
Abstract
Vasculogenesis/angiogenesis is one of the earliest processes that occurs during embryogenesis. ETV2 and SOX7 were previously shown to play a role in endothelial development; however, their mechanistic interaction has not been defined. In the present study, concomitant expression of Etv2 and Sox7 in endothelial progenitor cells was verified. ETV2 was shown to be a direct upstream regulator of Sox7 that binds to ETV2 binding elements in the Sox7 upstream regulatory region and activates transcription. We observed that SOX7 over-expression can mimic ETV2 and increase endothelial progenitor cells in embryonic bodies (EBs), while knockdown of Sox7 is able to block ETV2-induced increase in endothelial progenitor cell formation. Angiogenic sprouting was increased by ETV2 over-expression in EBs, and it was significantly decreased in the presence of Sox7 shRNA. Collectively, these studies support the conclusion that ETV2 directly regulates Sox7, and that ETV2 governs endothelial development by regulating transcriptional networks which include Sox7.
Introduction
V
The Ets gene family was shown to have essential roles in diverse biological processes, including angiogenesis, vasculogenesis, hematopoiesis, neurogenesis, and myocardial development [14 –19]. In situ hybridization and RT-PCR techniques demonstrated that Etv2 expression is restricted to the endocardium/endothelium of the E8.5 and E9.5 embryo, and that this expression is extinguished by E11.5 [14]. The Etv2 homozygous mutant embryos were lethal by E9.5, and lacked the endocardial/endothelial lineage [14]. Recent in vivo and in vitro results further support the hypothesis that ETV2 is a critical factor for the specification of the endocardial/endothelial and hematopoietic lineages [15,20 –23].
The Sry-related HMG box (Sox) gene family encodes transcription factors that regulate specification and differentiation of various developmental processes. Sox7, Sox17, and Sox18 comprise the Sox-F family, which was shown to play a critical role in formation of the endodermal lineage, hematopoietic stem cell regulation, cardiovascular development, and, more recently, arterial specification [5,24 –30]. All three Sox-F members were shown to be expressed in vascular endothelial cells [24,25,27,31,32]. Sox18-null mice were not viable after significant back-crossing onto a pure C57BL6 background and died at E14.5 [33]. Other reports showed that point mutations in Sox18 in both humans and mice cause syndromes with cardiovascular and hair follicle defects; however, previously generated Sox18-null mice were viable and display no obvious cardiovascular defects [34]. These results may be due to the generation of a dominant negative protein by the point mutations, while defects in the Sox18-null mice may be rescued by the compensatory action of other Sox-F members [5,27,34 –36]. Previously described redundancy of Sox17 and Sox18 as well as Sox7 and Sox18 in vascular development, arterial-venous identity, and cardiovascular development further support this theory [5,27,34 –36]. SOX7 was recently shown to be involved in the transcriptional regulation of the hemogenic endothelium [37], and it has been implicated in human cardiovascular development [38]. Although the downstream effects of the Sox-F family have begun to be elucidated, the upstream regulation of the Sox-F family members has yet to be defined.
In the present study, we demonstrate that ETV2 regulates the expression of Sox7 by directly binding to its upstream regulatory region. We further establish that the Etv2 developmental pathway results in the formation of endothelial progenitor cells, and it increases angiogenic potential of embryonic bodies (EBs). Collectively, these studies enhance our understanding of molecular networks that govern discrete stages of early endothelial development.
Materials and Methods
Generation of embryonic stem cell lines
Standard techniques were used for isolation and propagation of embryonic stem (ES) clones [39]. Etv2 mutant ES cells were derived from day 3 Etv2 mutant and WT blastocysts as previously described [40]. ES cells (A2loxCre) with doxycycline-inducible expression of Etv2 or Sox7-Myc were generated as previously described [41]. Briefly, the expressing vector pLox was integrated into the X chromosome of the Ainv15 ES cells placing the cDNA under the control of tetracycline-responsive element (TRE). In this system, the addition of doxycycline (dox) causes the reverse tetracycline transactivator to bind to TRE, resulting in over-expression.
