
Abstract
Mesenchymal stem cells (MSCs) are a population of cells harboring in many tissues with the ability to differentiate toward many different lineages. Unraveling the molecular profile of MSCs is of great importance due to the fact that these cells are very often used in preclinical and clinical studies. We have previously reported the expression of insulin-like growth factor 2 mRNA binding protein 1 (IGF2BP1) an oncofetal mRNA-binding protein—in different stem cell types such as bone marrow (BM)-MSC and umbilical cord blood (UCB)-hematopoietic stem cells. Here, we demonstrate that MSCs of adipose tissue, BM, and UC origin have a differential pattern of IGF2BP1 and ten-eleven-translocate 1/2 (TET1/2) expression that could correlate with their proliferation potential. Upon IGF2BP1 interference, a significant reduction of cell proliferation is observed, accompanied by reduced expression of c-MYC and GLI1 and increased p21. We also present, for the first time, evidence that IGF2BP1 is epigenetically regulated by TET1 and TET2 demethylases. Specifically, we show that TET1 directly binds to the promoter of IGF2BP1 gene and affects the hydroxymethylation status of its promoter. These results indicate that IGF2BP1 and TET1/2 contribute to the stemness of MSCs, at least regarding their proliferative potential.
Introduction
M
We have previously reported the expression of the insulin-like growth factor 2 mRNA-binding protein 1 (IGF2BP1) in different stem cell populations, specifically BM-MSC and UCB-hematopoietic stem cells (UCB-HSC) [4,5]. IGF2BP1 is a member of a highly conserved gene family of RNA-binding proteins. It was first discovered by its ability to specifically bind to c-MYC mRNA and protect it from endonucleolytic cleavage, thus increasing its half-life [6]. Multiple studies have established an important role for IGF2BP1 in embryo development. Knockdown of the gene caused high levels of perinatal death (60%), while the animals that were finally born, were significantly smaller with hypoplastic tissues in almost every organ [7]. It was shown that IGF2BP1 and its homologues in chicken or Xenopus, bind to various mRNAs affecting their half-life, their translatability, and their subcellular localization. Among these targets are c-MYC [8], IGFII-leader3′ [9], H19 [10], tau, β-actin [11], and GLI1 [12]. All these genes have a key role in embryo development, in tissue-specific differentiation or even in cellular proliferation, differentiation, and motility. We have previously shown that this gene is under epigenetic control in BM-CD34+ isolated progenitors [4].
Methylation is a major mechanism through which the spatiotemporal pattern of expression of genes involved in cell development and differentiation is controlled. Defects in methylation patterns change the molecular profile and produce aberrant phenotypes in embryonic stem cells (ESCs) and embryos by causing loss of lineage restriction and transdifferentiation toward the extraembryonic trophoblast lineage [13]. It is noteworthy that the degree of methylation at the promoter loci of these “embryonic” genes in MSCs is greater than that in ESCs but less in fibroblasts, which suggests that the biology of MSCs as a group may be epigenetically restricted [14].
Recently, the ten-eleven-translocate (TET) family of proteins was identified as a new family of enzymes that alter the methylation status of DNA [15,16] and, therefore, may have an impact on the expression of genes crucial to pluripotency or cellular differentiation. As demonstrated by Ito et al., mouse ESC (mESC) express only Tet1 and Tet2 and knocking down of Tet1, resulted in significant reduction of Nanog expression in preimplantation embryos, diminished ESC proliferation, and overt ESC differentiation [17]. Another report by Koh et al. states that Tet1 depletion in mESC correlated with decreased expression of the Nodal antagonists Lefty1 and Lefty2, increased expression of the mesoderm/endoderm transcription factors Brachyury and Foxa2, and upregulated expression of the trophoectoderm transcription factors Cdx2, Eomes, and Elf5, which all underlie the role of TET proteins as key regulators of early embryonic development and differentiation that control lineage determination at critical decision points [18].
On the other hand, various studies reported that TET2 loss leads to increased replicating capacity in vitro and enhanced HSC function in vivo, suggesting that TET2 restrains aberrant self-renewal and expansion of HSC [19,20]. Thus, the TET family of proteins could influence a plethora of genes and play different roles in many cell populations at different time points.
In this study, we wanted to gain an insight into the role of IGF2BP1 and TET demethylases in human MSCs. Our data demonstrate coordinated regulation of IGF2BP1 by TET1 and TET2 suggesting a possible role for all these genes in the physiology of MSCs.
Materials and Methods
MSC isolation and culture
Isolation of MSCs from any source was performed within 24 h and under sterile conditions. All donors were informed and had signed a consent form. All procedures had prior approval from the Institutional Review Board of Saint Savas Cancer Hospital and from the Research Ethics Committee of the University of Athens. Furthermore, at the time of sample collection, all donors were disease free.
BM-MSCs were isolated as previously described [21]. AT-MSCs were isolated according to Zuk et al. [22]. Briefly, 5–10 mL of processed lipoaspirate were washed thrice using phosphate-buffered saline (PBS)-Dulbecco (Biochrom AG, Berlin, Germany,
UC-MSC isolation was as follows: received UC was rinsed in PBS-Dulbecco (Biochrom AG, Berlin, Germany) with 1× antibiotic-antimycotic (Gibco). A hematoma-free piece of 5 cm was dissected and the blood vessels were removed. The remaining tissue was cut into small pieces and plated in a T175 culture flask (Greiner Bio-One GmbH) in complete medium as explants.
MSC culture conditions were similar, independent of tissue origin. Two to 3 weeks upon initial plating (depending upon the colonies observed), cells were trypsinised using 0.25% trypsin-ethylenediaminetetraacetic acid (EDTA) (1×; Gibco) and re-plated in complete medium with a cell density of 250/cm2. Medium was replenished twice a week and further passage was performed when cells reached 80% confluence.
Phenotypic and functional characterization
Calculation of doubling time (T D)
The population doubling time was calculated as previously described [5] using the following equation: T D=ln2×Dt/(lnN t−lnN 0), where Dt is the time for which the culture was allowed to grow and N 0 is the inoculum cell number while N t is the cell harvest number.
Calculation of population doublings
The population doubling (PD) was calculated by the following equation: PD=(logN t−logN 0)/log2, where N 0 is the inoculum cell number and N t is the cell harvest number.
