
Abstract
A major challenge facing the development of effective cell therapies is the efficient differentiation of pluripotent stem cells (PSCs) into pure populations. Lowering oxygen tension to physiological levels can affect both the expansion and differentiation stages. However, to date, there are no studies investigating the knock-on effect of culturing PSCs under low oxygen conditions on subsequent lineage commitment at ambient oxygen levels. PSCs were passaged three times at 2% O2 before allowing cells to spontaneously differentiate as embryoid bodies (EBs) in high oxygen (20% O2) conditions. Maintenance of mouse PSCs in low oxygen was associated with a significant increase in the expression of early differentiation markers FGF5 and Eomes, while conversely we observed decreased expression of these genes in human PSCs. Low oxygen preconditioning primed mouse PSCs for their subsequent differentiation into mesodermal and endodermal lineages, as confirmed by increased gene expression of Eomes, Goosecoid, Brachyury, AFP, Sox17, FoxA2, and protein expression of Brachyury, Eomes, Sox17, FoxA2, relative to high oxygen cultures. The effects extended to the subsequent formation of more mature mesodermal lineages. We observed significant upregulation of cardiomyocyte marker Nkx2.5, and critically a decrease in the number of contaminant pluripotent cells after 12 days using a directed cardiomyocyte protocol. However, the impact of low oxygen preconditioning was to prime human cells for ectodermal lineage commitment during subsequent EB differentiation, with significant upregulation of Nestin and β3-tubulin. Our research demonstrates the importance of oxygen tension control during cell maintenance on the subsequent differentiation of both mouse and human PSCs, and highlights the differential effects.
Introduction
P
Embryonic stem cells (ESCs) are derived from the inner cell mass (ICM) of blastocysts [6] and one approach has been to attempt to recapitulate the in vivo microenvironment that surrounds these cells during normal development. A microenvironmental factor gathering interest in terms of enhancing ESC process productivity is oxygen tension. During normal development physiological oxygen levels sensed by the developing embryo are considerably lower than those routinely used to culture PSCs in the laboratory. It has been previously shown in our laboratory that atmospheric oxygen levels in standard incubators equate to about 20% O2 [7]. This is significantly higher than the low oxygen environment experienced by developing human (∼3%) and mammalian embryos (rabbit 8.7% and monkey 1.5%) in vivo [8,9]. Physiological oxygen levels have been shown to maintain proliferation and pluripotency of PSCs [10], and decrease spontaneous differentiation [11,12]. Additionally, low oxygen has enhanced the differentiation efficiency of both mouse and human PSCs into cardiac [13,14], endothelial [15], chondrogenic [16], hepatic [17], neuronal [18,19], and retinal progenitor [20,21] cell types. Although there have been a number of studies into the effects of low oxygen at either the maintenance or differentiation stage in isolation, the knock-on effects of preconditioning PSCs at physiological oxygen levels on subsequent high oxygen lineage commitment remain unclear. In this study, we investigated whether the prior maintenance of PSCs at low oxygen affects their commitment to early germ layer formation during subsequent high oxygen differentiation protocols. Both mouse and human PSCs were maintained for three passages at 2% O2, before assessing germ layer bias using an embryoid body (EB) differentiation protocol. Our results show that the implementation of a prior low oxygen maintenance step has a significant impact upon the subsequent differentiation of PSCs.
Materials and Methods
Cell lines and undifferentiated cell culture
Mouse embryonic stem cells
E14Tg2a and OCT4-GFP mouse embryonic stem cell (mESC) lines were kindly donated by Stem Cell Sciences Ltd (Cambridge, United Kingdom) and routinely cultured between P48–P56, and P28–P31 respectively, in sterile conditions in a Dual Heraeus HERAcell 150i incubator (Thermo Fisher Scientific, Waltham, MA) at 37°C/5% CO2. Cells were grown without a feeder layer in Nunc T25 tissue culture-treated flasks (Thermo Fisher Scientific) coated with 0.1% (w/v) gelatin (Sigma-Aldrich, St Louis, MO), in the presence of 5 mL culture media. Culture media were composed of 450 mL of Dulbecco's modified Eagle's medium (DMEM) supplemented with 1 mL 50 mM 2-β mercaptoethanol, 5 mL non-essential amino acids, 5 mL of 200 mM Glutamax (all Life Technologies, Carlsbad, CA), and 15% fetal bovine serum (SLI, Crawley, United Kingdom), and supplemented with 0.5 mL of 1×106 units/mL leukemia inhibitory factor (ESGROW® LIF; Millipore, Billerica, MA) after filter sterilization. Growth medium was exchanged every 24 h, and cells passaged every 3 days at a ratio of 1:8–1:12, depending on perceived confluency. Split ratios were adjusted in low oxygen cultures to 1:3–1:5, to ensure consistent seeding densities between conditions. Briefly, cells were washed with Dulbecco's phosphate-buffered saline (DPBS) without Ca2+/Mg2+ (Sigma) and detached by incubation with 0.25% Trypsin-EDTA (Life Technologies) for 3 min at 37°C. The trypsin was quenched and the resultant single cell solution resuspended in fresh growth media and re-inoculated into a new flask.
Human induced pluripotent stem cells
Human induced pluripotent stem cell (hiPSC) line MSUH001 was grown between passages P61–P69, with kind permission by Professor Jose B. Cibelli (Michigan State University), on Mitomycin-C (1 mg/mL; Sigma-Aldrich) inactivated mouse embryo fibroblasts (MEFs). MEFs were plated in Nunc T25 tissue culture-treated flasks (Thermo Fisher Scientific) coated with 0.1% (w/v) gelatin. Media consisted of knockout DMEM supplemented with 20% (v/v) knockout serum replacement, 1 mM
Oxygen tension control
In these studies we investigate the effects of a low oxygen step of 2% O2 on PSCs. An oxygen tension of 2% O2 was chosen for experimentation based on results that the oxygen tension in the uterus of several mammals ranged from about 1.5% to 8.7% O2 [8]. As a proof of concept study it was decided to initially investigate the lower end of this range. Ambient oxygen levels in the laboratory (20% O2) [7] are referred to here on in as “high oxygen” conditions, whereas “low oxygen” experimental conditions refer to 2% O2, and these were maintained using the oxygen control of the Dual Heraeus HERAcell 150i incubator. Cells were subjected to unavoidable ambient oxygen levels during feeding and passaging but care was taken to keep these steps to a minimum. Previous work from our laboratory indicated that it took ∼5 h for the oxygen tension at the growth surface to equilibrate to 2% O2 following a media change [7]. We also showed that differentiating mESCs within an automated platform operating under low oxygen conditions during both the cell-culture and liquid-handing phases eliminated the transient oxygen tension shifts, but critically yielded comparable cell number, viability and differentiation efficiency, when compared to the equivalent manual process [22].
