at the level of satellite cell-derived myoblasts, which are directly responsible for muscle repair. Furthermore, we identified that MMP-10 might mediate the effect of CXCR4/SDF1 on injured muscle, supporting the existence of a functional interaction between these systems. Collectively, our findings reveal a new area of investigation, which could contribute to the development of novel therapies for muscle injury and/or dystrophic diseases.
Materials and Methods
Mice
Wild-type (WT; C57BL/6J) mice were purchased from Harlan Iberica. MMP-10 knockout (KO) mice were provided by Dr. W.C. Parks from the University of Washington. Genotypes for MMP-KO mice were determined through PCR analysis of mouse ear DNA using specific primers for MMP-10 (sense 1, 5′-TGTGTAGTGCCTACACTAAGCCA-3′; sense 2, 5′-TGCCTCGTCCTGCAGTTCATTC-3′; antisense, 5′-TAAGGGTGTGAGTCTTCATGGAT-3′). All animals were used at 8 weeks of age.
Model of muscle injury and in vivo treatments
Muscle damage was induced by an intramuscular injection of 10 μL (10 μg/mL) of Notechis scutatus scutatus notexin (Latoxan) into the left tibialis anterior (TA) muscle, leaving the right TA as a control [phosphate-buffered saline (PBS)]. In some experimental conditions, notexin was delivered to both TA muscles. After 24-h notexin treatments, we intramuscularly injected SDF1 (100 ng/mL; R&D), AMD3100 (10 μM; Sigma), or rhMMP-10 (100 ng/mL; R&D) into TA muscles as indicated, leaving contra-lateral muscles as controls.
A pair of specific siRNA sequences was pooled for each target, including SDF1 (siSDF1), CXCR4 (siCXCR4), and MMP-10 (siMMP-10) (Stealth siRNA; Invitrogen). These siRNAs were prepared in atelocollagen (Koken) according to the manufacturer's instructions and injected intramuscularly into notexin-treated left TA, whereas right injured TA was injected with empty atelocollagen as a control. Control siRNA (siControl) was also prepared and delivered under the same conditions (Invitrogen). The following specific siRNA sequences were used: SDF1, 5′-AGUGUGCAUUGACCCGAAAtt-3′ and 5′-UUUCGGGUCAAUGCACACUtg-3′; CXCR4, 5′-GGUACUUUGGGAAAUUUUUtt-3′ and 5′-AAAAAUUUCCCAAAGUACCag-3′; MMP-10, 5′-GACAGAUAACAGAUGAUUUtt-3′ and 5′-CGGAGACUUUUACCUUUUtt-3′.
Mice were sacrificed at 7 and 14 days after treatment by cervical dislocation and their TA muscles were removed.
Isolation and culturing of satellite cells
WT mice were killed by cervical dislocation, and the extensor digitorum longus (EDL) muscles were carefully removed. Myofibers were then isolated from EDL muscles as previously described (40). For suspension culture, myofibers were incubated in DMEM with 10% horse serum (PAA Laboratories), 0.5% chick embryo extract (ICN Flow), 4 mM
In vitro treatments
Satellite cell-derived myoblasts were grown to 30% confluency in six-well plates and transfected with siRNA. A second transfection was performed 24 h later. The siRNA duplexes were diluted in OptiMEM (Invitrogen) at 20 pmol/well and incubated with Lipofectamine 2000 (Invitrogen) in OptiMEM, according to the manufacturer's instructions. For myogenic differentiation, cells were transferred to eight-well chamber slides (Lab-Tek) immediately after the second transfection and maintained in differentiation medium (DMEM plus 2% horse serum).
Mouse SDF1 cDNA was cloned into pcDNA3.1/V5 plasmid (Invitrogen; K4800-01) and transfected into the COS-1 cell line (ATCC; CLR-1650) using Fugene-6 (Roche). Transfected cells were selected with geneticin (50 ng/mL; Invitrogen) and plated in six-well plates (100,000 cells/well) a day before isolating myofibers. Some COS-1 cells were transfected with the empty pcDNA plasmid, serving as control cells. The secretion of SDF1 into the media (10 ng/mL) was confirmed using an SDF1 ELISA Kit (Roche). Single EDL myofibers, along with their associated satellite cells, were cocultured with SDF1-transfected COS-1 cells in transwell plates (0.4 μm pore size; Corning) for 72 h. Some of them underwent a 24-h pretreatment with AMD3100 (10 μM). In addition, SDF1 (R&D) was sometimes added to the media of plated satellite cells or suspended single myofibers (10 ng/mL).
