
Abstract
The mechanisms responsible for the inhibitory effects of mesenchymal stem cells (MSCs) on dendritic cells (DCs) are still poorly understood. Our investigation of the potential signaling pathways revealed for the first time that human umbilical-cord-derived MSCs (UC-MSCs) instruct DCs to acquire tolerogenic phenotypes through the IL-6-mediated upregulation of SOCS1. This subset of MSC-DCs exhibited a tolerogenic pattern, with a clear decrease in the expression of co-stimulatory molecules and the capacity to stimulate CD3+ T cell proliferation and inflammatory factor secretion, and a significant increase in the production of inhibitory cytokine IL-10 and the ability to induce Treg cells and Th2 responses. Adoption of this tolerogenic pattern required the activation of SOCS1, which blocked DC maturation by impairing TLR4 signaling. The effects of UC-MSCs on SOCS1 activation were essentially mediated by the JAK-STAT pathway via IL-6 secretion. In summary, our data identify a new mechanism, involving the IL-6-mediated upregulation of SOCS1, by which UC-MSCs instruct DCs to acquire tolerogenic phenotypes.
Introduction
M
Though MSCs were first isolated from bone marrow, it has since been shown that MSCs can be derived from different adult tissues (eg, cord blood and adipose tissue) as well as from umbilical cord (UC-MSCs) [17 –19]. Compared with MSCs isolated from other sources, UC-MSCs offer distinct advantages, including easier accessibility, more primitive properties, higher proliferation capacity, and lower immunogenicity. Further, UC-MSCs have the potential to become an ideal source of stem cells for clinical applications [20,21]. Studying the immunoregulatory mechanisms of UC-MSCs is therefore important to promote their clinical applications.
Dendritic cells (DCs), the most potent antigen-presenting cells (APCs), are uniquely able to activate naive T cells and thereby initiate an immune response [22]. Previous studies demonstrated that DCs could also induce tolerance or downregulate the immune response, depending on their maturation state and the specific subset of DCs involved [23]. Tolerogenic DCs usually express low or intermediate levels of maturation markers while overexpressing IDO or IL-10 [24]. In addition, tolerogenic DCs can be induced in vitro by incubation with IL-10, TGF-β, hepatocyte growth factor (HGF), and vasoactive intestinal peptide [25 –27,28]. Recent studies suggest that BM-MSCs may influence the differentiation and function of DCs through the secretion of prostaglandin E2 (PGE2), IL-10, and IL-6 [29 –32]. Additionally, BM-MSCs may induce adult CD34+ hematopoietic progenitor cells to differentiate into regulatory DCs through the activation of the Notch pathway [33]. However, the underlying immunoregulatory effectors involved in the regulation of DCs by UC-MSCs have not been reported.
The JAK-STAT pathway is the primary signaling mechanism utilized by a wide array of mammalian cytokines and growth factors. JAK activation stimulates cell proliferation, differentiation, migration, and apoptosis. Various cytokines, including IL-6, can activate JAK by binding to its transmembrane receptor, thereby inducing the phosphorylation of STAT (STAT1/3) [34]. Phosphorylated STAT then translocates into the nucleus, where target genes such as SOCS1 are subsequently activated [35]. Accumulating evidence suggests that SOCS1, a member of the SOCS gene family, can regulate various cells of the immune system. SOCS1-knockout (SOCS1−/−) mice die within 3 weeks of birth from a syndrome characterized by severe lymphopenia, peripheral T cell activation, fatty degeneration and necrosis of the liver, and macrophage infiltration of major organs [36,37]. Using T cell-specific conditional knockout mice, the Tanaka et al. group demonstrated that most SOCS1−/− CD4 naive T cells differentiate into Th1 cells, even under nonskewing conditions, whereas Th17 differentiation is strongly suppressed [38]. In addition, recent studies reported that SOCS1 appeared to suppress DC maturation and function and prevent the development of systemic autoimmunity [39 –41]. DCs treated with SOCS1 small interfering RNA (siRNA) exhibited enhanced cytokine responses to lipopolysaccharide (LPS) and augmented polyfunctional cytotoxic T-lymphocyte responses [39].
Although recent studies have shown that BM-MSCs can regulate the differentiation and maturation of DCs through the secretion of specific cytokines and the formation of cell–cell contacts in vitro, the underlying mechanisms are still poorly understood. In the present study, we found that UC-MSCs instruct DCs to acquire tolerogenic phenotypes through the IL-6-mediated upregulation of SOCS1. This subset of MSC-DCs exhibited tolerogenic characteristics, with a clear decrease in the expression of co-stimulatory molecules and the capacity to induce CD3+ T cell proliferation and inflammatory factor secretion and an increase in the production of the inhibitory cytokine IL-10 and the ability to induce Treg cells and Th2 responses. Adoption of this tolerogenic pattern required the activation of SOCS1, which blocked DC maturation by impairing the TLR4 signaling. The effects of UC-MSCs on SOCS1 activation were mediated by the JAK-STAT pathway through the secretion of IL-6.
In summary, our data reveal a new mechanism, involving the IL-6-mediated upregulation of SOCS1, by which UC-MSCs instruct DCs to acquire tolerogenic phenotypes. The results of our study may help to explain the effects of UC-MSCs on DCs and may ultimately facilitate the translation of UC-MSC research into its clinical application.