EB culture and expression profiles
EBs were prepared using the hanging drop technique and the appropriate mES cell lines, and then cultured in suspension on a rotating plate beginning on day 2. Dox (1 μg/mL final concentration) was added to the culture at designated time points to induce protein expression [41]. Gene expression at each time point was analyzed using quantitative RT-PCR using 7900 Applied Biosystems sequence detection system (Foster City, CA). All Taqman gene expression assays were purchased from Applied Biosystems. Taqman gene expression assays included Gapdh (Mm99999915_g1), Etv2 (Mm 00468389_m1), Sox7 (Mm00776876_m1), Cdh5 (Mm00486938_m1), Pecam1 (Mm00476702_m1), and Tek (Mm01256904_m1).
Single-cell qRT-PCR
EBs were prepared using the hanging drop technique with control mES cells (A172lox), and then cultured in suspension on a rotating plate beginning on day 2. Day 5 EBs were harvested and dissociated as previously described [41]. Briefly, EBs were incubated in Collagenase Type I (17100-017; Gibco, Grand Island, NY) and triturated multiple times over a 10 min interval. Cells were washed, filtered, and stained with anti-Flk1-PE (12-58212-83; eBioscience, San Diego, CA), and anti-Cdh5-APC (17-1441-80; eBioscience). Double positive (Flk1+/Cdh5+) were sorted on an FACSAria (BD Biosciences, San Jose, CA). Sorted cells were stained with LIVE/DEAD viability/cytotoxicity kit (L-3224; Life Technologies, Grand Island, NY) and loaded on a 10–17 μm Auto Prep Integrated Fluidic Circuit (IFC; Fluidigm, San Francisco, CA) to capture single cells using the C1 Single cell Auto Prep System (Fluidigm).
Cells were loaded at 250,000 cells/mL according to the manufacturer's protocol. The IFC was imaged on a Nikon Tie Deconvolution microscope system to determine capture efficiency before proceeding to cDNA synthesis using the Ambion single cell to CT kit (PN 4458237; Life Technologies), and targeted amplification was performed for 18 cycles on a C1 Single Cell Auto Prep system (Fluidigm) with Taqman gene expression assays Gapdh (Mm99999915_g1), Etv2 (Mm 00468389_m1), and Sox7 (Mm00776876_m1). Fourty live cells were selected to load on a Dynamic Array IFC 48.48 chip (Fluidigm), and Etv2, Sox7, and Gapdh were quantified in quadruplicates on a Biomark HD system (Fluidigm) using Taqman Fast Universal PCR Master Mix (4352042; Applied Biosysystems, Austin, TX) according to the standard manufacturer's protocol, using the same Taqman probe used in the preamplification. The data were analyzed using Fluidigm's real-time PCR analysis software.
Protein analysis by western blot
Protein expression was determined by western blot analysis as previously described [42]. Antibodies used were anti-human Sox7 (AF2766; R&D Systems, Minneapolis, MN), anti-PECAM-1 (M-20) (sc-1506; Santa Cruz Biotechnology, Santa Cruz, CA), anti-CDH5 (C-19) (sc-6458; Santa Cruz Biotechnology), anti-SOX17 (S-20) (sc-17355; Santa Cruz Biotechnology), anti-SOX18 (H-140) (sc-20100), anti-α-tubulin (T5168; Sigma, St. Louis, MO), and anti-c-myc (Cat #11667149001; Roche, Indianapolis, IN).
Chromatin immuno-precipitation assays
EBs were prepared as described earlier using ETV2-HAX3 over-expressing cells and maintained in differentiation medium. Over-expression was induced by dox at day 3.0, and EBs were harvested at day 4.0. Chromatin DNA isolation from EBs and protein-DNA complex immuno-precipitation using anti-HA antibody (Y-11, sc-805×) or rabbit IgG as control (Santa Cruz Biotechnology) were performed as previously described [43]. SYBR Green qRT-PCR was performed using PCR master mix from Applied Biosystems (Cat #4309155) and primers specific to the mouse Sox7 upstream regions: Region I Fwd 5′ CGCTCCTCACCCAAATGTAT 3′, Region I Rev 5′ AAGAATGACTGGGTCAAGGAAA 3′; Region II Fwd 5′ TGAGACCTAGGGAGCTGATGC 3′, Region II Rev 5′ GTTGCTATTGGCTTGCTCCAC 3′; Region III Fwd 5′ TATCGCCGGGTTTTAGGATTA 3′, Region III Rev 5′ GCTTTAGACACACCCCACTGT 3′; Gapdh Fwd 5′ TGACGTGCCGCCTGGAGAAA 3′, and Gapdh Rev 5′ AGTGTAGCCCAAGATGCCCTTCAG 3′.