Immunostaining
Skeletal muscles were frozen in liquid nitrogen-cooled isopentane and serial 9-μm cryosections were collected at 200-μm intervals through the entire muscle. Sections were blocked with 20% goat serum and immunostained with mouse anti-embryonic myosin heavy chain (eMyHC) (1:100; DSHB), anti-Pax7 (1:100; DSHB), and anti-Ki67 (1:100; NeoMarkers). For all nuclear staining, 4,6-diamidino-2-phenylindole (DAPI) was used.
Cells and myofibers were fixed with 4% paraformaldehyde/PBS and permeabilized with 0.5% (v/v) Triton X-100/PBS. After blocking with 20% normal goat serum, cells and myofibers were incubated with the following primary antibodies (overnight, 4°C): rat anti-BrdU (1:300, clone BU1/75; Abcam), mouse anti-myosin (1:100, clone MF20; DSHB), anti-Pax7 (1:100; DSHB), and rabbit anti-MyoD (1:50; Santa Cruz). Primary antibodies were detected and visualized with fluorochrome-conjugated secondary antibodies (1:200; Amersham) before mounting in DakoCytomation Faramount fluorescent mounting medium containing DAPI.
Image capture and quantification
Immunostained cells, myofibers, and tissue sections were viewed on a Zeiss Axiophot epifluorescence microscope. Digital images were acquired with an AxioCamMR3 camera (Zeiss). In addition, AxioVision software and ImageJ were used to analyze tissue sections or cells (ie, whole muscle transversal sections or entire areas occupied by cells). Positive staining for eMyHC was calculated, in each of the treated and control muscles, as the percentage of stained area divided by the total area of muscle. The percentage of positive area for eMyHC in treated TA was related to that found in their respective controls and expressed as a fold change in the % of eMyHC expression. The number of proliferating and nonproliferating satellite cells was calculated by counting the total number of Pax7+/Ki67+ and Pax7+/Ki67− satellite cells, which were then related to the total muscle area (mm2). Fusion index was calculated by counting the number of nuclei within myosin+ myotubes and expressed as a percentage. In addition, the mean number of nuclei composing each myotube was calculated by counting the DAPI+ nuclei in at least 50 myotubes in each experimental condition. The amount of proliferating satellite cells was determined by counting the number of BrdU+cells in relation to the number of DAPI+ cells. To establish the percentage of satellite cells with each phenotype, we analyzed at least 15–20 individual myofibers (along with their associated satellite cells). In at least four independent experiments, we counted Pax7−/MyoD+, Pax7+/MyoD+, and Pax7+/MyoD− satellite cells, which were subsequently related to the total number of stained cells within the same fiber.
Immunoblotting
TA muscles and cells were homogenized in ice-cold lysis buffer (1% Triton X-100), which was supplemented with a mixture of phosphatase inhibitors (50 mM sodium pyrophosphate, 0.1 M NaF, and 10 mM sodium orthovanadate). Protein concentrations were determined using the BCA Protein Assay Kit (Pierce).
Proteins in muscle homogenates (40 μg), cell lysates (15 μg), and cell supernatants (20 μg) were separated via SDS-PAGE (10% gels) electrophoresis and subsequently transferred to nitrocellulose membranes (Bio-Rad Laboratories). The membranes were then blocked for 1 h at room temperature in Tris-buffered saline (10 mM Tris-HCl, 150 mM NaCl, and pH 8) containing 0.05% Tween 20 (TBS-T) and 10% nonfat dry milk. The blocked membranes were then probed with the following primary antibodies in blocking buffer (overnight, 4°C): mouse anti-Pax7 (DSHB), anti-MyoD (DSHB), anti-myosin (DSHB), anti-PCNA, (Santa Cruz Biotechnology), rabbit polyclonal anti-SDF1 (Cell Signaling), and anti-CXCR4 (Abcam). Mouse antibodies against β-actin (Sigma) or β-tubulin (Promega) were used as loading controls. The antigen–antibody complexes were detected using peroxidase-labeled goat anti-rabbit or anti-mouse antibodies (Amersham) in TBS-T containing 2.5% nonfat dry milk for 1 h at room temperature. The results were visualized using the enhanced chemiluminescence ECL Plus detection system (Amersham), and the bands were analyzed via densitometric analysis with the Gel Doc-Quantity One 4.5.0 (Bio-Rad).