Materials and Methods
Culture of human UC-MSCs
This experiment was approved by the Research Ethics Committee of the Third Affiliated Hospital of Sun Yat-sen University. Fresh human umbilical cords were obtained after birth, with the written consent of the parents, and collected in phosphate-buffered saline (PBS) containing 100 UI/mL penicillin and streptomycin (Life Technologies) at 4°C. The fresh umbilical cords were then processed within the optimal processing period of 6 h. The cords were washed twice with PBS containing penicillin and streptomycin, and the cord blood was removed during this process. The rinsed cords were cut into 0.5-cm pieces and floated in Dulbecco's modified Eagle's medium with low glucose (Life Technologies) containing 10% FBS (Life Technologies), 5% HS, penicillin, and streptomycin at 37°C in a humidified atmosphere with 5% CO2. The medium was changed every 2 days, and nonadherent cells were removed by washing after 7 days. When well-developed colonies of fibroblast-like cells appeared after 10 days, the cultures were trypsinized and transferred (without dilution) into a new flask for further expansion. The medium in this flask was also changed every 2 days.
Differentiation of human CD14+ monocytes
Peripheral blood mononuclear cells (PBMCs) were isolated from healthy blood by Ficoll-Paque (1.077 g/mL; Invitrogen) density gradient centrifugation. Monocytes (more than 90% CD14+) were purified from PBMCs using the MACS Monocyte Isolation kit (Miltenyi Biotec, Bergisch Gladbach), and 106 cells per milliliter were cultured in 3 mL of RPMI 1640, 10% FBS, recombinant GM-CSF (rGM-CSF) (50 ng/mL; PeproTech), and rIL-4 (50 ng/mL; PeproTech) in six-well flat-bottom plates. Cytokines were added to the cultures every 3 days. At day 6, DCs were harvested and cocultured with MSCs. To induce DC maturation, LPS (100 ng/mL; Sigma-Aldrich) was added for another 48 h.
Coculture experiment
Once human UC-MSCs reached 50% confluence, DCs were seeded onto MSC monolayers at a density of 106 cells per 4 mL per well in six-well plates, and MSC culture medium was replaced with RPMI 1640 supplemented with 5% FBS. LPS (100 ng/mL) was added at day 5. DCs were separated from MSCs by spinning the cells in suspension and then washing them. Neutralizing IL-6 monoclonal antibodies (mAbs) were added at days 0, 3, 5, and 7 at a concentration of either 5 or 10 μg/mL (R&D Systems). For the siRNA assay, DCs were transfected with siSOCS1.
Flow cytometry
The Abs used for flow cytometry were FITC-, PE-, or APC-conjugated mouse anti-human CD73, CD90, CD105, CD45, CD34, CD44, CD166, CD3, CD4, HLA-DR, CD1a, CD14, CD80, CD86, and CD83. Anti-CD73, CD90, CD105, CD45, CD34, CD44, and CD166 were purchased from eBioscience, while anti-CD3, CD4, HLA-DR, CD1a, CD14, CD80, CD86, and CD83 were purchased from BioLegend. The cultured cells were collected, washed twice, and resuspended in 100 μL of PBS containing 0.1% BSA. These cells were stained and labeled with either specific Abs or the appropriate isotype controls. The cells were incubated on ice for 30 min, washed with PBS containing 0.1% NaN3 and 0.5% BSA, and then fixed with a 1% paraformaldehyde solution. Analyses were performed by FACScan using CellQuest software (BD Bioscience). For the intracellular staining of T cells, peripheral blood lymphocytes (PBLs) were stimulated for 5 h with phorbol 12-myristate 13-acetate (PMA) plus ionomycin (Sigma-Aldrich). Brefeldin A (10 mg/mL; Sigma-Aldrich) was added during the last 3 h. The cells were washed and stained with mAbs specific for CD4 for 20 min at 4°C. After being washed in PBS, the cells were fixed and permeabilized with fixation/permeabilization solution (eBioSciences) for 15 min at room temperature. Following further washing, the cells were stained with mAbs against IFN-γ and IL-4 (eBioSciences) for 20 min at room temperature.
Assay for cytokines
Coculture supernatants were collected on days 3, 5, 7, and 9. The concentrations of IL-4, IL-6, IL-8, IL-10, IL-12p40, IL-17, HGF, VEGF, and TNF-α in the supernatants were determined using the Bio-PlexH Suspension Array System. In some experiments, cells were cultured for 24 h with or without LPS (100 g/mL) stimulation. Culture supernatants were collected, and the concentrations of IL-2, IL-10, and IL-12p70 were determined using ELISA kits (R&D Systems). Cytokines, such as IL-10, TNF-α, TNF-β, IL-17, IL-1a, and IL-1ra, were detected in the supernatants at day 5 of the mixed lymphocyte reaction (MLR) using the Bio-PlexH Suspension Array System.
Endocytosis assay
To determine the phagocytic ability of DCs, DCs or MSC-DCs with or without LPS were incubated for 30 min at 37°C, or at 4°C as a negative control, with FITC-conjugated OVA (Sigma-Aldrich) at a final concentration of 100 μg/mL in RPMI 1640 containing 10% FBS. The cells were then washed twice with ice-cold PBS containing 0.1% NaN3 and 0.5% BSA and resuspended in ice-cold PBS for immediate analysis by flow cytometry.
Migration assay
Cell migration assays were performed in 24-well Transwell chambers (Corning Costar) using polycarbonate membranes with a pore size of 5 μm. Approximately 2×105 DCs were loaded into the upper chamber. CCL21 (100 ng/mL; R&D Systems) was added to the lower chamber. After 3 h of incubation at 37°C with 5% CO2, the number of cells in the lower chamber was determined by flow cytometry. The cells were resuspended in exactly 100 μL of PBS, and a fixed amount of fluorescent beads was added. As it is nearly impossible to measure equal volumes of different samples, we used equal numbers of beads so that we could correct for the amount of migrated cells in each sample.
Mixed lymphocyte reaction
Once labeled with carboxyfluorescein diacetate succinimidyl diester (CFSE) (Invitrogen) and suspended in RPMI 1640 supplemented with 10% FBS, 100 U/mL penicillin, and 100 U/mL streptomycin, allogeneic PBLs from PBMCs were cultured at a concentration of 1×105 cells per 200 μL per well in 96-well U-bottom plates containing 2×104 DCs treated with mitomycin. After 5 days, cells were harvested and sorted by FACS.
