
Abstract
Human amniotic fluid mesenchymal stem cells (hAFMSCs) are promising for therapeutic applications in bone damage. Calcium sensing receptor (CaSR), a G protein-coupled receptor, plays a physiological role in the regulation of bone metabolism. Thus, the bone CaSR could be targeted by calcimimetic agonists, which may be potentially helpful in treating bone diseases. The aim of our study was to characterize CaSR expression in hAFMSCs and to assess the activity of calcimimetic R-568 during in vitro osteogenesis. Using western blotting, immunofluorescence, and flow cytometry, we consistently observed constitutive CaSR in osteo-differentiating hAFMSCs. Notably, both R-568 and calcium significantly enhanced hAFMSC osteogenic differentiation after exposure to osteogenic medium. To provide further evidence of the involvement of CaSR in osteogenesis, we correlated its expression with that of established osteogenic markers, that is, alkaline phosphatase (ALP), runt-related transcription factor 2 (Runx2), and osteopontin (OPN), and novel, not yet completely defined regulators of osteogenesis. Among these are β-catenin and Slug, which are mediators of Wnt signaling, and nuclear factor of activated T cells c1 (NFATc1), which plays a critical role in calcium/calcineurin signaling. Taken together, our results demonstrate that CaSR is expressed in hAFMSCs, positively correlates with osteogenic markers, and is activated by R-568. Notably, downregulation of CaSR by RNA interference supports the conclusion that CaSR activation plays a central role in hAFMSC osteogenesis. Thus, this study provides significant information on the mechanisms of hAFMSC osteogenesis, which could provide additional molecular basis for the use of calcimimetics in bone regenerative medicine.
Introduction
H
Recent evidence points to the role of the calcium sensing receptor (CaSR) in osteogenic differentiation [9,10]. This G protein-coupled receptor is activated by millimolar concentrations of extracellular Ca2+, its main direct agonist. A variety of direct (type I agonist) and indirect allosteric (type II agonist) regulators increase the sensitivity and responsiveness of CaSR to extracellular calcium concentrations ([Ca2+]o). Moreover, polyvalent cations, such as Mg2+, Cd2+, Ba2+, La3+, and Gd3+ and some peptides act as CaSR agonists [11].
This receptor was first cloned from bovine parathyroid glands [12], where it controls parathyroid hormone secretion in response to changes in extracellular calcium levels. Subsequently, CaSR was found to be expressed in the kidney [13], and in gastrointestinal mucosal cells [14], blood vessels [15,16], bone marrow [17], osteoclasts [18], and osteoblasts [19]. Recently, we reported the first evidence of CaSR expression in ovine amniotic fluid-derived cells [20]. Accumulating in vitro and in vivo data indicate that this receptor plays a physiological role in the regulation of bone metabolism [21,22]. CaSR mainly regulates the recruitment, differentiation, and survival of osteoblasts and osteoclasts through activation of several intracellular signaling pathways [18,21].
Recently, Gabusi et al. [23] demonstrated a clear phenotypical and functional effect of [Ca2+]o on human osteoblasts and the data clearly support the hypothesis of a direct role by this cation in bone remodeling processes [24,25]. Interestingly, recent studies have reported that elevated [Ca2+]o significantly stimulates osteogenic differentiation of mesenchymal, adipose-derived, and human periosteal-derived stem cells [26 –28]. Whether this occurs in hAFMSCs as well, and whether the process is CaSR-dependent, remains to be determined.
In this report we evaluate CaSR expression and its regulation during osteogenic differentiation of hAFMSCs. We also examine the role of R-568, a type II CaSR agonist, and that of its enantiomer S-568 in in vitro hAFMSC osteoblastic differentiation.
To dissect the molecular mechanisms evoked by CaSR activation, we focused on Wnt and calcium/calcineurin signaling pathways, which are known to regulate bone mass and osteoblast-specific gene expression [29 –31]. To this end, we examined cell localization and expression of β-catenin, Slug (a zinc finger transcription factor belonging to the Snail family of developmental proteins) and NFATc1 (a nuclear factor belonging to the NFAT family that encompasses five proteins evolutionally related to the Rel/NFκB family).
Materials and Methods
Chemicals
Powered R-568-HCl and S-568-HCl were provided by Amgen (Amgen, Inc., Thousand Oaks, CA), resuspended in dimethyl sulfoxide (DMSO) at a 10 mM concentration and stored at −20°C. Calhex-231 and NPS-2143 hydrochloride (Santa Cruz Biotechnology, Santa Cruz, CA) were suspended in ethanol and DMSO, respectively, at 10 mM concentration and stored at −20°C.