Quantitative real-time PCR
Total RNA was isolated from cells and purified using UltraSpec reagent (Biotecx Laboratories). The total RNA was then reverse transcribed using Superscript II RNase H reverse transcriptase (Invitrogen), and cDNA was amplified using the TaqManH Universal PCR Master Mix (Applied Biosystems). Transcript levels were quantified by real-time PCR (7300 Real-Time PCR System; Applied Biosystems) under the following conditions: 95°C for 3 min followed by 40 cycles consisting of 60 s at 90°C, 60 s at 60°C, and 60 s at 72°C. Calibration curves (1, 2, 5, and 10 ng of target cDNA) were included to specifically analyze CXCR4 and SDF1 expression. All measurements were related to the curves and normalized to GAPDH (Applied Biosystems). Specific primers and probes were designed using the Primer3 program (version 0.4.0) for CXCR4 (forward, 5′-ACGGCTGTAGAGCGAGTGTT-3′; reverse, 5′-AGGGTTCCTTGTTGGAGTCA-3′; probe, 5′-CATGGAACCGATCAGTGTGA-3′) and SDF1 (forward, 5′-CCAAACTGTGCCCTTCAGAT-3′; reverse, 5′-AAGTCCTTTGGGCTGTTGTG-3′; probe, 5′-TGTGCATTGACCCGAAATTA-3′).
Statistical analysis
All statistical analyses were performed using SPSS 15.0 (SPSS). The Kolmogorov–Smirnov test was used to verify the normality of the samples. If the variables passed the test, then we applied the parametric Student's t-test and one-way ANOVA followed by least significant difference (LSD) and honestly significant difference tests. However, when variables did not follow the normality criteria according to the Kolmogorov–Smirnov test, we applied the nonparametric Kruskal–Wallis and Mann–Whitney U tests. Data were expressed as means±standard error of the mean. Statistical significance was defined as P<0.05.
Results
The CXCR4/SDF1 axis is required for muscle regeneration
First, we confirmed that our experimental model of injury induction led to increases in CXCR4 and SDF1. We intramuscularly injected notexin into TA muscles of WT mice and assessed protein levels throughout the course of the degeneration–regeneration process via immunoblot (Supplementary Fig. S1A–C; Supplementary Data are available online at
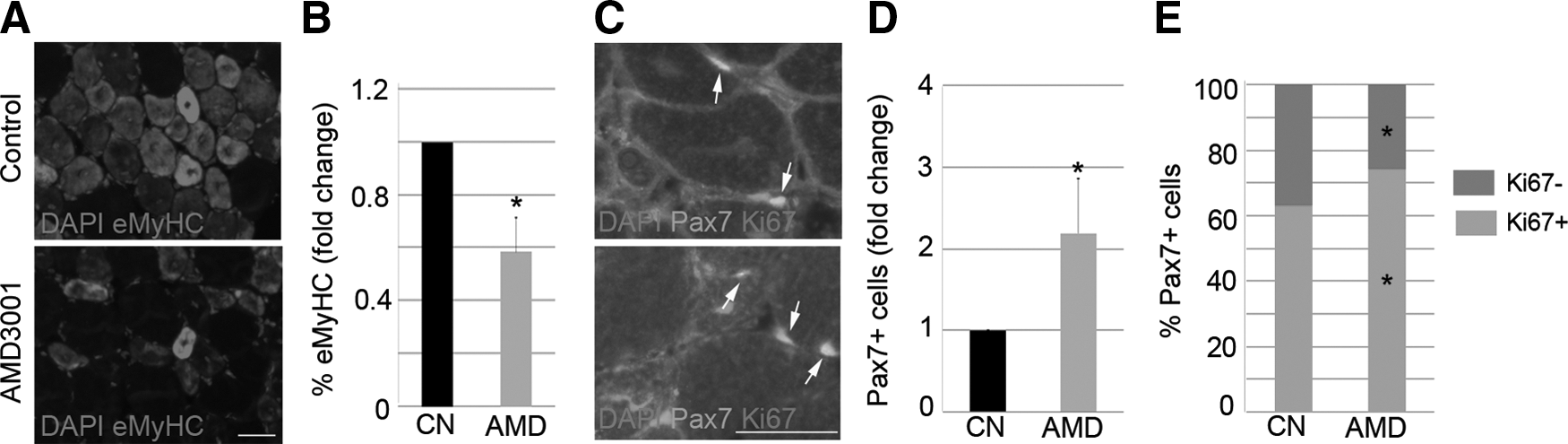
Inhibition of the CXCR4/SDF1 axis delays muscle regeneration. TA muscles of WT mice were injected with notexin. One day later, the right TA was treated with AMD3001 (AMD), and the left TA received PBS (Control, CN). The animals were sacrificed 7 days later, and the muscles were removed. Representative images of eMyHC
To assess whether delayed muscular repair is permanent, we analyzed injured TA 14 days after treatment with AMD. Despite finding a very few eMyHC+ regenerating fibers in injured muscles treated with AMD but not in the TA from controls (Supplementary Fig. S2A), the architecture of muscles was roughly similar, which, in turn, demonstrates that AMD temporally delays muscular regeneration.