Electrophoretic mobility shift and transcriptional assays
The electrophoretic mobility shift assays (EMSA) were performed using the Gel Shift assay Core Kit (E3050; Promega, Madison, WI). HA-tagged Etv2 protein was in vitro translated by TNT-coupled transcription rabbit reticulocyte translation system (L5010; Promega). HA-ETV2 was incubated with 32P-labeled synthetic oligonucleotides containing putative EBE (Etv2 binding element) sequences at room temperature for 10 min, and separated on a 4% acrylamide nondenaturing gel in Tris/Borate/EDTA buffer. For the supershift assays, anti-HA antibody (sc-805; Santa Cruz Biotechnology) was added to the reaction for 20 min at room temperature after the formation of the protein-DNA complex. Synthetic nucleotides generated were as follows: EBE-1: 5′ CTGAGACTTCCTGAAGTT 3′, EBE-2: 5′ TCCTCTGGGAAATGGCCC 3′, EBE-3: 5′ CCTGGACTTCCT 3′, EBE-1mut: 5′ CTGAGACcggcTGAAGTT 3′, EBE-2mut: 5′ TCCTCTGgtccATGGCCC 3′, EBE-3mut: 5′ CCTGGACcggcT 3′.
Recombinant reporter constructs and cell transfection
The recombinant reporter gene construct was obtained by cloning a PCR-amplified 241 bp fragment Region II of the Sox7 promoter (Fwd: 5′ CCATTTGATTTCAGCGTCCAGGC, Rev: 5′ CCACCTACAGGAAGTCCAGGATGAGC 3′) into the SacI and XhoI sites of the pGLT vector (pGL3basic; Promega, with hsp70 TATA boxes inserted at the HindIII site). Mutation of the EBEs was performed by site-directed mutagenesis. Luciferase constructs and increasing amounts of HA-tagged-Etv2, Ets1, or Erg expression vectors were transfected into C2C12 myoblasts using the Lipofectamine/Plus reagent method according to the manufacturer's protocol (Invitrogen, Carlsbad, CA) and incubated for 24 h. Cells were then harvested in passive lysis buffer and assayed using the Dual-Glo Luciferase Assay System (E2920; Promega).
Quantification of endothelial progenitor cells by flow cytometry
Day 6 EBs were harvested and dissociated as previously described [41]. Briefly, EBs were incubated in Collagenase Type I (17100-017; Gibco) and triturated multiple times over a 10 min interval. Cells were washed, filtered, and stained with anti-Flk1-PE (12-58212-83; eBioscience) and anti-Cdh5-APC (17-1441-80; eBioscience). Stained cells were analyzed on an FACSAria (BD Biosciences) to quantify double positive (Flk1+/Cdh5+) endothelial progenitors cells (EPCs) [41].
ShRNA knockdown of Sox7
The shRNA vector pLKO.1 containing the mouse Sox7-specific RNAi sequence 5′ CCTGGCTTTGACACCTTGGAT 3′ (TRCN0000086052) was introduced into the Etv2 over-expressing mES cells mentioned earlier by lentiviral transduction after replacement of the puromycin resistance gene in pLKO.1 with eGFP. Infected mES cells were sorted twice while selecting for GFPbright cells, resulting in an mES cell line with constitutive Sox7 shRNA expression and inducible Etv2 over-expression.
EB culture in Type I Collagen gel for endothelial sprouting assessment
EBs were prepared by the hanging drop method using the indicated mES lines, and cultured in suspension on a rotating plate for 7 days. After 7 days, the EBs were transferred to Collagen Type I-containing medium (1.25 mg/mL, catalog #354249; BD Biosciences, Bedford, MA) at a concentration of 150–200 EBs per p30 Petri dish, according to the protocol described by Feraud et al. [44]. EBs were dosed with vascular endothelial growth factor (rhVEGF, catalog #293-VE; R&D Systems, Minneapolis, MN) at a concentration of 50 ng/mL, incubated for 72 h, and scored for sprouting.
Statistical analysis
All P-values were calculated using Student's t-test analysis.