Isolation and culture of human amniotic fluid mesenchymal stem cells
hAFMSCs were isolated from amniotic fluid (AF) of pregnant women during routine amniocentesis between 16 and 18 weeks of gestation. Women received detailed information about the experimental protocol, which was approved by the Ethics Committee of the University of Chieti, and each participant gave written consent for the study. Two or 3 mL of AF were obtained from six patients and cells were immediately isolated by centrifugation at 1,200 rpm for 10 min at room temperature (RT). The supernatant was discarded and the cell pellet was suspended in standard medium (SM) composed of low glucose Dulbecco's modified Eagle's medium (PAA Laboratories, Dartmouth, MA) supplemented with 20% fetal bovine serum (PAA Laboratories), 5 ng/mL recombinant human basic fibroblast growth factor (R&D Systems, Minneapolis, MN), 1% penicillin/streptomycin, and 1%
In vitro osteogenic differentiation and treatment with calcimimetics
hAFMSCs were seeded at 3,000 cells/cm2 in six-well plates and osteogenic differentiation was induced by treating cells for up to 21 days with appropriate osteogenic medium (OM), containing 0.05 mM ascorbic acid-2-phosphate (Sigma-Aldrich, St. Louis, MO), 10 mM β-glycerophosphate (Sigma-Aldrich), and 100 nM dexamethasone (Sigma-Aldrich) and in the presence or absence of CaCl2 (2.5 mM; Sigma-Aldrich), calcimimetics R-568, and S-568 (1 μM). The concentration of calcium present in both SM and OM is 1.8 mM; then, we added the amount of calcium necessary to reach the final 2.5 mM concentration. In some experiments, the inhibitor Calhex-231 (a negative allosteric modulator of the CaSR receptor, 1 μM) or the selective inhibitor NPS-2143 (1 μM) was given to cells 30 min before the addition of treatments.
Osteogenic differentiation was assessed using Alizarin Red S (ARS) staining (Sigma-Aldrich), the alkaline phosphatase (ALP) assay (Sigma-Aldrich) and quantitative RT-PCR (qRT-PCR) for the expression of ALP, runt-related transcription factor 2 (RUNX2), osteopontin (OPN), type I collagen (COL I), and osteocalcin (OC).
Quantitative RT-PCR
Total RNA from hAFMSCs, cultured in SM or OM, was isolated using the RNeasy Plus Universal Mini Kit (Qiagen, Inc., Valencia, CA) according to the manufacturer's instructions. The quality of total RNA was assessed by spectrophotometric measurements of the A260/280 ratio. For reverse transcription, M-MLV Reverse Transcriptase Reagents (Sigma-Aldrich) were used. qRT-PCR was carried out with the ABI Prism 7900 Sequence Detection System (Applied Biosystems, Foster City, CA). hAFMSC expression of ALP, RUNX2, OPN, COL I, and OC was evaluated at 7, 14, and 21 days.
Commercially available TaqMan Gene Expression Assays (ALP, Hs01029144_m1, RUNX2, Hs00231692_m1, OPN Hs00959010_m1, COL I Hs00164004_m1, OC Hs01587813_g1, and COL II Hs00264051_m1) and the TaqMan Universal PCR Master Mix (Applied Biosystems) were used according to standard protocols. Beta2 microglobulin (B2M, Hs99999907_m1; Applied Biosystems) was used for template normalization. Duplicates were set up for each sample.
ARS and ALP assays
Osteogenic differentiation was assessed by ARS staining and ALP assay at respectively 14 and 7 days of exposure to OM, using established protocols [28,32]. For the ARS assay, cells were fixed with 4% paraformaldehyde (PFA) for 15 min. After rinsing with distilled water, cells were stained with 40 mM ARS (pH 4.2) for 20 min at RT, then washed and analyzed [32]. For quantification of ARS staining, 800 μL 10% (v/v) acetic acid was added to cells for 30 min at RT under gentle shaking. The monolayer was then collected using a cell scraper and diluted with 10% (v/v) acetic acid. Subsequently, 500 μL mineral oil (Sigma-Aldrich) was added to samples, which were heated to 85°C for 10 min and then placed for 5 min on ice. The samples were then centrifuged at 20,000 g for 15 min. Supernatants (500 μL) had 200 μL of 10% (v/v) ammonium hydroxide added and aliquots (150 μL) were read in triplicate at 405 nm.
ALP staining was performed using the ALP colorimetric assay purchased from Sigma-Aldrich.
