
Abstract
Hepatocellular carcinoma (HCC) is a malignant tumor associated with a generally poor prognosis and a high rate of recurrence. HCC usually develops in the context of chronic liver diseases, and long-lasting premalignant conditions precede cancer development. A promising therapeutic approach is to eliminate precancerous cells, which are considered as the precursors of cancer stem cells, to prevent further malignant transformation. In this study, we identified a subpopulation of precancerous cells in a rat liver carcinogenesis model, which were enriched in CD133+CD44+CD45−HIS49− cells that formed part of the hepatic oval cells fraction. Prospective isolation of the precancerous cells using flow cytometry identified stem cell properties such as the ability to expand clonally and differentiate into bi-lineage cell types. Furthermore, an acyclic retinoid, which was recently shown to improve overall survival after HCC resection, directly inhibited the extensive expansion of the isolated precancerous cells in vitro and decreased the emergence of the precancerous cells and their progeny in vivo. Long-term follow-up after the acyclic retinoid treatment confirmed reduction in precancerous changes, ultimately resulting in suppression of HCC development. These findings, together with data from recent clinical trials showing marked reduction in intrahepatic recurrence, suggest that acyclic retinoid directly prevents de novo HCC by inhibiting the development of precancerous cells. Given recent advances in diagnostic techniques and the establishment of surveillance programs, the targeting of precancerous cells may have a huge impact on preventative cancer therapies.
Introduction
H
Numerous studies have shown that several cancers are initiated from cancer stem cells in various solid tumors [2
–8]. Since the precancerous lesions or changes are observed in almost all types of cancers [9], at this very early stage of cancer development, the precancerous cells, also referred to as precancerous stem cells or progenitors, are considered as the precursors of cancer stem cells [10] and are promising therapeutic targets for early cancer detection and prevention [11,12]. The cellular pathology of precancerous cells exhibits abnormal development, which is indicative of an early precancerous change at the cellular level. The natural history of cancer remains unclear, but multistep neoplastic progression probably involves multiple somatic mutations. It is possible to transform normal hepatic stem cells into cancer stem cells with the potential to form tumors [13]. Thus, it is hypothesized that the transformation of normal cells to cancer cells accompanies precancerous changes in epithelial neoplasms, such as uterine cervical, HCC, gastric, colorectal, pancreatic, cutaneous, and oral carcinomas (summarized in Supplementary Fig. S1; Supplementary Data are available online at
A prospective clinical study showed that oral administration of acyclic retinoids significantly inhibits HCC recurrence [26,27]. The inhibitory effect of acyclic retinoids was supported by the finding of lower vitamin A (all-trans retinol) levels in human HCC samples compared with surrounding tissues [28]. Studies in rats and mice have also shown similar results [29], and a causal relationship between the hepatic loss of retinoic acid function and the onset of liver tumors [30]. Furthermore, acyclic retinoids persistently inhibit HCC development even after they are no longer administered, which is potentially a result of the clonal deletion of latent malignant cells [31]. Thus, acyclic retinoid-based therapy is a promising treatment option for HCC.
However, the cells targeted by acyclic retinoids and their therapeutic mechanism of action remain unclear, which is a hindrance to the adoption of this approach in clinical practice. The elucidation of the cellular origin of human HCC and development of novel therapeutic targets is obviously critical. In this study, we attempted to identify the subpopulation of precancerous cells that contribute to HCC and the mechanism by which acyclic retinoids suppress HCC development.
Materials and Methods
Animals and study design
Eight-week-old male Fischer 344/N Slc rats were purchased from Japan SLC (Shizuoka, Japan). All animal experiments were performed according to protocols approved by the Laboratory Animal Resource Center of Yokohama City University (No. 10–34). To induce the development of hepatic oval cells and hepatocarcinogenesis, the animals were fed a diet containing 2-acetylaminofluorene (2-AAF) at a dose of 12 mg/kg body weight/day. Partial hepatectomy (PH) was performed, and all animals were subjected to surgical removal of two-thirds of the liver after treatment with 2-AAF for 1 week. PH was performed under inhalational anesthesia using Escain isoflurane (Mylan, Osaka, Japan). The acyclic retinoid peretinoin (Kowa, Tokyo, Japan) was administered intragastrically each day. Rats were randomly assigned to one of four treatment groups, and peretinoin was administered at a dose of 0, 40, 60, or 80 mg/kg body weight for 240 days. The dose was temporarily decreased to 40 mg/kg for 1 week after PH in the group that received 80 mg/kg for more than 2 weeks (Supplementary Fig. S2).
Immunohistochemistry
Cryostat sections and cells cultured in dishes were fixed using methanol:acetone (1:1), and cytospin analysis was performed with sorted cells, which were fixed in acetone before immunohistochemical staining. Primary antibodies specifically detected cytokeratin 19 (CK19; Progen, Heidelberg, Germany), cytokeratin 7 (CK7; Dako; Glostrup, Denmark), glutathione S-transferase placental form (GSTP; MBL, Nagoya, Japan), OV-6 (R&D Systems, Minneapolis, MN), CD44 (BD Biosciences, Franklin Lakes, NJ), CD133 (Abcam, Cambridge, UK), EpCam (Abcam), or Ki67 (Abcam). Alexa Fluor 488-, Alexa Fluor 555-, or Alexa Fluor 647-conjugated goat anti-mouse IgG, IgG2b, or goat anti-rabbit IgG (Invitrogen Molecular Probes, Eugene, OR) were used as secondary antibodies. GSTP-positive foci were counted with WinROOF software (ver. 6.1; Mitani, Tokyo, Japan).
Real-time RT-PCR
Total RNA was extracted from freshly sorted cells or liver tissue using Isogen reagent (Nippon Gene, Toyama, Japan). One milligram of total RNA was converted into cDNA using SuperScript III Reverse Transcriptase (Invitrogen, Carlsbad, CA). cDNA was amplified with SYBR Premix ExTaqII (Takara Bio, Otsu, Shiga, Japan) using an ABI PRISM 7700 System (Applied Biosystems, Foster City, CA). Quantitative PCR was performed using ABI TaqMan Gene Expression Assays (Applied Biosystems) for α-fetoprotein (Afp; assay ID: Rn00560661_m1) and with other primer sets from Takara Bio (Table 1).