Results
Etv2 and Sox7 expression patterns in EBs
Transcriptome analysis performed on E8.5 Etv2-mutant embryos compared with wild-type littermates revealed decreased expression of Sox-F family transcription factors in the Etv2 mutant embryo [45]. To further assess the temporal relationship between Etv2 and Sox7, gene expression patterns were followed in developing EBs, beginning on day 2 until day 6. Etv2 mRNA expression had a narrow window of induction peaking at day 3.5 followed by rapid downregulation to baseline levels by EB day 4.5 (Fig. 1A). Sox7 expression was initiated at day 3.5 with a gradual increase through EB day 6 (Fig. 1B). The onset of Sox7 mRNA after Etv2 upregulation corroborates the hypothesis that Sox7 is a potential direct downstream target of ETV2. In addition, analysis of EBs from ES cells generated from Etv2-mutant and WT-littermate blastocysts showed complete absence of Sox7 mRNA in the Etv2-mutant EBs (Fig. 1C); while the pattern of Sox7 mRNA expression in EB differentiation assays of the WT littermate cells was similar to the Ainv18 ES cell line (Fig. 1B), and it simply shifted 0.5 days due to a variation in embryonic staging and ES cell derivation. Although both Etv2 and Sox7 have previously been described in the EPC population, to confirm that Etv2 and Sox7 were present in the same cell, single-cell qRT-PCR was performed using FLK1+/CDH5+ EPCs derived from EBs harvested at day 5. This demonstrated that Sox7 and Etv2 were co-expressed in all 40 cells assayed (Fig. 1D). Finally, a dox-inducible ETV2 over-expressing ES cell line was utilized to show that over-expression of ETV2 in EBs before the appearance of endogenous ETV2 resulted in significant induction of Sox7 mRNA compared with the control EBs (Fig. 1E). Over-expression of Etv2 in engineered ES cells, after induction with dox, was observed at the mRNA level (data not shown) and at the protein level by western blot analysis (Fig. 1E).

Etv2 and Sox7 expression patterns in embryonic bodies (EBs).
ETV2 binds and activates the Sox7 promoter
The 10 kb upstream regions of the mouse, human, and rat Sox7 gene were aligned, and conserved regions between species were identified. In addition to the homology of the proximal promoter, three additional upstream regions had conserved sequences: region I (−5272 to −5512 bp from the transcriptional start site), region II (−3616 to −3835), and region III (−910 to −1240). Putative EBEs corresponding to the sequence G/C C/G C/a G A G/a T/c [46] were identified in all three regions, (Fig. 2A). All three regions of the Sox7 upstream region were tested for ETV2 binding using the chromatin immuno-precipitation (ChIP) technique from cells expressing HA epitope-tagged ETV2. Enrichment of each region, after immunoprecipitation with anti-HA antibody, was determined by qRT-PCR and compared with a Gapdh control (Fig. 2B). The fragment containing three putative EBEs, from −3616 to −3835 upstream of the transcriptional start site, was significantly enriched 4.4±0.6-fold, suggesting that ETV2 binds to this region of the Sox7 promoter. EMSA revealed that ETV2 binds to two of the three putative EBEs identified in region II of the Sox7 promoter (Fig. 2C). This binding could be supershifted with an HA antibody that recognized the ETV2-HA fusion construct, and it could be competed with WT synthetic oligonucleotide but not with mutated synthetic oligonucleotide (Fig. 2C). The ability of this regulatory fragment of Sox7 to confer ETV2-dependent transcriptional activation was tested by fusing the 241 bp fragment containing the three EBE sequences (−4002 bp to −3761 bp) to the luciferase reporter and performing transcriptional assays in C2C12 myoblasts. As shown in Fig. 2D, co-transfection of the reporter plasmid with increasing amounts of ETV2-expressing vector resulted in a dose-dependent activation of luciferase activity (4.38±0.13-fold, 4.92±0.50-fold, and 6.63±0.26-fold (P<0.05, n=3)) compared with the control (no ETV2-expressing plasmid). Moreover, the reporter plasmid in which all three EBE sequences were mutated had no activation, even with maximal amounts of ETV2 (1.03±0.07-fold change; P<0.05, n=3; Fig. 2D). Transcriptional assays using expression plasmids for ETS1 and ERG (ETS-family proteins expressed during endothelial development) showed that the 241 bp region containing the EBE sequences specifically responded to ETV2 but not to other closely related proteins (Fig. 2E). In combination, these data show that ETV2 binds two EBEs of the Sox7 promoter (EBE-1: −3936 to −3926 and EBE-3: −3778 to −3768), and it confers ETV2-dependent activation. Based on these data, we conclude that Sox7 is a direct downstream target of ETV2.