Cell number and growth kinetics
Cell concentrations were measured using a VI-CELL Cell Viability Analyzer (Beckman Coulter, Brea, CA) according to manufacturer's instructions. For growth kinetic analysis, cells were plated at 1×105 cells/cm2 in Nunc six-well dishes (VWR, Radnor, PA). Three wells were harvested every 24 h over three passages of maintenance at either high or low oxygen and analyzed. The errors presented are standard deviations of three independent measurements and were calculated using Excel's data analysis software.
EB differentiation
Mouse embryonic stem cells
Undifferentiated mESCs were washed with DPBS without Ca2+/Mg2+, and single cells harvested as described above. Trypsin/EDTA was quenched using culture media minus LIF, and the cell suspension centrifuged. The supernatant was discarded and the cell pellet resuspended in culture media minus LIF and cells counted. About 2.25×106 cells were equally distributed between the wells of non-tissue culture-treated square Petri dishes (Sterilin, Newport, United Kingdom), where they were left to aggregate. Every 48 h EBs were resuspended in fresh media and transferred to fresh Petri dishes.
Human induced pluripotent stem cells
Undifferentiated hiPSCs were washed with DPBS without Ca2+/Mg2+ and single cells harvested. Formation of human EBs was carried out using AggreWell™ 400Ex plates; see manufacturer's guidelines (Stem Cell Technologies, Vancouver, Canada). About 4.7×106 cells were pipetted into each well, which equates to ∼1,000 cells per EB.
Directed cardiac differentiation
Cardiac differentiation was based on the methodology of Mummery et al. [23]. Briefly, E14 mESCs were trypsinized, washed, and diluted to 40,000 cells/mL in DMEM complete culture media minus LIF. The base of a 10 cm diameter bacterial dish was filled with PBS (for humidification). Approximately one hundred 20 μL drops of mESC suspension (equivalent to 800 cells/20 μL) were placed on the lid of the dish. The lid was inverted and placed on the PBS-filled base. Cells were incubated at 37°C in 5% CO2 for 5 days. After 5 days the single EBs that had formed in each droplet were each transferred to a single well of a gelatin-coated Nunc 96-well dish (VWR). Media were changed every 2 days and cultures observed for cardiomyocytes beating.
Metabolite analysis
Spent media were recovered from cultures, centrifuged to remove debris, and frozen until analysis could be performed. Glucose and lactate concentrations were measured using a multi-parameter Nova BioProfile analyzer (Nova Biomedical, St, Waltham, MA). These measurements were used to calculate specific rates of glucose consumption (GCR), lactate production (LPR), and the yield of lactose from glucose.
Calculation methods
Specific growth rate (μ)
μ values were calculated during the exponential growth phase of each culture condition using Equation (1):
where μ corresponds to the specific growth rate value at any given time interval Δt (in days, between sampling points i−1 and i), and X is the viable cell concentration at each corresponding time point. The exponential phase of each culture condition allowed the calculation of three specific growth rate values from which the average and standard deviation were calculated and presented in reciprocal days (day−1).
Specific rates of glucose consumption and lactate production
The specific GCR and LPR were calculated using Equation (2):
where qMet (pmol/cell/day) represents specific GCR or LPR. ΔMet is the variation in metabolite concentration during the time period Δt, and ΔXv the average viable cell concentration during the same period. The apparent yield of lactate from glucose (Y′ lactate/glucose) was calculated as the ratio between q lactate and q glucose. The errors presented are standard deviations of three independent measurements and were calculated using Excel's data analysis software.
Statistical analysis
Unless otherwise stated all cell culture experiments were conducted in triplicate (n=3). Data points are presented as the mean of three repeats, and error bars represent one standard error of the mean above and below the mean. Significant differences between data points from two conditions were calculated using a two-tailed, paired Student's t-test (Microsoft Excel). Where tests were carried out on log-normal data, such as fold change calculations during quantitative real-time polymerase chain reaction (qPCR) analysis, data were log transformed to assume a normal distribution of data, for which parametric tests were appropriate [24]. Significant differences between the means of three or more groups were tested using a one-way repeated measures ANOVA, followed by a post-hoc Tukey's Multiple Comparison Test.
For information on immunocytochemistry, flow cytometry and qPCR, please refer to the Materials and Methods section in the supplementary material. (Supplementary Data are available online at
Results
The effects of maintenance at low oxygen on mESCs
We cultured mESCs in parallel in either high (20%) or low (2%) oxygen conditions for three passages, and have reported here on data for the third and final passage unless otherwise stated. Low oxygen inhibited the proliferative capacity of mESCs and resulted in cells entering the stationary phase much earlier than high oxygen cultures (Fig. 1A). After 3 days, cell density in high oxygen was significantly higher (5.8±0.06×105 cells/cm2) than that at low oxygen (1.39±0.08×105 cells/cm2). This was associated with a significantly lower specific growth rate (μ) for mESCs maintained in low oxygen (0.36±0.07 day−1, compared to 0.81±0.06 day−1 at high oxygen; P<0.01) (Fig. 1B). During the first passage of low oxygen exposure there was inconsistent inhibition of cell growth, with final cell numbers after passage 1 significantly different from final cell numbers after passages 2 and 3, despite identical inoculation cell densities (Supplementary Fig. S1) (P<0.05). With growth appearing to stabilize after three passages, this supported the decision for a three passage-preconditioning step.

The effects of three passages of maintenance at 2% O2 on mESCs. Cells were grown in parallel at high oxygen (20%) or low oxygen (2%) tensions over three passages and then characterized to assess the effects of low oxygen on key stem cell characteristics.
Cell morphology analysis revealed notable differences. Both conditions yielded classic tightly packed colonies characteristic of mESCs (Fig. 1C), though cultures in low oxygen also comprised a substantial subpopulation. These cells were flatter with sharper edges (yellow arrows) and, although observed in ambient cultures, predominated at low oxygen.
Cultures were analyzed for pluripotent marker expression by immunocytochemistry (ICC) and both conditions were predominantly positive for Oct4, Nanog, and UTF1 (Fig. 1C). However, the flatter, spikier cells (yellow arrows) displayed decreased fluorescence intensities; an effect more pronounced at 2% O2. To investigate whether these effects were cell line-specific we subjected the Oct4-GFP mESC line to the same conditions. Interestingly, a similar population of flatter cells with sharper edges emerged and again predominated in response to low oxygen (Supplementary Fig. S2). These cells also showed a decrease in Oct4-GFP expression (yellow arrows).