Fourier transform infrared spectroscopy
The extracellular matrix composition was analyzed by Fourier transform infrared (FT-IR) spectroscopy. Cell layers were collected with ammoniated water (50 mM ammonium bicarbonate, pH 8.0), lyophilized, and analyzed by an FT-IR Spectrometer (Spectrum 100; PerkinElmer, Salem, MA). Absorption spectra were collected from 4,000 to 600 cm−1 [33].
Western blotting
CaSR protein levels were evaluated at day 14 on undifferentiated and osteogenic differentiated hAFMSCs by western blotting (WB) as previously described [16]. Briefly, confluent cell monolayers were collected by scraping and suspended with ice cold lysis buffer containing 50 mM Tris-HCl, pH 7.4, NP 40 1%, 150 mM NaCl, and a cocktail of protease and phosphatase inhibitors (1 μg/mL each of aprotinin, leupeptin and pepstatin, 1 mM each of PMSF, Na3VO4, and NaF). Under nondenaturing (not preheating at 100°C) and reducing conditions, 100 μg of protein was used and resolved by sodium dodecyl sulfate–polyacrylamide gel electrophoresis on 8% polyacrylamide gels.
After incubation with blocking solution (5% dry milk in TBS Tween 20 0.1%) for 3 h at RT, the blots were exposed overnight at 4°C to monoclonal anti-CaSR antibody (1:1,000; Thermo Scientific, Milan, Italy) and then to goat anti-mouse IgG (1:10,000; Calbiochem, San Diego, CA). The blots were washed and protein bands were detected using ECL Prime Western Blotting Detection Reagent (Amersham Pharmacia Biotech, Buckinghamshire, England).
Immunostaining
For identification of the CaSR receptor, undifferentiated and differentiated hAFMSCs were cultured on glass coverslips for 14 days, fixed with 4% PFA (Sigma-Aldrich) for 10 min at RT, and permeabilized for 30 min at RT in Net-Gel (NaCl 150 mM, EDTA 5 mM, TRIS-HCl 50 mM pH 7.4, NP-40 0.05%, λ-Carrageenan 0.25%, and NaN3 0.02%). After permeabilization, cells were incubated for 1 h at RT with primary monoclonal antibody against CaSR (1:100; Thermo Scientific), and then stained for 1 h at RT with FITC-conjugated secondary antibody (1:200; Invitrogen, Milan, Italy).
Nuclei were stained using DAPI solution (0.5 μg/mL; Sigma-Aldrich). A similar procedure was performed using antibodies against active-β-catenin (1:100; Millipore, Bedford, MA), NFATc1 (1:50; Santa Cruz Biotechnology), and SLUG (1:100; Cell Signaling Technology, Inc., Danvers, MA). Alexa-488 fluorophore-conjugated secondary antibody (1:200; Invitrogen) was used as the secondary antibody [34]. Slides were observed with a 50i Eclipse microscope (Nikon) and images were acquired with a Cool-SNAPcf digital CCD camera (PhotoMetrics, Huntington Beach, CA). Digital acquisition, processing, measure, and analysis of fluorescence were performed by Meta Image Series 7.5 (MetaMorph, Metafluor, MetaVue) software obtained from Molecular Devices [35].
Flow cytometry
Antibodies
FITC-conjugated anti-CD13, anti-CD44, anti-CD45, anti-CD105, anti-CD166, and PE-conjugated anti-CD29 were obtained from Ancell (Bayport, MN); anti-CD14-FITC was obtained from Miltenyi Biotec (Bergisch Gladbach, Germany); anti-HLA-DR-PE, anti-HLA-ABC-Alexa488, anti-CD90-FITC, anti-CD73-PE, anti-Sox-2-Alexa488, anti-SSEA-4-FITC, anti-OCT3/4-PE, anti-CD117-APC, and anti-CD146-FITC were purchased from Becton Dickinson (BD Biosciences, San Jose, CA); anti-CD144-FITC was purchased from Acris Antibodies (Herford, Germany); anti-CD34-PE was obtained from Beckman Coulter (Fullerton, CA); anti-h-TERT was purchased from Calbiochem (Dermastadt, Germany); and secondary FITC-conjugated was purchased from Jackson ImmunoResearch Laboratories (West Grove, PA).
hAFMSC flow cytometric phenotype
Briefly, sample cells were washed and stained for surface or intracellular antigens, as previously described [36]. Surface antigens were evaluated in samples incubated with specific antibodies for 30 min at 4°C in the dark and then incubated for 5 min in 0.5% PFA at RT. Intracellular antigens were evaluated in fixed (Lysing solution; BD Biosciences) and permeabilized cells treated with Perm II (BD Biosciences). Then, samples were incubated with specific antibodies for 30 min at 4°C in the dark and for 5 min in 0.5% PFA at RT [36].