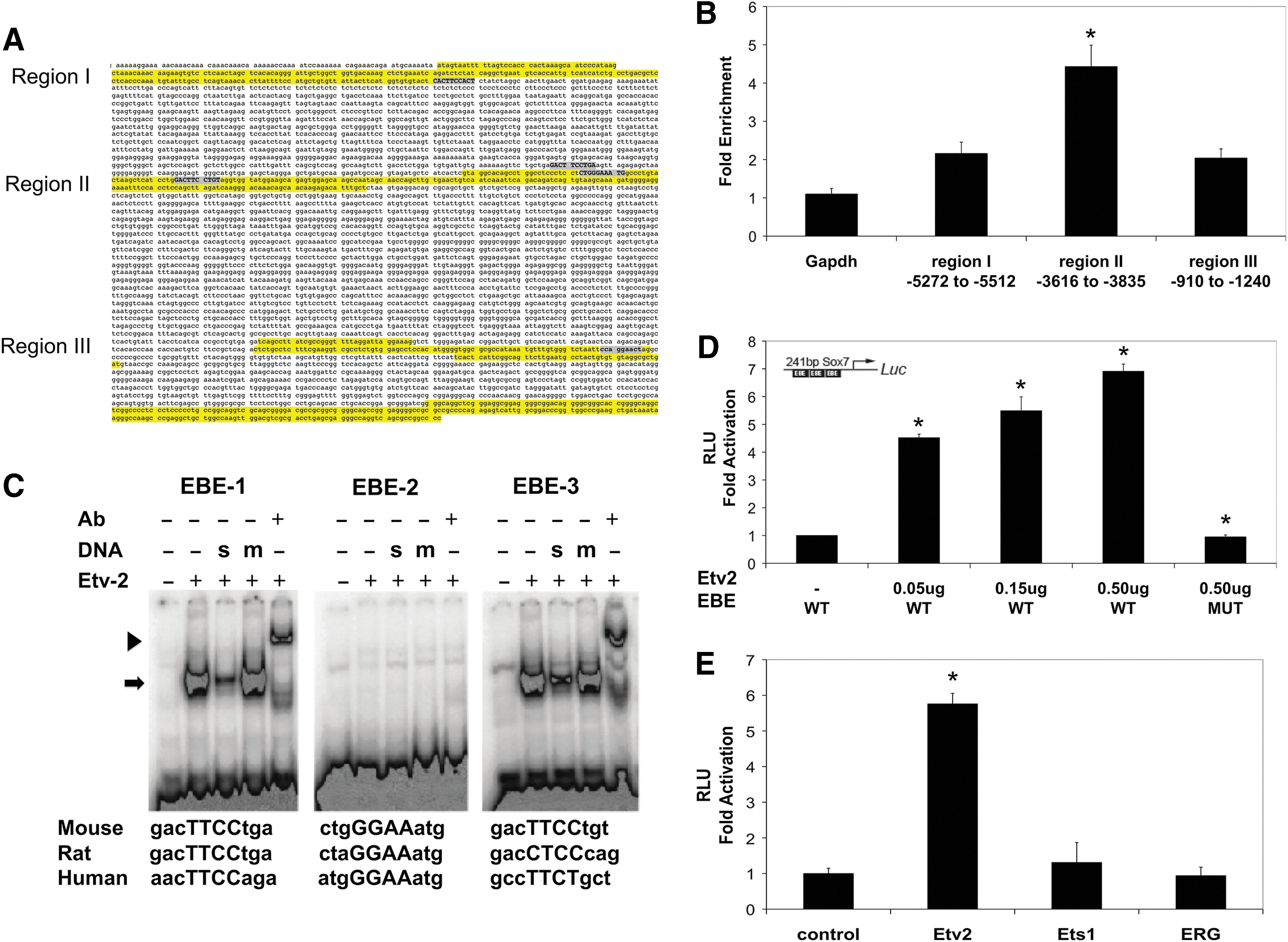
ETV2 binds and activates the Sox7 promoter.
