
Abstract
Pancreatic cancer (PC) is one of the most deadly cancers. The higher mortality is in part due to treatment resistance and early onset of metastasis. The existence of cancer-stem-like cells (CSLCs) has been widely accepted to be responsible for tumor aggressiveness in PC. Emerging evidence suggests that CSLCs have the capacity for increased cell growth, cell migration/invasion, metastasis, and treatment resistance, which leads to poor clinical outcome. However, the molecular role of CSLCs in tumor development and progression is poorly understood. Therefore, mechanistic understanding, and targeted killing of CSLCs may provide a newer therapeutic strategy for the treatment of PC. It has been well accepted that microRNAs (miRNAs) play critical roles during tumor development and progression through deregulation of multiple genes. Moreover, deregulated expression of miRNAs may also play a key role in the regulation of CSLC characteristics and functions. Here we show that isolated CD44+/CD133+/EpCAM+ cells (triple-marker-positive cells) from human PC cell lines, MiaPaCa-2 and L3.6pl cells, display aggressive characteristics, such as increased cell growth, clonogenicity, cell migration, and self-renewal capacity, which is consistent with overexpression of CSLC signatures/markers. We also found deregulated expression of over 400 miRNAs, including let-7, miR-30, miR-125b, and miR-335, in CSLCs. As a proof-of-concept, knockdown of miR-125b resulted in the inhibition of tumor cell aggressiveness of CSLCs (triple-marker-positive cells), consistent with the downregulation of CD44, EpCAM, EZH2, and snail. These results clearly suggest the importance of miRNAs in the regulation of CSLC characteristics, and may serve as novel targets for therapy.
Introduction
P
In this study, we isolated and characterized CD44+/CD133+/EpCAM+ cells (triple-positive cells) from human PC cell lines, MiaPaCa-2 and L3.6pl cells, which showed CSLC characteristics. These CSLCs (triple-positive cells) display aggressive phenotypes and functions, such as increased cell growth, clonogenicity, cell migration, and self-renewal capacity, all of which are consistent with overexpression of CSLC signatures/markers, compared with parental cells and triple-marker-negative cells. Microarray expression profile data showed differential expression of more than 400 miRNAs, including let-7, miR-30, miR-125b, and miR-335, in CSLCs (triple-marker-positive cells). As a proof-of-concept, the knockdown of miR-125b by its siRNA inhibitor resulted in the inhibition of aggressive phenotypes and functions of these triple-marker-positive cells, which was consistent with the downregulation of CD44, EpCAM, EZH2, and Snail. These results suggest that targeting miRNAs could become a novel strategy for the killing of CSLCs, and thus such a strategy would be useful for improving the treatment outcome of patients diagnosed with PC.
Materials and Methods
Cell lines and culture conditions
The CSLCs (triple-marker-positive; CD44+/CD133+/EpCAM+) and triple-marker-negative (CD44−/CD133−/EpCAM−) cells were isolated from human PC, MiaPaCa-2 and L3.6pl cells, by fluorescence-activated cell sorting (FACS) technique and maintained in 5% fetal bovine serum (FBS)–Dulbecco's modified Eagle's medium (DMEM) medium at 37°C in a standard culture condition. The CSLCs established from four CD44+/CD133+/EpCAM+ cells were maintained in sphere formation medium (FBS-free, 1:1 DMEM:F-12K media plus B27 and N2 supplements; Invitrogen) to maintain its undifferentiated status. CD44+, CD133+, and EpCAM+ are known as cancer stem cell (CSC) surface markers [5 –7], and these triple-marker-positive cells are reminiscent of CSLCs.
Sphere formation assay
CSC (CSLCs) self-renewal capacity was evaluated by sphere formation assay, as described previously [8]. Briefly, 1,000 single-suspended cells were plated on ultra-low-attachment wells of Costar six-well plates (Corning Incorporated) in 2 mL of sphere formation medium. After 7 days of incubation, the sphere cells were collected by centrifugation (300 g for 5 min), and the number of spheres (pancreatospheres) was counted under a microscope.
Cell survival assay
The cell survival was evaluated by MTT assay. About 5,000 cells per well were plated in a 96-well plate. After 3 days of incubation, the cells were harvested for the standard MTT assay, as described in the previous publications [9,10].
Colony formation assay
Clonogenicity was evaluated by colony formation assay, as described previously [8,10]. Briefly, 1,000 single viable cells were plated in 10 mL of 5% FBS–DMEM medium in 100-mm Petri dishes. The cells were then incubated for 14 days at 37°C in a tissue culture incubator. Colonies were stained with 2% crystal violet, washed with water, and counted.
Wound healing assay
Cell migration capacity was evaluated by wound healing assay, as described previously [8,10]. Briefly, the cells were seeded in six-well plates in 5% FBS–DMEM medium. When cells reached 95% confluency, they were scratched with a 200 μL of pipette tip. The wound healing capacity was evaluated and photographed with a camera-equipped inverted microscope after 20 and 48 h of incubation.
