
Abstract
The embryonic stem cell (ESC) microenvironment can promote the proliferation of terminal cells and reduce the invasiveness of tumor cells. However, implanting ESCs directly in vivo can result in tumorigenicity, immune rejection after differentiation, and graft-versus-host reaction. Therefore, safety is very important in the clinical application of ESCs. We injected ESCs modified with a suicide gene into a leukemia mouse model through peripheral blood to observe the treatment effectiveness. In addition, according to the pre-test, we set the time point of differentiation after transplantation and then activated the suicide gene to kill the ESCs that we had initially implanted, hoping to avoid the risks mentioned earlier. Our results indicated that the body weight and survival rates of mice treated with an ESC microenvironment increased, and leukemic cells in peripheral blood and bone marrow decreased compared with untreated mice. There was no obvious teratoma in mice that received ESC therapy and induced the suicide gene at the proper time during the observation period, while an apparent teratoma was observed in the lungs of mice which received ESC therapy and never induced the suicide gene. Therefore, the ESC microenvironment can promote self-healing of the in vivo microenvironment. Inducing the suicide gene at the appropriate time can reduce or even avoid tumorigenicity and immune rejection after transplantation of ESCs in vivo and improve the safety of their clinical application.
Introduction
E
The microenvironment can induce ESCs to differentiate into various cells. The ESC microenvironment can also have a positive impact on cells in contact with it. The embryonic microenvironment has a strong positive influence on cells. As shown by the birth of the Dolly sheep, a mature breast cell in an embryonic environment can yield a new individual. Our previous studies also demonstrated that in ESC-conditioned medium, ESCs co-cultured with corneal epithelial cells or corneal endothelial cells could significantly promote the proliferation of terminal cells and even make them express surface markers of some precursor cells [18,19], which indicated that an ESC microenvironment could reverse the terminal differentiation of these cells. The ESC microenvironment can even influence the epigenetic effect of tumor cells. In one study, ESCs were cultured on an artificial basement membrane for 3–4 days; then, ESCs were removed and cultured melanoma cells on the matrix, and the melanoma cells were less invasive [20]. All these effects depend on the embryonic microenvironment provided by ESCs, which requires the ESCs to maintain an undifferentiated state. After differentiation, ESCs not only lose their microenvironmental therapeutic effect but also induce immune rejection due to the expression of other genes and surface antigens [21,22]. It is difficult to remove ESCs after transplanting them in vivo. Therefore, many studies on stem cell transplantation treat the recipient with immunosuppressive drugs, but once the recipient's immune system is suppressed, immune cells derived from ESCs may have advantages and lead to graft-versus-host reactions [21,23]. In addition, immunosuppression often results in increasing the risk of severe infection.
To both maximize the therapeutic effect of ESCs and avoid tumor formation, immune rejection, graft-versus-host reactions, and other defects after transplantation, our study attempted to inject ESCs modified with a suicide gene [thymidine kinase (TK)] into a mouse leukemia model and observed the influence of the embryonic microenvironment on the progression of the leukemia. Moreover, when ESCs began to differentiate and the ESC microenvironment weakened, the suicide gene was induced to kill the ESCs and cells derived from the ESCs, and then fresh undifferentiated ESCs were injected into the body to sustain the embryonic microenvironment. The treatment was examined for avoiding the adverse effects mentioned earlier.
Materials and Methods
Animals and cells
Severe combined immunodeficient (SCID) mice (female, aged 6 weeks and weighing 20 g) were bought from the company of Vital River (Beijing, China). All animal experiments were performed with permission from the Medicine Ethics Committee of the Zhongshan Ophthalmic Center, Sun Yat-Sen University.
Mouse ESC-E14s were generously provided by Professor P. Xiang in Sun Yat-Sen University, China [24]. ESC-E14s were plated at a density of 400/cm2 in 0.1% gelatin-coated tissue culture dishes (Sigma, St. Louis, MO) containing mouse ESC culture medium (Invitrogen, Carlsbad, CA).
The human K562 leukemia cell line was bought from American type culture collection and suspended in bacterial culture dishes.
Constructing stable transfected ESC-GFP-TK
The construction of ESC-GFP-TK was according to the standard procedure [25]. Using the Gateway recombination reaction, pUp-promoter (EF1α) and pDown-TK or pDown-green fluorescent protein (GFP) were recombined into the pDest-puromycin vector and pDest-blasticidin following the protocol for the LR recombination reaction to construct lentiviral expression vectors designated EF1α-TK and EF1α-GFP.
The lentiviral particles were produced by transient cotransfection of 293FT cells with the expression lentiviral vectors mentioned earlier and the ViralPower™ Lentiviral packaging mix (Invitrogen) using transfection reagent (Roche, Indianapolis, IN). The supernatant containing lentivirus was harvested at 72 h after transfection and concentrated by ultracentrifugation (50,000 g for 120 min at 4°C).
For lentiviral transduction, dissociated ESC clumps were transduced in suspension with lentivirus at a multiplicity of infection of 50 and then plated onto 0.1% gelatin-coated plates in knockout medium with 5 μg/mL polybrene (Sigma). Three days after lentivirus transduction, 1 μg/mL puromycin or 10 μg/mL blasticidin (Sigma) was added to the culture medium and maintained for 5 days.
Immunofluorescence staining
The phenotype of the ESCs was determined by immunofluorescence staining according to the standard procedure [18]. For immunofluorescence staining, undifferentiated ESCs were fixed with 4% paraformaldehyde for 20 min, and then permeabilized with 0.1% Triton X-100, following blocking of nonspecific binding with goat serum. Then, the cells were incubated with primary antibody overnight at 4°C. After washing with phosphate-buffered saline (PBS), the fluorescence-conjugated secondary antibody was added for 40 min at room temperature. The nucleus was counterstained with Hoechst 33258 (blue; Invitrogen) for 15 min. The primary antibodies used were rabbit polyclonal anti-OCT4 Ab (1:100; Abcam, Cambridge, United Kingdom) and mouse monoclonal anti-SSEA1 Ab (1:100; Abcam). The secondary antibody used was Alexa Fluor-conjugated goat-anti mouse lgG (green and red; Invitrogen). An examination was performed with a laser scanning confocal microscope (LSM 510 META; Carl Zeiss, Hamburg, Germany). Cells incubated with PBS instead of the primary antibody were used as negative controls.
Western blotting
Expression of OCT4 and TK was detected in ESCs and ESC-GFP-TKs using western blotting according to the standard procedure [18]. In short, cells were washed twice with PBS, scraped off using a cell scraper, and homogenized in sample buffer [60 mM Tris, pH 6.8, 2% (w/v) sodium dodecyl sulfate, 100 mM 2-ME, and 0.01% (w/v) bromophenol blue] for 30 min on ice. The homogenate was centrifuged at 16,000 rpm for 25 min. The concentration was checked by a Bio-Rad protein assay kit (Bio-Rad, Mississauga, Canada). Total protein of 50 μg was loaded to a lane of 12% polyacrylamide gradient gel, and electrophoresis was performed for 40 min. Then, the gel proteins were transferred to PVDF membranes (Millipore, Billerica, MA) for 1 h at 100 V. Membranes were blocked with 5% nonfat dry milk in PBS containing 0.1% Tween-20 for 1 h at room temperature, then incubated with primary antibodies at 4°C overnight. After washing three times with Tris-Buffered-Saline with Tween (TBST), membranes were incubated with horseradish-peroxidase-linked secondary antibody for 2 h at room temperature and washed with TBST. Localization of antibodies was detected by chemiluminescence using ECL kit (Amersham, Piscataway, NJ) following the manufacturer's instructions. The primary antibody used was mouse anti-OCT4 (1:5,000) and mouse anti-TK (1:5,000). β-Actin (1:3,000; Cell signaling, Billerica, MA) was used as a positive control.