We next analyzed the expression levels of the following genes by qPCR: Rex1 (ICM [25]); Nanog, Oct4, Sox2, and Utf1 (PSCs); FGF5 (pluripotent epiblast [26]); and Eomes (epiblast/primitive streak [27]) (Fig. 1G). Relative to high oxygen cultures there was significant downregulation of Rex1 by 38% (P<0.01), while Utf1, Nanog, and Sox2 revealed no significant difference. Although not significant there was a small increase in Oct4 expression (1.62±0.09-fold). Indicative of progression to commitment, Fgf5 and Eomes were significantly upregulated in low oxygen with a 6.37±0.4-fold (P<0.01) and 2.18±0.1-fold (P<0.01) increase respectively. Flow cytometry was used to further quantify the expression of Oct4, FGF5 and Eomes at the protein level (Fig. 1D–F). Although there was no significant difference in total percentage of Oct4-positive cells, analysis of fluorescence intensity revealed a shift in the Oct4-positive population after exposure to 2% O2 (Fig. 1D). Under standard conditions the positive population expressed predominantly high levels of Oct4 protein, as indicated by the sharp peak in fluorescence. Meanwhile, low oxygen exposure was associated with the pronounced expansion of a subpopulation of cells expressing lower levels of Oct4 (“Low”; Fig. 1D). We were able to arbitrarily gate on the separation between high-and low-expressing subpopulations, and saw that low oxygen induced a decrease in the high Oct4-expressing population from 76%±0.4%, to 63%±6%; and an increase in the proportion of lower-expressing Oct4-positive cells. It is possible that this lower Oct4-expressing population could correspond to the spikier cells with reduced Oct4 expression analyzed by ICC (Fig. 1C and Supplementary Fig. S2). Decreased oxygen tensions caused a notable increase in FGF5 and Eomes protein expression. At low oxygen we observed 48%±27% of cells expressing FGF5, compared with 30%±19% at 20% O2 (Fig. 1E). Low oxygen also caused a significant increase in Eomes expression from 50%±16% of cells at 20% O2 to 65%±17% at 2% O2 (P<0.05) (Fig. 1F). In summary, these data suggest that low oxygen is priming mESCs for progression to early commitment.
We proceeded to investigate how low oxygen had impacted hypoxia-inducible factor (HIF) gene regulation and metabolism of the cells. qPCR was used to analyze the transcription of HIF genes and their downstream targets. As expected, relative to standard cultures, there was significant upregulation of HIF genes in cells maintained in low oxygen following the three passage step at low oxygen (Fig. 1H). There was also a clear increase in target genes VEGF, Erythropoietin (EPO) and PGK-1. To investigate whether this was a transient response HIF gene transcription was measured over each passage of the three passage step (Supplementary Fig. S3). These data suggest a potentially transient state during passage 1 and stabilization thereafter between passage 2 and 3—supporting the choice for a three passage-preconditioning step. Cellular metabolism was also affected in low oxygen (Table 1). GCR was significantly raised (7±1 pmol/cell/day) compared with high oxygen (1.54±0.09 pmol/cell/day) (P<0.01). We also observed a significantly higher LPR (14±2 pmol/cell/day) at low oxygen compared with at high oxygen (2.5±0.2 pmol/cell/day) (P<0.001) and the apparent yield of lactate from glucose (Y′lactate/glucose) was higher under low oxygen at 1.8±0.4 compared with 1.6±0.1, suggesting an expected increase in glycolytic metabolism. Together, these results demonstrate that mESC cells responded to low oxygen maintenance as would be expected for most cell types in terms of HIF expression, HIF-responsive gene expression, and cellular metabolism.
Specific GCR and LPR (pmol/cell/day) were calculated, along with the apparent yield of lactate from glucose (Y′ lactate/glucose). Errors represent one standard deviation about the mean of three independent data points (n=3). A paired Student's t-test was used for statistical analysis: a P<0.01; b P<0.001.
GCR, glucose consumption rate; LPR, lactose production rate; mESC, mouse embryonic stem cell; ns, not significant.
The effects of low oxygen on subsequent EB differentiation of mESCs
To investigate the effects of low oxygen preconditioning on subsequent early differentiation we allowed mESCs to form EBs. EBs promote spontaneous differentiation of PSCs into all three germ layers enabling analysis of early formation of endodermal, ectodermal, and mesodermal lineages. As with our previous experiments, E14Tg2a mESCs were expanded in standard high oxygen conditions and then subjected to either low oxygen or high oxygen for three passages. Cells were then allowed to form EBs and left to differentiate for up to 72 h (Fig. 2A). We were particularly interested in the first 72 h of differentiation to observe the effects that could be directly attributed to the preconditioning step immediately prior. Notably, there were no significant differences in EB size or shape between the two conditions (Supplementary Fig. S4).
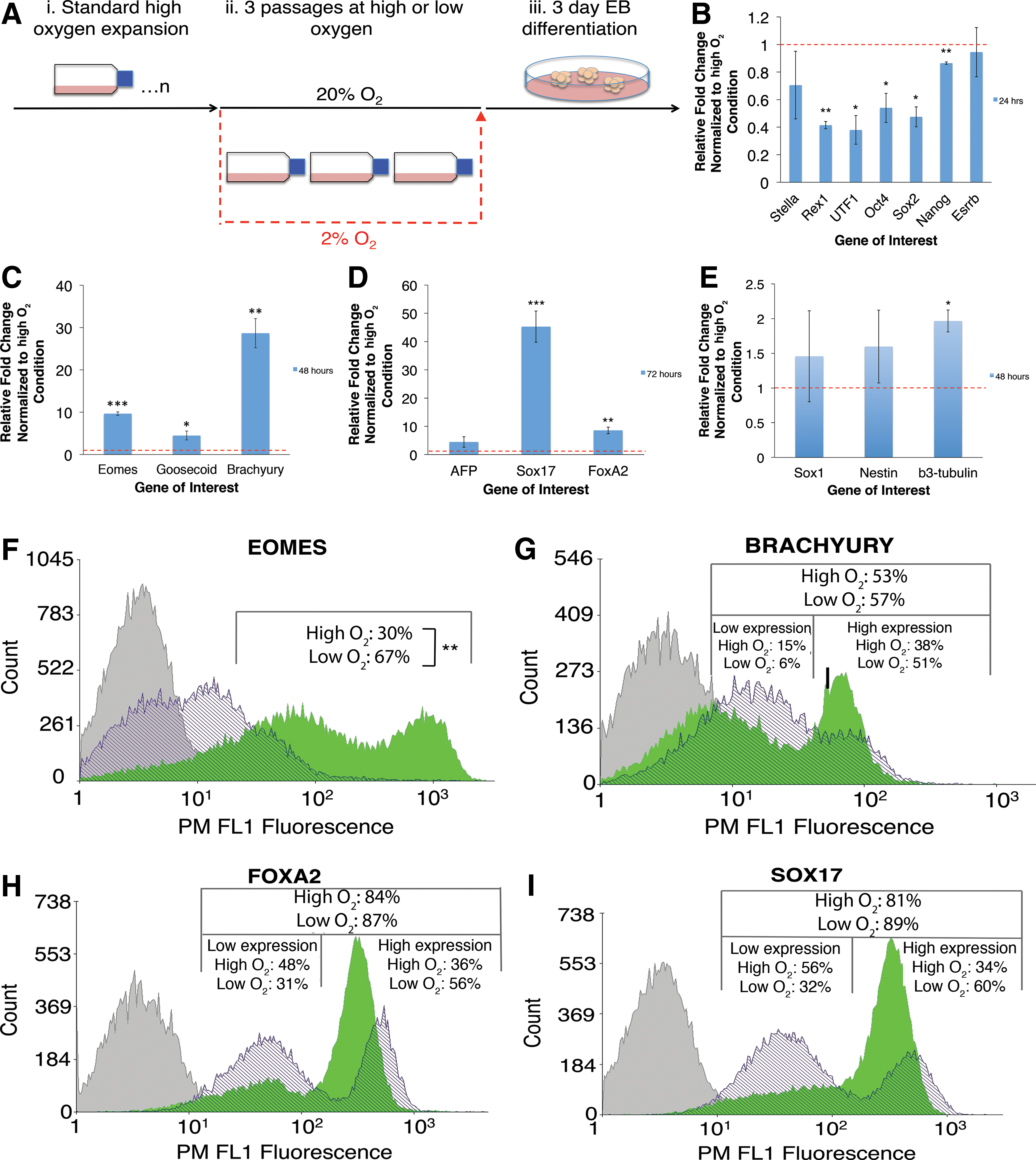
The effects of prior maintenance at 2% O2 on subsequent EB differentiation of mESCs.