Inhibition of CXCR4/SDF1 components impairs muscle regeneration
In order to verify the importance of the CXCR4/SDF1 axis in muscle regeneration and to better understand the individual roles of CXCR4 and SDF1 in the repair process, we depleted each of these components in damaged muscles. For this, both TA muscles of WT mice were treated with notexin. After 24 h, pooled siRNA duplexes (10 μM) against either CXCR4 (siCXCR4) or SDF1 (siSDF1) were intramuscularly delivered to the right damaged TA, while the left TA received control siRNA (siControl). Animals were sacrificed 7 days later, and the loss of CXCR4 and SDF1 proteins was confirmed by western blotting (Fig. 2A, E). Muscle regeneration was also analyzed.
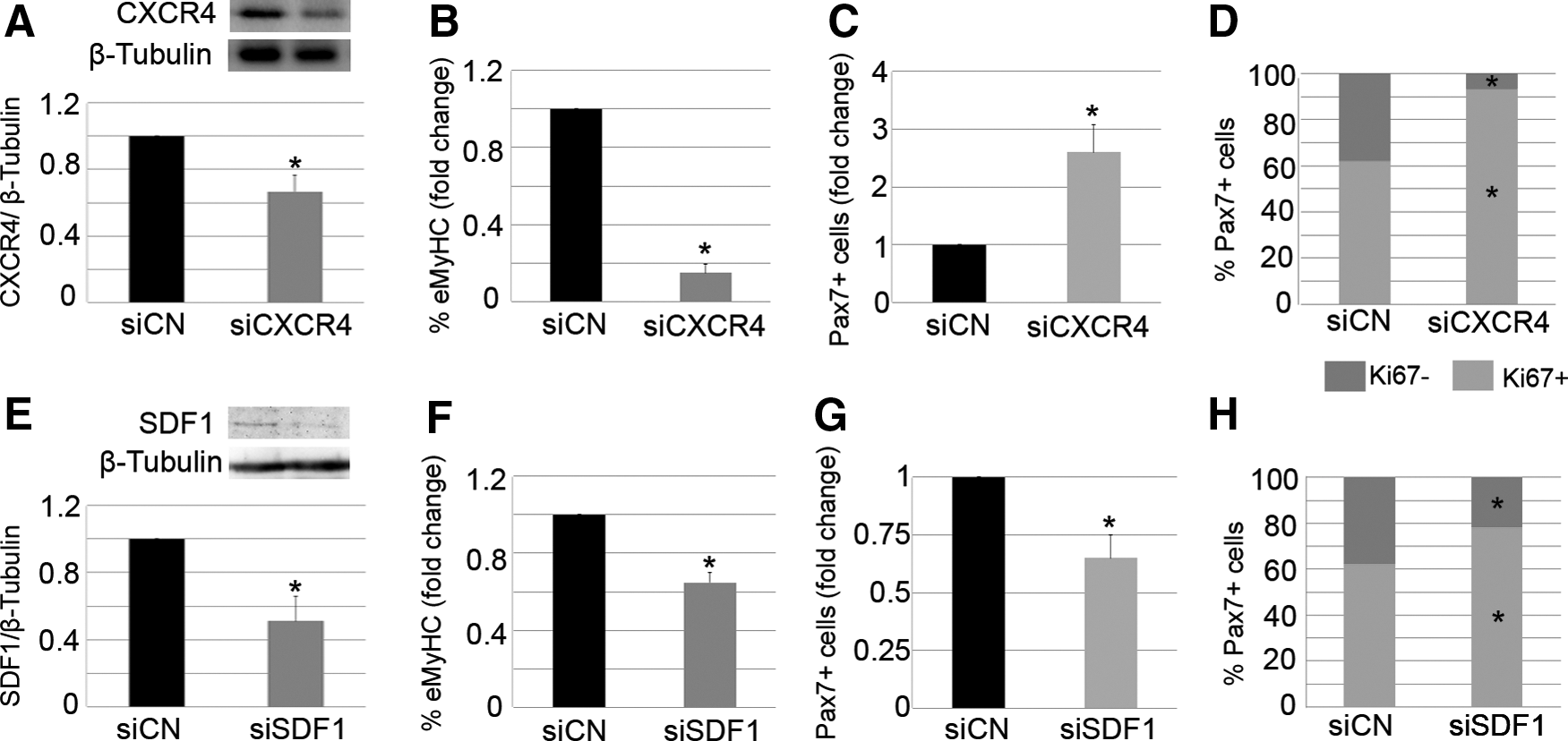
Local modulation of CXCR4 or SDF1 impairs muscle regeneration. Right injured TA muscles of WT mice were treated with specific siRNA duplexes against SDF1 (siSDF1) or CXCR4 (siCXCR4), while contra-lateral damaged muscles received control siRNA (siCN). Seven days later, the animals were sacrificed. Western blot analysis revealed down-regulation of target genes
Similar to what we found in injured muscles treated with AMD, the administration of siCXCR4 into notexin-injured muscles led to a significant reduction of the percentage of new regenerating myofibers, as determined by the presence of eMyHC expression (Fig. 2B). In addition, the total number of Pax7+ satellite cells in siCXCR4-treated muscles was higher than that found in controls (Fig. 2C). An increased percentage of proliferating Pax7+ satellite cells was observed (Fig. 2D), suggesting a delay in muscle repair.
Interestingly, despite the fact that siSDF1-treated injured muscles displayed a lower percentage of eMyHC+ regenerating fibers compared with controls (Fig. 2F), and that the majority of Pax7+ satellite cells expressed Ki67 on SDF1 depletion (Fig. 2H), the number of total Pax7+ satellite cells was significantly decreased compared with the controls (Fig. 2G). These results suggest that SDF1 silencing impairs the differentiation of satellite cell progeny and is required to properly maintain the pool of satellite cells after muscle injury.