Tumor formation in SCID mice
ESCs and ESC-GFP-TKs (1×106) were injected subcutaneously into the hind leg of six SCID mice. Tumor growth was examined every week, and a tumor was selected and sliced for an analysis of its components at 5 weeks after an injection.
Flow cytometry analyses
Apoptosis analyses
We used allophycocyanin (APC; Invitrogen) and propidium iodide (PI; Invitrogen) to evaluate the relationship between the proliferation of ESC-GFP-TKs and the ganciclovir (GCV) concentration (0, 0.01, 0.1, 1, 10, 100, and 1,000 μg/mL). Data analysis was conducted using CellQuest software (BD Biosciences, Mountain View, CA).
Analyses of OCT4 expression
To determine OCT4 expression in ESC-GFP-TKs, flow cytometry was performed as previously described [26]. In short, ESC-GFP-TKs were fixed with Reagent A (Fixation Medium; Molecular Probes, Eugene, OR) for 15 min, and then permeabilized with Reagent B (Permeabilization Medium; Molecular Probes) for 15 min. The cells were incubated with primary antibody for 30 min at room temperature. After washing with PBS, the fluorescence-conjugated secondary antibody was added for 30 min at room temperature. The primary antibody used was a rabbit anti-OCT4 Ab (1:50; Abcam). The secondary antibody used was Alexa Fluor-conjugated donkey anti-rabbit IgG (1:100, red color; Invitrogen). The final suspension was analyzed using an FACSCalibur flow cytometer (BD Biosciences), and the results were analyzed using CellQuest Pro software (BD Biosciences).
MTT proliferation assay
The ESCs and ESC-GFP-TKs were seeded at 300 cells/well in 96-well plates, and cell proliferation curves were generated according to the optical density (OD) value measured at 490 nm during 7 days (Power Wave XS; Bio-Tek, Winooski, VT). All samples were assayed in five duplicates.
To further determine the sensitivity of GCV to ESC-GFP-TKs, we used the MTT assay to assess the relationship between the proliferation of ESC-GFP-TKs and the GCV concentration. ESC-GFP-TKs were seeded at 300 cells/well in 96-well plates. Twenty-four hours after seeding, the cells were treated with different concentrations of GCV (0, 0.01, 0.1, 1, 10, 100, and 1,000 μg/mL). Untransfected ESCs were used as controls. The average number of cells was counted, and a cell proliferation curve was generated according to the OD measured at 490 nm (Power Wave XS; Bio-Tek). All samples were assayed in five replicates.
Developing a leukemia mouse model
A leukemia mouse model was developed using the human leukemia K562 cell line [27,28]. K562 cells (1×106) were injected into 6-week-old SCID mice through the tail vein after 2 Gy irradiation.
Animal experiment
Four weeks after the injection of K562 cells, the mice were divided into the untreated group and the ESC+GCV group according to body weight. All animals were divided into the following three groups: (1) the blank control group (normal SCID mice), (2) the untreated group (leukemia mouse model without treatment), and (3) the ESC+GCV group (leukemia mouse model with ESC-GFP-TKs and GCV treatment). The ESC+GCV group received ESC-GFP-TKs on the first and fourth days and received GCV on the 11th and 12th days every 12 days.