We used qPCR to analyze gene expression during early EB differentiation. We observed a significantly faster downregulation of pluripotent genes in cells that had been previously maintained in low oxygen for three passages. After 24 h of differentiation there were significant decreases in Rex1, Utf1, Oct4, Sox2, and Nanog (Fig. 2B).
We went on to investigate the effects of prior low oxygen preconditioning on the subsequent expression of a panel of early differentiation markers. Firstly, confirming differentiation was occurring in both conditions, we observed an upregulation of mesoderm marker Brachyury after 24, 48 and 72 h in both conditions relative to undifferentiated mESCs. We also observed an upregulation of endoderm marker Sox17 after all time points in the low oxygen-preconditioned cells, and after 72 h in the high oxygen cells, relative to undifferentiated mESCs (Supplementary Fig. S5). We then compared the high oxygen EBs with the low oxygen-preconditioned EBs for the expression of a panel of germ layer markers. Low oxygen-preconditioned EBs showed significant upregulation of all mesoderm and endoderm markers, at all time points tested. Most significant upregulation of mesodermal markers Eomes, Goosecoid, and Brachyury was at 48 h showing a 9.67±0.4-fold (P<0.001), 4.49±1.1-fold (P<0.05), and 28.69±3.4-fold (P<0.01) increase respectively (Fig. 2C). The most significant upregulation of endodermal genes was observed at 72 h with AFP, Sox17, and FoxA2 showing a 4.44±1.9-fold, 45.2±5.6-fold (P<0.001), and 8.54±1.2-fold (P<0.001) increase respectively (Fig. 2D). Low oxygen preconditioning had a significantly less pronounced effect on ectodermal gene expression at all time points. Representative data are shown from 48 h, with Sox1, Nestin, and β3-tubulin showing just a 1.53±0.6-fold, 1.71±0.4-fold, and 1.97±0.2-fold (P<0.05) increase respectively (Fig. 2E).
To assess mesoderm and endodermal marker expression at the protein level, we employed flow cytometry to analyze Eomes, Brachyury, FoxA2, and Sox17 expression after 72 h (Fig. 2F–I). There was a significant increase in Eomes-positive cells from 31%±8% in standard conditions, to 67%±7% in cultures having been exposed to low oxygen prior to EB differentiation (P<0.01) (Fig. 2F). There were small increases in the total percentage of cells expressing Brachyury, FoxA2, and Sox17. However, further analysis of fluorescence intensity revealed a bimodal distribution that was comprised of high- and low-expressing subpopulations, which we gated on arbitrarily based on their clear separation. Cultures maintained in low oxygen prior to EB differentiation exhibited an increased proportion of cells expressing high levels of Brachyury (51%±3% compared with high oxygen cultures with 38%±9%) and a decreased proportion of cells expressing lower levels (6%±3% compared with 14%±4%) (Fig. 2G). Endodermal markers displayed similar expression patterns. Cultures pre-exposed to low oxygen exhibited an increased proportion of cells expressing high levels of FoxA2 (56%±9% compared with 36%±8%) and a decreased proportion of cells expressing lower levels (31%±5% compared with 48%±6%) (Fig. 2H). Finally, cells exposed to low oxygen showed increased levels of highly expressing Sox17-positive populations (60%±9% compared with 34%±11%), and decreased levels of lower-expressing positive populations (32%±9% compared with 56%±7%) (Fig. 2I). In summary, the qPCR and flow cytometry results revealed that low oxygen preconditioning of mESCs caused an increase in endodermal and mesodermal marker expression at both a transcriptional and translational level during subsequent high oxygen early EB differentiation.
The effect of 2% O2 preconditioning on subsequent directed cardiac differentiation
Results so far had indicated that prior exposure to low oxygen could speed up the onset of differentiation, in addition to influencing the lineage potential of mESCs. This led us to investigate whether three prior passages at low oxygen could impact the subsequent formation of more mature mesodermal lineages such as cardiomyocyte progenitors, using an established high oxygen protocol [23]. After 12 days of differentiation there was no clear difference in morphology, although further analysis by ICC revealed that cells that had been previously maintained in low oxygen no longer expressed any Oct4 protein (Fig. 3A). Meanwhile, cells passaged in high oxygen contained residual pockets of Oct4-expressing cells (yellow arrow) (Fig. 3A). Cells cultured in standard conditions had also retained diffuse expression of the early mesoderm marker Brachyury, whereas cells exposed to low oxygen displayed a much lower expression of Brachyury as indicated by fluorescence intensity, with only isolated pockets of cells retaining the higher expression level seen at high oxygen (yellow arrow) (Fig. 3A). These results indicate that prior maintenance of mESC at 2% O2 led to lower levels of immature pluripotent and mesodermal marker expression.