Cell cycle analysis
Propidium iodide (PI) staining and flow cytometry assay was performed to examine cell cycle parameter of cells in culture. After being washed, the cells were fixed with 75% ethanol at −20°C overnight. The fixed cells were washed and reacted with PI-RNase A solution (Invitrogen) at 4°C for 3 h, followed by flow cytometric analysis.
Western blot analysis
The relative levels of p21 and p27 proteins were measured by western blot analysis, which has been described previously [9]. The antibodies against p21 and 27 were obtained from Cell Signaling Technology.
Quantitative real-time RT-PCR of miRNAs and mRNAs
Real-time RT-PCR assay was performed to measure miRNAs and mRNAs, as described previously [8,10]. Reverse transcription (RT) reactions of total RNAs were then performed using TaqMan miRNA, and cDNA Assay kits (Applied Biosystems), respectively, following the manufacturer's instructions. Polymerase chain reactions (PCRs) of miRNAs and mRNAs were performed using TaqMan PCR Master Mix and CYBR Green PCR Master Mix kits (Applied Biosystems), respectively. The miRNA primers and probes were purchased from Applied Biosystems. The primers of cDNAs were synthesized from Invitrogen. The sequences of the primers are listed in the Supplementary Table S1 (Supplementary Data are available online at
Immunostaining and confocal imaging microscopy
After the cells reached 50%–70% confluent that were cultured in four-chamber culture glass slides in 5% FBS–DMEM medium, the cells were then washed, fixed with 4% formaldehyde solution, and permeabilized with 0.05% Tween-20 solution. Antibodies against CD44, EpCAM, Notch-1, EZH2, and Snail (Cell Signaling) were used for immunostaining, following the manufacturer's protocol. DAPI was used to stain the nucleus of the cells as a control. The antibody-labeled cells were examined and photographed under a confocal imaging microscope (100× magnification; EVOS Imaging Systems).
miRNA microarray assay and pathway/network enrichment analysis
Purified total RNAs were isolated by using mirVana miRNA Isolation kit (Ambion, Inc.), following the manufacturer's instruction. The miRNA microarray assay was performed by LC Sciences. The miRNA expression profiling analysis was performed by miRBase version 16 (LC Sciences). The values of log [2] of each miRNA from data comparisons were used for the fold-change levels. Data were normalized by using selected housekeeping genes. System network analysis was performed by using the web-based bioinformatics tool Ingenuity Pathway Analysis software (Ingenuity Systems) for predicting functional network.
Transfections of anti-miR125b siRNA inhibitor
Anti-miR-125b siRNA (Applied Biosystems) was transfected into the CSLCs (triple-marker-positive cells) using DharmaFECT transfection reagent (Thermo Scientific) following the manufacturer's instructions, as described previously [8,10]. The scramble siRNA was used as the negative control. The transfected cells were used for sphere formation, wound healing, colony formation, real-time RT-PCR, or confocal imaging microscopy assays, as described previously.
Statistical analysis
All the data were prepared and presented as mean and standard deviation by using GraphPad Prism and Excel software, as described previously [8,10]. Student's t-test was conducted to test a significant difference between two groups (P≤0.05 was considered as statistically significant).
Results
Isolation of CSLCs (triple-marker-positive cells) and triple-marker-negative cells of MiaPaCa-2 cells
The CSLCs (triple-marker-positive cells) were isolated, established from four of CD44+/CD133+/EpCAM+ cells (four absolute number of the cells) of human PC MiaPaCa-2 cells by using FACS technique (Fig. 1A, B). The triple-marker-negative cells were isolated and established from 78,000 (absolute number) (CD44−/CD133−/EpCAM− cells) of MiaPaCa-2 cells by FACS technique. Flow cytometry assay shows that only 0.3% was triple-marker positive (CD44+/CD133+/EpCAM+) in MiaPaCa-2 cells (the data not shown). The triple-marker-negative cells were <0.01% in CD44+/CD133+/EpCAM+ cells (the data not shown), suggesting minor contamination. However, CSLCs (triple-marker-positive cells) consist, at least, 60% CD44+/CD133+/EpCAM+ cells (the data not shown). After 4 weeks of incubation, four of isolated CSLCs (triple-marker-positive cells) in the sphere forming assay survived and grew to become more than one million cells (Fig. 1A). However, four (absolute number) of cells, either MiaPaCa-2 or isolated triple-marker-negative cells, failed to survive, and thus they did not grow in the sphere formation medium (Fig. 1A).

Characteristics of CSC triple-positive cells.