Western blot
Equal amounts of protein (30–60 μg) from each sample were separated by 12% SDS-PAGE and transferred to PVDF membranes (Millipore). The proteins were then incubated overnight at 4°C with primary antibodies against human SOCS1, phospho-JIN1/2, JNK1/2, phospho-ERK1/2, ERK1/2, phospho-P38, P38, phospho-AKT, AKT, NF-κB, phospho-STAT3, STAT3 (Cell Signaling Technology), and β-actin (Santa Cruz Biotechnology). Following a 30-min wash, the membranes were incubated with a horseradish-peroxidase-conjugated secondary antibody for 1 h at room temperature. After being washed for 30 min, the membranes were visualized by enhanced chemiluminescence (ECL; Millipore) and recorded on Kodak film.
Real time–polymerase chain reaction
Total RNA was extracted using TRIzol (Invitrogen) and then reverse transcribed using a QuantiTect RT kit (Qiagen). PCR products were analyzed by electrophoresis on 2% agarose gel and photographed after ethidium bromide staining. In some experiments, gene expression was evaluated using an SYBRGreen real-time PCR kit (Takara). Data were normalized to the reference gene β-actin. The forward primer for SOCS1 was GGAAGGAGCTCAGGTAGTC, and the reverse primer was AGCTTCGACTGCCTCTT.
siRNA silencing assay
Three pairs of SOCS1 siRNAs (3′dTdT GGUCUUGGAAGGAGGAGAA 5′, 3′dTdTUGUGCGUGAAGGCGUGUAA5′, and 3′dTdTUGCGUGAAGGCGUGUAAG 5′) and a negative control were synthesized by Guangzhou RIBOBIO Company. siRNAs were transfected into DCs using Lipofectamine RNAiMAX (Invitrogen), according to the instructions.
Statistical analysis
Statistical analyses were performed using Student's t test. Data are depicted as mean±SEM. P values≤0.05 were considered significant. *P≤0.05, **P≤0.01, and ***P≤0.001. All statistical analyses were conducted using the Statistical Program for Social Sciences 13.0 software program (SPSS, Inc.).
Results
Characteristics of human UC-MSCs
Human UC-MSCs were isolated as previously described and identified based on flow cytometric analysis and by their differentiation capacity. The UC-MSCs formed colonies of adherent, fibroblast-like cells (Fig. 1B) expressing CD90, CD73, CD105, CD44, and CD166 but not hematopoietic cell markers, such as CD34 and CD45 (Fig. 1A). The UC-MSCs also did not express CD14 and HLA-DR (Fig. 1A). These results are consistent with the previously reported surface antigen expression profiles of BM-MSCs.

Characterization of human umbilical cord mesenchymal stem cells (MSCs).
To further identify UC-MSCs, we determined the differentiation capacity of the expanded MSCs. As shown in Fig. 1C, the UC-MSCs could be differentiated into classical mesenchymal lineage cells, such as osteoblasts (Fig. 1C, left) and adipocytes (Fig. 1C, right).
MSC-DCs phenotypically resemble tolerogenic DCs
DCs were seeded onto UC-MSC monolayers to evaluate the effects of MSCs on DC morphology and phenotype. During coculture, DCs aggregated vigorously and adhered to the UC-MSCs (Fig. 2A). In contrast to DCs, which scatter and have many long dendrites in the presence of LPS, UC-MSC-cocultured DCs still gather together and display fewer and shorter dendrites after LPS treatment, similar to monocytes in the presence of GM-CSF and IL-4 (Fig. 2A). Moreover, FACS analysis showed that, in contrast to DCs or DCs before coculture/culturing alone (B-DCs), UC-MSC-cocultured DCs expressed relatively low levels of functional markers, such as the antigen-presenting molecule CD1a; the costimulatory molecules HLA-DR, CD80, and CD86; and the maturation marker CD83. However, UC-MSC-cocultured DCs expressed higher levels of CD14 compared with DCs or B-DCs (Fig. 2B–H). Surprisingly, LPS treatment scarcely increased the expression of the aforementioned functional markers in UC-MSC-cocultured DCs compared with DCs or B-DCs (Fig. 2B–H), suggesting that these UC-MSC-instructed DCs (called MSC-DCs) acquired tolerogenic phenotypes.