Sox7 increases EPCs in EBs
Etv2 is essential for the development of the endothelial lineage [14]; we, thus, hypothesized that ETV2 functions through a SOX7-dependent pathway in endothelial development. The effect of SOX7 over-expression on the production of FLK1+/CDH5+ EPCs was assessed in EBs harvested at day 6 [42,45,47]. Induction of SOX7-MYC for 72 h (day 3–6, Fig. 3A) significantly increased FLK1+/CDH5+ EPCs as compared with the control (12.2%±2.7% vs. 5.0%±2.8%, P<0.05, n=9; Fig. 3B); however, the fold increase was consistently lower compared with the changes seen with ETV2 over-expression (twofold vs. threefold) (Supplementary Fig. S1; Supplementary Data are available online at
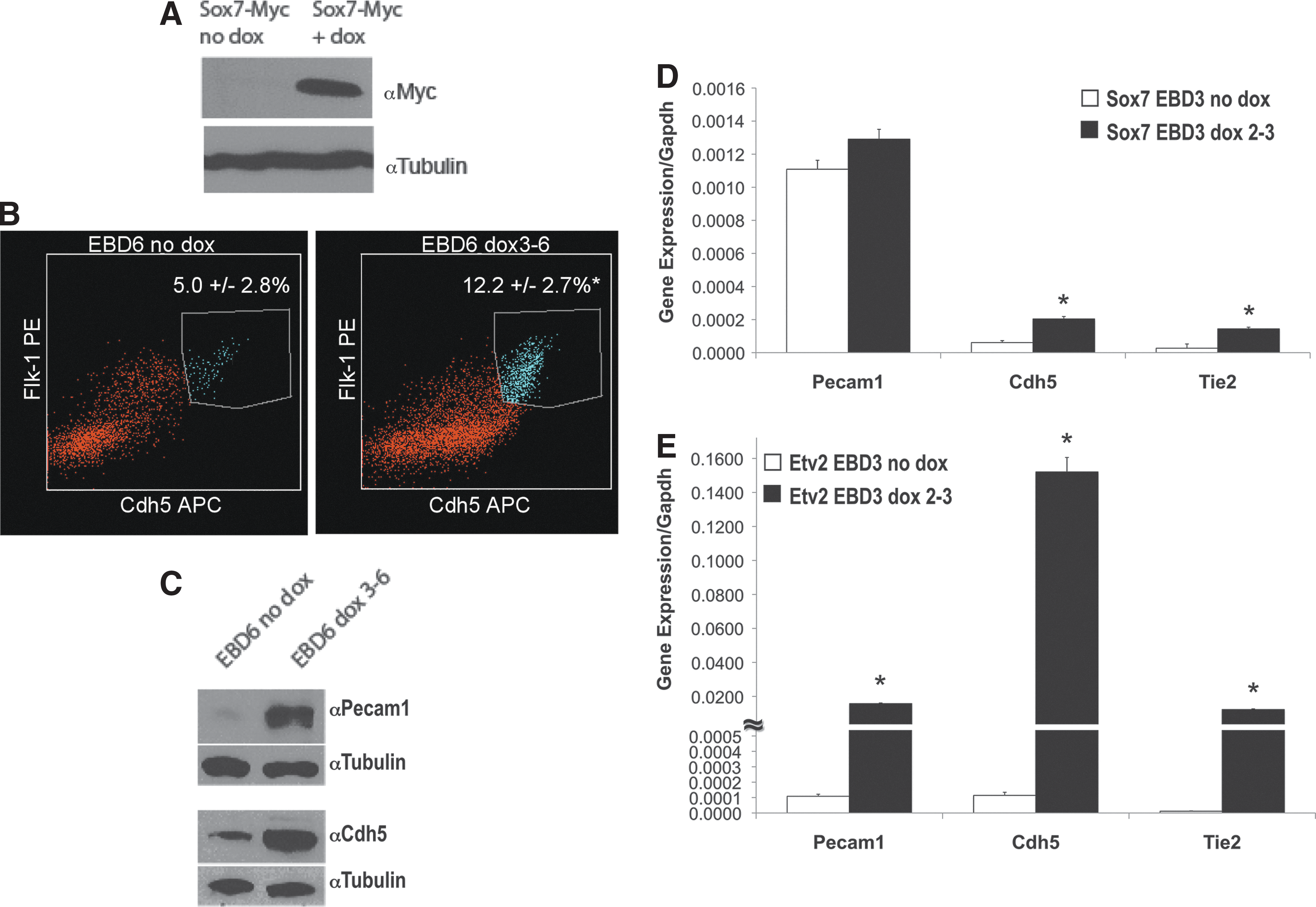
SOX7 over-expression results in increased levels of endothelial progenitors cells (EPCs) and endothelial gene expression.