We recorded the changes in body weight, peripheral blood, and bone marrow in each group during the observation period.
Statistical analysis
All values are presented as the mean±SE. All statistical analyses were performed with SPSS software version 13.0 using one-way analysis of variance, and a probability value of 0.05 was considered statistically significant.
Results
Phenotype of mouse ESCs
The morphology of the mouse ESCs was a small, bright, round, high nucleoplasm ratio under a light microscope. Immunohistochemistry indicated that the ESCs expressed the stem cell markers OCT4 and SSEA-1. Approximately 99.8% of ESCs expressed OCT4 as evaluated by flow cytometry. Western blotting showed the expression of OCT4 in ESCs (Fig. 1).
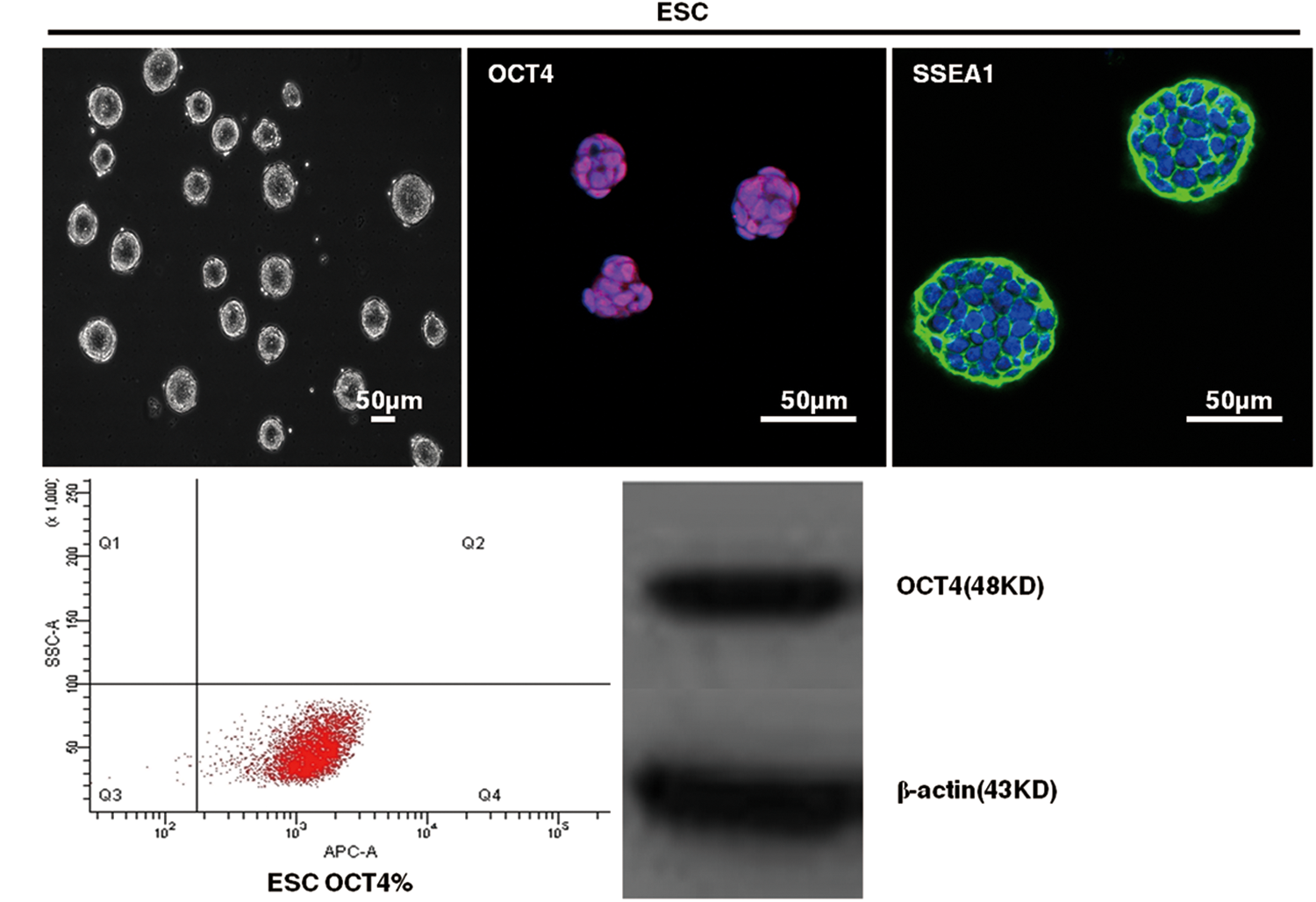
Phenotype of mouse embryonic stem cells (ESCs). Representative images of morphology by phase microscopy and the expression of OCT4 and SSEA1 in ESC by immunofluorescent staining. The expression of OCT4 by flow cytometry and western blot. Color images available online at
Generation of GFP- and TK-modified ESCs
Two plasmids encoding the TK and GFP genes were under the control of a housekeeping gene promoter (EF1α) (Fig. 2A). After puromycin and blasticidin selection, four purified, genetically modified cell lines were attained, which we called ESC-GFP-TKs. Individual colonies were picked and grown in culture for further analysis. Immunofluorescent staining and flow cytometry results demonstrated that ESC-GFP-TKs were successfully and stably transfected with GFP gene and ∼99.8% of ESC-GFP-TKs were GFP positive as evaluated by flow cytometry (Fig. 2B). Western blotting results showed that TK was expressed in ESC-GFP-TKs and not in ESCs, while β-actin level in ESCs was unaffected (Fig. 2C).
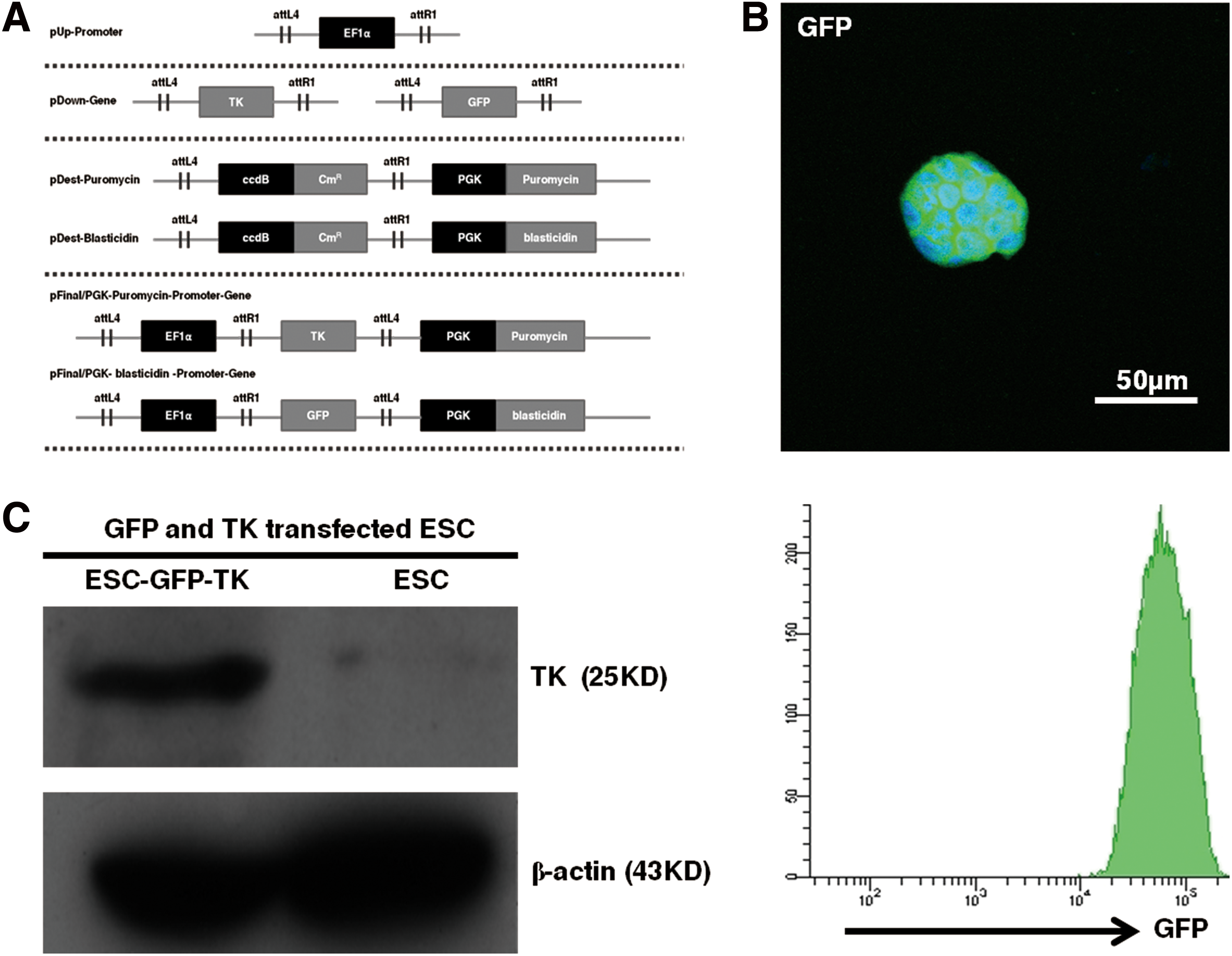
Generation of green fluorescent protein (GFP)- and thymidine kinase (TK)-modified ESCs.
Characteristics of ESC-GFP-TKs
ESCs transfected with the GFP and TK genes maintained high self-renewal capacity, high proliferation (Fig. 3A), and expressed the stem cell marker OCT4 by western blot, immunofluorescent staining, and flow cytometry (99.8%) (Fig. 3B, C). MTT proliferation assays showed that the proliferation rates of ESC-GFP-TKs (0.031±0.026, 0.063±0.026, 0.109±0.039, 0.283±0.079, 0.485±0.148, 1.575±0.29, 2.786±0.458) and ESCs (0.048±0.031, 0.079±0.018, 0.154±0.04, 0.306±0.048, 0.529±0.134, 1.216±0.213, 2.365±0.290) were not significantly different during 7 days (n=5, P>0.05) (Fig. 3D). When ESC-GFP-TKs and ESCs were injected into SCID mice subcutaneously, there was no difference in the time to tumor formation (9 days), tumor size (14.8±0.2 mm and 14±0.7 mm, respectively), or the components of the teratoma (containing of the three germ layers) (n=3, P>0.05), which was consistent with the results of the MTT proliferation assay. The teratoma derived from ESC-GFP-TKs expressed GFP under a confocal microscope (Fig. 3E).