Cell isolation and flow cytometric sorting
Hepatic nonparenchymal cells were isolated from rats administered 2-AAF 7 days after PH using a standard 2-step collagenase perfusion method [32]. Antibodies were used to detect CD45 (BD Biosciences), erythroid cells/HIS49 (HIS49 clone; BD Biosciences), and CD133 (Abcam). Fluorescein isothiocyanate (FITC)-conjugated CD44 (BD Biosciences), Alexa 647-conjugated anti-rabbit IgG (Invitrogen), and phycoerythrin (PE)-conjugated streptavidin (Invitrogen) were also obtained. Analysis and sorting were performed using a MoFlo sorter (Beckman Coulter, Glostrup, Denmark).
Clonal colony assay in vitro
We used a low-density clonogenic culture system [33,34]. Cells were treated with the acyclic retinoid peretinoin after the first 24 h of culture. In all cultures, the final concentrations were 30 μg/L peretinoin and 0.03% dimethyl sulfoxide (the same concentration was used in the controls). We assessed clonogenesis and proliferation in the fractionated cells by assessing the colony size and the relative frequency of colony formation after 1 week of culture (only colonies containing >20 cells were counted).
Statistical analyses
All data are expressed as the mean±SD or SE. The results were compared using Mann–Whitney U-tests for two groups, with Bonferroni correction when multiple groups were compared. P-values less than 0.05 were considered significant.
Results
Development of hepatic oval cells is accompanied by emergence of precancerous cells
Hepatic stem/progenitor cells contribute to liver regeneration if hepatocyte proliferation is impaired. To induce this process, the rats were treated with 2-AAF and subjected to PH, after which responses to liver injury were examined. Oval and ductal cells were identified using CK19 in and around the periportal region (Fig. 1A and Supplementary Fig. S3A). We also identified foci positive for GSTP, a marker protein for precancerous changes in the liver that is undetectable in normal rat livers [35 –37]. Foci were observed from day 28 (Fig. 1A).
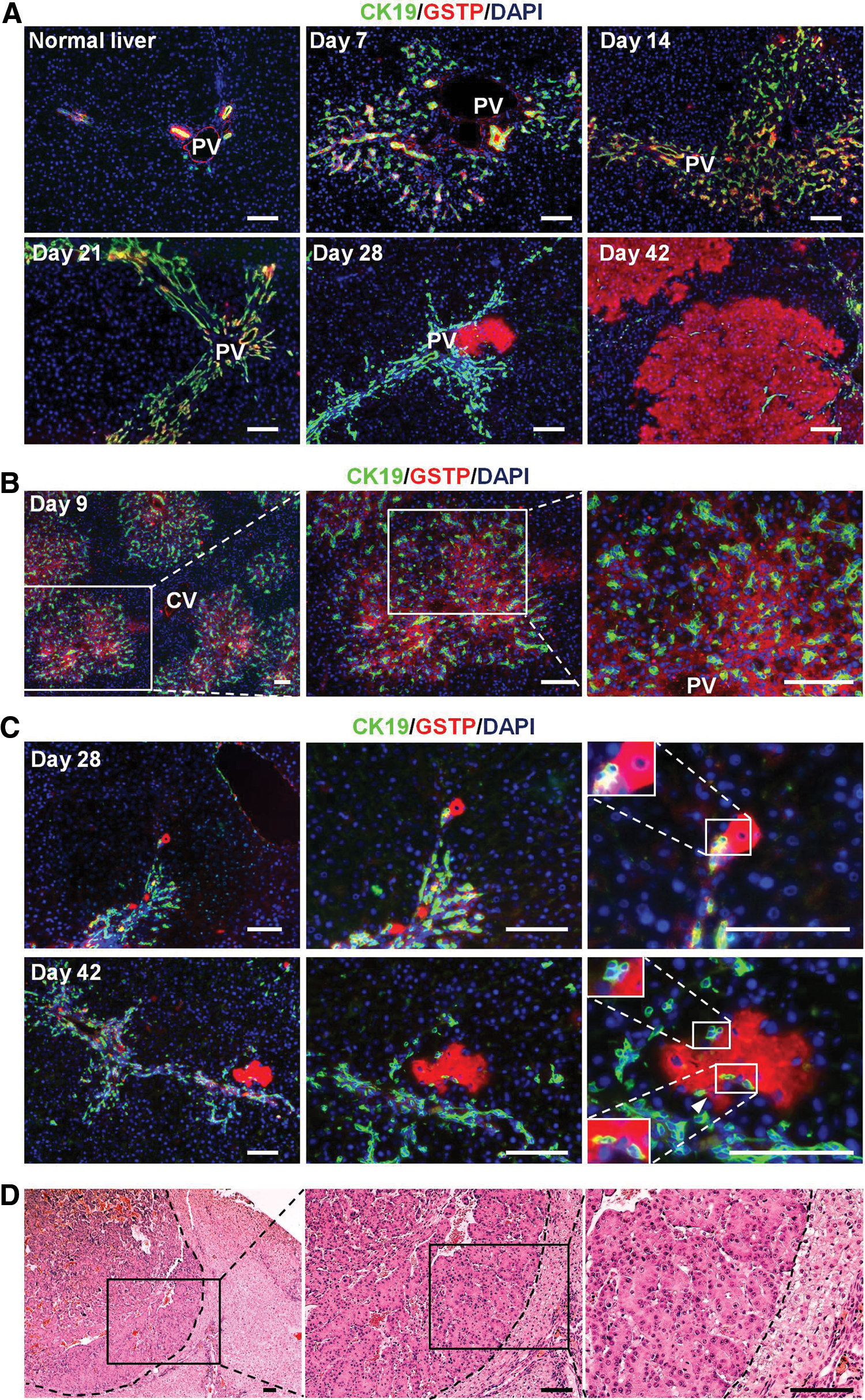
Development of HCC in a rat model. Development of hepatic oval cells, GSTP-positive cells, and foci, and HCC in rats treated with 2-AAF and subjected to PH.