Calculation of growth index and cumulative GI
The growth index (GI) was calculated using the following equation: GI=N t/N 0, where N 0 is the inoculum cell number and N t is the cell harvest number. The cumulative GI (cGI) was calculated using the following equation: cGI=GIn×GI(n−1), where n is the passage number.
Surface staining
Phenotypic characterization of MSCs took place in every other passage. MSCs were stained, fixed, and analyzed, as previously described [5] on a FACSCalibur Flow Cytometer (BD Biosciences, San Jose, CA,
In vitro differentiation
Adipogenic and osteogenic lineage differentiation assays were performed on passages 2 and 4, as previously described [21]. Differentiation was assessed by OilRedO staining and Alizarin Red S staining, as previously described [5], for adipogenic and osteogenic differentiation respectively. All chemicals used in both stains were purchased from Sigma-Aldrich Co.
RNA isolation and quantitative RT-PCR analysis
Total RNA was extracted from MSCs using either the Nucleospin RNA II kit (Macherey-Nagel GmbH&Co, Dueren, Germany,
Quantitative RT-PCR (qPCR) was performed on a Rotor-Gene 3000 (Corbett Research, Sydney Australia,
Furthermore, the ΔΔCt method was used for data analysis of gene expression levels. GAPDH mRNA levels were used for internal normalization. NANOG levels were normalized to MCF-7, a breast cell line overexpressing embryonic markers. In addition, IGF2BP1, its targets, cell cycle genes, TET1 and TET2 levels were normalized to K562, a leukemic cell line.
RNA interference
Small RNA interference (siRNA) transfection experiments were performed using siIMPORTER™ (Upstate, Milton Keynes, United Kingdom, Upstate,
Control-Alexa Fluor 488-labeled siRNA was used to monitor the cellular uptake of each experiment (Qiagen Ltd, West Sussex, United Kingdom,
Western blotting
Whole-cell lysates were prepared from cells transfected with control or IGF2BP1 siRNAs for 24, 48, and 72 h. Cells were lysed in ice-cold RIPA buffer containing protease inhibitors. Protein concentration of each lysate was determined using a DC protein assay (Bio-Rad, Hercules, CA). Ten micrograms of total protein of each sample was analyzed in a NuPage 4%–12% Bis-Tris acrylamide gel and transferred to a PVDF membrane. Western blotting was performed using the following primary antibodies: IGF2BP1 (sc-166344) and β-TUBULIN (sc-5274) from Santa Cruz Biotechnology, Inc. (Dallas, TX,
Immunocytochemistry
MSCs were cultured in Lab-Tek slides (Nalge NUNc International, Rochester, NY,
Proliferation assays
Thymidine incorporation
MSCs were seeded at a density of 6,000 cells/cm2, in 96-well plates (Greiner Bio-One GmbH) 24 h prior to siRNA transfection. After 48 h of incubation, cells were pulsed with 1 μCi/well [methyl-3H] thymidine (specific activity: 53.0 Ci/mmol, 1 mCi/mL; Amersham, GE Healthcare, Amersham, United Kingdom,
Colony forming units-fibroblast assay
To evaluate the self-renewing capacity of MSCs, 150 cells/cm2 were seeded at passage 2 onto a six-well plate (Greiner Bio-One GmbH) in complete medium. At day 14, the medium was removed; cells were washed twice with PBS and stained with 0.5% crystal violet (Merck &Co, Inc., Whitehouse Station, NJ,
Hydroxymethylated DNA immunoprecipitation
Initially the promoter of the IGF2BP1 gene was retrieved using the Ensembl Genome Browser,
The hMeDIP kit (Cat No. AF-110-0016) and the MeDIP kit (Cat No. mc-green-001) from Diagenode (Seraing, Belgium,
Chromatin immunoprecipitation
For the chromatin immunoprecipitation (ChIP) assay, the SimpleChIP® Enzymatic Chromatin IP Kit (#9003; Cell Signalling, Danvers, MA) was used, according to the manufacturer's instructions. Briefly, 5×106 UC-MSCs were fixed with formaldehyde, to “preserve” the protein-DNA interactions. Cells were then lysed and chromatin was harvested and fragmented using a Dounce homogenizer and a bench-top sonicator (Ultrasonic, BRANSON200). The chromatin was then subjected to immunoprecipitation using TET1-specific Ab (GeneTex, GTX627420). DNA sequences associated with the protein of interest co-precipitated as part of the cross-linked chromatin complex and the relative amount of that DNA sequence was enriched by the immunoselection process. After immunoprecipitation, the protein-DNA cross-links were reversed and the DNA was purified. qPCR was used to detect the enrichment of the sequence of interest. The Histone H3 Rabbit mAb serves as a positive control IP for almost any locus studied and Normal Rabbit IgG serves as a negative control IP. In addition, ribosomal protein L30 (RPL30) primers were used as positive control.
Statistics
The Student's t-test with a 95% confidence interval was performed using GraphPad Prism version 5.00 for Windows, GraphPad 5 Software (
Results
IGF2BP1 and TET1/2 expression in MSCs of different origin
IGF2BP1 expression has been previously detected by our group in HSC and BM-MSCs, with indications of epigenetic regulation [4,5]. However, the normal role of this protein in stem cells has not yet been described. Recent publications assign to TET1 and TET2 DNA demethylases an important role in ESC physiology [17,18,23] by demethylating the Nanog gene promoter [17].
To explore the role of IGF2BP1 and TET1/2 in MSCs, we initially tested populations of fetal and adult origin for their expression pattern. All cell preparations met the phenotypic and functional criteria for MSC definition (Supplementary Table S4). As shown in Figure 1, IGF2BP1, TET1, and TET2 (A, B, and C respectively) mRNA levels of expression were considerably higher in UC-MSCs, as compared to both adult, BM, and AT. These differences in mRNA were also reflected in the corresponding protein levels (Supplementary Fig. S1).