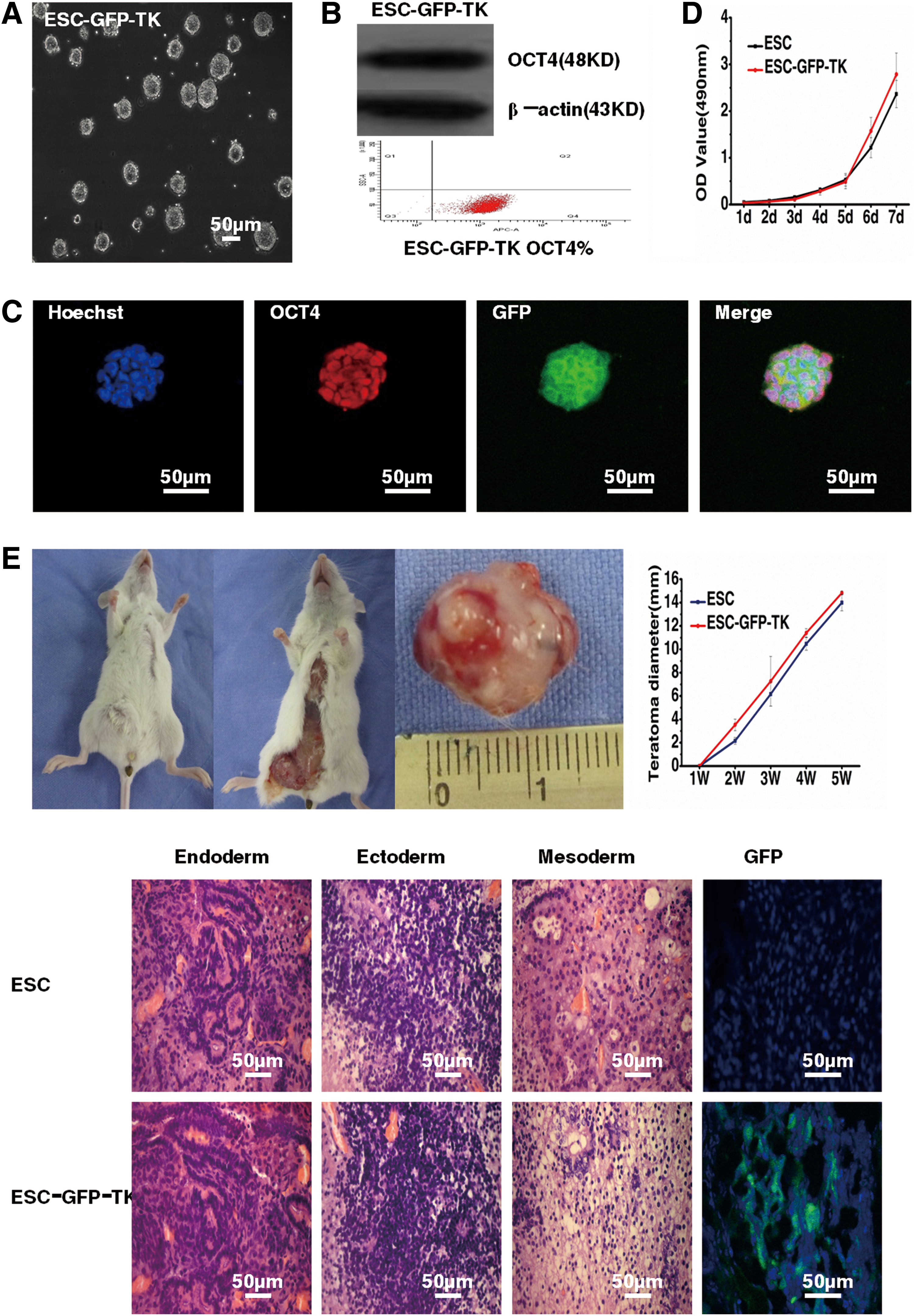
Characteristics of ESC-GFP-TKs.
Sensitivity of ESC-GFP-TKs to GCV
To assess their sensitivity to GCV, ESC-GFP-TKs were exposed to different concentrations of GCV (0, 0.01, 0.1, 1, 10, 100, and 1,000 μg/mL) for 2 days and then subjected to the APC/PI apoptosis assay. Untransduced ESCs were used as the control. The rates of apoptosis of the ESCs and ESC-GFP-TKs were 2.4%±0.1% and 4.4%±0.3%. Apoptosis of the ESC-GFP-TKs increased slightly to 10.9%±1.3% at a GCV concentration of 0.01 μg/mL. When the concentration of GCV was 0.1 μg/mL, the rate of apoptosis was 65.6%±2.1%. With increasing concentrations of GCV, apoptosis gradually increased. The viability of TK+ cells significantly decreased at a GCV concentration of 1 μg/mL (81.4%±5.5%), and almost no viable TK+ cells were found at 10 μg/mL (88.4%±0.5%), 100 μg/mL (92.2%±0.8%), or 1,000 μg/mL (94.3%±0.7%) on day 2 (Fig. 4A). These results indicated that GCV could induce TK+ cell apoptosis in a highly efficient and dose-dependent manner.