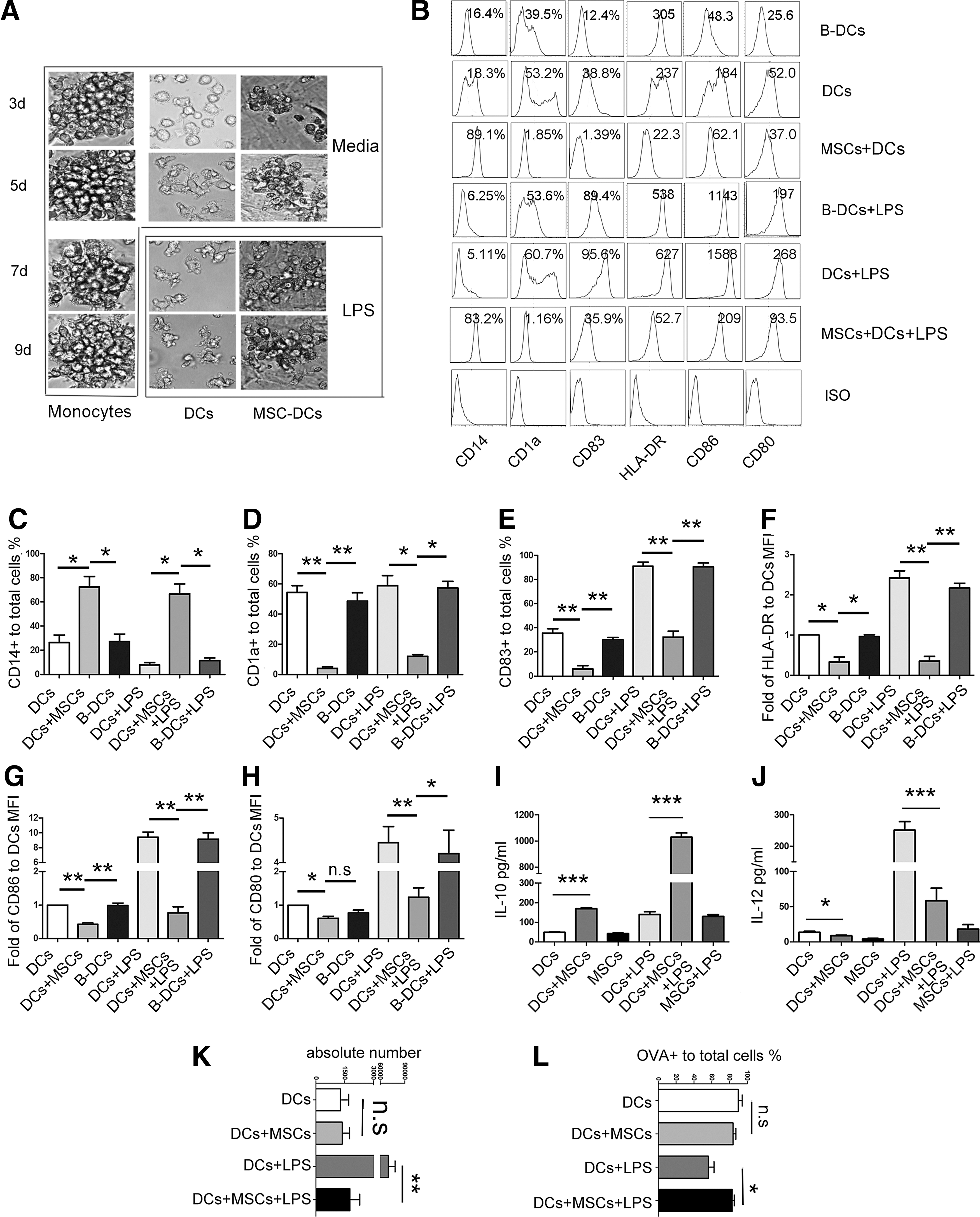
MSC-dendritic cells (DCs) phenotypically resemble tolerogenic DCs.
To further confirm the tolerogenic characteristics of MSC-DCs, we determined their cytokine expression patterns using ELISA. In contrast to DCs, DCs cocultured with UC-MSCs spontaneously secreted more IL-10 and less IL-12 (Fig. 2I, J). This pattern was strongly enhanced following LPS stimulation (Fig. 2I, J). As shown in Fig. 2K and L, UC-MSC-cocultured DCs still possessed greater phagocytic but lower migration capacities after LPS treatment. These results demonstrate that DCs acquire tolerogenic phenotypes in presence of UC-MSCs.
MSC-DC functions exhibit tolerogenic characteristics
Because UC-MSCs appeared to instruct DCs to acquire tolerogenic phenotypic feature, we next evaluated the effects of UC-MSCs on DC function. PBLs with or without CFSE were used as responders and cocultured with either allogenic DCs, or MSC-DCs, at a ratio of 5:1, and T cell proliferation and cytokine production were evaluated after 5 days.
Notably, UC-MSCs not only induced tolerogenic DC-like phenotype, but also reduced the capacity of DCs to stimulate T cell proliferation (Fig. 3A, B and Supplementary Fig. S1A; Supplementary Data are available online at
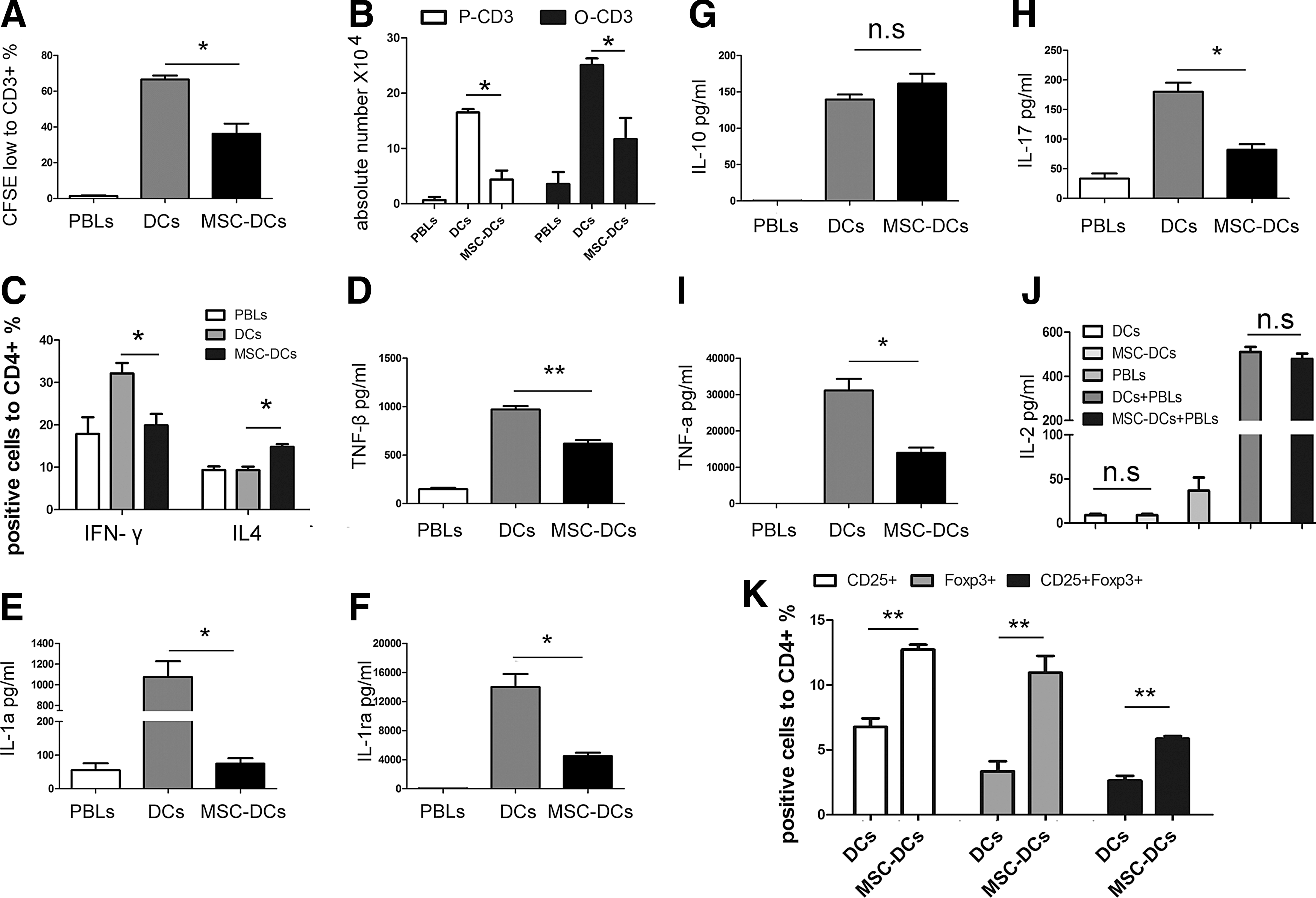
MSC-DCs exhibit tolerogenic functional characteristics.
UC-MSCs instruct DCs to acquire tolerogenic characteristics by upregulating SOCS1
To evaluate how UC-MSCs instruct DCs to acquire tolerogenic phenotypes, we analyzed the expression of SOCS1, which has been shown to be a key negative regulator of DCs. DCs exhibited significantly increased SOCS1 expression in the presence of UC-MSCs (Fig. 4A). To determine whether the induction of tolerogenic DCs by UC-MSCs was dependent on SOCS1 upregulation, DCs were transfected with SOCS1-siRNA (siSOCS1) and then cocultured with UC-MSCs. To select the most specific siRNA against SOCS1 out of three SOCS1-targeted sequences, we analyzed SOCS1 mRNA and protein levels via real-time PCR and western blotting, respectively. Figure 4B and C shows the knockdown efficiency of