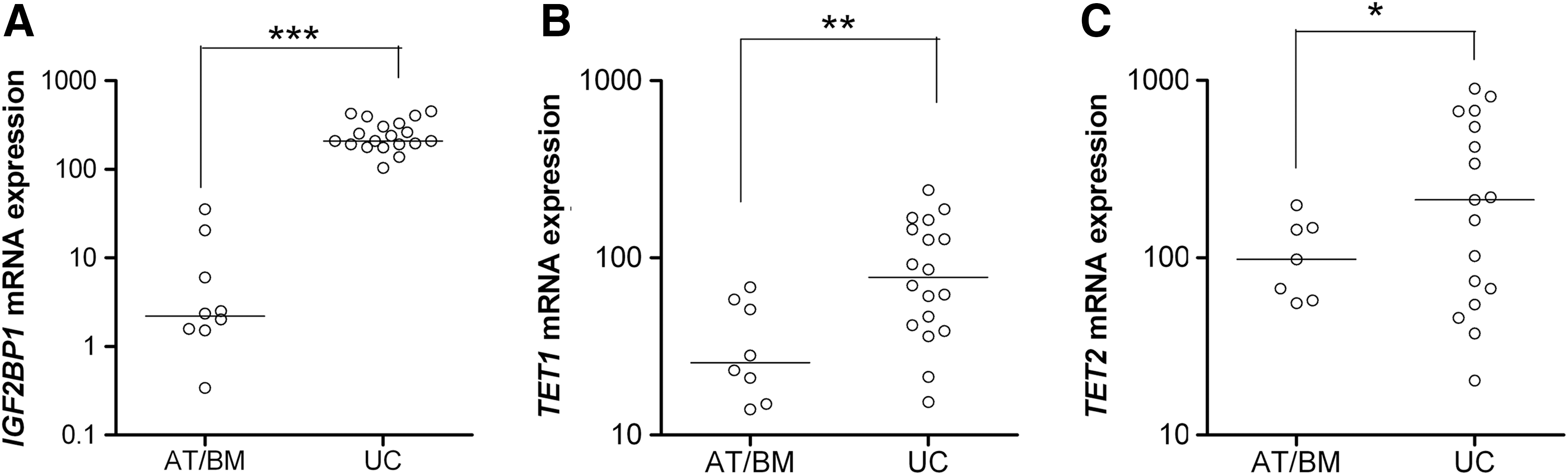
IGF2BP1
In line with other reports, we found that adult MSCs have a lower in vitro proliferative capacity compared with UC-derived MSCs [24] (Fig. 2A). It is also known that in vitro changes in MSC physiology, including their proliferative potential, can be attributed to changes in their gene expression pattern [25]. In this point we should underline that the different conditions used in cell isolation of each source, do not allow direct comparisons of the in vitro proliferative history spanning from seeding to first passage. Therefore, we decided to investigate the effect of the number of population doublings (PD)—as a measure of the proliferation rate—on gene expression within the same source, that is, the UC-derived MSCs that possess the highest proliferative capacity. As seen in Figure 2B, initially UC-MSCs proliferate fast [passages 1–3, with a median doubling time (T D) of 1.46 days] whereas after passage 6 they start to slowdown (median T D=5.11 days). To this end, we compared gene expression of UC-MSCs at passages 1–2 (PD estimated from P0=5–15) versus passages >7 (PD>30) (Fig. 2B). Indeed, the number of PD significantly affected IGF2BP1 mRNA levels, although never reaching the levels observed in adult MSCs (Fig. 2C). TET1 expression was significantly decreased to levels comparable to adult MSCs (Fig. 2D), while no significant decrease was observed in TET2 expression in relation to UC-MSC PDL (Fig. 2E). Altogether, these results are an indication that IGF2BP1 might be implicated in the regulation of UC-MSC proliferation.
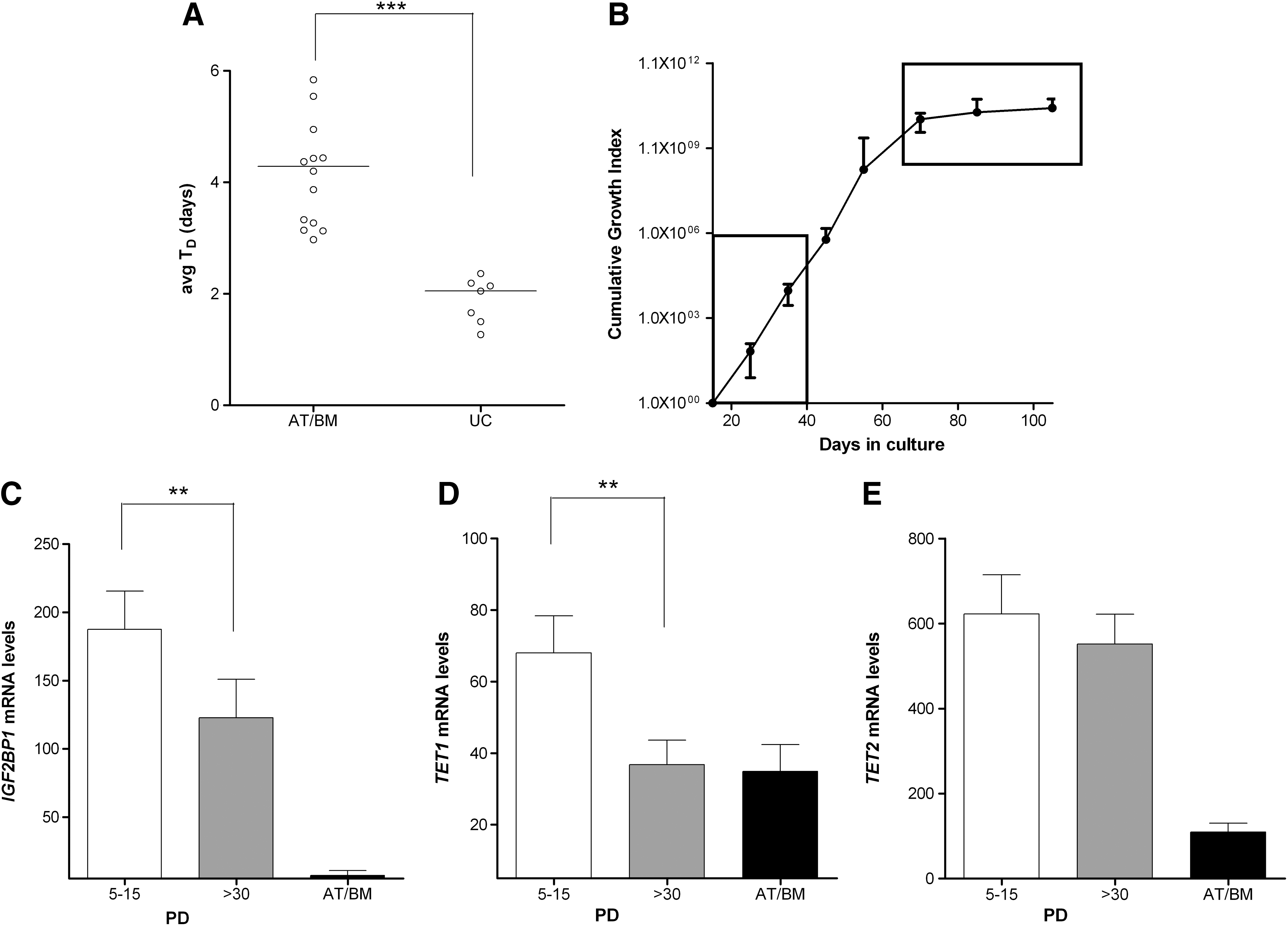
Proliferative capacity of human MSCs and expression of IGF2BP1, TET1, and TET2.