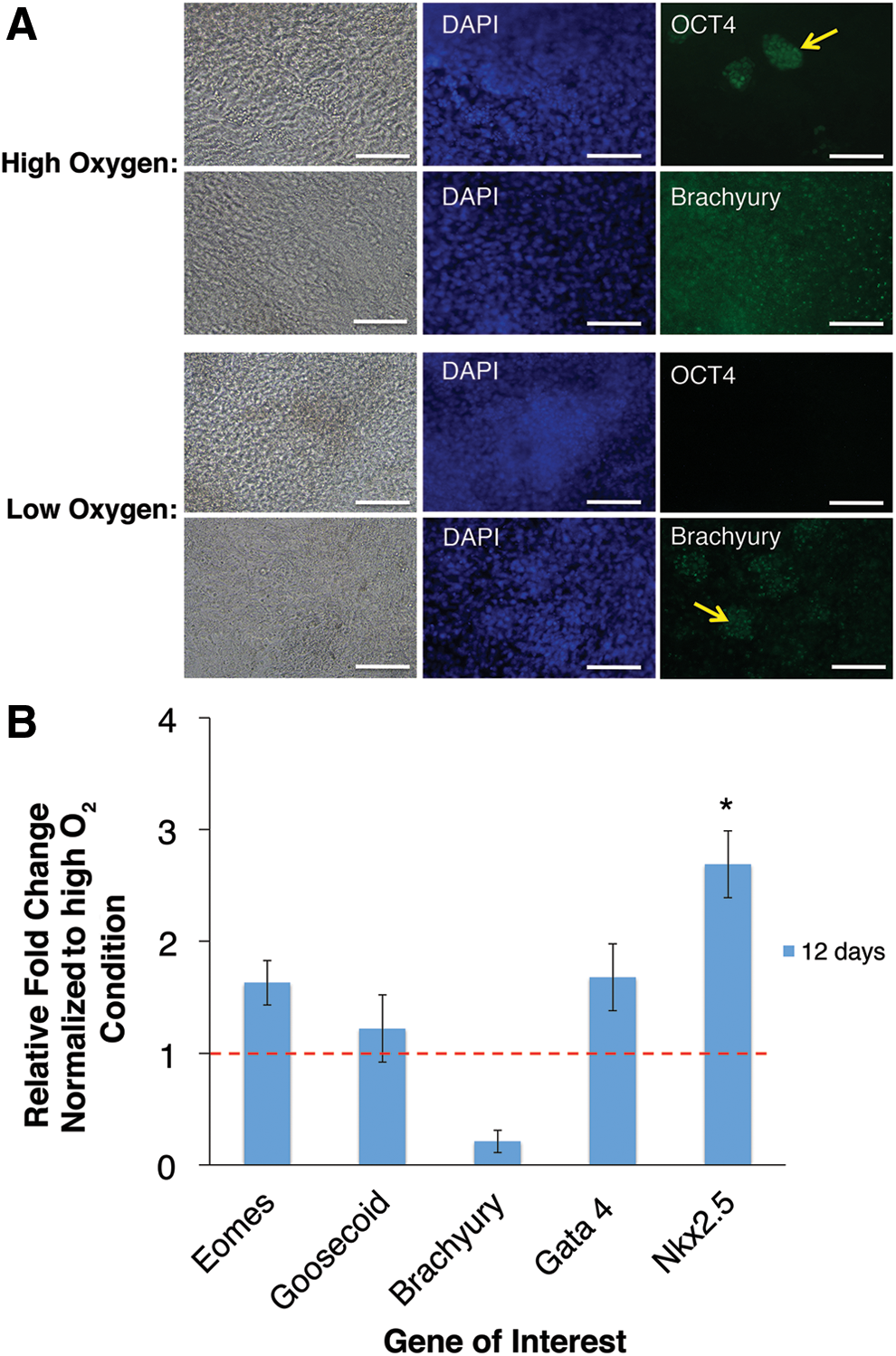
The effects of 2% O2 preconditioning on subsequent directed cardiac differentiation. mESCs subjected to three passages of prior maintenance at high or low oxygen were subsequently differentiated into cardiomyocyte progenitors using an established normoxic protocol.
Having observed marked differences in protein expression we went on to analyze the expression of a wider panel of mesodermal (Eomes, Goosecoid, and Brachyury) and early cardiac (Gata4 and Nkx2.5) genes by qPCR. Cultures that had been previously maintained for three passages in low oxygen showed some upregulation of Eomes and Goosecoid and downregulation of Brachyury reinforcing the ICC results. Cardiac markers were also upregulated with Gata4 showing a 1.7±0.3-fold increase, and Nkx2.5 significantly increased by 2.7±0.3-fold (P<0.05) (Fig. 3B). These results indicate that the maintenance of mESC under low oxygen conditions can have a significant impact upon the formation of more mature cell types during subsequent longer differentiation processes.
The effects of low oxygen on human PSCs
As a means of comparison we went on to study the effects of low oxygen exposure on human PSCs. For these experiments we used hiPSCs, which were grown on a feeder layer of MEFs for three passages at either 20% O2 or 2% O2. In agreement with others our results showed that hPSCs cultured at atmospheric oxygen were inclined to spontaneously differentiate [12] (yellow arrow) (Fig. 4A). In contrast, cultures maintained at low oxygen for three passages showed less spontaneous differentiation, and displayed more clearly defined borders against the MEF background (Fig. 4A). In agreement with others [28] there was a decrease in the specific growth rate of 0.14±0.09 day−1 at low oxygen, compared with 0.42±0.08 day−1 in standard conditions (Fig. 4B) (P<0.05).