To assess whether modulation of CXCR4/SDF1 axis alters muscle repair kinetics, we analyzed TA 14 days after injury. Injured muscles treated with siCXCR4 or siSDF1 had a few eMyHC+ fibers as well as injured muscles treated with siControl (Supplementary Fig. S2B, C). These results suggest that transient alteration of CXCR4 and SDF1 expression in injured muscle alter muscular regeneration kinetics but not permanently.
Expression of CXCR4 and SDF1 is required for in vitro myogenesis
To characterize the role of the CXCR4/SDF1 axis in satellite cells, we cultured satellite cell-derived myoblasts and differentiated them through mitogen withdrawal. Throughout the course of myogenic differentiation, from myoblasts to myotubes with the ability to spontaneously contract, we observed increased expression and secretion of SDF1 by quantitative real-time PCR and western blot (Supplementary Fig. S3A–C). In contrast, we found that CXCR4 expression was down-regulated as myoblasts fused into myotubes (Supplementary Fig. S3D, E). These primary satellite cells were subsequently transfected with siCXCR4, siSDF1, or siControl. After 6 days in culture, the depletion of CXCR4 and SDF1 was confirmed by western blot (Fig. 3A, D). Notably, compared with controls, loss of CXCR4 or SDF1 led to a decreased number of BrdU+ satellite cells (Fig. 3B, E; arrows) and reduced ability of cells to fuse into myotubes, as determined by the fusion index (Fig. 3C, F). In addition, the mean number of nuclei in each myotube showed that CXCR4 and SDF1 silenced myoblasts had a defect in cell fusion (Fig. 3C, F), suggesting that these proteins are required for properly growth of myotubes. These findings were verified by immunoblotting for PCNA, MyoD, Pax7, and MyHC (Supplementary Fig. S4A–H). Collectively, these data confirm that both CXCR4 and SDF1 are required for efficient in vitro myogenesis.