Inhibition of IGF2BP1 expression in UC-MSCs
To further investigate whether IGF2BP1 is involved in the regulation of UC-MSC proliferation, we applied RNA interference. Initially, we determined the efficiency of the assay at different time points. As depicted in Figure 3A, IGF2BP1 mRNA levels significantly reduced compared with control cultures for 48 h post-transfection and then gradually increased, but still remained at lower levels compared with control cultures 72 h post-transfection. This reduction in the mRNA levels was further confirmed at the protein level by western blot analysis (Fig. 3B).
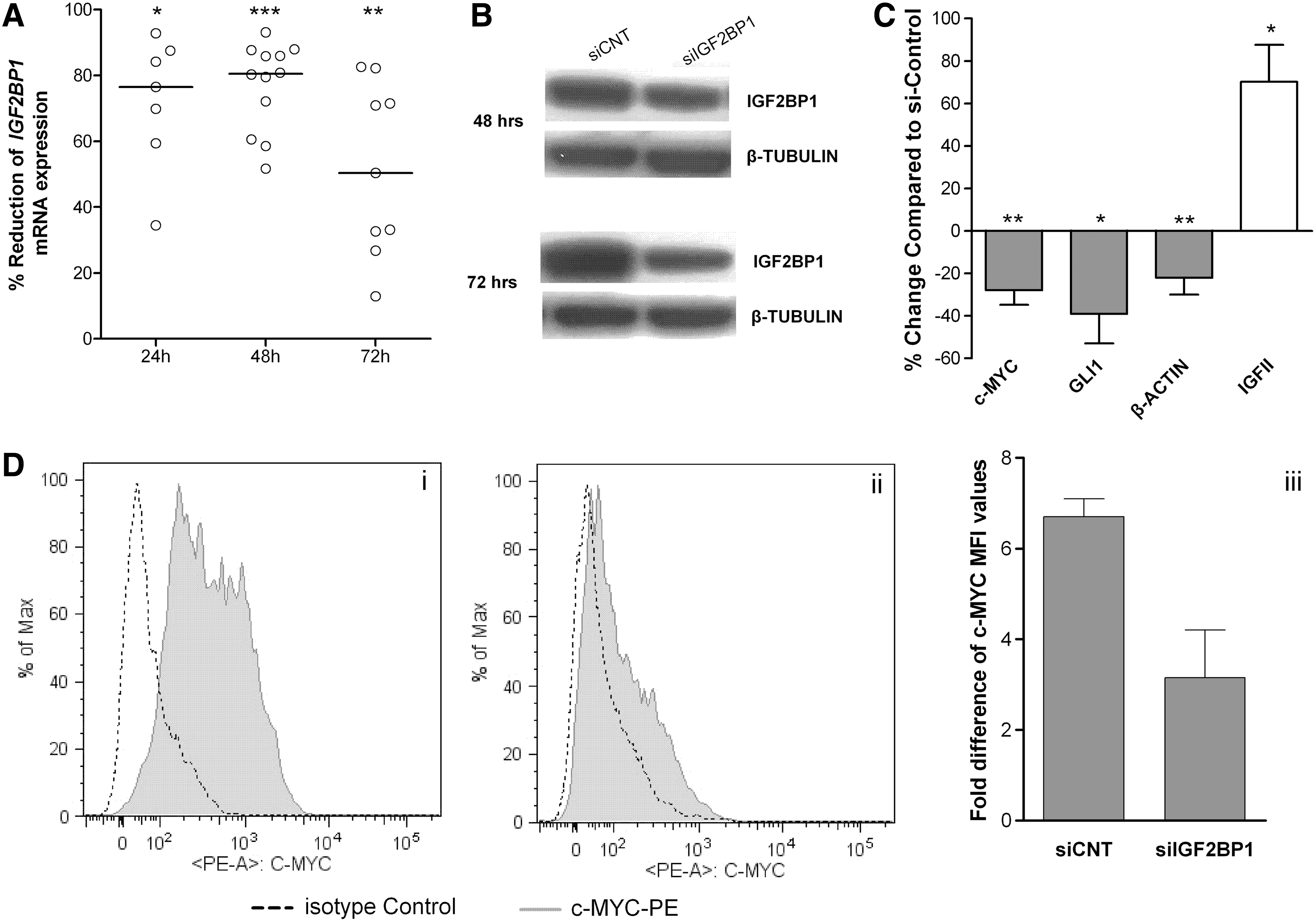
siRNA mediated reduction of IGF2BP1 levels of expression in human UC-MSCs.
Then, we examined the expression of certain genes previously reported to be targets of IGF2BP1 in other cell types, to verify whether the reduction of IGF2BP1 could result in a biological effect [12,26]. As shown in Figure 3C, c-MYC, β-ACTIN, and GLI1 mRNA levels were reduced and IGFII-leader3 mRNA levels were increased. The effect of IGF2BP1 knockdown was further controlled at the protein level of c-MYC, as its mRNA and protein molecules have a short half-life. Indeed, a significant reduction of c-MYC mean fluorescent intensity values—which represent the antigen intensity per cell—was observed (Fig. 3D).
Having established that RNA interference has the potential to elicit a biological effect, we further tested the outcome of IGF2BP1 interference on MSC proliferative capacity by 3H-TdR incorporation and colony forming units-fibroblast formation. Indeed, decreased IGF2BP1 expression caused a significant reduction in 3H-thymidine incorporation in cultures of UC-MSCs (Fig. 4A). In addition, a decrease in colony formation was observed (Fig. 4B and Supplementary Fig. S2).