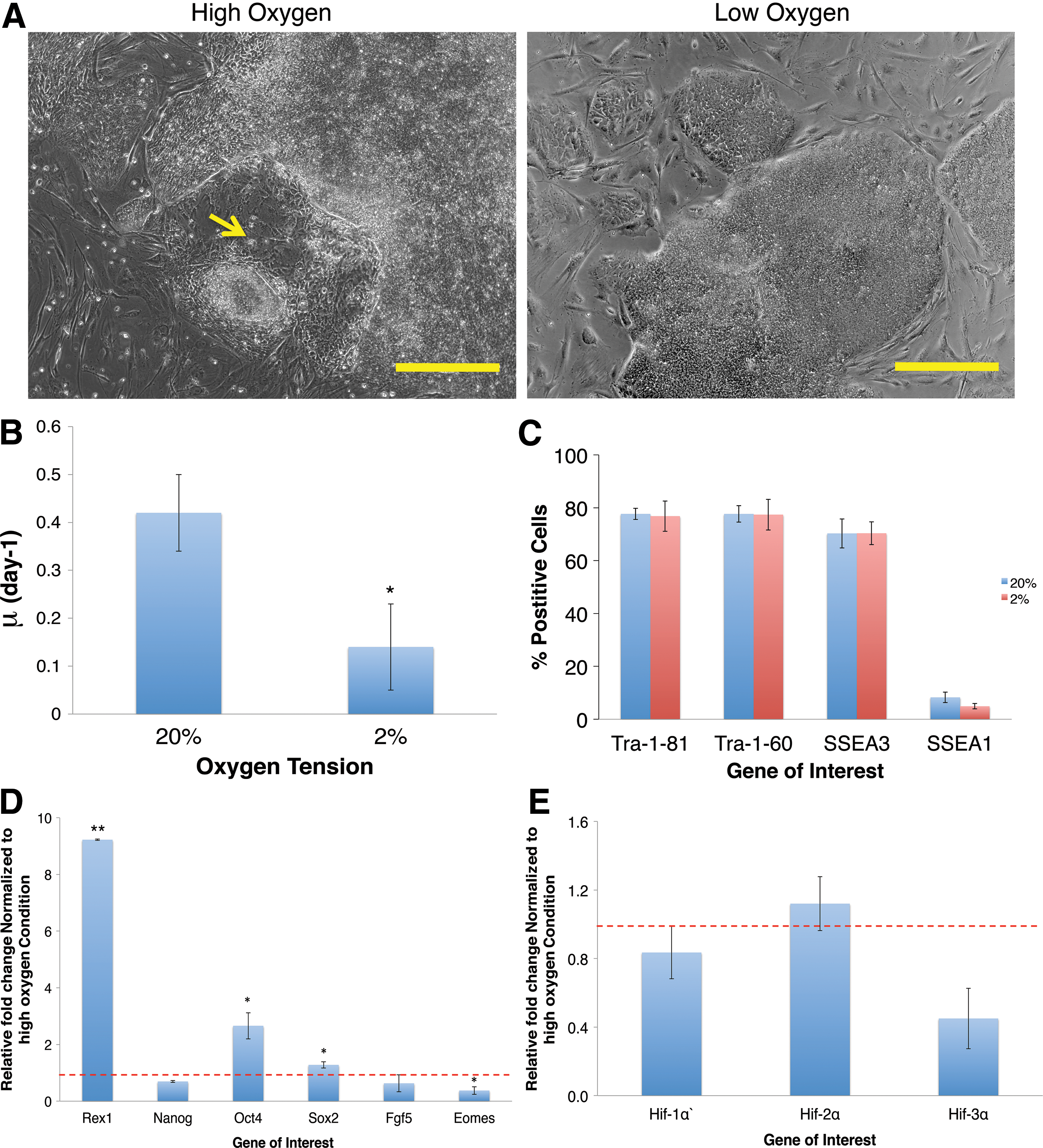
The effects of three passages of maintenance at 2% O2 on human PSCs. Human induced pluripotent stem cells were grown in parallel at high oxygen (20%) or low oxygen (2%) tensions over three passages and then characterized to assess the effects of low oxygen on stem cell characteristics.
Cells maintained under both conditions were analyzed by flow cytometry for pluripotent cell surface antigens Tra-1-81, Tra-1-60, and SSEA3, and early differentiation marker SSEA1 (Fig. 4C). Expression of Tra-1-81, Tra-1-60, or SSEA3-positive cells was maintained at a similar level in both conditions, although there was a notable though insignificant decrease in SSEA1 expression in cells exposed to low oxygen (high: 9%±2%; low: 5%±0.2%).
We next analyzed the expression levels of a panel of pluripotent and early differentiation genes by qPCR (Fig. 4D). Low oxygen significantly upregulated Rex1 expression by 9±0.03-fold (P<0.01). There was also significant upregulation in Oct4 (2.66±0.46-fold, P<0.05) and Sox2 (1.28±0.11-fold, P<0.05) expression. Meanwhile, low oxygen caused a downregulation of FGF5 and Eomes expression by 27% and 62% (P<0.05) respectively. These results were in contrast to mESCs where low oxygen had a mixed impact upon pluripotent marker expression and caused a clear increase in FGF5 and Eomes expression.
We proceeded to investigate how low oxygen had impacted HIF regulation and metabolism of the hPSCs. qPCR was used to analyze the transcription of HIF genes following the three passage preconditioning step. In agreement with others, there was no significant effect on HIF gene transcription following exposure to low oxygen [11,29,30] (Fig. 4E). However, similar to mESCs, cellular metabolism was affected in low oxygen (Table 2). GCR was raised to 2.83±2.24 pmol/cell/day compared with high oxygen levels of 0.8±0.24 pmol/cell/day. We also observed a significantly higher LPR in low oxygen (6.58±2.74 pmol/cell/day) compared with that in high oxygen (1.52±0.52 pmol/cell/day) (P<0.05) and the apparent Y′lactate/glucose was higher under low oxygen at 2.32±0.89 compared with 1.9±0.45, suggesting, alongside upregulation of pluripotent markers in the human cells, there was an expected increase in glycolytic metabolism [31]. Together, these results demonstrate similar responses to those reported elsewhere of hPSCs to low oxygen, in terms of pluripotency, HIF expression, and cellular metabolism.
Specific GCR and LPR (pmol/cell/day) were calculated, along with the apparent yield of lactate from glucose (Y′ lactate/glucose). Errors represent one standard deviation about the mean of three independent data points (n=3). A paired Student's t-test was used for statistical analysis: a P<0.05; hiPSC, human induced pluripotent stem cell.
The effects of low oxygen on subsequent differentiation of hPSCs
Having studied the effect of low oxygen on the culture of hiPSCs, we looked to analyze the knock-on effects during subsequent EB differentiation in atmospheric oxygen. In contrast to mESCs, hiPSCs having been preconditioned for three passages at low oxygen, retained higher levels of pluripotent gene expression, with Rex1, Oct4, and Nanog all significantly upregulated by 2.9±0.3-fold (P<0.05), 1.78±0.3-fold (P<0.05) and 2.93±0.5-fold (P<0.05) respectively after 5 days, relative to standard cultures (Fig. 5A).