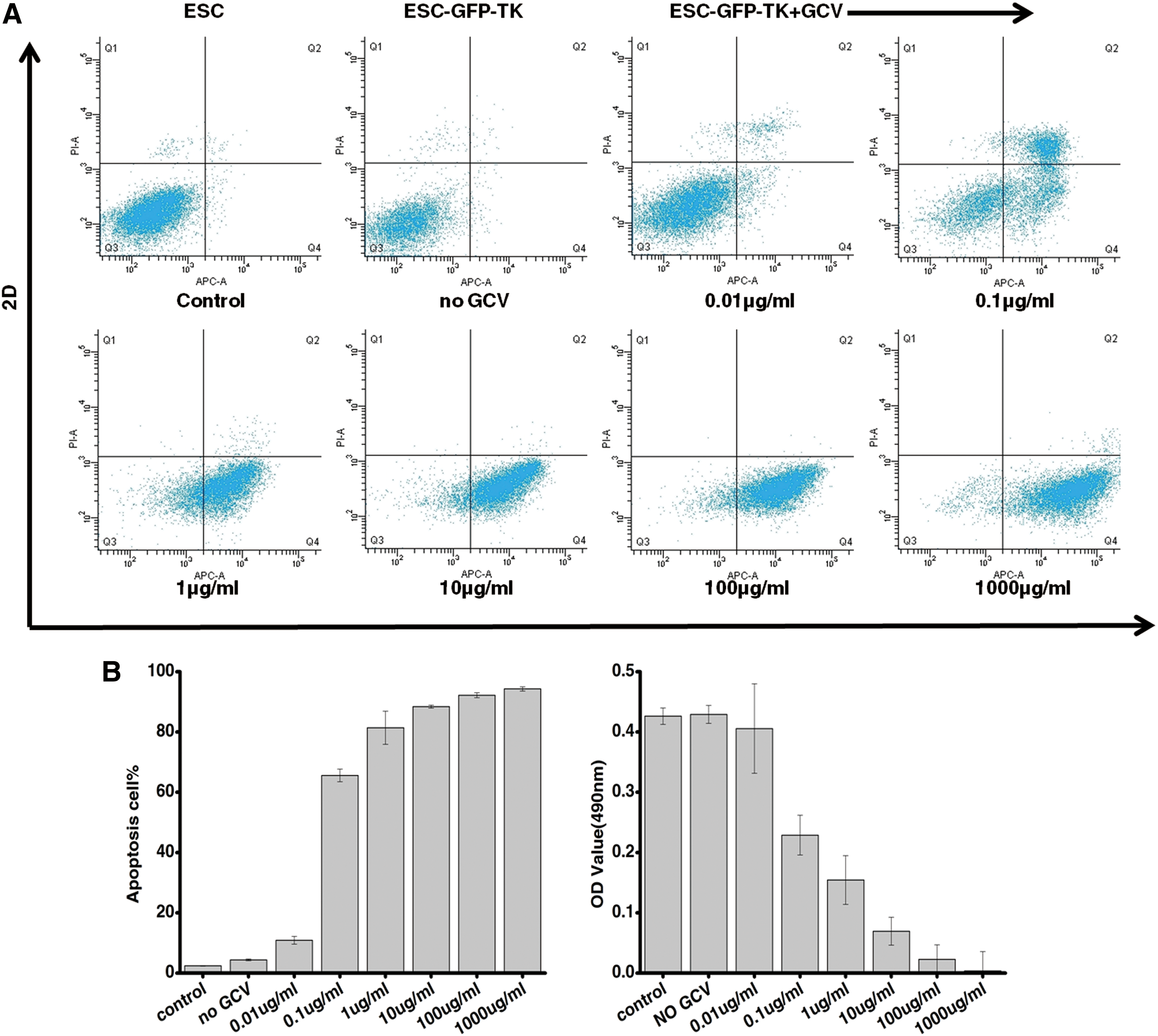
Sensitivity of ESC-GFP-TKs to ganciclovir (GCV) in vitro.
To further observe the sensitivity to GCV, we used the MTT proliferation assay to evaluate the relationship between ESC-GFP-TKs proliferation and the GCV concentration. The result showed that the OD values of ESCs and ESC-GFP-TKs were 0.426±0.014 and 0.429±0.015, and with increasing concentrations of GCV, the OD values gradually decreased. When the concentration of GCV was 0.01, 0.1, 1, 10, 100, and 1,000 μg/mL, the OD values were 0.406±0.074, 0.229±0.033, 0.155±0.04, 0.07±0.023, and 0.023±0.024, 0.004±0.03 respectively (Fig. 4B).
A leukemia mouse model generated by an injection of K562 cells
A leukemia mouse model was developed using the human leukemia K562 cell line, which exhibits significant atypia with varied morphology and a high nucleoplasm ratio compared with normal cells in peripheral blood and bone marrow (Fig. 5C). K562 cells (1×106) were injected into 6-week-old SCID mice through the tail vein after 2 Gy irradiation. Body weight was significantly decreased in mice injected with K562 cells (−3.9±1.6 g), while it increased in the controls (2±0.7 g) at the end of 4 weeks (Fig. 5A). Due to the pretreatment of the mice with 2 Gy irradiation, the mice exhibited significant bone marrow suppression in the first 3 weeks. The number of white blood cells in the peripheral blood dramatically increased in the leukemia mouse model (0.36±0.16×109, 0.4±0.13×109, and 8.16±4.23×109), while it remained unchanged in normal SCID mice (12.31±1.16×109) in the first 3 weeks (Fig. 5B). Nucleated cells were significantly increased in the bone marrow of leukemia mice, especially hematopoietic islands (Fig. 5D). In the fourth week, the white blood cell count in the peripheral blood of leukemia mice continued to rise (13.84±5.57×109) (Fig. 5B), and K562 cells could be found in the bone marrow of leukemia mice (Fig. 5D).

Development of a leukemia mouse model.
Evaluation of the effectiveness of ESC-GFP-TKs treatment
To evaluate the effectiveness of ESC treatment, SCID mice were divided into the following three groups: (1) blank control group (normal SCID mouse), (2) ESC+GCV group (leukemia mice receiving ESC-GFP-TKs treatment and induction of the suicide gene at the proper time), and (3) untreated group (leukemia mice without treatment).
The ESC+GCV group showed great improvement in general characteristics (Fig. 6A).

Evaluation of the effectiveness of ESC-GFP-TKs treatment.
Weight loss
After 1 week of ESC-GFP-TKs treatment, the ESC+GCV group had lost 1.5±0.6 g body weight, which was significantly less than the untreated group (2.6±0.9 g) (n=40, P<0.05). After 2 weeks of ESC-GFP-TKs treatment, the weight loss of the ESC+GCV group was 3.2±1 g and 4.1±1.1 g for the untreated mice. Although weight of both the ESC+GCV group and untreated group decreased, the weight loss of the untreated group was more obvious, but the difference was not statistically significant (n=20, P>0.05). After 3 weeks of treatment, the mice's weight continued to decrease, but the weight loss of the ESC+GCV group (3.7±1.2 g) was less than that of the untreated group (4.6±0.7 g), and the difference was not statistically significant (n=10, P>0.05). During the observation period, the weight of normal mice slightly declined (0.5±0.3 g) compared with the untreated group and the ESC+GCV group, and the difference in weight loss was statistically significant (n=10, P<0.05) (Fig. 6B).
Peripheral blood and bone marrow
The number of red blood cells was slightly reduced in untreated mice compared with ESC+GCV mice and normal mice. The proliferation of the bone marrow of untreated mice was obvious compared with ESC+GCV mice and normal mice, and a large number of leukemia cells were found in the untreated group. Bone marrow cells exhibited obvious atypia in the untreated group. The number of leukemia cells in the ESC+GCV group decreased in the bone marrow, proliferation of nucleated cells was reduced, and cell atypia was not obvious. The proliferation of bone marrow cells in both the ESC+GCV group and the untreated group was higher than in normal mice (Fig. 6C).
Survival rate
Compared with the untreated group (0%), the ESC+GCV group (12.5%) had a higher survival rate at 8 weeks, while the survival rate of normal mice was 100% (Fig. 6D).
In order to observe what happened to the mESC after an injection in vivo, we detected the percentage of OCT4 and the number of ESC-GFP-TK in mouse blood by flow cytometry. As shown in Figure 7 and Table 1, on the first day, we injected undifferentiated ESC-GFP-TK (1×106) in vivo; the OCT4% was 97.5%±0.4% and the cell number was 99.5063±0.2079(104) by flow cytometry. On the third day, the OCT4% declined to 2.9%±0.3% and the cell number was 198.9835±11.9551(104). Due to the decline in OCT4%, we injected ESC-GFP-TK (1×106) into mouse blood once more on the fourth day; the OCT4% increased to 24.4%±0.4%; and the cell number was 177.8729±12.0618(104). On the 7th day and the 10th day, the OCT4% was 2%±0.5% and 0.9%±0.2%; the cell number was 18.392±6.8087(104) and 9.6552±14.3983(104). On the 11th and 12th day, the mouse received GCV (1000 μg) from blood to induce apoptosis of ESC-GFP-TK. On the 13th day, the OCT4% was 0.1%±0.1% and the ESC-GFP-TKs were only 0.0805±0.05(104) left (Fig. 7A, B).