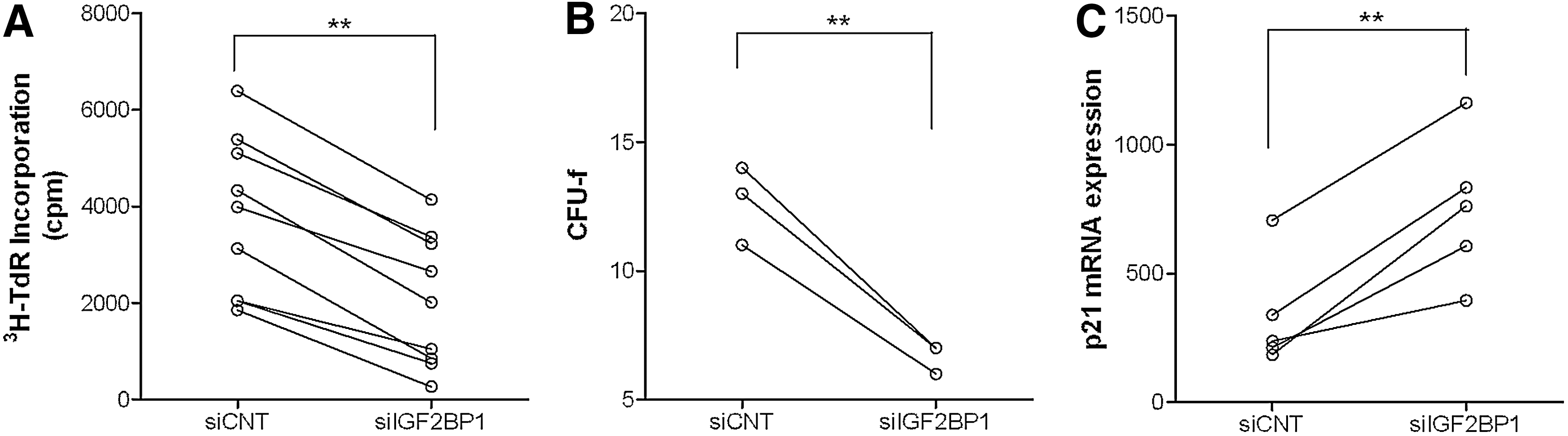
Effect of IGF2BP1 inhibition on UC-MSC proliferation.
We have previously shown that inhibition of proliferation in MCF-7 breast cancer cells after IGF2BP1 knockdown was related to reduction of c-MYC and elevation of cyclin-dependent kinase inhibitor 1A (p21) [4]. Bearing this in mind, siRNA-treated UC-MSC cultures were also tested for p21 expression. As expected, the mRNA levels of this cell cycle regulator were increased between 48 and 72 h in all samples tested (Fig. 4C).
Similar results after IGF2BP1 knocking down were also obtained in adult MSCs (Supplementary Fig. S3A). Furthermore, UC-MSCs at passage >7, exhibiting lower IGF2BP1 levels compared with their counterparts at passages 1–2, also expressed decreased GLI1 and p21 levels (Supplementary Fig. S3B).
To ascertain that these effects on proliferation were not due to apoptosis, we tested MSC cultures with Annexin-V/7AAD staining. No difference was observed in the percent of positively staining cells between control and siRNA-treated cultures (data not shown).
Taken altogether, these findings could point to the possible involvement of IGF2BP1 in MSC proliferative capacity.
TET1 and TET2 downregulation in UC-MSCs
To test our hypothesis that IGF2BP1 may be a downstream target of TET1 or TET2 demethylases, we knocked down each one of the two demethylases and evaluated any changes on IGF2BP1 mRNA levels by qPCR. As shown in Supplementary Figure S4A and D, TET1 and TET2 expression was significantly reduced, 48 h post-tranfection. Gene expression changes were also accompanied by decrease in protein expression levels (Supplementary Fig. S4B, E).
Prior to analyzing the relation of IGF2BP1 with TET proteins, we tested whether NANOG gene expression is affected by TET1 downregulation in UC-MSCs, as initially reported by Ito et al. in mESC [17]. Indeed, MSCs treated with siTET1 exhibited a significant reduction in NANOG expression (Supplementary Fig. S4C), while siTET2 resulted in elevation or no alteration of NANOG expression (Supplementary Fig. S4F).
To analyze the effect of TET1 or TET2 interference on IGF2BP1 and on MSC proliferation, we performed a series of experiments in UC-MSCs. As depicted in Figure 5B and C respectively, there was a significant reduction in IGF2BP1 mRNA levels upon exposure to siTET1, while a milder reduction was observed upon TET2 siRNA treatment. These findings indicate for the first time, to our knowledge, that IGF2BP1 gene could be a direct target of TET1.