Reduction of CXCR4 and SDF1 expression in myoblasts affects myogenesis. Satellite cell-derived myoblasts were transfected with siCXCR4 or siControl and cultivated for 6 days. Loss of CXCR4
Administration of SDF1 improves muscle regeneration
Based on our findings, we hypothesized that the CXCR4/SDF1 axis directly participates in the regenerative processes occurring in the muscle. In support of this theory, we injected notexin into both TA muscles of WT mice. One day later, we administered SDF1 (100 ng/mL) into the left muscle, while the right muscle served as the control. Strikingly, SDF1 treatment tripled the expression of eMyHC by day 7 (Fig. 4A), whereas the number of total Pax7+ satellite cells (Fig. 4B, C; arrows) was significantly decreased in comparison to controls. Accordingly, the administration of SDF1 led to a lower percentage of proliferating Pax7+satellite cells in damaged muscle (Fig. 4D). Overall, these results suggest that SDF1 directly accelerates muscle repair after injury.

SDF1 accelerates muscle repair after injury. Right notexin-injured TA muscles of WT mice were treated with SDF1, while contra-lateral damaged muscles received PBS (control). Animals were sacrificed 7 days later. We quantitated the percentage of eMyHC+ cells
Exogenous SDF1 accelerates in vitro myogenic kinetics
To understand the effects of the SDF1 gain of function during muscle repair, we examined the role of SDF1 during the earlier steps of myogenic progression. To this end, single myofibers, along with their attached satellite cells, were cocultured with SDF1-secreting COS-1 cells (SDF1: 10 ng/mL). In addition, other fibers were cocultured with control COS-1 cells. After 72 h in suspension, SDF1 increased the percentage of Pax7+/MyoD+ satellite cells (Fig. 5A, C; arrows), while those with the Pax7−/MyoD+ phenotype were reduced (Fig. 5A, C; arrowheads). These findings support the notion that SDF1 participates in satellite cell proliferation rather than differentiation. In addition, similar data were obtained when exogenous SDF1 (10 ng/mL) was added to single myofiber-associated satellite cells (Fig. 5B). Notably, we found that the effect of SDF1 on myogenic progression was completely abolished on pretreatment with AMD (Fig. 5D). These findings demonstrate that the earlier steps of myogenic progression are dependent on the CXCR4–SDF1 interaction.

Exogenous SDF1 accelerates the myogenic kinetics of myoblasts.
In order to investigate the involvement of SDF1 in the later steps of myogenic progression, such as cell fusion, we added exogenous SDF1 (10 ng/mL) to primary satellite cell-derived myoblasts. Compared with control cells, treatment with SDF1 for 6 days led to a doubling in the number of proliferating myoblasts (Fig. 5E) and augmented fusion into myotubes (Fig. 5F). In addition, the increased mean number of nuclei in myotubes in SDF1- treated cell cultures showed that SDF1 improved growth of myotubes (Fig. 5G). These results correlated with higher protein levels of PCNA, MyoD, Pax7, and MyHC in SDF1-treated myoblasts compared with controls (Supplementary Fig. S5A–D). However, it remains possible that the effects of SDF1 on the later steps of myogenic differentiation are simply a consequence of increased proliferation during early myogenesis.
The CXCR4/SDF1 axis requires MMP-10 activity to induce muscle regeneration
It was previously reported that various MMPs, such as MMP-2 or MMP-9, could interact with the CXCR4/SDF1 axis during the regulation of specific cellular processes [30,42 –45]. We recently described that the modulation of MMP-10 protein levels in injured muscle affects muscular regeneration [40]. In fact, the delivery of siMMP-10 into injured muscles delayed muscle regeneration; whereas rhMMP-10 accelerated the repair process. In addition, MMP-10 KO mice have delayed muscular regeneration compared with WT animals [40].
Since we had previously described a role for MMP-10 in muscle regeneration, we next sought to analyze whether MMP-10 modulation could affect the ability of the CXCR4/SDF1 axis to regulate repair. First, we assessed CXCR4 and SDF1 expression in injured muscles, which were treated with siMMP-10 or rhMMP-10 to delay or accelerate repair, respectively [40]. Western blot analysis revealed that siMMP-10 led to a significant reduction in CXCR4 and SDF1 levels (Supplementary Fig. S6A–C), while these proteins were significantly higher in rhMMP-10-treated muscles (Supplementary Fig. S6D–F). In addition, we assessed the expression of CXCR4 and SDF1 in muscles from MMP-10 KO mice. Indeed, the protein levels of both the receptor and ligand were significantly lower in MMP-10 KO TA compared with WT muscles (Supplementary Fig. S6G–I). These findings suggest that MMP-10 could mediate the effect of the CXCR4/SDF1 axis during muscle repair.