different SOCS1-targeted sequences; of the three SOCS1-targeted sequences, siSOCS1-2 clearly showed the highest knockdown efficiency in terms of both mRNA and protein levels. Thus, siSOCS1-2 and the relevant NC groups were chosen for further functional analysis. The expression of CD1a, CD80, CD86, and CD83 in siSOCS1-2-MSC-DCs increased significantly compared with the untreated MSC-DCs (Fig. 4D and Supplementary Fig. S2). The decreased expression of CD14 and increased expression of HLA-DR were not statistically significant (Fig. 4D and Supplementary Fig. S2). Moreover, in contrast to MSC-DCs, siSOCS1-2-MSC-DCs exhibited significantly increased IFN-γ expression by CD4+ T cells (Fig. 4F). However, the decreased expression of IL-4 was not statistically significant (Fig. 4F). Additionally, in contrast to MSC-DCs, siSOCS1-2-MSC-DCs partially recovered the ability to stimulate CD3+ T cell proliferation (data not shown). Finally, we studied the effect of SOCS1 on the induction of Treg cells and found that the induction of Treg-cell subsets by siSOCS1-2-MSC-DCs decreased significantly compared with MSC-DCs (Fig. 4E and Supplementary Fig. S1D). These results indicate that the tolerogenic characteristics of MSC-DCs are dependent on SOCS1 activation.

MSCs educate DCs to acquire tolerogenic phenotype by upregulating SOCS1.
SOCS1 mediates the impairment of TLR4 signaling in MSC-DCs
Given that MSC-DCs responded only slightly to LPS stimulation (Fig. 2B), we investigated the TLR4 signaling pathway that has been shown to activate and mediate the function of DCs in the presence of LPS. In contrast to DCs, MSC-DCs exhibited reduced SAKP/JNK, P38, and ERK phosphorylation after LPS stimulation (Fig. 5A, B). IRF3 was also phosphorylated to a lesser extent in MSC-DCs (Fig. 5C, D). A weaker NF-κB band was detected in MSC-DC nuclear extracts compared with those of DCs (Fig. 5E, F). As shown in Fig. 5A and B, MSC-DCs exhibited distinct increases in Akt phosphorylation (Fig. 5A, B). Thus, TLR4 signaling was impaired in MSC-DCs, not only in the MyD88-dependent but also in the MyD88-independent signaling pathway.