Self-renewal capacity of CSLCs (triple-positive cells)
The sphere formation assay was performed to evaluate the self-renewal capacity of CSLCs. The results show that the CSLCs (triple-marker-positive cells) of MiaPaCa-2 cells had significantly higher number of spheres (pancreatospheres) after 7 days of incubation in the sphere formation medium, compared with either parental MiaPaCa-2 cells or triple-marker-negative cells (Fig. 1B). Similarly, the CSLCs (triple-marker-positive cells) of L3.6pl cells had significantly higher number of pancreatospheres, compared with the triple-marker-negative L3.6pl cells (Supplementary Fig. S1A). These findings suggest that the CSLCs (triple-marker-positive cells) have remarkably higher self-renewal capacity.
Cell survival capacity of CSLCs (triple-marker-positive cells)
MTT assay was performed to evaluate cell survival capacity of CSLCs (triple-marker-positive cells) in 5% FBS–DMEM medium. The results showed that CSLCs (triple-marker-positive cells) of MiaPaCa-2 cells had increased cell growth capacity after 3 days of incubation, compared with triple-marker-negative cells (P<0.05; n=3) (Fig. 1C).
Cell cycle parameters of CSLCs (triple-marker-positive cells)
PI staining and flow cytometry assay was performed to evaluate the cell cycle parameters of CSLCs (triple-marker-positive cells). The results showed that CSLCs (triple-marker-positive cells) had significantly decreased G0-G1 phase, and had increased G1-G2 phase and S phase, compared with parental MiaPaCa-2 cells and triple-marker-negative cells (Table 1). These findings suggest that CSLCs (triple-marker-positive cells) have different pattern of cell cycle parameters. However, there was no significant difference in cell cycle parameters between the parental MiaPaCa-2 cells and triple-marker-negative cells (Table 1). According to a previous report, Sox2-overexpressing L3.6pl cells had a long S-phase [11], which is different from our observation in this study. This difference of S-phase may be due to different subgroups of CSLCs used in our studies. We used the isolated CSC triple-marker-positive cells (CD44/CD133/EpCAM) in this study, instead of single-marker- (Sox2) positive cells [11].
Cell cycles were analyzed by propidium iodide staining and flow cytometry assay. Mean±SD represents mean and standard deviations. n=3 represents 3 independent experiments.
P≤0.05, compared with MiaPaCa-2 cells and triple-marker-negative cells.
CSLCs, cancer-stem-like cells.
The relative levels of p21 and p27 in CSLCs
We also examined the relative levels of CDK inhibitors p21 and p27 in the CSLCs (triple-marker-positive cells) of MiaPaCa-2 cells by western blot analysis. The results show that the CSLCs have significantly higher p21 protein levels (Fig. 1D), which is similar to the previous findings in Sox2-overexpressing L.36pl cells reported by other investigators [11].
Clonogenicity of CSLCs (triple-positive cells)
The colony formation assay was performed to evaluate the clonogenic potential of cells. The results show that CSLCs (triple-positive cells) had a significantly increased capacity of colony formation in 5% FBS–DMEM medium, compared with either parental MiaPaCa-2 cells or triple-marker-negative cells (Fig. 1E) (P<0.05; n=3). There was no significant difference in colony formation between parental MiaPaCa-2 cells and triple-marker-negative cells (Fig. 1E). These findings suggest that CSLCs (triple-marker-positive cells) display a significantly higher clonogenicity.
Cell migration of CSLCs (triple-marker-positive cells)
The wound healing assay was performed to evaluate the cell migration capacity of CSLCs (triple-marker-positive cells). The results show that CSLCs (triple-marker-positive cells) have remarkably higher wound healing capacity after 20 and 48 h of incubation in 5% FBS–DMEM medium, compared with either parental MiaPaCa-2 cells or triple-marker-negative cells (Fig. 1F). There was no substantial difference in the wound healing capacity between the parental MiaPaCa-2 cells and triple-marker-negative cells. We also examined the effect of the conditioned media obtained from the triple-marker-negative cells on cell migration in the CSLCs (triple-marker-positive cells). The results show that the conditioned media obtained from the triple-marker-negative cells decreased the wound healing capacity of the CSLCs (triple-marker-positive cells) of MiaPaCa-2 (Fig. 1G) and L3.6pl cells (Supplementary Fig. S1B), compared with the control cells. These data suggest that CSLCs (triple-marker-positive cells) have a higher cell migration capacity.