The role of SOX7 in the ETV2 pathway was further investigated by engineering cells in which ETV2 could be over-expressed in the presence of Sox7 inhibition. ShRNA technology was combined with the dox-inducible over-expressing ES-system to obtain such an mES cell line, and the ability to produce EPCs in EBs was assessed. EBs were made using hanging drop technology and cultured in suspension for 6 days. ETV2 was over-expressed with dox from day 3 to 6, and shRNA for Sox7 was constitutively expressed. To show that knockdown by Sox7 shRNA was effective, protein levels were analyzed by western blot. SOX7 and CDH5 a downstream target of Sox7 [37], protein levels were induced by dox in control cells (in the absence of shRNA), and clearly decreased in the presence of Sox7 shRNA. However, the closely related Sox-F family members SOX17 and SOX18 did not show changes in protein levels in the presence of Sox7 shRNA, indicating knockdown specificity of the Sox7 shRNA to Sox7 (Supplementary Fig. S2). Having validated our system, EBs were collected on day 6 and FLK1+/CDH5+ cells were quantified. ETV2 over-expressing EBs containing control pLKO.1 vector had percentages of FLK1+/CDH5+ cells similar to those previously observed in ETV2 over-expressing cells not infected with lentivirus with a threefold increase in double-positive cells on over-expression (no dox: 5.5%±1.4%, dox: 14.2%±3.3%, P<0.05, n=6; Fig. 4B). In the presence of Sox7 shRNA, the ETV2 over-expressing cells had a significantly lower percentage of FLK1+/CDH5+ cells when compared with ETV2 over-expressing cells containing the control pLKO.1 vector (6.4%±1.4% vs. 14.2%±3.3%, P<0.05, n=6; Fig. 4B). Sox7 inhibition significantly decreased the level of FLK1+/CDH5+ in un-induced cells (5.5%±1.4% vs. 3.5%±2.0%, P<0.05, n=10, Fig. 4B). To further examine the effects on mesodermal commitment, we analyzed FLK1+ cells in EBs collected at day 4 (Table 1). The data indicate that while ETV2 over-expression increases the FLK1+ cells at day 4 (66.5%±3.5% vs. 90.5%±5.5%, P<0.05, n=4), these cells were significantly decreased in SOX7 over-expressing cells (73.1%±6.1% vs. 60.2%±5.5%, P<0.05, n=4). Knockdown of Sox7 generated some variability in the responses and resulted in statistically insignificant differences in the day 4 FLK1+ population.

Sox7 shRNA decreases endothelial progenitor cell production and angiogenic sprouting in ETV2 over-expressing EBs.
P<0.05, n=4.
P<0.05, n=4.
ns=not significant.
Functional role of Sox7 in the ETV2 pathway
Having established an ETV2-Sox7 pathway in progenitor cells, an angiogenic assay was performed to further define the functional role of SOX7 in the ETV2 pathway in vascular development. EBs were prepared using the hanging drop technique and mES with inducible Etv2 with or without Sox7 shRNA. EBs were cultured in suspension for 7 days, before transfer to secondary collagen culture for 72 h in medium containing rhVEGF. Over-expression of ETV2 significantly increased angiogenic sprouting in EBs from 54.6%±8.4% to 80.4%±7.2% (P<0.05, n=4; Fig. 4C, D), and these values were consistent with previously observed values [48]. Sox7 knockdown significantly decreased the number of sprouting EBs to 27.2%±10.8% and 40.9%±14.3% in EBs with or without ETV2 over-expression, respectively (P<0.05, n=4; Fig. 4C), further supporting the hypothesis that ETV2 regulates transcriptional networks which include Sox7 to carry out its functions during vascular development.
Discussion
It was previously demonstrated that Etv2 is expressed early during embryogenesis in the endocardial/endothelial lineage and that Etv2-deficient embryos are lethal, lacking endocardial/endothelial and hematopoietic lineages [14,21,23,45]. In the zebrafish model system, Etv2 has also been shown to be important for vascular development, initially for EPC specification, and later in hemogenic endothelial cells [49,50]. Genetic fate mapping analysis utilizing an Etv2-cre transgenic mouse model confirmed that ETV2-expressing cells give rise to both hematopoietic and endothelial lineages [45].