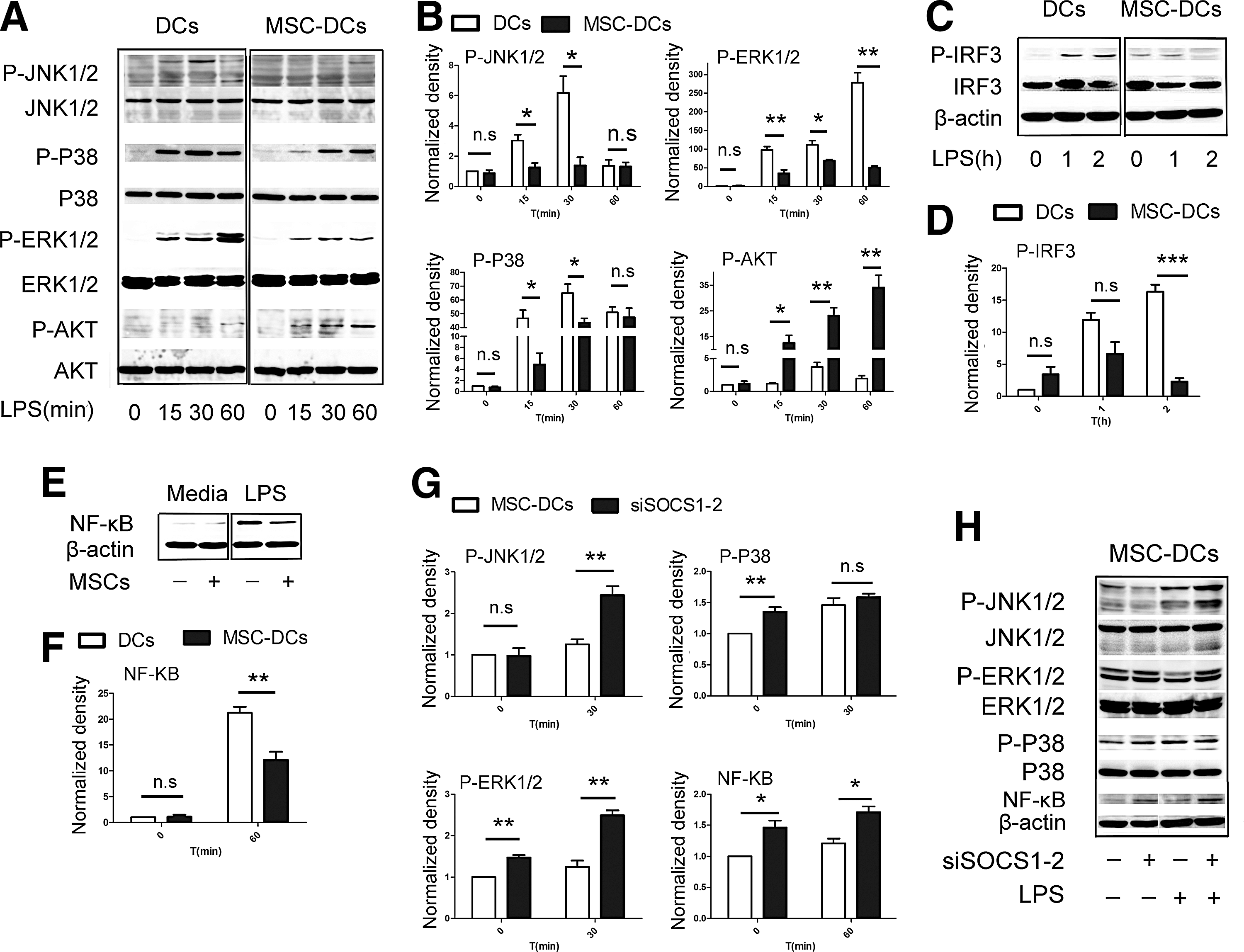
SOCS1 impairs TLR4 signaling in MSC-DCs.
Because the tolerogenic characteristics of MSC-DCs were dependent on SOCS1 activation, we investigated the effect of SOCS1 on TLR4 signaling. Our findings showed that the phosphorylation of SAKP/JNK, P38, and ERK distinctly increased in siSOCS1-2-MSC-DCs compared with MSC-DCs (Fig. 5G, H). Moreover, the nuclear translocation of NF-κB was also markedly increased in siSOCS1-2-MSC-DCs (Fig. 5G, H). Finally, DCs treated with the inhibitor of SAKP/JNK, P38, and ERK still expressed lower levels of CD1a, HLA-DR, CD80, and CD86 after LPS stimulation, which was in similar to MSC-DCs (data not shown). Thus, the activation of SOCS1 interfered with the maturation of MSC-DCs by inhibiting TLR4 signaling, which may represent a potential mechanism underlying their tolerogenic phenotype.
IL-6 upregulates SOCS1 via the JAK-STAT pathway
To identify the key factors responsible for the activation of SOCS1 by UC-MSCs, soluble factors present in the supernatants at various time points were measured. DCs cocultured with UC-MSCs had higher levels of VEGF, HGF, IL-6, and IL-8 and the anti-inflammatory factor IL-4 and IL-10 but lower levels of the pro-inflammatory factors IL-12p40, IL-17, and TNF-α compared with DCs alone (Fig. 6A–H). Moreover, our data showed that the expression of CD1a, CD80, CD86, HLA-DR, and CD83 decreased significantly whereas that of CD14 increased markedly in IL-6-treated DCs (Supplementary Fig. S3). These functional markers recovered somewhat when NA-IL-6 was added to the UC-MSC and DC cultures (Supplementary Fig. S3), suggesting that IL-6 influences the differentiation of DCs. Thus, we hypothesized that IL-6 interfered with the development of DCs by upregulating SOCS1. As expected, the expression of SOCS1 increased substantially when IL-6 was added to the culture medium but decreased significantly when NA-IL-6 was added (Fig. 6I). However, HGF and IL-10 could not appreciably upregulate the SOCS1 expression by DCs in our study (Supplementary Fig. S4).