CaSR flow cytometry
As previously described, cells were stained under permeabilizing and nonpermeabilizing conditions to visualize both total and extracellular membrane CaSR. Briefly, only for permeabilized samples, the staining procedure followed a previous step of membrane fixation and permeabilization, as previously described [20]. Then, in both cases, the staining was performed incubating 5×105 cells/sample with primary monoclonal antibody against CaSR (1:100; Thermo Scientific) for 45 min on ice, followed by staining with Alexa488-labeled secondary antibodies (1:200; Jackson ImmunoResearch Laboratories) for 30 min on ice. To evaluate cell viability, 7-aminoactinomycin D (7-AAD), was used (BD Biosciences). To assess nonspecific fluorescence, we used samples stained with the corresponding secondary antibody alone. Samples were analyzed by flow cytometry. Data are indicated as a percentage of positive cells or mean fluorescence intensity for CaSR in live and nonpermeabilized cells.
Flow cytometry measurement
Cells were analyzed on a FACSCanto II flow cytometer (Three Laser, eight-color configuration; BD Biosciences), using FACSDiva software v.6.1.3 (BD Biosciences).
Quality control included regular checkup with Rainbow Calibration Particles (BD Biosciences). Debris was excluded from the analysis by gating on morphological parameters; 20,000 nondebris events in the morphological gate were recorded for each sample. To assess nonspecific fluorescence we used specific irrelevant controls or secondary antibodies. All antibodies were titrated under assay conditions and optimal photomultiplier gains were established for each channel [37]. Data were analyzed using FlowJo™ software (TreeStar, Ashland, OR).
RNA interference and cell transfection
siRNA duplexes for CaSR were purchased from Eurogentec based on the sequences kindly provided by Dr. Romuald Mentaverri upon request [38]. Briefly, siRNAs (18.75 nM final concentration for each duplex) were transfected in hAFMSCs with Hiperfect (Qiagen, Hilden, Germany), according to the manufacturer's instructions advised for transfection of human fibroblast cell lines. As control siRNA duplexes we employed nontargeting scrambled sequences previously described [39]: Sc-A, 5′GCU UCC UAC CAC AAU UUC U dTdT and Sc-B, 5′GGU AAU CAA CUA AUC UUA A dTdT. Cells were transfected every 72 h up to 7 days, and the suppression of CaSR expression was assessed by flow cytometry.
Statistical analysis
Data are reported as mean±standard deviation. Statistical comparisons were made using Student's t-test. A difference was considered significant at P<0.05.
Results
hAFMSC isolation and characterization
Adherent cell populations from human amniotic fluid mesenchymal stem cells (hAFMSCs) were generated [35] and immunophenotypized as reported in Table 1.
Expression: −, negative; +/−, low; +, positive; ++, high.
Phenotypic FACS analysis showed that hAFMSCs at passage 3 stably expressed mesenchymal markers (CD90, CD73, and CD105) and surface adhesion molecules (CD13, CD29, CD44, CD146, and CD166), while they did not display any hematopoietic molecular markers (CD14, CD34, and CD45). They were positive for the intracellular stemness markers SSEA-4, Sox-2, and h-TERT and expressed a low level of OCT3/4, while CD117 and surface endothelial marker CD144 were not expressed. In addition, hAFMSCs were positive for HLA-ABC (MHC class I antigen) while HLA-DR (MHC class II antigen) expression was not detected.
Osteoblastic differentiation of hAFMSCs
hAFMSCs were analyzed for their capacity to differentiate into osteogenic lineage (Fig. 1). During osteogenic differentiation, we evaluated the expression of the osteogenic markers ALP, RUNX2, OPN, COL I, and OC at 7, 14, and 21 days, by qRT-PCR analysis (Fig. 1A). As expected, ALP was abundant in early stages of differentiation and then decreased. Runx2 strongly increased at day 14 of differentiation and was then downregulated to promote osteogenesis that, on the contrary, is accompanied by a progressive increase in OPN. COL I increased at day 21 of differentiation while OC increased by day 7 and significantly at day 14 and 21.

Estimation of osteogenic differentiation of hAFMSCs.
As confirmation of osteogenic differentiation, hAFMSCs grown for 14 days in OM changed their morphology with the formation of cellular aggregates (see bright field images in Fig. 1B, C), produced mineral matrix (see ARS staining in Fig. 1D–G), and showed an increase in ALP activity (Fig. 1H–M). In addition, the possible presence of chondrogenic markers such as collagen type II and cartilage-specific proteoglycans was excluded by reverse transcriptase-polymerase chain reaction (RT-PCR) and Alcian Blue staining respectively (data not shown), confirming the specificity of osteogenic differentiation.