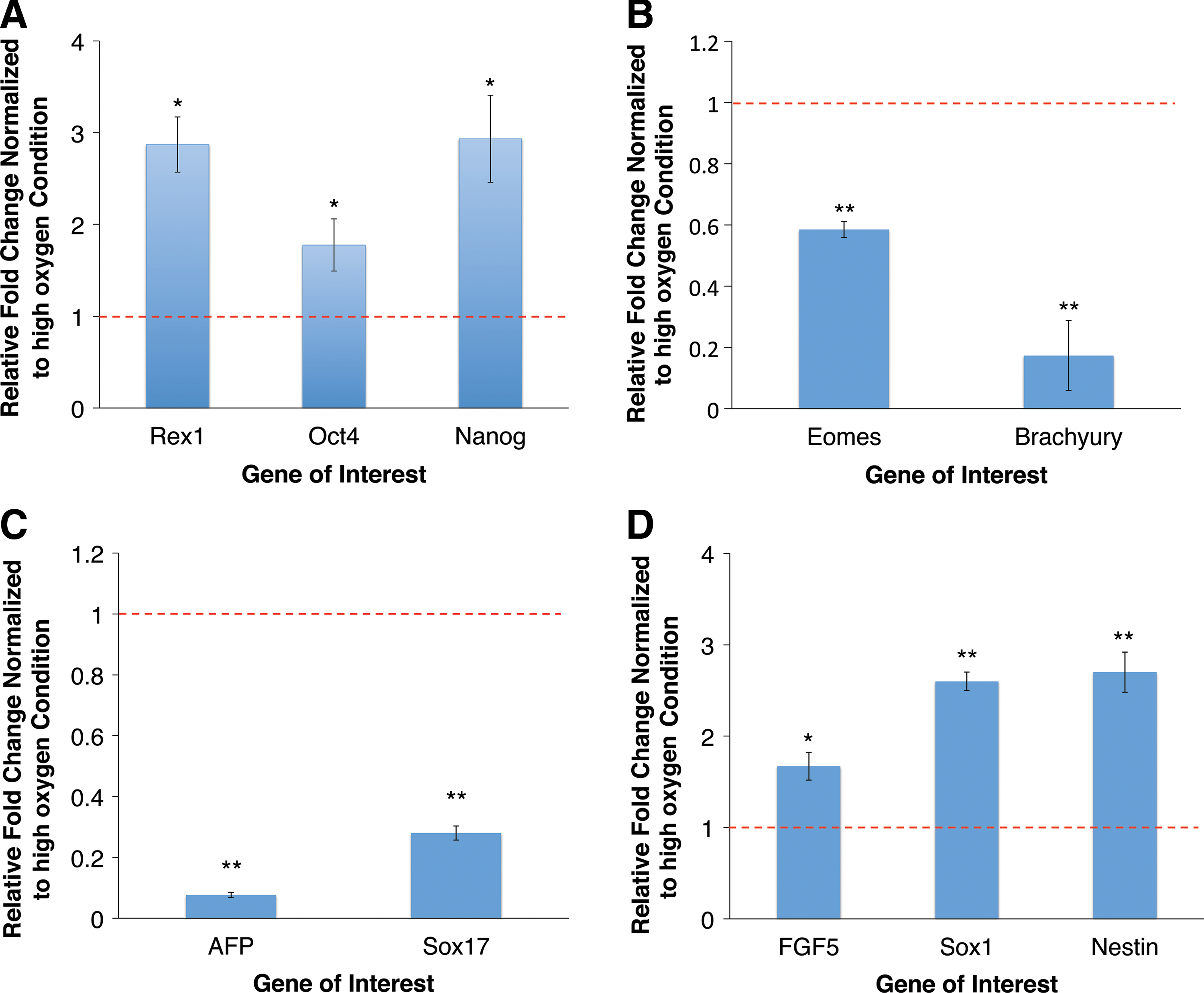
The effects of prior maintenance at 2% O2 on subsequent EB differentiation of hiPSCs.
We next observed the effects on lineage commitment during the 5 day EB differentiation. Contrary to results in mouse, prior exposure to low oxygen resulted in significantly lower levels of mesoderm and endodermal gene expression relative to high oxygen control cultures. Eomes and Brachyury showed 41% (P<0.01) and 83% (P<0.01) lower expression levels respectively (Fig. 5B), while AFP and Sox17 showed 92% (P<0.01) and 72% (P<0.01) lower expression levels respectively (Fig. 5C). Conversely, there was significant upregulation in early ectodermal gene expression (Fig. 5D). FGF5, Sox1, and Nestin were upregulated by 1.67±0.15-fold (P<0.05), 2.6±0.1-fold (P<0.01), and 2.7±0.2-fold (P<0.01) respectively, relative to high oxygen cultures. These results revealed that prior low oxygen preconditioning resulted in an increased propensity for ectodermal gene upregulation, and a decrease in mesoderm and endoderm marker expression during subsequent differentiation—the opposite to what was observed in mouse. To assess whether priming at low oxygen resulted in the inhibition of hiPSC differentiation we compared the expression levels of Rex1, Nanog, and Eomes in day 5 EBs with pre-differentiation hiPSCs (Supplementary Fig. S6). After 5 days of differentiation, pluripotent marker Rex1 was downregulated in both conditions but remained higher in cells that had been preconditioned at low oxygen. However, the levels of Nanog were very similar in both preconditioned and high oxygen conditions with a modest drop in the expression of this pluripotent marker when compared with undifferentiated cells. Levels of the early mesodermal differentiation marker Eomes were lower in preconditioned cells when compared with high oxygen and undifferentiated cells. Taken together, our results show that low oxygen priming results in the preferential formation of ectoderm over mesoderm and endoderm lineages. However, further studies using directed differentiation protocols will be required to uncover whether priming inhibits differentiation during the formation of specific lineages.
Discussion
In recent years scientists have scrutinized the surrounding environment of the ICM cells for clues to the optimal culture conditions of their in vitro counterparts. Physiological oxygen levels have been demonstrated to be significantly lower than the atmospheric oxygen tensions routinely used since PSCs were first derived [8,9]. Recent research into oxygen tension in stem cell culture has focused on either the expansion [11,12,29 –40] or the subsequent differentiation [7,13,16,17,19 –21,41 –45] of PSCs in isolation. There appears to be, however, a lack of studies investigating the knock-on effect of low oxygen preconditioning of PSCs on subsequent lineage commitment. To further understand these interactions we investigated how prior maintenance in low oxygen conditions affected the subsequent differentiation of PSCs.
Culturing cells at 2% O2 was associated with clear population changes in mESCs. Prior to low oxygen exposure mESCs were Rex1 +, indicative of a naïve pluripotent state [6,46]. Following three passages at 2% O2 qPCR and flow cytometry analysis revealed maintenance of pluripotency but with a decrease in Rex1 mRNA expression, and upregulation of primed, primitive ectoderm markers FGF5 [26] and Eomes [27]. This was accompanied by the emergence of a second population of flatter cells, with sharper edges indicative of onset of differentiation [33,34]. These low oxygen-induced effects have been reported by others in addition to an inhibited proliferation rate, and an increase in glycolytic metabolism [32 –34,36,37,47]. It is known that mESCs are comprised of a dynamic heterogeneous population that spans the pluripotent state from naïve ICM-like cells through to primed epiblast or primitive ectodermal equivalents [25,48 –50]. With this in mind there are parallels between this observed low oxygen-induced population and the “primed state” of ESC pluripotency [6,34,46]. It is known that the ICM and the epiblast can be distinguished by the upregulation of Rex1 and FGF5 respectively [51]. Meanwhile, Toyooka et al. have proposed that sorted Rex1 +/FGF5− and Rex1−/FGF5+ mESC populations have features of the pre- and postimplantation embryo respectively [25]. This supports the notion that reduced oxygen is skewing the dynamic pluripotent equilibrium toward a primed, epiblast-like fate. Indeed, based on gene expression, morphology, and proliferative capacity, our cells show some similarities to pluripotent epiblast stem cells (EpiSCs) [52,53]. Moreover, it has been shown that the conversion of mESCs to mEpiSCs through the removal of LIF and the provision of bFGF can be enhanced by exposure to low oxygen [54]. In summary, our results are broadly consistent with previous studies and show for the first time that the maintenance of mESCs in the presence of LIF under low oxygen conditions alone is sufficient to prime cells toward an EpiSC-like phenotype.