The Sox-F family members (Sox7, Sox17, and Sox18) have been shown to be important in the formation of the endodermal lineages, regulation of hematopoietic cells, and cardiovascular development [24,27,31,32,35,36,51]. In the endothelial system, recent studies have implicated Sox7 as an important factor in hematopoietic specification in addition to its importance in endothelial precursors [52]. Zebrafish studies also demonstrated that Sox7 and Sox18 were expressed predominately in the vasculature, and reduced expression of both genes resulted in defective vasculature development [25,36,53]. Gandillet et al. have shown that knockdown of Sox7 in EB differentiation assays severely affected both endothelial and hematopoietic precursors [54]. Recently, it was shown that continued enforced expression of SOX7 in hemangioblast-derived blast colonies resulted in sustained expression of endothelial markers while impairing hematopoietic differentiation [37]. This same study implicated Cdh5, a gene known to be downstream of ETV2, as a direct downstream target of SOX7 [21,55]. Although there is significant overlap of Etv2 and Sox7 function in early endothelial and hematopoietic development, as well as apparent overlap of transcriptional pathways, a direct linkage between Etv2 and Sox7 has not previously been described.
Given that the Sox-F family of genes is known to be expressed in the endothelial lineage, and that previous studies have established that Etv2 is required for the establishment of hemangiogenic mesoderm and the induction of early endothelial progenitor cells [15,20 –23], our finding of the downregulation of the Sox-F family of genes in the absence of Etv2 could have been predicted (in situ hybridization data not shown). However, it was quite striking that the expression of Sox7, but not all Sox-F family members, was completely abolished in the absence of Etv2. Single-cell expression analysis confirmed that Etv2 and Sox7 are co-expressed in EPCs. These findings supported our initial data revealing that Sox7 could be a direct downstream target of ETV2, and that the Sox-F family members have specific roles in endothelial development. The upstream regulation of Sox7 is poorly understood. Research in cancer biology has implicated both p38MAPK and β-catenin/Wnt pathways as regulators of Sox7 expression [56]. However, the developmental regulation of Sox7 has not been previously described. Our findings from rigorous promoter analysis and protein-DNA interaction, gel shift, and transcriptional assays confirm that ETV2 directly activates Sox7. It is important to note, however, that Sox7 alone does not seem to induce the early endothelial lineage as efficiently as Etv2. This is not surprising given the critical role of Etv2. It is likely that Etv2 also contributes to endothelial induction through Sox7-independent pathways. However, a functional role of this newly described ETV2-Sox7 activation was also substantiated. Reduction of Sox7 in the presence of ETV2 activation significantly reduced both the percentage of endothelial progenitors and the angiogenic potential of embryoid bodies. Furthermore, perturbation of Sox7 expression at even basal levels of Etv2 expression also significantly reduced the angiogenic capacity of embryoid bodies, further confirming that Etv2 regulates endothelial development via transcriptional pathways which include Sox7. These results from our current studies provide further insights into the transcriptional pathways of the endothelial lineage.
Initial work focused on the role of the Sox-F family members in cardiogenesis and angiogenesis and demonstrated that the family largely has redundant roles in these systems [5,24 –28,32,34,35]. Although there are areas of overlap in the family member's functional roles, studies have begun to focus on the identification of the unique aspects of the developmental pathways of each individual member. Recent work has shown that each member also possesses specific functions as illustrated by the importance of Sox18 in lymphangiogenesis, the role of Sox17 in fetal hematopoietic stem cell proliferation and definitive endoderm formation, and the requirement of Sox7 for primitive endoderm formation [57]. In the present study, we also highlight the individual role of Sox7 in endothelial development. Additional studies will be necessary to completely define the comprehensive role of each Sox-F family member in endothelial development.
In this article, a previously unknown direct link between ETV2 and Sox7 is established. We provide evidence that Sox7 is one of the targets of ETV2 necessary to regulate the endothelial pathway, resulting in endothelial progenitor cell upregulation and increased angiogenic sprouting. Since the role of Sox7 in endothelial development is generating intense interest, future studies that investigate downstream targets of Sox7 and their role in endothelial development will further illuminate the importance of this pathway.
Footnotes
Acknowledgments
The authors thank Michael Kyba for providing the Tet-on inducible Ainv18 ES cell line. They also acknowledge Michelina Iacovino and Alessandro Magli for critical discussions and technical assistance. They recognize the University of Minnesota Genomic Center for the single-cell capture qRT-PCR work. This work was supported by the National Institutes of Health (K08 HL102157-01).
Part of this work was presented at the American Heart Association (AHA) 2012 Scientific Sessions, Los Angeles, CA.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.