CSLCs (triple-marker-positive cells) display high expression of CSC-associated genes
Real-time RT-PCR assay showed that CSLCs (triple-marker-positive cells) of MiaPaCa-2 cells had increased mRNA levels of CSC signatures and other markers, such as CD44, CD133, EpCAM, Oct4, Nanog, Sox2, EZH2 (enhancer of zeste homolog 2), Akt, Notch-1, and Snail, compared with triple-marker-negative cells of MiaPaCa-2 cells (Fig. 2A). Confocal imaging microscopy study also showed that CSLCs (triple-marker-positive cells) had increased protein expression of EpCAM and EZH2, compared with parental MiaPaCa-2 and triple-marker-negative cells (Fig. 2B). CD44, CD133, and EpCAM are well-known stem cell surface markers [5 –7]. Nanog and Sox2 are also known CSC marker genes [12,13]. EZH2, Akt, Notch-1, and Snail are reportedly involved in tumor development, and have been considered as potential CSC mediators, and thus they are also considered as CSC markers [5,14 –16]. Similarly, CSLCs of L3.6pl cells had the high expression of CSC markers EpCAM, EZH2, and Snail, compared with the triple-marker-negative cells (Supplementary Figs. S1C and S2A). Our data suggest that CSLCs (triple-marker-positive cells) have higher expression of CSC signatures and associated markers.

The expression of CSC signatures/mediators in CSLCs (triple-positive cells) of MiaPaCa-2 cells.
Differential expression profiling of miRNAs of CSLCs (triple-marker-positive cells)
Microarray analysis was performed to examine differential expression of miRNAs of CSLCs (triple-marker-positive cells), compared with either the parental MiaPaCa-2 cells or triple-marker-negative cells. We found 438 miRNAs that were differentially expressed in CSLCs (triple-marker positive) versus triple-marker-negative cells (Supplementary Table S2). Among these miRNAs, 191 miRNAs were upregulated and 247 miRNAs were downregulated in CSLCs (triple-marker-positive cells), compared with triple-marker-negative cells. The heatmap of CSLCs (triple-marker positive) versus triple-marker-negative cells is presented in Supplementary Fig. S3. Moreover, we found 486 miRNAs that were differentially expressed between CSLCs (triple-marker positive) and the parental MiaPaCa-2 cells (Supplementary Table S2). Among those miRNAs, 243 miRNAs were upregulated and 243 miRNAs were downregulated. The heatmap of parental MiaPaCa-2 cells versus CSLCs (triple-marker-positive cells) is shown in Supplementary Fig. S4. However, there were only 180 differentially expressed miRNAs between triple-marker-negative cells and the parental MiaPaCa-2 cells (Supplementary Table S2). Among those miRNAs, 108 miRNAs were upregulated and 72 miRNAs were downregulated.
RT-PCR assay of selected miRNAs
We analyzed the miRNA expression levels of six miRNAs because they were either upregulated or downregulated in CSLCs (triple-marker-positive cells) compared with parental MiaPaCa-2 and triple-marker-negative cells as described in Supplementary Table S3, and validated their comparative expression by RT-PCR. The results show that let-7f, let-7i, miR-30a, and miR-30c were downregulated and miR-125b and miR-335 were upregulated in CSLCs (triple-marker-positive cells), compared with MiaPaCa-2 and triple-marker-negative cells (Fig. 3). All these miRNAs have been reportedly found to be associated with tumor development [17 –25]. Similarly, the CSLCs (triple-marker-positive cells) of L3.6pl cells showed similar results like MiaPaCa-2 cells with respect to these miRNAs (Supplementary Fig. S2B).

Validation of the expression of selected miRNAs in MiaPaCa-2, CSLCs (triple positive), and triple-negative cells by real-time RT-PCR. RNU48 was used as the control. The bars represent standard deviation. *P≤0.05 (n=3), CSLCs (triple-positive cells) compared with triple-negative cells. miRNAs, microRNAs.
Pathway network analysis of miRNAs in CSLCs (triple-marker-positive cells)
We further performed ingenuity modeling analysis of the miRNA profiling to better understand the pathways that are involved in conjunction with their respective target genes. The algorithms of networks of selected miRNAs that are involved were generated based on their pathway connectivity. The results show that the pathway-enrichment analyses of CSLCs (triple-marker positive) versus triple-marker-negative cells are similar to that of CSLCs (triple-marker positive) versus the parental MiaPaCa-2 cells (Fig. 4A). There were 10 biological functional groups that showed such connectivity, including cancer, gastrointestinal (GI) disease, and genetic disorder (Fig. 4A). These suggest that differential expression of miRNAs in CSLCs (triple-marker-positive cells) is highly associated with the development and progression of tumors, especially GI tumors. The network analysis of selected miRNAs in CSLCs (triple-marker positive) versus triple-marker-negative cells was similar to that of CSLCs (triple-marker positive) versus MiaPaCa-2 cells (Fig. 4B). The network analysis also showed that many miRNAs were regulated through each other either directly or indirectly, which were also regulated by several target genes. We found upregulation of miR-20a, miR-21-3p, miR-23a, miR-34a-5p, miR-193a, miR-294, miR-296, miR-335, miR-432, and miR-1913, as well as the downregulation of let-7a, miR-21-5p, miR-30a,c, miR-92a, miR-130a, miR-330, miR-532, and miR-660 in CSLCs (triple-marker-positive cells), compared with MiaPaCa-2 and triple-marker-negative cells (Fig. 4B). Interestingly, miR-30a,c, miR-193a, miR-294, miR-296, miR-423, miR-532, miR-660, and miR-1913 target insulin signaling, especially miR-34a-5p is a target of insulin via inflammatory cytokines whereas miR-92a is a target of MAP2K1/2 (Fig. 4B).