We found that low oxygen preconditioning of mESCs for three passages resulted in the earlier onset of germ layer formation during subsequent EB differentiation (Fig. 2). This was in agreement with studies showing that partially primed intermediate epiblast stem cells undergo increased rates of differentiation when compared with ground state mESCs [48]. We specifically saw an increase in mesoderm and endodermal differentiation. It has been shown that co-stimulation of mESCs by LIF and BMP4 is required to sustain self-renewal and multilineage differentiation potential via a differentiation blockade of opposing lineages [55]. HIF-1α inhibits LIFR and thus partially downregulates LIF-STAT3 signaling [32], while BMPs block neural differentiation [56,57]. Therefore, in low oxygen conditions with LIFR partially inhibited, overriding BMP4 signaling may inhibit ectoderm and promote mesoderm and endodermal differentiation. This may also explain why low oxygen expansion prior to neural differentiation did not lead to an increase in the yield of GABAergic neurons [58]. Additionally, we observed a small increase in Oct4 mRNA expression after three passages at 2% O2. Others have shown that a <2-fold increase in Oct4 such as this promoted differentiation into mesoderm and endoderm [59].
Aside from a similar decrease in proliferation, we observed a stark difference in the response of hPSCs to low oxygen compared to mouse. In agreement with others, we have shown that low oxygen decreases spontaneous differentiation in hPSCs while upregulating pluripotent genes [11,31,39,45,60], though it shows no obvious effect on HIF gene regulation [11]. For the first time we have shown that 2% O2 significantly increases the expression of Rex1, an ICM marker, and significantly downregulates the expression of lineage committed marker Eomes. Westfall et al. did observe a similar trend reporting a small increase in Rex1 and a sporadic decrease in Eomes at low oxygen. They also observed very low FGF5 levels with no significant differences between 20% and 4% O2. However, here we have observed a notable, though insignificant, decrease in FGF5 expression at 2% O2. The differences in potency of effect between the two sets of experiments may be attributable to differences in oxygen tension tested. They investigated effects at 4% O2, whereas we looked at the effects of 2% O2. The low oxygen-induced push back toward a more naive pluripotent state described here is the opposite to what was observed in mESCs. Given that Hif-2α directly upregulates Oct4 expression in both human [11] and mouse [61], it is possible that the self-renewal promoting effects of low oxygen seen in hPSCs and other stem cells types [11,62 –64] are counterbalanced by Hif-1α-mediated inhibition of LIFR/STAT3 signaling in mESCs [32,33], leading to early priming (Supplementary Fig. S7). It has been shown that ground state pluripotency in mouse can be maintained in the absence of LIF via inhibitors of FGF4/ERK and GSK3 signaling [65,66]. Combinations of these inhibitors in reduced oxygen mESC culture have been shown to maintain pluripotency and proliferative capacity, and critically eliminate induction of primed markers supporting the notion that the priming effect observed in mESCs is a LIF-dependent trait [34,37]. Indeed, LIF-independent hPSCs are instead primed toward a naïve stem cell fate when exposed to low oxygen, opposing the natural tendency to drift toward differentiation (for illustrative diagram see Supplementary Fig. S7). This in turn may have led to a preference for ectodermal differentiation in subsequent EB differentiation (Fig. 5). Increased Oct4/Sox2 expression during low oxygen culture of hPSCs may explain this ectodermal bias. Their downstream target Lefty2 is increased in reduced oxygen [30] and via Nodal inhibition [67] is known to prevent differentiation toward mesoderm and endodermal lineages. With substantial residual pluripotent gene expression 5 days into EB differentiation we hypothesize that the default differentiation pathway becomes ectoderm.
This research has clear implications for stem cell processing, and it demonstrates the importance of recapitulating the in vivo niche as a basis for stem cell bioprocess development [5]. Research in our laboratory has specifically examined the role of oxygen tension in stem cell processes and confirmed that differentiating cells in low oxygen showed a significant upregulation in the production of neural cell types [7]. As oxygen tension varies both spatially and temporally during mammalian development [68] it has became apparent that step changes in oxygen tension during differentiation are necessary in improving the yield and efficiency of a process [19]. Our current work has now shown that to fully optimize a process it is critical to take into account the prior maintenance of PSCs to influence subsequent differentiation. Supporting this notion is our differentiation of mESCs into cardiomyocytes. Through the implementation of a low oxygen-preconditioning step prior to the onset of cardiac differentiation, we induced a significant upregulation in cardiomyocyte marker Nkx2.5 (Fig. 3). We also observed a decrease in the number of contaminant pluripotent cells. The concept of low oxygen preconditioning has been studied in other cell types such as human adipose tissue mesenchymal stem cells where an enhanced ability to differentiate into adipocytes and osteocytes was reported [64]. To our knowledge, however, the preconditioning step has not yet been implemented with PSCs.
Conclusions
• Maintenance of mESCs under low oxygen conditions results in the inhibition of proliferation and early priming toward an EpiSC-like phenotype.
• Low oxygen-induced priming of mESCs results in the promotion of mesoderm and endodermal differentiation during subsequent differentiation.
• In contrast, hPSCs cultured at low oxygen are pushed toward a more naïve pluripotent state, and they are primed for subsequent ectoderm differentiation.
• Through investigation of oxygen tension, we have shown that control of microenvironmental factors at the maintenance stage can have knock-on effects for subsequent lineage commitment. This is critical for future bioprocess development.
Footnotes
Acknowledgments
The authors would like to thank Professor Jose B. Cibelli (Michigan State University) for permission to use MSUH001 hiPS cell line for this research and the Spanish Stem Cell Bank for providing the cell line. Also Professor Peter Andrews (University of Sheffield) for kind donation of the SSEA1 antibody. We appreciate the advice of Tomas Adejumo from UCL concerning flow cytometry analysis. K.F. would like to thank the BBSRC for a PhD studentship.
Author Disclosure Statement
The authors indicate no potential conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.