Immunocytochemical tracing was performed to determine whether the emergence of hepatic oval cells and GSTP-positive cells and foci were correlated. In CK19-expressing oval cells, GSTP-positivity was cytologically distinct with a relatively low frequency (25.4%±3.8%, Fig. 1B) after initial treatment with 2-AAF and PH on day 7. In addition to oval cell development, high-intensity GSTP-positive foci emerged, and they frequently overlapped or were adjacent to oval cells expressing CK19 (Fig. 1C). After GSTP-positive foci emerged on day 28, a significantly higher percentage of the foci were adjacent to oval cells compared with that on day 42 (80.0%±9.5% vs. 67.8%±11.0%, P<0.05; Table 2). Furthermore, the spatial relation images produced using serial sections showed that most of the GSTP-positive foci colocalized and/or made contact with hepatic oval cells (Supplementary Fig. S3B), suggesting that the GSTP-positive foci were probably derived from the GSTP-positive subpopulation of hepatic oval cells.
P<0.05, Mann–Whitney U-test.
GSTP, glutathione S-transferase placental.
Pathological diagnoses of hepatocarcinogenesis were confirmed on the basis of architectural and cytological features in rats at 300 days after PH (Fig. 1D and Supplementary Fig. S3C). The results suggested that hepatic precancerous changes and HCC followed GSTP-positive hepatic oval cell development in this rat model.
Hepatic precancerous cell isolation and characterization
One week after PH, several OV-6-positive cells (also recognized as CK19 [38]), which are rare in normal livers, also expressed CD133, CD44, and EpCam. Furthermore, most of the CD44-positive cells expressed the hepatic oval cell marker OV-6 (Supplementary Fig. S4).
We isolated and characterized the new cell population in the precancerous, changed livers using flow cytometric cell sorting with hepatic nonparenchymal cells and the surface markers CD133 and CD44 after hematopoietic cells were removed with antibodies specific for CD45 and HIS49 (Fig. 2A). One week after PH, the percentage of CD133+CD44+CD45−HIS49− cells was 1.00%±0.61% in livers from rats treated with 2-AAF and subjected to PH, which was significantly higher than that observed in normal rat livers (0.05%±0.03%, P<0.01). We determined the cell population that was hepatic precancerous cell-enriched by assessing the gene expression profiles of perfused total liver cells, liver parenchymal cells, liver nonparenchymal cells, and the subpopulations isolated using flow cytometry (CD133−CD44−CD45−HIS49−, CD133+CD44−CD45−HIS49−, and CD133+CD44+CD45−HIS49−; Fig. 2A). Compared with nonparenchymal liver cells, CD133+CD44+CD45−HIS49− cells had significantly higher expression levels of oval cell markers such as Afp, Krt19, Dlk1, Epcam, Gstp1, and Gpc3 (Fig. 2B). Furthermore, immunocytochemical examination of freshly sorted cells indicated that >90% of the CD133+CD44+CD45−HIS49− cells coexpressed the hepatic oval cell markers EpCam and OV-6 at significantly higher levels than all other liver cell populations (P<0.01; Fig. 2C, D and Supplementary Fig. S5A).
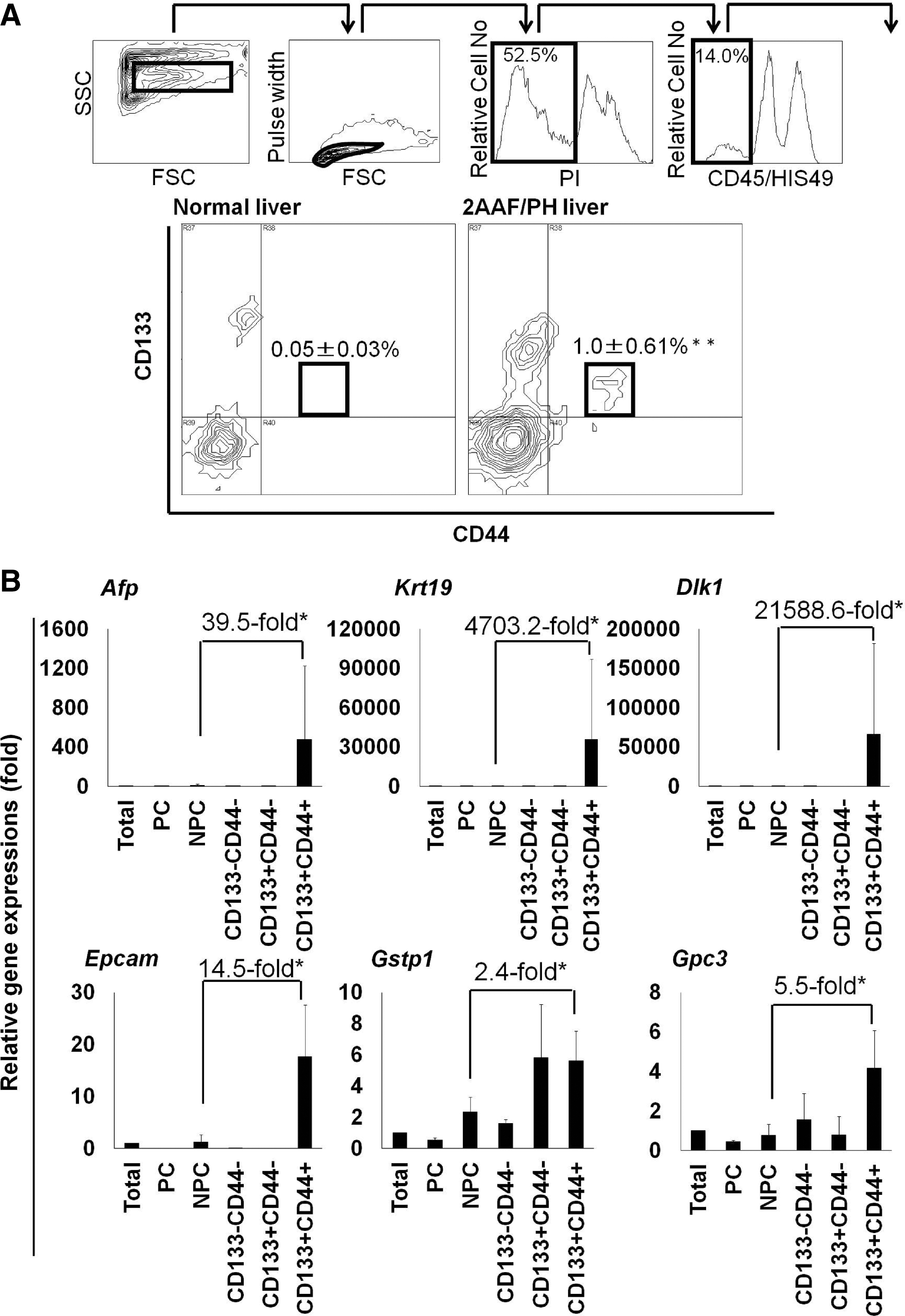
Isolation of hepatic precancerous cells among oval cells.