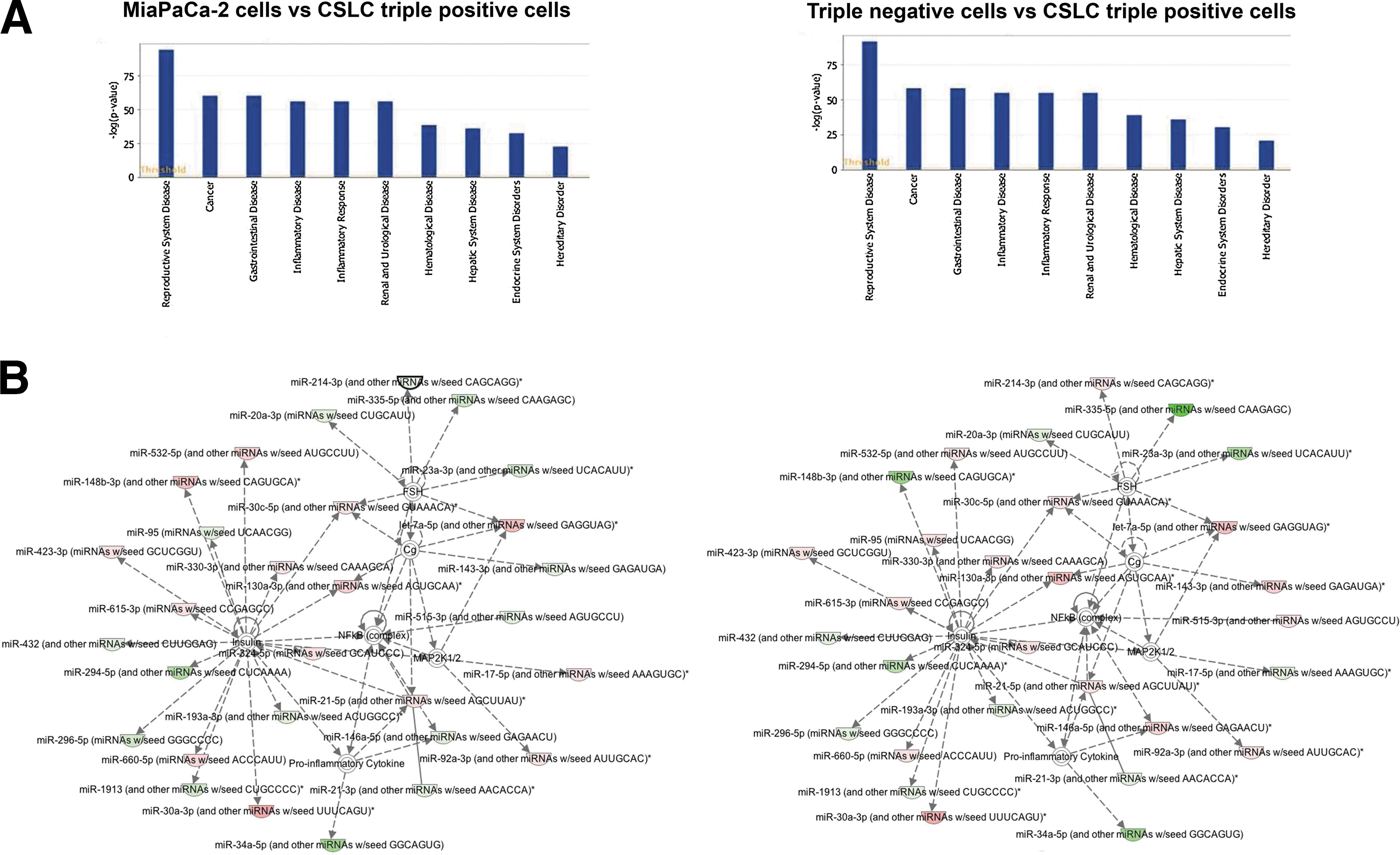
Pathway/network analysis of miRNA expression in MiaPaCa-2 cells, CSLCs (triple-marker-positive cells), and triple-marker-negative cells.