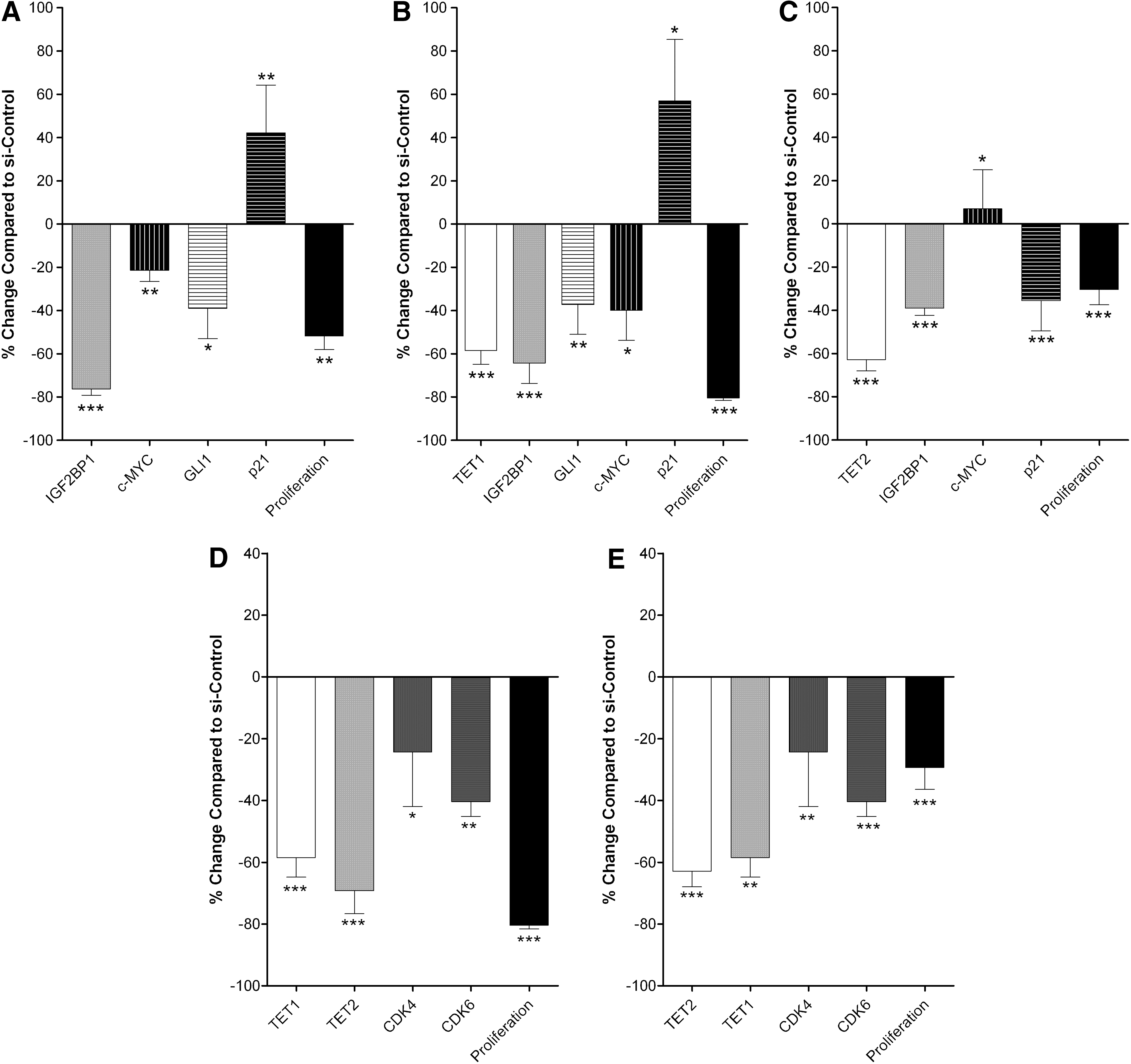
IGF2BP1, TET1, or TET2 interference and proliferation of UC-MSCs. Effect of IGF2BP1
We next investigated how decreased TET1 or TET2 gene expression influences UC-MSC proliferation, compared to IGF2BP1 siRNA treatment. As shown in Figure 5, there was a significant reduction in 3H-thymidine incorporation by the siRNA treated UC-MSC cultures for TET1 (Fig. 5B) and for TET2 (Fig. 5C), similar to siIGF2BP1 treatment (Fig. 5A). Subsequently, we wanted to test whether IGF2BP1 reduced levels of expression and the reduced proliferation caused by TET1 or TET2 gene interference correlate with the same molecular events.
A common pattern of events is observed between IGF2BP1 and TET1 interference (Fig. 5A, B respectively). Specifically, TET1 inhibition reduces IGF2BP1 mRNA to similar levels with those after treatment with siIGF2BP1. Levels of c-MYC are downregulated, while p21 is upregulated by both siIGF2BP1 and siTET1. On the other hand, TET2 reduction resulted in slightly increased c-MYC expression and decreased p21 expression (Fig. 5C).
Since inhibiting both, TET1 or TET2, resulted in reduced proliferation, although at varying levels, we proceeded to explore whether other molecules involved in cell cycle were differentially affected during this process. We observed an interaction between the two demethylases, since, siRNA against TET1 diminished TET2 mRNA levels and vice versa. Further exploring the molecular mechanisms for the reduced proliferation upon TET1 or TET2 interference, we found that cyclin-dependent kinases 4 and 6 (CDK4/6) mRNA levels significantly declined (Fig. 5D, E respectively).
Epigenetic status of IGF2BP1 gene in UC-MSCs
A number of studies report that only TET1 protein includes a CXXC domain, which is responsible for DNA binding, while TET2 molecule lacks this and the ability of direct binding [27].
In this context, to evaluate the epigenetic status of IGF2BP1 gene, relative levels of hydroxy-methylated IP-DNA were calculated in the genomic DNA of UC-MSCs treated with siTET1 versus UC-MSCs treated with Control si. As shown in Figure 6B, TET1 gene interference caused a significant reduction of hydroxymethylation levels in the tested CpG regions of the promoter (Fig. 6A), 1,000 bp upstream of the gene TSS. This reduction was mirrored by an increase in the methylation levels of the IGF2BP1 promoter in at least two of the rich CpG regions tested (Supplementary Fig. S5).