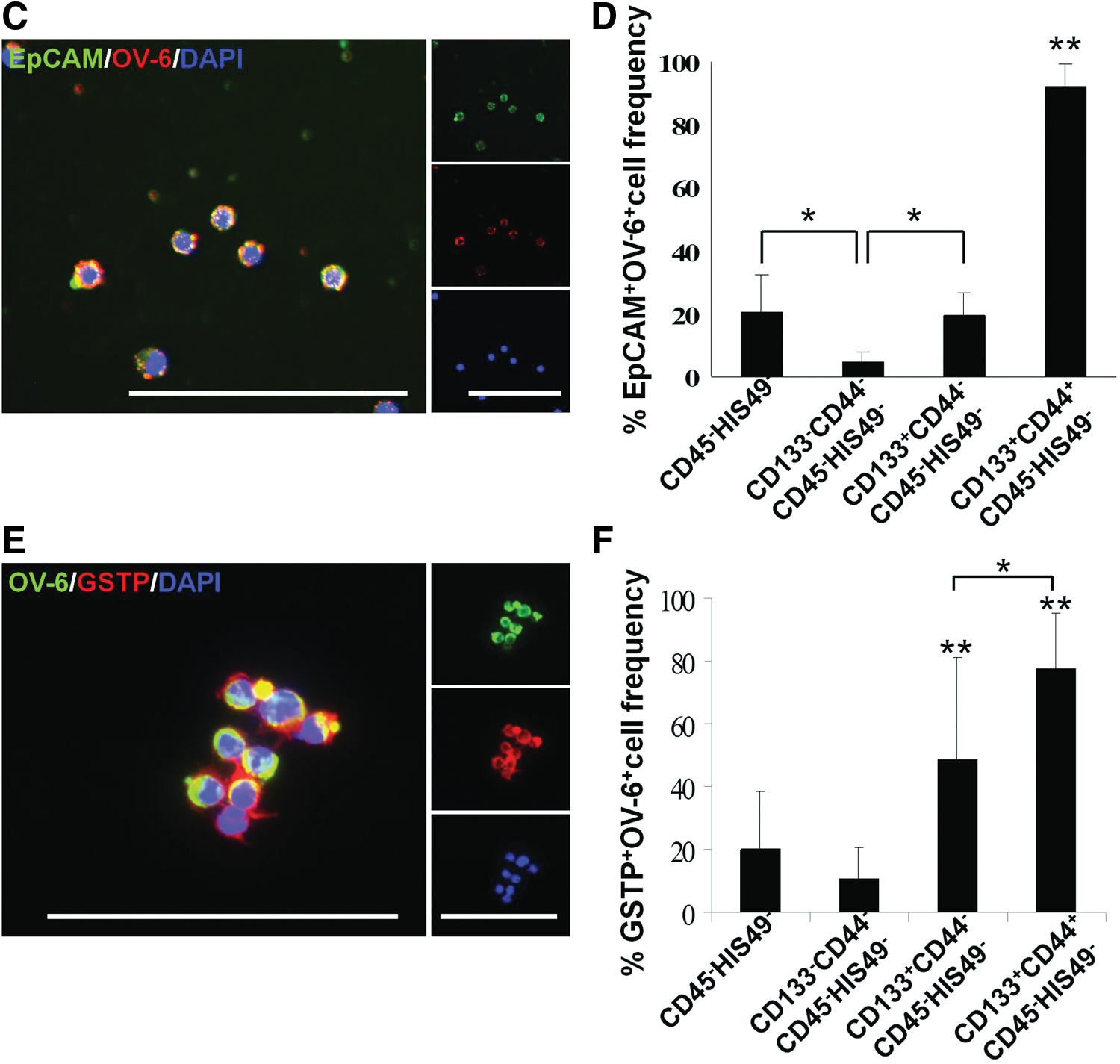
Cells positive for EpCam, CK19, and/or OV-6 were also detected in CD133+CD44+CD45−HIS49− cell-derived colonies (Supplementary Fig. S5B), which indicated that hepatic oval cells were present in this cell population. Cells with the capability to form clonal colonies (representative cell tracing results are shown in Fig. 3A) in in vitro cell culture at a significantly high frequency were restricted to the CD133+CD44+CD45−HIS49− subpopulation (P<0.01; Fig. 3B). Immunocytochemistry showed that the clonal colonies also contained albumin- and CK7- or CK19-positive cells, in addition to cells that were positive for both proteins, which suggested that the cells could differentiate into at least 2 cell types (Fig. 3C).