The role of miR-125b in the regulation of CSLC phenotypes and functions
The alterations in the expression of miR-125b have reportedly been found to be associated with poor clinical outcome of several tumors [25]. The data from experimental studies have suggested that miR-125b may play an important role in tumor development [26,27]. In our study, CSLCs (triple-marker-positive cells) showed overexpression of miR-125b, compared with MiaPaCa-2 cells and triple-marker-negative cells, as described earlier (Fig. 3). To further explore the potential role of miR-125b in the regulation of CSC phenotypes and functions, we performed functional loss of expression study of miR-125b by transfecting anti-miR-125b siRNA into the CSLCs (triple-marker-positive cells). The results show that the transfection of anti-miR-125b siRNA significantly decreased the levels of miR-125b in the CSLCs (triple-marker-positive cells) of MiaPaCa-2 cells (Fig. 5A). Knockdown of miR-125b by transfecting anti-miR-125b siRNA also resulted in a significant decrease in the formation of pancreatospheres, clonogenic potential, and wound healing capacity of CSLCs of MiaPaCa-2 cells (Fig. 5B–D), suggesting that downregulation of miR-125b could be clinically important for the treatment of PC. Confocal imaging microscopy study showed decreased protein expression of EpCAM, EZH2, and Snail in CSLCs (triple-marker-positive cells) by knockdown of miR-125b (Fig. 6A). RT-PCR assay showed that knockdown of miR-125b led to decreased mRNA levels of CD44, EpCAM, EZH2, and Snail in the CSLCs (triple-marker-positive cells) of MiaPaCa-2 cells (Fig. 6B). Similarly, anti-miR-125b also decreased the formation of pancreatospheres, and reduced wound healing capacity, consistent with reduction in the expression of CSC markers in CSLCs (triple-marker-positive cells) of L3.6pl cells (Supplementary Figs. S1 and S2A). These findings clearly suggest that miR-125b may play an important role in the regulation of CSLC phenotypes and functions, and as such miR-125b could become a novel therapeutic target in PC.

Effect of anti-miR-125b siRNA transfection on the expression of miR-125b—self-renewal capacity, clonogenicity, and cell migration of CSLCs (triple-marker-positive cells) of MiaPaCa-2 cells.
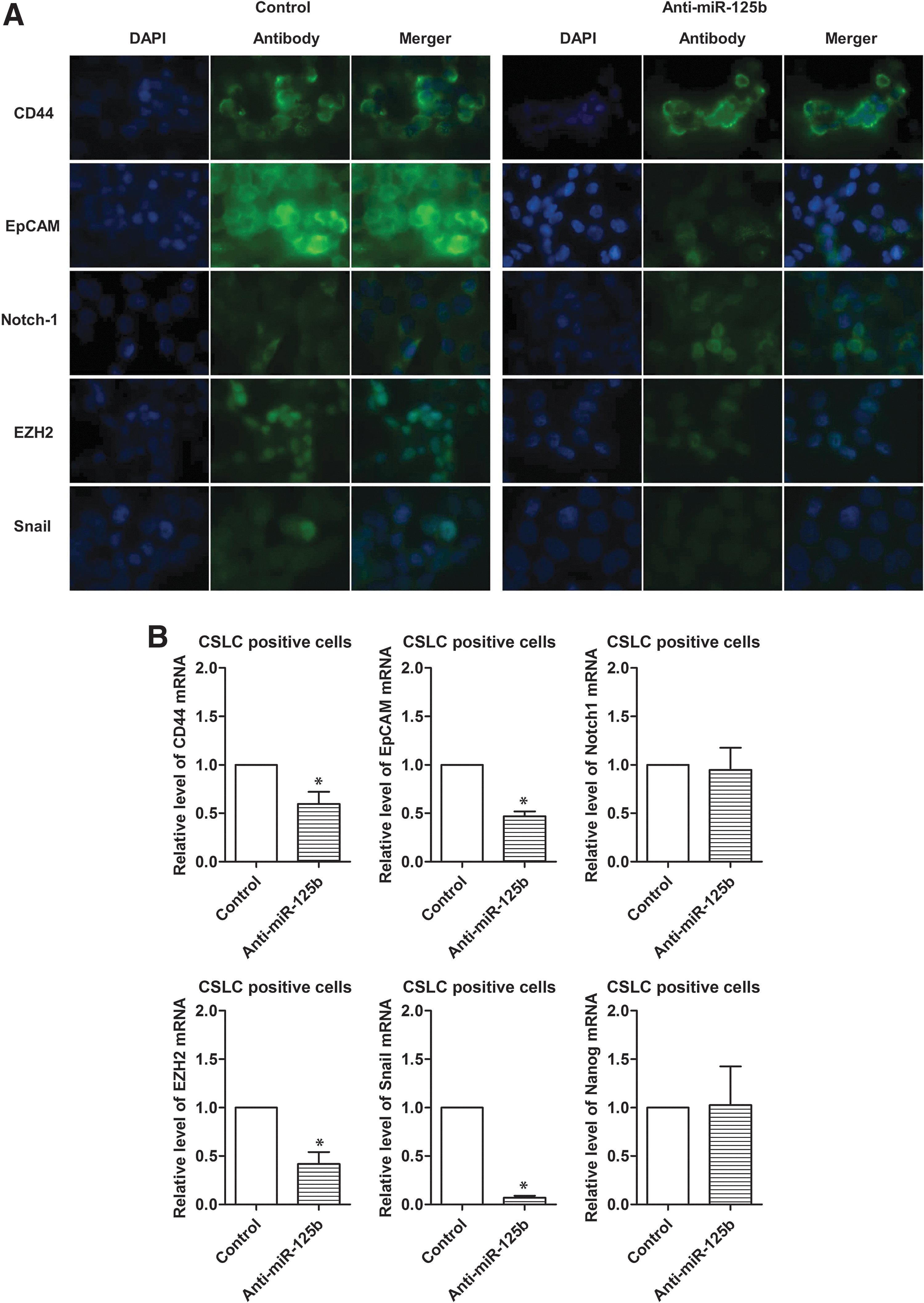
Effect of knockdown of miR-125b on CSC signatures/mediators in CSLCs (triple-marker-positive cells).