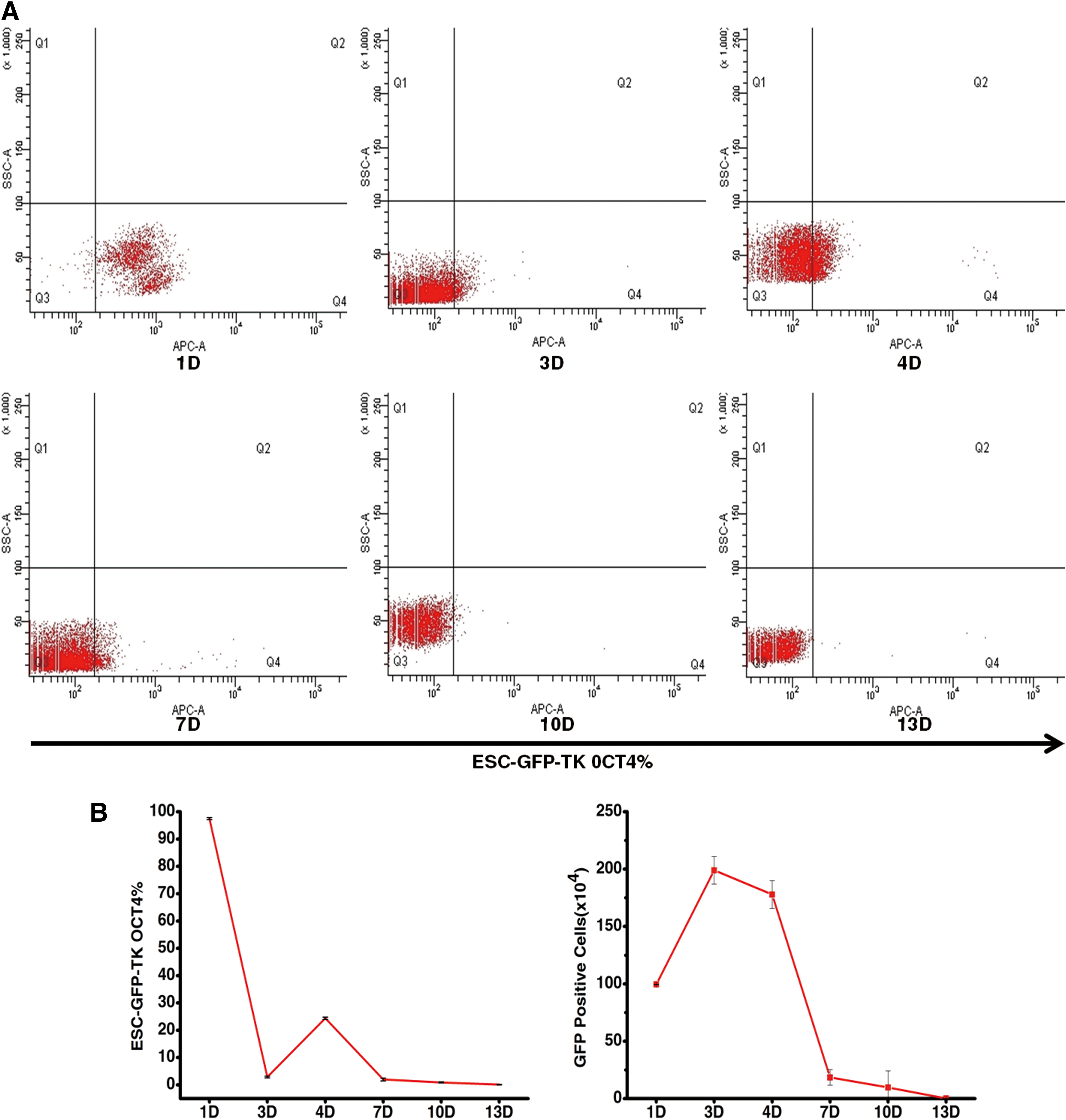
The percentage of OCT4-positive ESC-GFP-TKs and the number of ESC-GFP-TKs in mouse peripheral blood over 13 days.
ESC, embryonic stem cell; GFP, green fluorescent protein; TK, thymidine kinase.
Safety evaluation of ESC-GFP-TKs in vivo
Although the sensitivity of TK+ ESCs to GCV was shown in vitro, it is important to further evaluate the tumor-preventing function of the TK gene in murine models. For this purpose, ∼1×106 undifferentiated ESC-GFP-TKs in PBS were injected into 6-week-old SCID mice through the tail vein. Flow cytometry was used to detect apoptosis of ESC-GFP-TKs after inducing the TK suicide gene in peripheral blood. The result showed that before inducing the suicide gene, 6.8%±0.9% ESC-GFP-TKs underwent apoptosis. Two days after injecting GCV through the tail vein, apoptosis was induced in 94.5%±0.2% of ESC-GFP-TKs (Fig. 8A). We subsequently divided these mice into two groups: the ESC group, which only received ESC-GFP-TKs on the 1st and 4th days every 12 days, and the ESC+GCV group, which received GCV (1,000 μg) on the 11th and 12th days in addition to ESC-GFP-TKs on the 1st and 4th days every 12 days. Six weeks later, the mice that received GCV every 12 days did not exhibit an obvious fluorescence signal. In contrast, the group that did not receive GCV showed an obvious fluorescence signal. After 8 weeks, a teratoma could be found in the lungs of mice in the ESC group (8/10). The tumor in the lung was closely linked to the lung and contained three layers of germ tissue. GFP fluorescence could be observed under confocal microscopy, while there were no visible tumors in the ESC+GCV group (0/10) (Fig. 8B).