Stem cell characteristics and in vitro proliferative inhibition of precancerous cells. Freshly isolated CD133+CD44+CD45−HIS49− oval cells after 6 days (or 7 days) in culture.
Immunocytochemical examination also showed that GSTP-positive cells were enriched at a high frequency in the freshly sorted CD133+CD44+CD45−HIS49− subpopulation (P<0.05; Fig. 2E, F and Supplementary Fig. S5C) and in the single cell-derived colonies after 1 week of culture (Fig. 3D). Thus, GSTP-positive precancerous cells were enriched in and were developed from CD133+CD44+CD45−HIS49− cells, which showed characteristics of hepatic oval cells.
Expansion inhibition of hepatic precancerous cells in vitro
We investigated whether hepatic precancerous cells could be inhibited in vitro after treatment with the acyclic retinoid peretinoin because this drug has been reported to suppress HCC [26,27], and to verify the relationship between oval cells and HCC [39 –41]. In the present study, we found that GSTP-positive precancerous cells comprised a small subpopulation of oval cells. To evaluate the effects of acyclic retinoid on cell expansion and the colony formation capabilities of isolated hepatic precancerous cells in vitro, we performed a single cell-based assay, as described previously [33,34]. Phase-contrast images showed that the acyclic retinoid markedly inhibited the clonal expansion of CD133+CD44+CD45−HIS49− cells (Fig. 3E). After 6 days of culture, the hepatic precancerous oval cell-derived colonies in the group treated with the acyclic retinoid were half the size of the control colonies (49.3%±9.1%, P<0.05; Fig. 3F), and the efficiency of clonal colony formation was also significantly lower than that of controls (43.9%±6.4%, P<0.05; Fig. 3F). These data showed that the acyclic retinoid inhibited the stem-like characteristics of hepatic precancerous oval cells in vitro, including clonal expansion and colony formation.
Inhibition of the development of hepatic oval cells and precancerous cells in vivo
Immunohistochemical experiments were performed to examine the effects of the acyclic retinoid during the initial stages of hepatic oval cell generation and precancerous subpopulation formation in vivo. Significantly fewer oval cells expressing CK19 were detected on day 7 in the groups that received the acyclic retinoid (Fig. 4A). Peretinoin at a dose of 60 mg/kg or 80 mg/kg significantly reduced the percentage of CK19+ oval cells compared with the control group (P<0.01 for both comparisons; Fig. 4B). Reduction in oval cells led to a decline in the subpopulation of GSTP-positive precancerous cells, while more importantly, the precancerous cell subpopulation frequency decreased (P<0.01; Fig. 4C, D). Quantitative PCRs for the oval cell-related genes Afp, Krt19, Dlk1, Epcam, and Gpc3 showed that the expression levels of these genes tended to decrease with increasing peretinoin doses. The control group had significantly higher Epcam and Gpc3 expression levels compared with the peretinoin-treated groups (P<0.01) and significantly higher expression levels of Krt19 compared with the group treated with 80 mg/kg peretinoin (P<0.05; Fig. 4E).
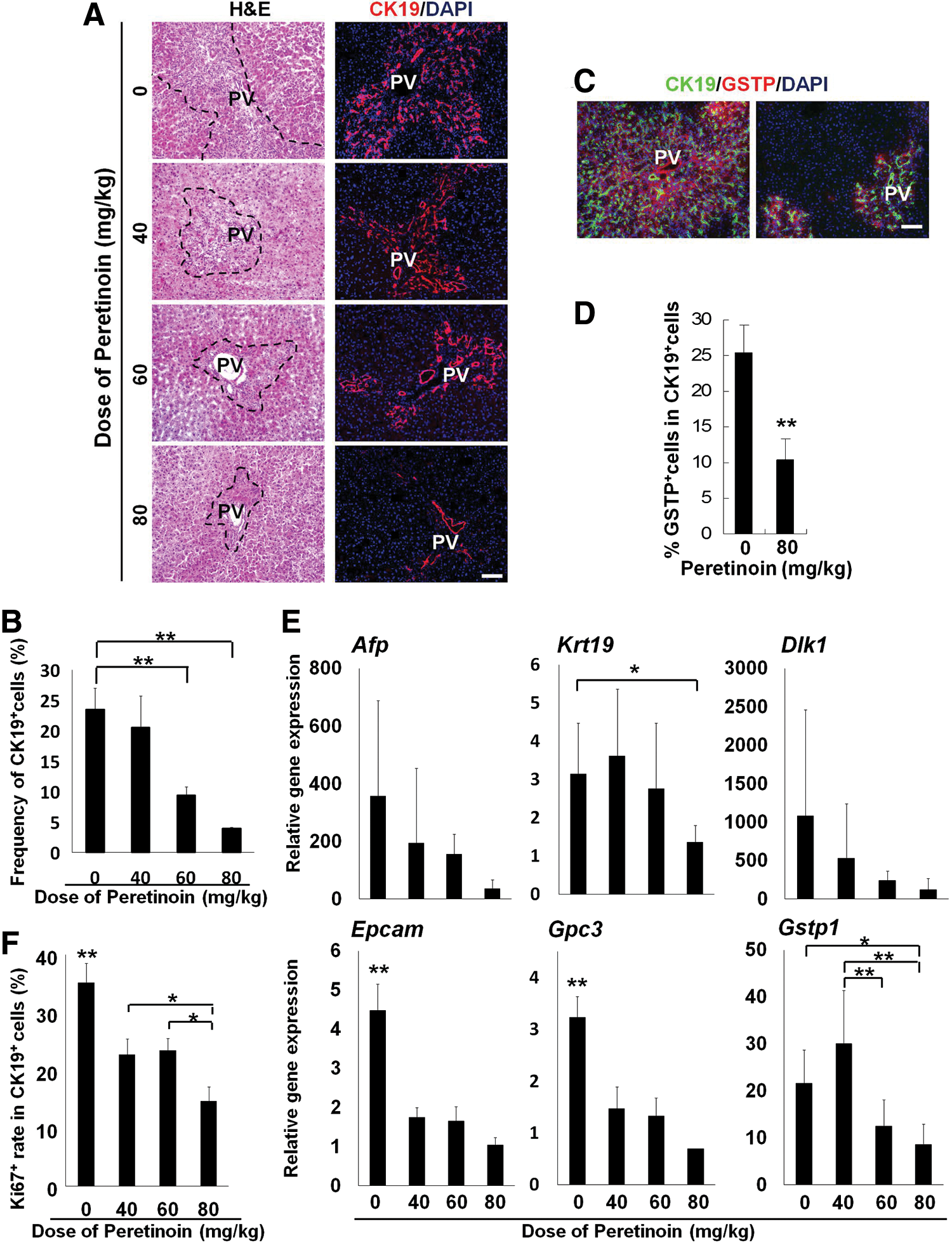
Inhibition of the development of GSTP-positive precancerous oval cells in vivo. Rats were examined on day 7.