To corroborate our hypothesis, we induced injury in both TA muscles of MMP-10 KO mice and intramuscularly injected the right muscles with AMD or SDF1 (after 1 day of damage). The left TA muscles received PBS and served as controls. Seven days later, eMyHC staining revealed that SDF1 administration did not increase the percentage of newly regenerating myofibers in injured KO muscles (Fig. 6A), which was contrary to results observed with WT mice. Similarly, AMD treatment did not alter the expression of new eMyHC+ regenerating myofibers in damaged muscles from MMP-10 KO animals (Fig. 6B). Taken together, these findings suggest that the CXCR4/SDF1 axis requires MMP-10 activity to induce muscle repair.
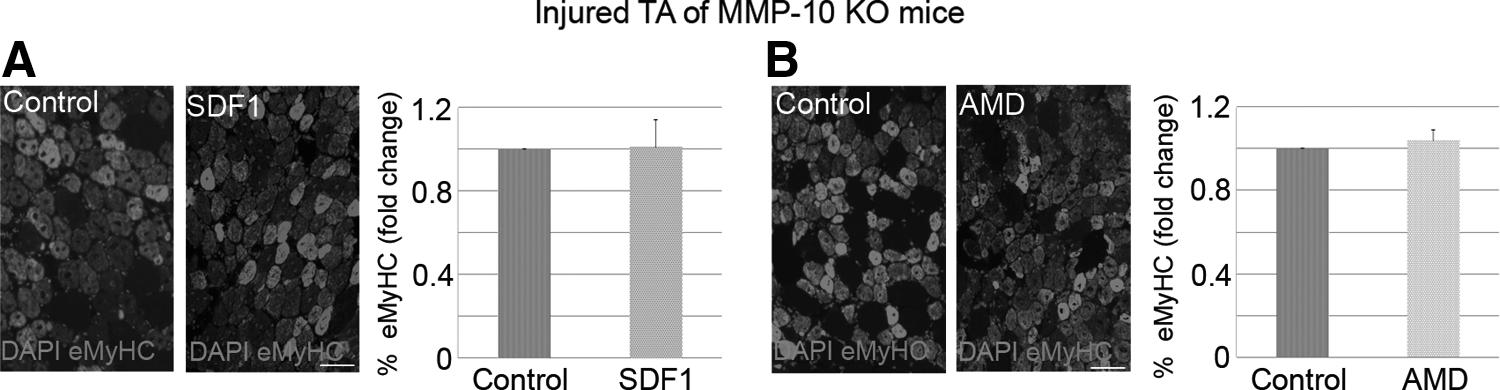
The CXCR4/SDF1 axis requires MMP-10 activity to induce muscle regeneration.
Discussion
In the present study, we have examined the role of the CXCR4/SDF1 axis in muscle regeneration after injury. As previously reported [31,46], our findings have confirmed that skeletal muscle responds to damage by enhancing the expression of CXCR4 and SDF1, suggesting their participation in the muscle repair process. We found that AMD3001, an antagonist of CXCR4 [41], delayed muscle regeneration, reducing the number of eMyHC+ myofibers and increasing the percentage of proliferating Pax7+ satellite cells. In addition, transient depletion of CXCR4 or SDF1 significantly delayed muscle regeneration, as shown by reduced expression of eMyHC. In contrast, exogenous SDF1 accelerated the repair process. Furthermore, it appears that SDF1 is required to properly maintain the satellite cell pool after injury, as the total number of Pax7+ satellite cells decreased in SDF1-depleted injured muscles. In line with this, it was reported that local elevation of SDF1 in injured organs promotes cell recruitment that supports tissue repair and regeneration [18,47,48], an effect that is abrogated by AMD3100 [49,50]. Thus, in our injury model, we speculate that increased SDF1 is required for the recruitment of Pax7+ satellite cells, which subsequently differentiate to restore/replace damaged fibers.