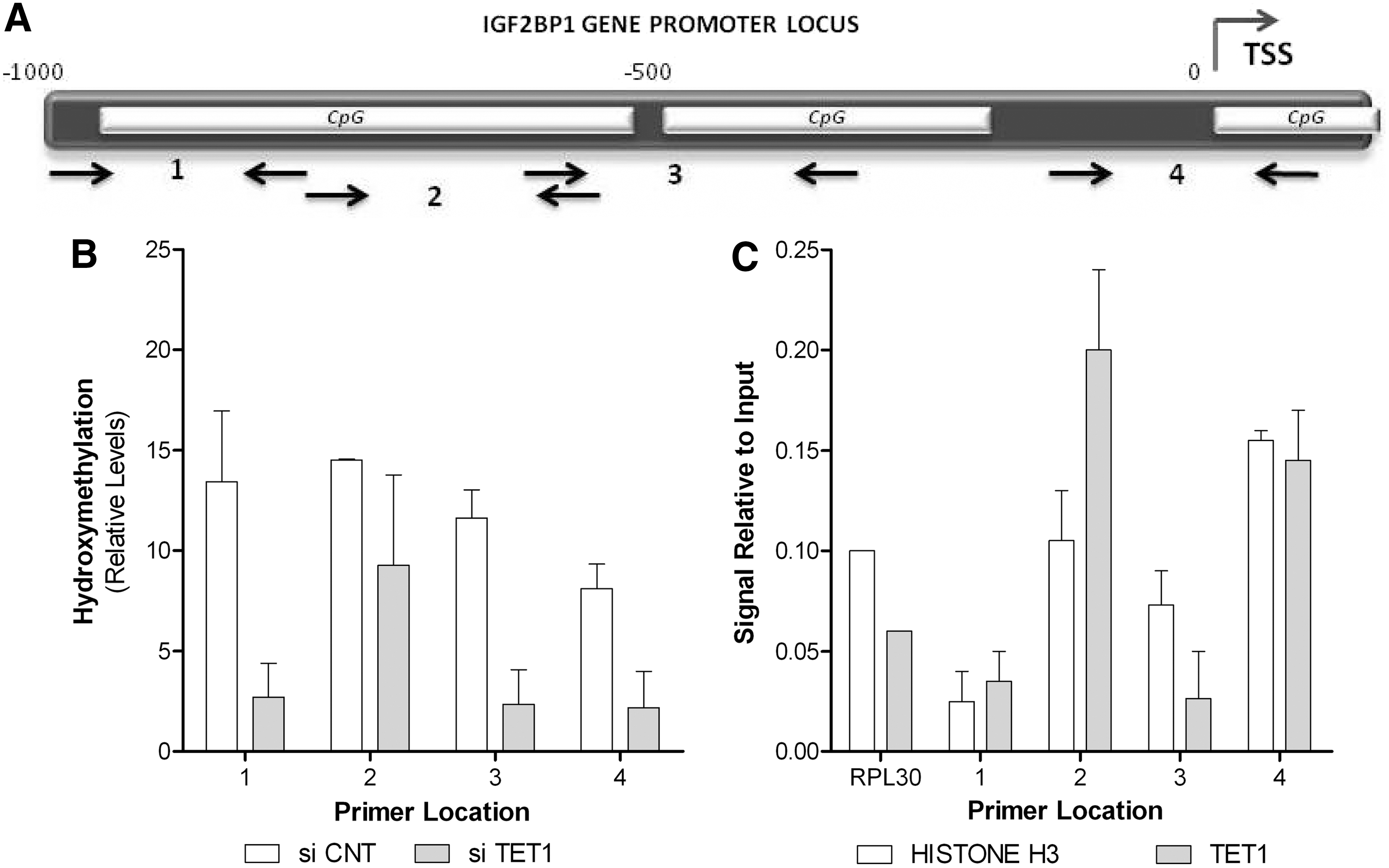
TET1 directly binds to IGF2BP1 and inhibits its methylation.
To further establish that TET1 controls IGF2BP1 methylation status by direct binding to its gene promoter, we performed ChIP analysis using a TET1-specific antibody (Fig. 6C). We detected significant enrichment of the IGF2BP1 promoter DNA sequences of all loci examined above for their hydroxymethylation status.
Discussion
ESC have been extensively investigated and are always used as a reference population for the deciphering of pathways involved in development and differentiation. Their characteristics comprise what is referred to as “stem” signature. A lot of scientific interest has focused on determining differences and similarities among stem cell populations, with MSCs being one of the most studied. They can be found in a multitude of tissues, other than the originally defined source of the BM [28]. In this study, we demonstrate the differential expression of important embryonic genes: IGF2BP1 and the TET1 and TET2 demethylases in human MSCs from different sources. We further look into the role of these genes and their possible interaction in regulating cellular proliferation.
IGF2BP1 is an mRNA-binding protein with a fetal pattern of expression [26]. We have previously reported that IGF2BP1 is expressed in stem cells with high renewal capacity like human UCB-HSC and mouse BM-MSCs [4,5]. Furthermore, we [4] and others [29] have assigned a role to IGF2BP1 in the control of cell proliferation in malignant cells.
In this study, we show that MSCs of fetal origin, UC-derived, which proliferate in vitro faster than their adult counterparts from BM and AT, also express significantly higher levels of IGF2BP1. Furthermore, UC-MSCs, which have undergone a limited number of PDL in vitro, also express higher levels of IGF2BP1 compared with their in vitro exhausted counterparts. This observation was further substantiated when we knocked down this gene and monitored MSC proliferation rate. Moreover, the significant reduction in IGF2BP1 expression resulted in a considerable reduction in the levels of c-MYC, GLI1, and p21. It is well established that c-MYC is a key regulator of cell cycle proliferation in almost all cell types [30]. Furthermore, a number of studies have reported c-MYC to be one of the major regulators of stem physiology [31], with prominent role in the production of induced pluripotent stem (iPS) cells, where the quantity and the exact time point of c-MYC transient expression are detrimental for the number of the iPS cells produced [32].
The GLI transcription factors—effectors of Hedgehog (Hh) signaling—are involved in cell fate determination, proliferation, and patterning in many cell types and most organs during embryo development, and in regulating stem cell maintenance and regeneration [33]. Moreover, GLI1 has been reported to be involved in the regulation of HSC proliferation [34]. Noubissi et al. [12] identified a novel mechanism, in colorectal tumorigenesis, by which Wnt signaling regulates the transcriptional outcome of Hh signaling pathway, via GLI1 mRNA stabilization by the RNA-binding protein IGF2BP1. Our data suggest that this mechanism might also be active in normal MSCs, partly to regulate their proliferation.
We have previously reported that knocking down of IGF2BP1 in MCF-7 breast cancer cells resulted in growth inhibition through c-MYC reduction and induction of p21 [4], a negative regulator of cell cycle, similarly to our current observations with MSCs.
In our previous work on IGF2BP1 expression in HSC, we showed that treatment of BM-CD34+ cells with 5-azacytidine resulted in de novo activation of the gene [4]. Additionally, it was recently reported that Tet1 and 2 demethylases play an important role in epigenetic control in mESC physiology [17,18]. These observations prompted us to investigate the possibility that these TET family members might also regulate IGF2BP1 expression through its methylation status in UC-MSCs.
Since no data exist regarding the pattern of expression of TET in MSC populations, we initially tested MSCs from all sources for TET1/2 expression. Thus, we report here for the first time, to our knowledge, that both these DNA demethylases are expressed in all samples tested, irrespective of their tissue of origin. However, it is worth mentioning that TET1 and TET2 are significantly higher in UC-MSCs, similar to IGF2BP1.