To examine the effects of the acyclic retinoid on oval cell proliferation after PH, tissues were stained with antibodies specific to Ki67 and CK19. Fewer proliferating oval cells were detected following the acyclic retinoid (peretinoin) treatment (Fig. 4F and Supplementary Fig. S6). Immunohistochemical analysis showed that the percentage of Ki67-positive oval cells relative to the total number of oval cells expressing CK19 was reduced at all peretinoin doses after 7 days (P<0.01; Fig. 4F).
GSTP-positive foci have been reported as a marker of precancerous changes in the liver [35 –37]. Our results suggested a relationship between GSTP-positive cells, foci, and oval cells. Indeed, the oval cell subpopulation could be the source of GSTP-positive foci. Immunohistochemical analysis showed that peretinoin at doses of 40–80 mg/kg significantly reduced the number of GSTP-positive foci on day 42 (P<0.01 compared with control samples; Fig. 5A, B). Quantitative PCR analyses showed that peretinoin (80 mg/kg) significantly reduced the Gstp1 mRNA expression levels (P<0.05 vs. control group; Fig. 4E). The proliferative activity of GSTP-positive foci in the Ki67-positive cell assay was also significantly reduced by peretinoin treatment (P<0.01; Fig. 5C, D). These data suggest that the acyclic retinoid peretinoin inhibits oval cell proliferation and development at an early stage, thus suppressing the generation of oval cell progeny and precancerous cells in vivo.