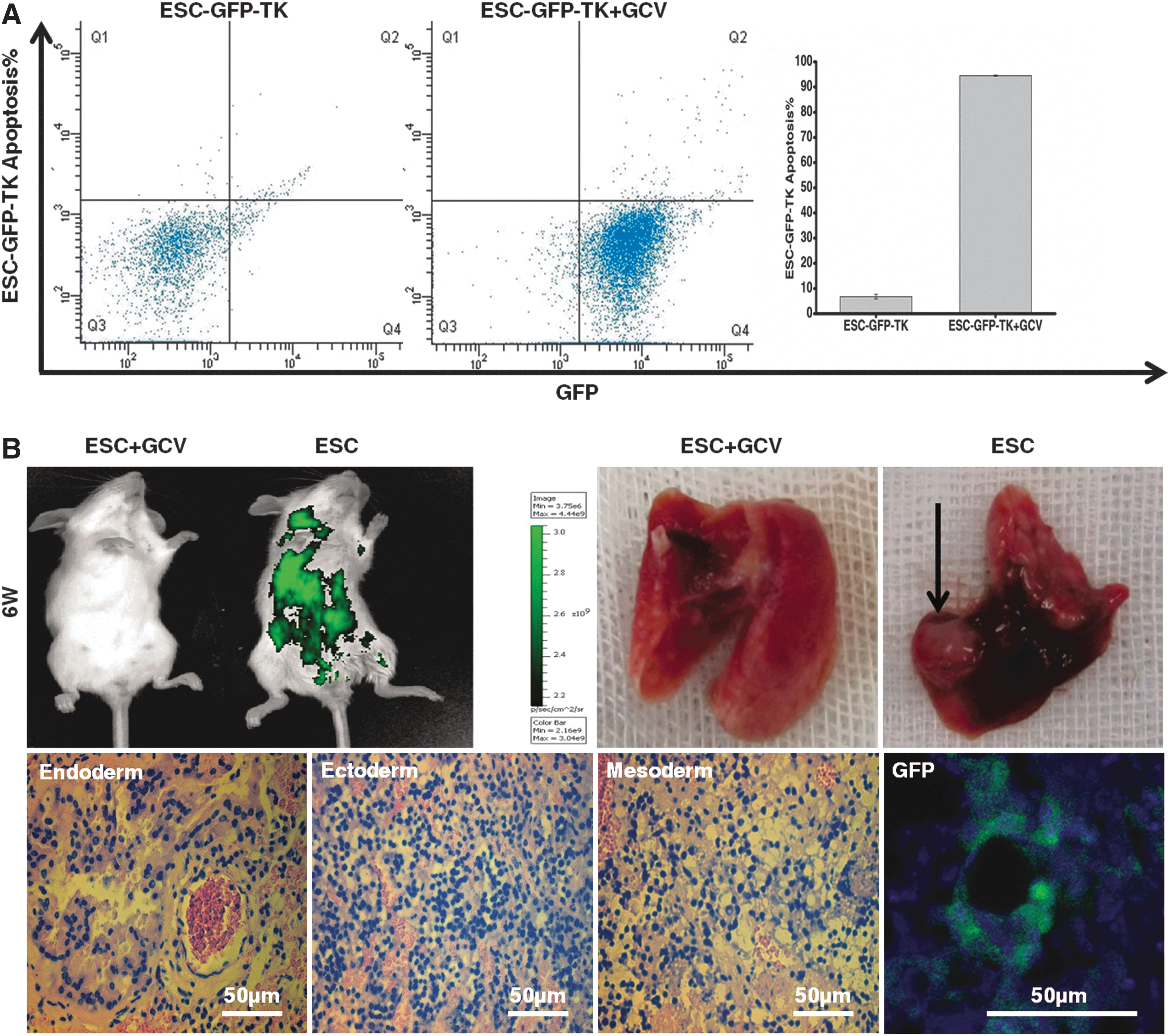
Safety evaluation of ESC-GFP-TKs in vivo.
Discussion
It is safe to use ESCs modified with TK for in vivo application
Safety is a prerequisite for the clinical application of stem cells. The major risks of ESC transplantation include tumor formation and immune reaction.
Reducing or avoiding the risk of tumorigenicity after transplantation
ESCs are undifferentiated totipotent cells that have the capacity to differentiate into all types of cells. Tumorigenicity is a manifestation of ESC pluripotency, but it is a major obstacle to clinical application. Although undifferentiated ESCs could differentiate into functional dopaminergic neurons after being injected into the striatum of a Parkinson mouse model, teratomas were found in 20% of those mice [16]. To reduce the tumorigenicity of ESCs in vivo, current research on stem cells often uses cells that were differentiated before an injection. First, ESCs are differentiated into specific progenitor cells to eliminate pluripotency, and then, progenitor cells derived from ESCs are implanted in the body to treat diseases. However, if the differentiation of ESCs is not complete, the risk of teratoma formation still exists. Moreover, the differentiated cells derived from ESCs often include cells that differentiated through other pathways. For example, ESCs were cultured on PA6 feeder-layer cells before in vitro introduction, and then differentiated cells were implanted into animals. In the first 5 weeks, a teratoma was found in 2 out of 15 recipients [17]. To reduce or avoid the risk of teratoma formation in clinical applications, we transfected ESCs with TK, then used GCV to control the lifetime of the ESC-GFP-TKs in vivo. When the implanted ESC-GFP-TKs began to differentiate, the suicide gene was induced to kill the implanted ESC-GFP-TKs in a timely fashion. The ESC-GFP-TKs were completely eliminated or reduced to a level below the amount required for tumorigenicity in vivo to reduce the possibility of tumor formation and immune response (rejection or graft-versus-host reaction). The ESCs were in a low-growth state in the first 2 weeks after implantation; later, they began to differentiate into the epithelium [2], which was also found in our previous studies. We have previously found that some ESC-GFP-TKs still express OCT4 after co-culturing with cornea epithelium for 14 days [29]. To avoid frequent injections and side effects, a suicide gene was induced on the 11th day after the implantation of ESC-GFP-TKs to maintain an ESC microenvironment and avoid immune responses and tumor formation.
The tumorigenicity of ESCs after an in vivo transplantation is dose dependent; that is, the rate and timing of tumor formation are positively correlated with the cellularity of ESCs injected into the body. Our present study found that before the suicide gene was induced, apoptosis of ESC-GFP-TKs was rare (6.76%±0.8905%) in peripheral blood based on flow cytometry, which was similar to apoptosis in ESC-GFP-TKs in vitro (2%±0.15%). After inducing the suicide gene, almost all ESCs were apoptotic in peripheral blood (94.52%±0.21679%), which showed that this method can effectively remove ESC-GFP-TKs injected into the body. Our present study also observed that teratomas were found in the lung of mice that received ESC-GFP-TKs treatment without induction of the suicide gene during 8 weeks (8/10) (the rate of tumor formation was 80%), which was similar to Dressel's result, in which the rate of teratoma formation was 95% after transplanting ESCs into homologous or allogeneic immunodeficient mice. No teratoma developed in mice that received the ESC/suicide gene induction (8 weeks) treatment, likely due to clearing of the ESC-GFP-TKs or reducing their levels in the body in a timely manner, thus effectively reducing or avoiding the tumorigenicity of ESC-GFP-TKs. Even though a small amount of ESC-GFP-TKs remained, the amount of ESC-GFP-TKs was insufficient to form teratoma in our observation time. Although the evidence is insufficient to prove that this method can completely eliminate the tumorigenicity of ESC-GFP-TKs, other research has shown that inducing a suicide gene has a therapeutic effect even if a teratoma had formed; ESCs modified with TK were implanted into SCID mice subcutaneously, and a teratoma was produced at 6–8 weeks, but the volume of tumor was significantly reduced after inducing the suicide gene [25].
In the present study, teratomas were only found in the lung of animals after implanting ESC-GFP-TKs through the tail vein, which might be because ESC-GFP-TKs aggregated in the lung, so it is necessary to closely observe changes in the lung to see whether ESC-GFP-TKs are injected into the body through the tail vein. However, the origin of the organ selectivity remains unknown.