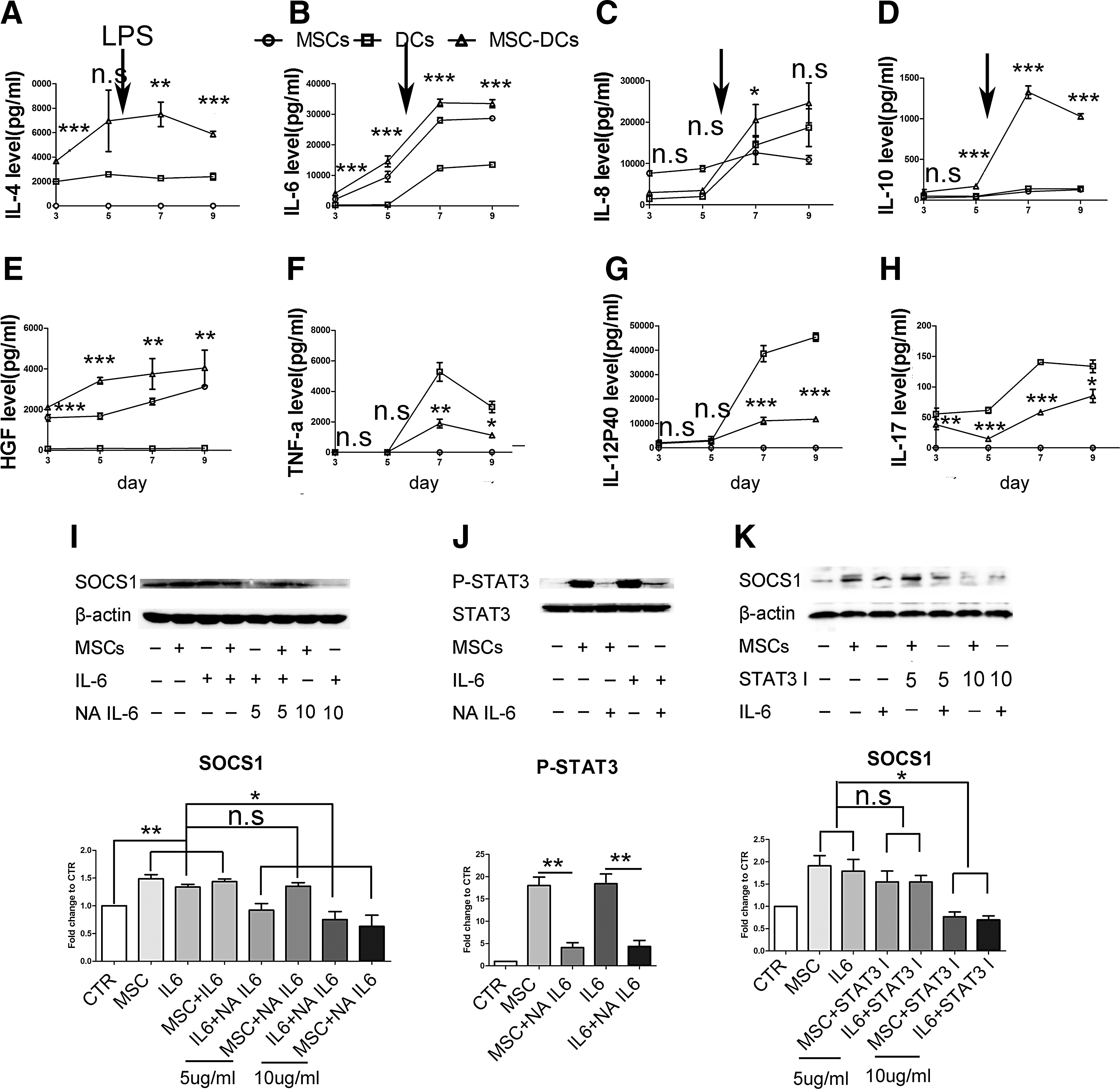
IL-6 upregulates SOCS1 via the JAK-STAT pathway.
To further explore the mechanisms underlying IL-6-mediated SOCS1 upregulation, we analyzed the JAK-STAT signaling pathway, which has been reported to induce the expression of SOCS1 in previous studies [42,43]. Our findings confirmed that the levels of phospho-STAT3 increased significantly in the presence of UC-MSCs or IL-6 but decreased considerably following the addition of NA-IL-6 (Fig. 6J). Further, the expression of SOCS1 was downregulated significantly when a STAT3 inhibitor (10 μg/mL) was added to the culture system (Fig. 6K). Thus, these data suggest that IL-6 upregulates the expression of SOCS1 via STAT3 activation.
Discussion
The inhibitory effect of MSCs on DCs is one of the ways by which UC-MSCs induce immunomodulation, but the mechanism has largely remained unclear. In the present study, we aimed to investigate the relevant signaling networks and found for the first time that UC-MSCs instruct DCs to acquire tolerogenic phenotypes through the IL-6-mediated upregulation of SOCS1.
In this study, we revealed that DCs cocultured with UC-MSCs exhibited reduced expression of various functional markers, spontaneously secreted higher levels of IL-10, and displayed a weakened capacity to induce lymphocyte proliferation and inflammatory factor secretion, along with an increased ability to induce Treg cell and Th2 responses. Thus, these MSC-DCs are very different from DCs, not only phenotypically, but also functionally. The characteristics of MSC-DCs were clearly comparable to those of tolerogenic DCs, as described in a previous report [24]. Therefore, this unique subset of MSC-DCs may in fact represent one form of tolerogenic DCs.
The SOCS gene family has been reported to regulate various cells of the immune system, particularly DCs. Recent studies have reported that SOCS1, a key physiological regulator of the immune system, appeared to suppress DC maturation and function and prevent the development of systemic autoimmunity [39 –41]. APCs obtained from SOCS1-deficient mice were hyper-responsive to LPS and produced large amounts of pro-inflammatory cytokines [44,45]. Using a DC-targeting approach to deliver siRNA to these cells, the Sandesh group found that SOCS1 silencing in myeloid-derived DCs resulted in enhanced cytokine responses to LPS and a strong MLR [46]. In the current study, we observed that DCs significantly increased the expression of SOCS1 and exhibited the tolerogenic characteristics after human UC-MSC treatment. These characteristics were lost when SOCS1 was knocked down, suggesting that the tolerogenic characteristics are dependent on the activation of SOCS1. Thus, SOCS1 is the key factor in the generation of MSC-DCs.