Discussion
Emerging evidence suggests that CSCs (or CSLCs) play important role in tumor development and progression because the existence of CSLCs in a tumor has significant clinical implication especially in drug resistance and treatment failure, which result in tumor recurrence. The minority subpopulations of cells (<0.1%) with CSLC characteristics have been identified in many tumors including PC, and the presence and persistence of these cells after conventional therapy are related to poor clinical outcome, such as reduced disease-free survival and increased mortality [6]. Different from tumor-associated non-CSC cells, such rare CSLC subpopulation displays unique features, such as highest capacity of self-renewal, prolonged survival potential, and greatest capacity of differentiation into multiple types of tumor cells, which is in part responsible for tumor cell heterogeneity. Mounting evidence suggests that these features are also involved in increased cell growth/proliferation, cell migration/invasion, treatment resistance, and metastasis, leading to poor clinical outcome in most solid tumors including PC [28]. In the current study, we have demonstrated that CD44+/CD133+/EpCAM+ cells (triple-marker-positive cells) isolated from human PC cell lines MiaPaCa-2 and L3.6pl behave like CSCs (CSLCs). These CSLCs (triple-marker-positive cells) have aggressive phenotypes and functions, such as increased capacity of clonogenicity, cell migration, and self-renewal, consistent with overexpression of CSC signatures/markers and tumor-associated mediators. Therefore, molecular understanding and targeting these CSLCs could provide a newer effective therapeutic strategy for the treatment of aggressive tumors such as PC. However, the regulation of CSLC characteristics during tumor development and progression is not well understood, and thus became the focus of our current investigation.
Without a doubt, miRNAs, small nonprotein-coding RNA molecules, are beginning to shed lights in understanding their roles in tumor development, tumor progression, and treatment failure through regulation of many target genes, including tumor-associated mediators [29 –33]. Moreover, emerging evidence suggests that miRNAs may also play a key role in the biology of CSCs or CSLCs during tumor development and progression; however, the precise molecular mechanisms in support of the role of miRNAs in the biology of CSLCs is not well understood.
Increasing number of evidence suggests that miR-30 may play an important role in tumor development and progression mediated by regulating several signaling networks such as epidermal growth factor and the processes epithelial-to-mesenchymal transition or in short epithelial-to-mesenchymal transition (EMT) [34]. However, the role of miR-30 in the regulation of CSC phenotypes and functions is not well understood. It has been shown that the expression of miR-30 is lost or downregulated during transforming growth factor (TGF)-β-mediated acquisition of EMT, a process being reportedly associated with drug resistance and CSC characteristics in human colon cancer cells [35]. The expression of miR-30 has been found to be inversely associated with the expression of CSC signature Lin-28 during differentiation and proliferation of embryonic stem cells, and CSC-like cells [36]. The levels of miR-30 are significantly downregulated in Lin-28-positive tumor (CSC-like) cells, compared with Lin-28-negative tumor (non-CSC-like) cells [36]. One experimental study has shown that breast cancer cells produced a remarkably low levels of miR-30 family, including a, b, c, and d, under CSC-like sphere forming assay conditions. The target screening analysis showed that miR-30 regulates the expression of apoptosis- and proliferation-related genes, such as AVEN, FoxD1, IDH1, and TFDP1. Interestingly, forced overexpression of miR-30a has been shown to inhibit self-renewal capacity of breast cancer cells, which was associated with reduction of breast tumor in vivo [37]. Another experimental study showed that CSC-like cells from breast cancer have very low levels of miR-30, consistent with the overexpression of ubiquitin-conjugating enzyme 9 (Ubc9) and integrin beta3 (ITGB3), both of which are known to be associated with tumor development and progression via regulating miR-30 targeting genes [23]. Forced overexpression of miR-30 in the CSC-like cells derived from breast cancer cells showed decreased self-renewal capacity by repressing Ubc9, and also induced apoptosis through inhibition of ITGB3. The knockdown of miR-30 through its siRNA inhibitor transfection restored self-renewal capacity in differentiated breast cancer cells. Moreover, forced overexpression of miR-30 reduced tumorigenesis and lung metastasis in a mouse xenograft model of CSC-like cells derived from breast cancer cells. Our data show that CSLCs (triple-marker positive; CD44+/CD133+/EpCAM+ cells) derived from PC MiaPaCa-2 cells have reduced levels of miR-30a,c, compared with both parental MiaPaCa-2 cells and triple-marker-negative cells. These data suggest that miR-30 may play an important role in the regulation of CSC characteristics during tumor development and progression in PC, and as such strategies by which the expression of miR-30 family could be induced (restored) would become a novel approach for the treatment of PC.