Reducing or avoiding the risk of immune response after ESC-GFP-TKs transplantation
The ESC microenvironment can repair and reconstruct the in vivo microenvironment, which depends on the undifferentiated state of ESCs because ESCs are at the lowest immunogenic stage at this time. Numerous studies have shown that undifferentiated ESCs do not express MHC I and MHC II antigens, so undifferentiated ESC do not generally activate the recipient's lymphocytes to induce immune rejection [30,31]. In addition, undifferentiated ESCs express high levels of FasL, which could induce apoptosis in host immune cells and therefore prevent rejection [32]. Once ESCs differentiate, other surface antigens are exposed [21,22], particularly the MHC II antigen, which increases the transformation of antigen-presenting cells to T cells, causing an immune rejection [31]. In the present study, we used modified ESCs with TK to achieve real-time control of ESC-GFP-TKs in vivo. ESC-GFP-TKs were killed before differentiating, which not only reduced or even prevented tumor formation in vivo but also reduced or avoided immune rejection related to ESC transplantation.
Reducing or avoiding graft-versus-host reaction
Hematopoietic stem cell therapy is the oldest and most widely used stem cell therapy. To reduce the rejection of the graft by the host immune system in vivo, the recipient should receive immunosuppression by radiation or chemotherapy before transplantation, which may increase the risk of graft-versus-host reaction [33], tumors, and infections. Teratomas form easily in immunocompromised recipients after ESC transplantation. When undifferentiated ESCs were directly injected into homologous or allogeneic immunodeficient mice, 95% of recipients developed a teratoma. If the same amount of ESCs were injected into immunocompetent allograft or xenograft recipients, there was no teratoma formation in any recipient in the same observation period, but if the heterogeneous recipients received cyclosporine A pretreatment, 61% of the recipients produced teratomas [34], suggesting that immunosuppression may increase the incidence of tumor formation.
Immune system preparation is achieved by destroying the patient's hematopoietic microenvironment through radiation or chemotherapy. In recent years, with a deeper understanding of the impact of microenvironment on cells, protection of the bone marrow microenvironment has received more attention. In one study, when undifferentiated ESCs were directly injected into immunosuppressed mice, the ESCs could not reconstitute hematopoiesis [15], most likely because of the damage to the hematopoietic microenvironment. Using the ESC microenvironment for treatment does not require immunosuppression. In the present study, recipients were not immunosuppressed before ESC transplantation and therefore had no damage to the bone marrow hematopoietic microenvironment, enabling the use of the ESC microenvironment to promote self-healing and reconstruction of in vivo microenvironment. Using the ESC microenvironment in treatment not only promotes self-healing of tissue but also, to some extent, reduces the incidence of graft-versus-host reaction, teratoma, and infection.
Appropriate cellularity is the basis for avoiding accidental death and achieving the best effect
Through pre-experiments, we found that after an injection of 1×108 ESCs into mice through the tail vein, the mice would immediately undergo convulsions, dyspnea, and cyanosis; they died within 5 min. Current studies generally use 1×106 cells as the therapeutic dose [35]. To prevent cell embolisms, our present study used 1×106 cells for a single cell injection dose to ensure that the dose did not affect the observation of in vivo tumorigenicity or the treatment effect.
In pre-experiments, we also observed that the animals' weight loss stopped or even began to increase over the first 3 days after ESC transplantation, then began to decrease on the fourth day. We speculated that the density of ESCs might decrease or that some began to differentiate 3 days after transplantation, which affected the maintenance of the ESC microenvironment. Thus, in our study, fresh ESCs were injected on the fourth day after the first transplantation. Fresh undifferentiated ESCs could strengthen the possibility of maintaining ESC microenvironment by the first injection of ESCs because of the increasing number of undifferentiated ESCs. In addition, the side effects of a single injection with a large number of ESCs were avoided.
In the present study, we observed that the number of ESC-GFP-TKs in peripheral blood significantly decreased from the 4th day to the 10th day by flow cytometry, which was possibly due to the ESC-GFP-TKs depositing in other organs (eg, lungs). In addition, the death of ESC-GFP-TKs could not be ruled out because of leaving the better environment in vitro; ESC-GFP-TKs in peripheral blood appeared dead.
In addition, we observed that animals rapidly lost body weight after inducing the suicide gene. We surmised that this result was caused by the death of ESCs in vivo and generated a 2-day window in which there were not enough ESCs. Therefore, we are planning to modify the ESCs with two types of suicide genes that can be induced by different drugs. ESCs carrying different suicide genes are injected in vivo at different times, and the suicide genes are induced at different times to make sure a certain amount of ESCs are present in vivo to avoid the window period caused by one suicide gene. Therefore, the ESC microenvironment will be more stable.
ESC microenvironment promotes self-healing
The cellular microenvironment plays a crucial role in cell growth, proliferation, and phenotype and it is the basis of transdifferentiation. Therapy using the ESC microenvironment uses ESCs to promote the proliferation of surviving cells in vivo and stop the differentiation of abnormal cells to generate self-healing and the reconstruction of the in vivo microenvironment. Our previous studies found that using ESC-conditioned medium, ESC co-culturing with corneal epithelial cells or corneal endothelial cells significantly improved the proliferation of terminal cells and even caused them to express some surface markers of precursor cells [18,19,29], which indicated that the ESC microenvironment could reverse cellular differentiation. The ESC microenvironment has been previously shown to change the epigenetics of melanoma cells and reduce their invasiveness [20]. Our experimental results indicated that the incidence of atypia and myelodysplastic peripheral blood and bone marrow were reduced after implanting ESCs into a leukemia model mice, and the survival rate increased. A total of 12.5% of the mice that received the ESC treatment survived at 8 weeks; in contrast, all the untreated mice were dead at 8 weeks. Leukemia is a malignant systemic disease. Although the survival rate of mice in this experiment did not significantly increase, the survival rate increased compared with the untreated group, which indicated the effectiveness of the ESC microenvironment at treating leukemia, but detailed treatment programs still need to be studied in further experiments.
Conclusions
In summary, taking advantage of the ESC microenvironment and low immunogenicity for in vivo transplantation can promote the proliferation of surviving normal cells and reduce the division of abnormal cells without destroying the original microenvironment in vivo. This study attempted to regulate the lifetime and retention of ESCs in vivo by inducing a transfected suicide gene. When ESC-GFP-TKs began to differentiate, lost the embryonic microenvironment, induced immune responses, or formed a teratoma, the suicide gene was induced to kill the ESC-GFP-TKs to avoid the danger of tumor formation and immune response to improve the safety in use of ESCs in clinical applications.
Footnotes
Acknowledgments
This work was supported by the Guangdong Natural Science Foundation (no. S2012010009113), the Natural Science Foundation of China (no. 81270971), the National High Technology Research and Development Program (863 Program) of China (no. 2012AA020507), and the Fundamental Research Funds of State Key Laboratory of Ophthalmology of China (2012PI05). The authors thank Professor Peng Xiang (Center for Stem Cell Biology and Tissue Engineering, Sun Yat-Sen University) for technical assistance.
Author Disclosure Statement
No competing financial interests exist.