To further delineate the role of TET1/2 proteins, we studied UC-MSCs upon gene knockdown. The data presented herein show that TET1 and TET2 knockdown significantly affected the proliferative capacity of UC-MSCs, though to a different degree. This observation is in accordance with the studies by Koh et al. [18] who report that the effects of Tet1 depletion in mESCs were dominant over an often milder effect of Tet2 depletion. TET1 knocking down led to significantly reduced proliferation by increased p21 levels and a concomitant reduction in c-MYC and GLI1 levels, similar to IGF2BP1 knockdown. A weaker, though significant, reduction in cell proliferation was also observed upon TET2 silencing despite a reduction in p21 levels and an elevation in c-MYC levels. On the other hand, TET1 and TET2 depletion caused a significant reduction in the levels of CDK4/6, key regulators of cell cycle. Thus, TET1 affects both, c-MYC/p21 and CDK4/6 pathways, explaining the more robust inhibition of cell proliferation upon its silencing, whereas the milder effects observed with TET2 targeting may be explained by its ability to downregulate only CDK4/6.
Moreover, TET1 or TET2 inhibition resulted in a significant reduction in the mRNA of IGF2BP1. However, TET1 knockdown brought about a nearly 2-fold higher reduction in IGF2BP1 levels than did TET2. These findings could suggest that IGF2BP1 may be directly targeted only by TET1; a hypothesis further substantiated by the direct binding of TET1 on the IGF2BP1 promoter and the analysis of its hydroxymethylation status. Furthermore, it was recently reported that TET1 and TET2 have a physical association with NANOG. However, only TET1 and NANOG co-occupy genomic loci of pluripotency and lineage commitment genes in ESC, possibly assigning a co-factor role for TET2. Among these genes, IGF2BP1 is at the third place of the top-ranked target loci [35]. Thus, our results come in accordance with the notion that TET1 affects gene expression more dramatically while TET2 acts a co-factor whose loss can be compensated, and therefore the effect of its inhibition is milder.
The data presented in this study denote that human MSCs share with embryonic tissues many common molecular regulatory pathways, including the TET1/2 DNA demethylation machinery and the post-transcriptional regulatory protein IGF2BP1 (Fig. 7). It appears that these factors are part of a highly complex molecular network comprising the specific “stem” profile of the MSC population and their crosstalk needs to be further clarified. Whether IGF2BP1 is only related to the proliferative potential of MSC, an essential feature of stem cells, or is more fundamentally implicated in the stem physiology, remains to be investigated.
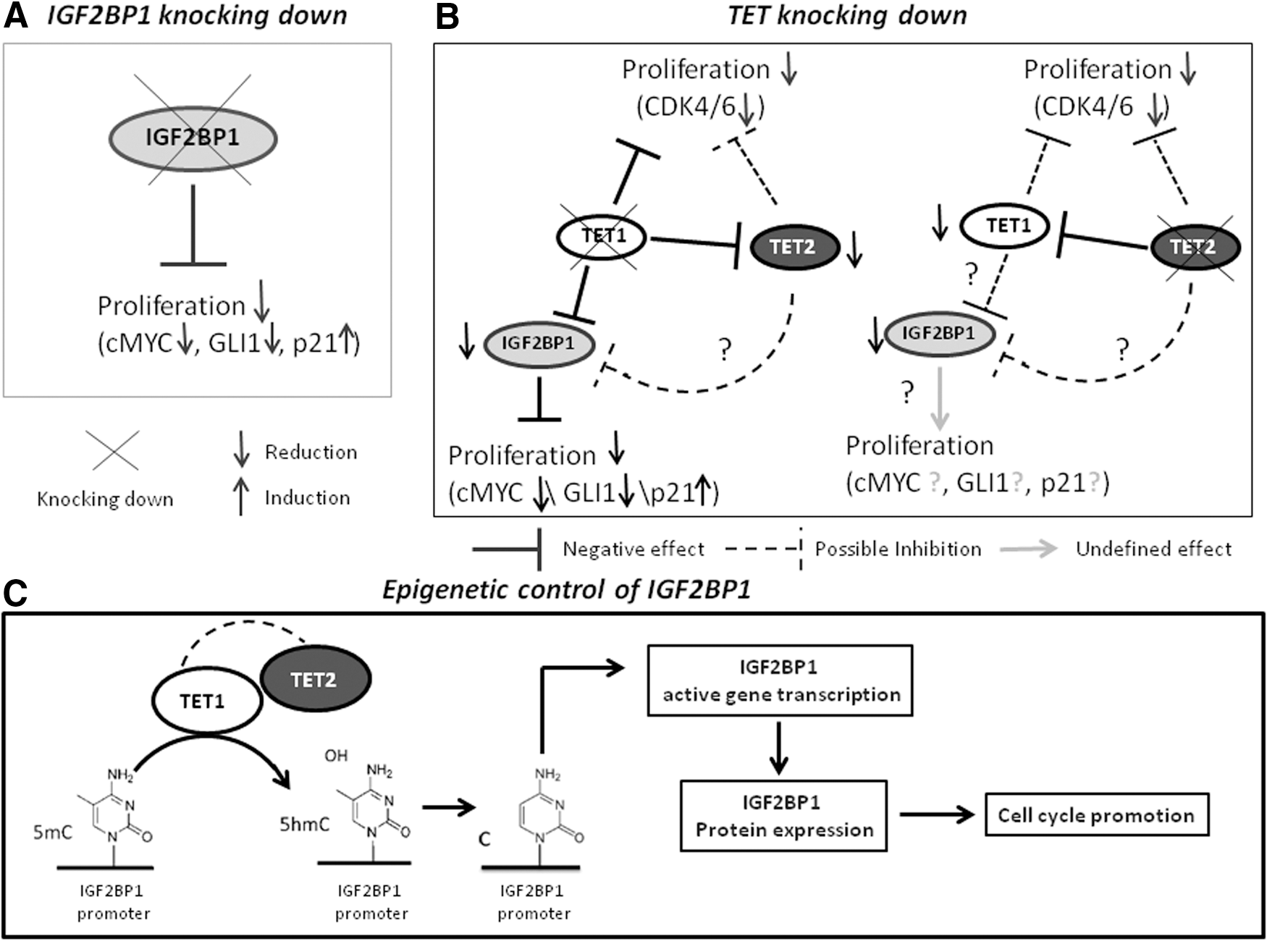
Schematic summary of the results obtained in this study.
Footnotes
Acknowledgments
We would like to thank E. Anastasopoulou for her assistance in flow cytometry and N. Cacoullos for her help in editing this article. We acknowledge a donation from OPAP SA to M.P.
Author Disclosure Statement
The authors indicate no potential conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.