It has been demonstrated that SDF1 expression at sites of injury can also recruit immune cells or endothelial progenitor cells, which can participate in muscle repair [51]. In addition, SDF1 participates in the release of ECM components, which are involved in muscle regeneration [52]. These factors might explain why our in vivo data were not efficiently reproduced in our in vitro models, as it is sometimes difficult to specifically attribute specific processes to particular cell types. In this regard, we have not determined which cells secrete SDF1 or which CXCR4+ cells are recruited to injured sites. However, our in vitro data confirmed that both CXCR4 and SDF1 had a direct effect on myogenic cells in the absence of other confounding cell types, and these myogenic cells are known to be directly responsible for muscle repair [4]. In addition, our data have verified that CXCR4 expression decreased during myogenesis, while expression and secretion of SDF1 increased [31,32]. These findings suggest that contractile myotubes produce SDF1 in order to attract satellite cells and facilitate fusion, which could mirror the process in vivo. Furthermore, we found that CXCR4 or SDF1 depletion diminished the ability of satellite cell-derived myoblasts to incorporate BrdU, subsequently reducing myotube differentiation. Accordingly, exogenous SDF1 increased satellite cell proliferation and fusion into myotubes. Therefore, since cell fusion constitutes a later event in the repair process, we propose that the CXCR4/SDF1 axis is required to satellite cells account the repair response of muscle after an episode of injury.
Although SDF1 mainly functions through CXCR4, it can also interact with CXCR7 [46,53]. Both CXCR4–SDF1 and CXCR7–SDF1 interactions regulate myogenesis, but at different stages of differentiation. SDF1 participates in the early steps of myogenic progression through CXCR4, whereas the CXCR7/SDF1 axis regulates later steps [46]. In suspension myofibers (ie, earlier steps of myogenic progression), we found that SDF1 increased the percentage of Pax7+/MyoD+ satellite cells, while the percentage of those with the Pax7−/MyoD+ phenotype decreased. This suggests that SDF1 influences satellite cell proliferation rather than differentiation. In addition, we found that this effect was abrogated on pretreatment of myofiber-associated satellite cells with AMD3100, confirming that SDF1 acts through CXCR4. These findings support a model in which increased satellite cell fusion into myotubes during the later steps of myogenic differentiation is secondary to enhanced proliferation.
Migration from distant myofibers to sites of injury requires satellite cells to cross the basal lamina of their resident myofibers [3], followed by directed movement to damaged areas [5]. This idea is in agreement with the notion that myogenic precursors not only respond to chemokines, but also should express and secrete MMPs [54]. In this regard, increased expression of MMPs and cytokines in damaged skeletal muscle seems to be indicative of their roles in muscle repair or disease progression [55 –57]. However, the concomitant up-regulation of these factors could also suggest a functional interaction between them. In fact, several pieces of data support this idea: (1) MMPs and cytokines participate in the same cellular processes (eg, migration) [58 –60]; (2) MMPs can regulate chemokine activity through direct proteolysis of chemokines; (3) and MMPs contribute to chemokine release from ECM [61 –63]. In addition, some chemokines can affect MMP function by directly or indirectly [64,65] altering MMP activity. Thus, MMPs have the ability to modulate the locomotion machinery of cells through interaction with chemokines.
We recently demonstrated that MMP-10 has a pivotal role in skeletal muscle maintenance and repair [40]. In the present study, we have extended these previous findings by identifying that alterations in MMP-10 levels had parallel affects on the expression of CXCR4 and SDF1. Furthermore, we found that AMD3100 and SDF1 lost their effects on muscle regeneration (delayed or accelerated repair, respectively) in muscles from MMP-10 KO mice. This suggests that the CXCR4/SDF1 axis requires MMP-10 activity to induce efficient repair of skeletal muscle. Although the biochemical pathway triggered by MMP-10 during CXCR4/SDF1-induced muscle regeneration requires further investigation, it is clear that cross-talk exists between these two systems.
Taken together, our data suggest that the CXCR4/SDF1 axis is pivotal for efficient muscle regeneration after an episode of injury. In addition, it is possible that this system functions through MMP-10 activity where MMP-10 might regulate CXCR4/SDF1-mediated muscle repair via chemokine modulation. These findings suggest an important molecular mechanism that could be exploited to develop novel therapies for dystrophic diseases.
Footnotes
Acknowledgments
This work was supported by grants from the Spanish Ministry of Science and Technology (SAF2009-12039); Caja de Ahorros de Navarra (Programa Tú Eliges: Tú Decides); the Spanish Ministry of Health (PI08/1919, PS09/00143 and RD12/0019/0031); the Ministerio de Economia y Competitividades (PLE2009-0116); European Union's Seventh Framework Program for Research, Program (INELPY); and the Unión Temporal de Empresas-Centro de Investigación Médica Aplicada. The authors thank Virginia Izuriaga and María Jiménez for their technical assistance. They also thank Parks for providing MMP-10 KO mice. They are grateful to the Developmental Studies Hybridoma Bank for the antibodies used in this study.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.