Our results showed that MSC-DCs responded only minimally to LPS stimulation. Therefore, we investigated the activation of the TLR4 signaling pathway in MSC-DCs, which involved in the phosphorylation of cytoplasmic components and nuclear transcription factors that are known to promote cytokine production after LPS stimulation. MSC-DCs exhibited little activation of TLR4 signaling not only in the MyD88-dependent but also in the MyD88-independent signaling pathway, as well as the impairment of NF-κB signaling after LPS stimulation. Previous reports showed that the activation of the TLR4 signaling pathway following LPS stimulation led to the NF-κB-mediated production of IL-12, a key cytokine driving the Th1 response [47]. Consistent with these findings, our data revealed that MSC-DC-mediated IL-12 secretion and Th1 responses decreased significantly, indicating that the impairment of NF-κB is responsible for the decreased IL-12 production and Th1 response. Moreover, DCs treated with SAKP/JNK, P38, and ERK inhibitors exhibited phenotypes similar to MSC-DCs, indicating that the TLR4 signaling is a key pathway involved in the tolerogenic characteristics of MSC-DCs. Given that our data showed that the tolerogenic characteristics of MSC-DCs were dependent on SOCS1 activation, we investigated the effect of SOCS1 on the TLR4 signaling pathway. We found that the inhibition of the TLR4 signaling pathway was significantly reversed when SOCS1 was knocked down, indicating that SOCS1 is responsible for the impairment of TLR4 signaling, which is in agreement with previous reports [48,49]. Thus, SOCS1 could maintain the tolerogenic characteristics of MSC-DCs by blocking their maturation through impaired TLR4 signaling.
Recent reports have shown that certain cytokines can activate the expression of SOCS1; therefore, we monitored the levels of soluble cytokines present in the cell culture supernatants at several time points. Encouragingly, our data showed that high levels of VEGF, HGF, IL-6, IL-8, and IL-10 were present in the coculture system; IL-6-treated DCs exhibited decreased CD1a, CD80, CD86, HLA-DR, and CD83 expression and increased CD14 expression. Thus, we focused on the effect of IL-6 on the expression of SOCS1. As expected, we observed that SOCS1 expression was considerably upregulated in DCs treated with IL-6. Moreover, decreased SOCS1 expression was observed when the coculture system was treated with an IL-6-neutralizing antibody. These data suggested that IL-6 secreted by UC-MSCs is responsible for the upregulation of SOCS1. Some studies have shown that IL-10 secreted by BM-MSCs could impair the maturation and function of DCs and that IL-10 could induce the production of tolerogenic DCs [27,28]. However, although IL-10 levels in the culture supernatants of UC-MSC-cocultured DCs were much higher than in the supernatants of DCs cultured alone at days 5, 7, and 9 in our study, UC-MSCs expressed only low levels of IL-10. These data indicate that IL-10 in the coculture supernatants was mainly derived from MSC-DCs rather than from UC-MSCs. Thus, we hypothesized that UC-MSCs instruct DCs to acquire tolerogenic phenotypes via IL-6 secretion and that these tolerogenic DCs (MSC-DCs) secreted IL-10 to further maintain their tolerogenic phenotypes. Moreover, we also explored the relationship between IL-10 and SOCS1 and found that IL-10 could not upregulate SOCS1 expression in DCs (Supplementary Fig. S4A, B), whereas IL-10 secretion was downregulated when MSC-DCs were transfected with siSOCS1-2 (Supplementary Fig. S4C). These data further support our hypothesis.
To further explore the mechanisms underlying the IL-6-mediated upregulation of SOCS1, we analyzed the JAK-STAT signaling pathway, one of the most important signal transduction cascades involved in the regulation of cytokine receptor signaling [35]. Our results showed that phospho-STAT3 levels increased in DCs after treatment with UC-MSCs or IL-6. Importantly, a neutralizing antibody against IL-6 has been shown to reduce the STAT3 activating effect of UC-MSCs and IL-6. Additionally, the expression of SOCS1 was downregulated significantly when a STAT3 inhibitor (10 μg/mL) was added to the coculture system. These results confirm that IL-6 upregulates SOCS1 expression via the JAK-STAT3 signaling pathway.
In summary, our results provide the first convincing evidence, showing that UC-MSCs instruct DCs to acquire tolerogenic phenotypes through the IL-6-mediated upregulation of SOCS1. Our present study provides novel information on the molecular mechanism of immunoregulatory effect of UC-MSCs on DC maturation and function (Fig. 7), which further supports that UC-MSCs may modulate the immune system not only through acting directly on T cells but also the initial step of antigen recognition by inhibiting DC differentiation and maturation. These findings may enhance the future prospects of the clinical application of UC-MSCs in new and broader immune-related applications; for example, in the transplantation of UC-MSCs to treat autoimmune diseases, graft rejection as well as severe graft-versus-host disease occurred after hematopoietic stem cell transplantation or solid organ transplantation. A thorough understanding of the mechanisms of action will facilitate the translation of basic knowledge of UC-MSC biology into new clinical applications.

A schematic showing the mechanism of UC-MSC-mediated immune tolerance.
Footnotes
Acknowledgments
This work was supported by National Natural Science Foundation of China (30972914, 81172036, 81170451, 81170452, and 81000190), 985 Project (82000-3281901), Fundamental Research Funds for the Central Universities (10ykpy05, 09ykpy21, and 12ykpy43), State Key Projects on Infection Diseases of China (2012ZX10002016-023, 2012ZX10002017-005, and 2012ZX10002010-001-007), Natural Science Foundation of Guangdong Province (10151008901000208 and S2012010008792), and key project of Medical Science in SYSU (10YKJC03).
Author Disclosure Statement
This work has no conflict of interest, including financial and other relationships.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.