A large number of studies have provided clear evidence in support of the role of miR-125b in tumor development and progression [25]; however, the precise mechanistic role of miR-125b in the regulation of CSC phenotypes and functions has not been well understood. It has been shown that CSC-like glioma stem cells of SU3 cells overexpress miR-125b, which exhibits oncogenic behaviors in vitro and in vivo [27]. The knockdown of miR-125b by its siRNA inhibitor resulted in the inhibition of invasive potential of glioblastoma cells by targeting PIAS3 (protein inhibitor of activated STAT) via inactivation of STAT3/MMP axis [26]. Moreover, another study found a positive relationship between Oct4 expression (a known CSC signature protein) and miR-125b in cervical cancer cells, in which Oct4 caused overexpression of miR-125b, resulting in the promotion of tumorigenesis in vivo [38]. Further, the overexpression of miR-125b has been identified in human glioblastoma CD133-positive (CSC-like) cells. Overexpression of miR-125b also led to increased cell migration, whereas the knockdown of miR-125b reduced cell migration in glioblastoma CSC-like cells, which was found to be associated with the expression of MMPs [39]. In contrast, one experimental study reported downregulation of miR-125b in PC Capan-1 and HPAC sphere-forming cells [40], and these results are opposite to our findings. We found overexpression of miR-125a in CSLCs (triple-marker-positive cells) derived from MiaPaCa-2 cells, which was consistent with tumor cell aggressiveness. The knockdown of miR-125b by its siRNA inhibitor resulted in the inhibition of clonogenicity, self-renewal capacity, and cell migration, which was consistent with the downregulation of CD44, EpCAM, EZH2, and Snail. EZH2 and Snail are known markers of aggressive tumor cells, and CSC regulators [5,41 –43]. These data clearly suggest that miR-125b may have a critical role in the regulation of CSC phenotypes and functions, and thus it could become a novel target for therapy.
Accumulating evidence showed that miR-335 plays an important role in tumor development and progression. Several clinical studies showed that miR-335 is upregulated and associated with poor clinical outcome of various tumors, such as prostate cancer, meningiomas, and glioma [20,44 –46]. In contrast, other studies have shown downregulation of miR-335, which was associated with poor clinical outcome of patients diagnosed with adrenocortical carcinoma, thyroid carcinoma, and prostate cancer [47 –49]. Another recent study showed reduced expression of miR-335 in the tumor tissues of 23 PC patients, compared with the normal pancreatic tissues of the same patients, and this was correlated with the expression of Sox2, a potential CSC marker [50]. It has been found that forced overexpression of miR-335 resulted in the inhibition of cell growth, clonogenicity, and cell migration in neuroblastoma cells, and liver metastasis in vivo [51]. The pathway/network analysis revealed that miR-335 may be involved in the regulation of various target genes such as pro-oncogenic signaling pathways, including p53, MAPK, TGF-β, Wnt, Rb, and mTOR [20]. Other experimental studies suggest that miR-335 may have tumor-promoting function [45]. These mixed findings suggest that the biological functions of miR-335 in tumor development and progression may be context dependent; however, the role of miR-335 in the regulation of CSC characteristics is not well understood. In our study, we found overexpression of miR-335 in CSLCs (triple-marker-positive cells) derived from PC MiaPaCa-2 cells, which supports some of the reported findings.
In summary, our data showed that CSLCs (triple-marker positive; CD44+/CD133+/EpCAM+ cells) display aggressive phenotypes and functions, such as increased cell growth, clonogenicity, cell migration, and self-renewal capacity, which is in part due to differential expression of a large number of miRNAs, including let-7f,i, miR-30a,b, miR-125b, and miR-335, compared with its parental MiaPaCa-2 cells and triple-marker-negative cells. The knockdown of miR-125b by transfecting siRNA inhibitor reduced clonogenicity, cell migration, and self-renewal capacity, which was consistent with downregulation of CSC signatures/mediators, such as CD44, EpCAM, EZH2, and Snail, suggesting that miRNA-targeted approach could become a novel therapeutic strategy for PC.
Footnotes
Acknowledgment
The authors thank Puschelberg and Guido Foundations for their generous financial contribution.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.