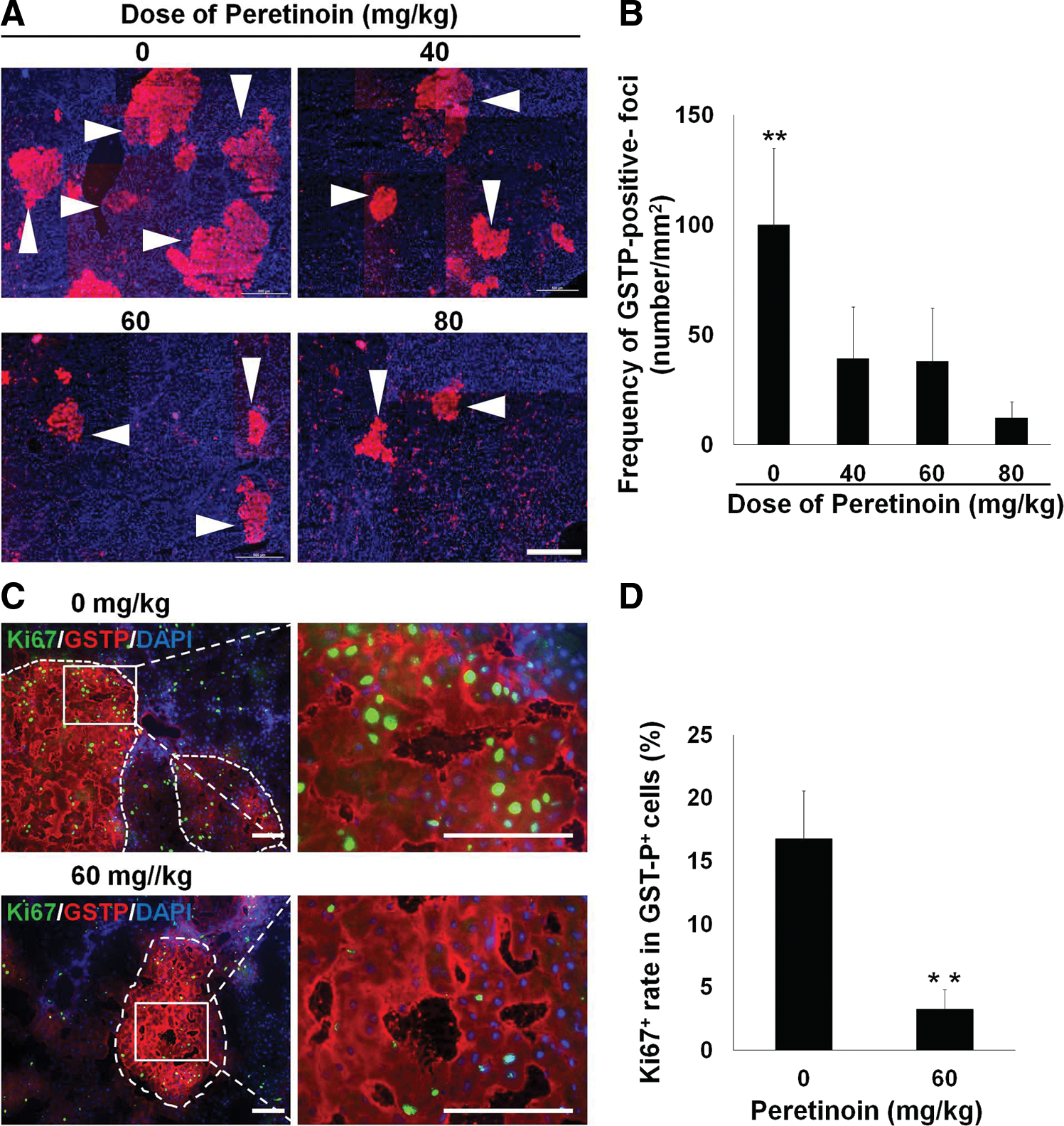
Therapeutic blocking precancerous oval cell also inhibits the development of GSTP-positive foci and HCC. Rats were examined on day 42
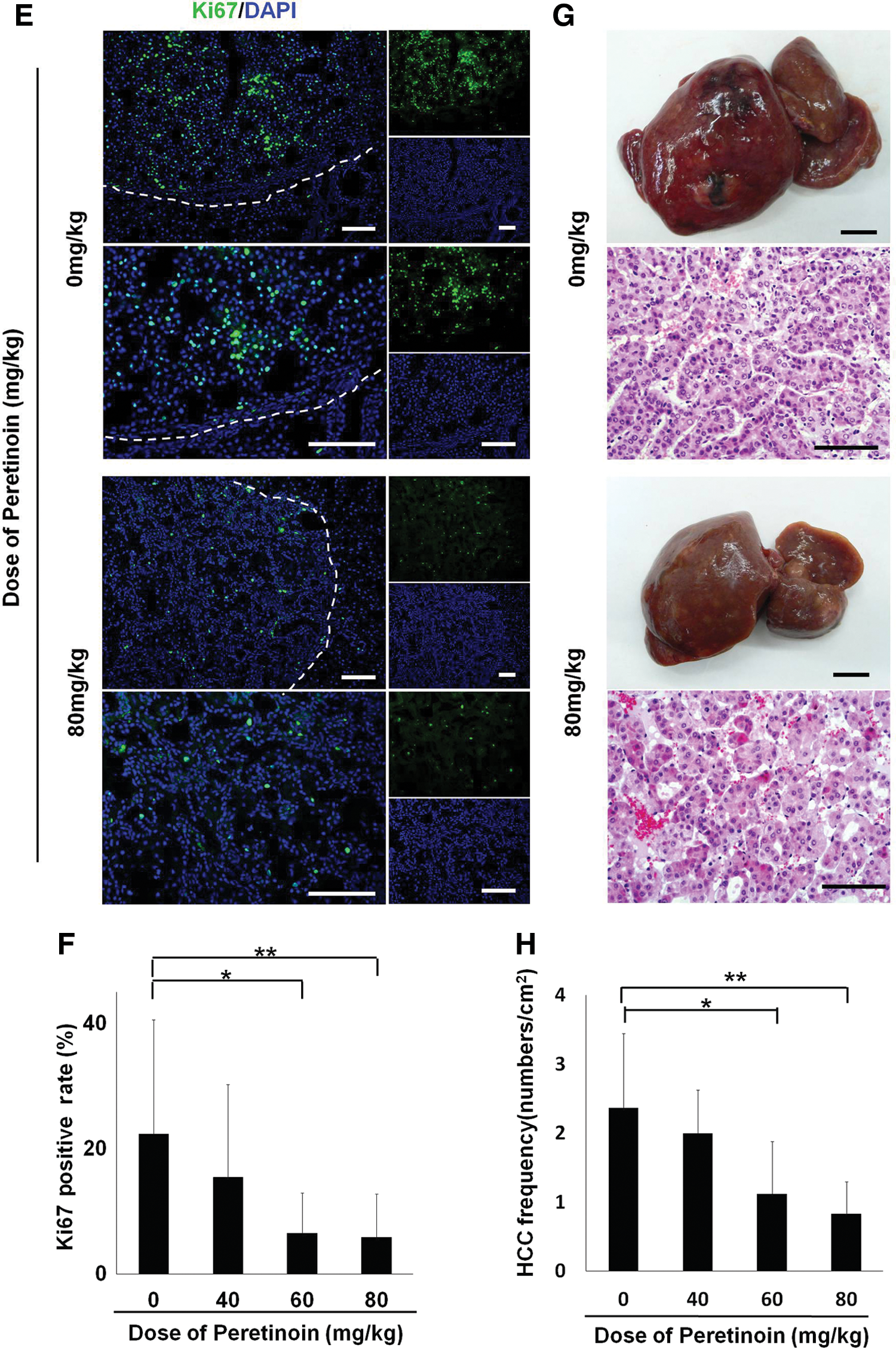
Direct suppression of precancerous cells also inhibits HCC in vivo
To further assess the consequences of GSTP-positive precancerous cell inhibition, we conducted long-term observations for 240 days. Immunohistochemical analysis of Ki67 showed that the number of proliferative liver cells was significantly reduced by treatment with the acyclic retinoid peretinoin at doses of 60 mg/kg or 80 mg/kg (P<0.01; Fig. 5E, F). HCC regions were identified on the basis of pathological diagnoses (Fig. 5G). Compared with the control group, the peretinoin-treated groups showed dose-dependent reductions in the number of HCC regions per unit area (Fig. 5H). There were significant differences at doses of 60 mg/kg (P<0.05) and 80 mg/kg (P<0.01). These data suggest that the generation of hepatic oval cells is inhibited by the acyclic retinoid, including the subpopulation of precancerous cells. Thus, HCC cell proliferation is reduced, which suppresses HCC formation.
Discussion
A comprehensive understanding of the origin of HCC is required to develop clinical strategies that target precancerous cells and prevent recurrence after initial therapy. Oval cells have been described as multipotent stem/progenitor cells that can give rise to hepatocytes and cholangiocytes. They have also been shown to replicate after liver injury and reside in the canals of Hering [42]. Previous reports suggest that HCC might originate from CK19-positive progenitor cells [43]. The number of oval cells increases with the severity of human chronic liver disease [44], which supports a causal role for oval cells in hepatocarcinogenesis. However, direct evidence for this relationship is still lacking. Indeed, most studies have relied on phenotypic analyses or analyses performed at a single time point [39 –41]. Furthermore, in our experiments, oval cells could be induced to emerge in all model animals but not in all the animals suffering from HCC disease. Only a small subpopulation of oval cells expressed the precancerous marker GSTP, which suggests that some event had occurred to transform a subpopulation of oval-like cells so that they had the appearance of precancerous cells, which could in turn increase the risk of their developing into HCC cells. To demonstrate the capability of precancerous cells to proliferate, differentiate, and transform, we combined in vitro precancerous cell isolation and characterization with in vivo dynamic tracing of cell fate to obtain relevant supporting evidence.
In the present study, liver carcinogenesis was traced and examined to identify the cellular origins of hepatic precancerous cells and HCC. GSTP-positive foci localized with oval cells in the animal model used in the present study, which demonstrated the link between oval cells and HCC development. GSTP-positive foci have been associated with hepatic precancerous changes [35], and the degree of hepatic precancerous change correlates with the incidence of HCC [45]. Furthermore, we found that GSTP-positive cells can be isolated from nonhematopoietic fractions containing hepatic oval cells that express CD133+CD44+, whereas GSTP-positive foci more frequently overlapped or were adjacent to oval cells. We also noted that only some of the hepatic oval cells expressed GSTP, and that the GSTP-positive cells expanded over time both in vivo and in vitro. The acyclic retinoid dose-dependently inhibited oval cell expansion and the subpopulation of precancerous cells in the livers of rats treated with 2-AAF and subjected to PH of liver tissue, which reflected the reduced numbers of GSTP-positive foci and HCC regions. Overall, our data show that GSTP-positive cells and foci are associated with hepatic oval cells and HCC occurrence. These results may have clinical significance for HCC if these precancerous cells or their progeny also can be found in humans, and thus therapeutically targeted.
The acyclic retinoid was shown to reduce the rate of HCC recurrence in clinical trials [26,27,31]. However, the therapeutic targets and mechanisms of action of acyclic retinoids remain unclear. Clonal deletion refers to the removal of latent malignant (or premalignant) cells from an organ in a hypercarcinogenic state [46,47]. Our results suggest that the therapeutic effects of the acyclic retinoid may reflect the deletion of precancerous cells. Acyclic retinoids suppress the clonal expansion of the hepatic oval cell subpopulation, which may further inhibit precancerous changes and prevent de novo carcinogenesis. Indeed, the acyclic retinoid inhibited the clonal expansion of CD133+CD44+CD45−HIS49− cells in vitro and the expression of stem or oval cell markers in vivo. As expected, a follow-up study based on our data showed that peretinoin inhibited HCC formation.
In a phase II/III clinical trial, the acyclic retinoid peretinoin significantly reduced the risk of HCC recurrence and death compared with placebo in patients with Child–Pugh grade A impairment and a major tumor diameter <20 mm prior to curative therapy, although there was no significant improvement in the study population as a whole [48]. These data suggest that the acyclic retinoid provides early benefits in preventing HCC recurrence in patients with preserved liver function who have undergone curative therapy for small tumors. HCC can arise from multiple cellular populations, and current therapies have limited efficacy for small tumors. Thus, our results suggest a possible effect of the acyclic retinoid peretinoin during the precancerous stage of liver cancer development, which mainly comprises precancerous cells and their progeny.
However, a recent report demonstrated distinctly different HCC progenitors from the oval cell-derived precancerous cells suggested in this study. The cells residing in dysplastic foci in DEN-mice for 3 months only developed into cancer cells when transplanted in a certain liver environment [49]. Although the cells shared several markers with hepatic stem cells/oval cells, the original cells were most likely derived from dedifferentiated hepatocytes in zone 3. These findings suggest multiple sources of HCC. The initiation of liver tumor formation is a very complex area of study, made even more complex by the fact that the markers, cell types, and initiating factors, and the microenvironment, considerably differ between different rodent models and human tumors. Furthermore, the effects on rodent oval cells do not necessarily translate directly into effects on human hepatic progenitor cells. Thus, conclusions based on rodent models should be interpreted with caution when extrapolating mechanisms to describe tumorigenesis in humans. Future clinical trials should examine cells in patient specimens directly when evaluating the prevention of HCC recurrence using the acyclic retinoid. If precancerous cells can be detected and evaluated prior to treatment during clinical practice, it may be possible to increase the confidence associated with achieving a specified exposure level to a medicinal product, such as the acyclic retinoid peretinoin, with desired therapeutic outcomes in patients. In conclusion, it is clear that a subpopulation of hepatic oval cells is precancerous and that these cells can lead to the development of HCC. Suppression of precancerous cell development by the acyclic retinoid peretinoin in rats treated with 2-AAF and PH inhibited HCC formation. Thus, precancerous cells that represent a transformed subpopulation of oval cells can be considered as therapeutic targets of the acyclic retinoid peretinoin. HCC is a complex disease in humans; thus, multiple solutions are probably required. The acyclic retinoid peretinoin may prevent de novo HCC carcinogenesis by targeting precancerous cells and their progeny, thereby providing a potential avenue for treatment and prevention in patients with HCC.
Footnotes
Acknowledgments
We thank A. Tanaka and H. Sato for performing the FACS analysis, and T. Yazawa and Y. Nagashima for providing diagnoses of hepatic pathology. We also thank M. Kimura, K. Takiguchi, and H. Tanaka for experimental assistance. This work was supported by Grants-in-Aid (18591421, 20591531, and 23591872) for scientific research from the Ministry of Education, Culture, Sports, Science, and Technology (MEXT) of Japan, and grants for the Strategic Promotion of Innovative Research and Development (S-innovation, 62890004) from the Japan Science and Technology Agency (JST).
Author Disclosure Statement
The authors disclose that N. Ishibashi and M. Imajima are employees of Kowa Co. (Japan). The remaining authors have no conflicts of interest to disclose.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.