CaSR identification
CaSR expression was evaluated by western blotting (Fig. 2A), under reducing and nondenaturing conditions. We observed a band of about 160 kDa consistent with results from previous studies with other experimental models [16,19,40], and likely corresponding to totally glycosylated monomeric mature form of the receptor. Notably, its expression strongly increased after 14 days of osteogenic differentiation (P<0.001).
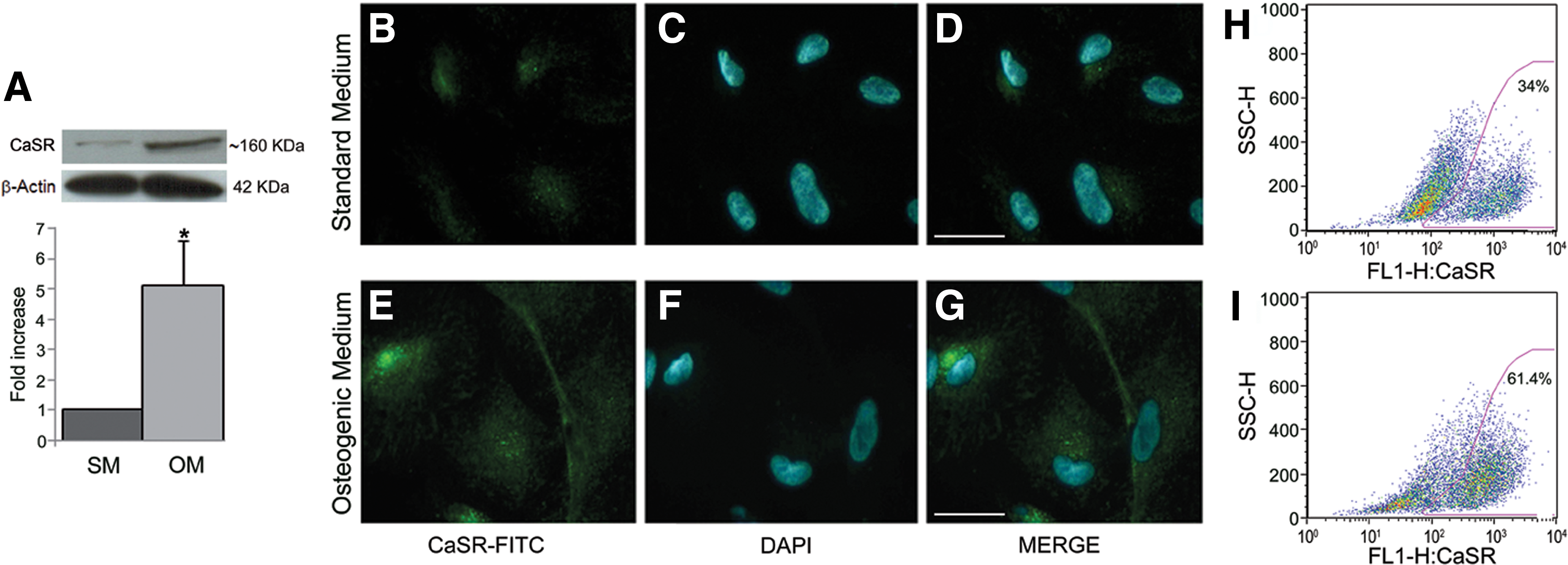
Expression of calcium sensing receptor (CaSR) in hAFMSCs.
Immunofluorescence microscopy clearly confirmed that CaSR levels increased after 14 days of osteogenic differentiation (Fig. 2E–G) as compared with undifferentiated cells (Fig. 2B–D), and demonstrated that CaSR is mostly localized on the cell surface. These data were confirmed by quantitative flow cytometric analysis (Fig. 2H, I) showing that CaSR cell surface expression significantly increased in osteogenic differentiated cells (P<0.05, n=3). In particular, the percentage of CaSR-positive cells was 64.4%±6.4% in OM (Fig. 2I) and 34.6%±0.9% in SM (Fig. 2H). As a whole, CaSR expression data indicate that the increase of functionally active form of CaSR is positively associated with osteogenic differentiation.
Effects of CaSR agonists on osteo-differentiating hAFMSCs
We next investigated the impact of intracellular calcium ([Ca2+]i) increase or the exposure to calcimimetics on hAFMSC osteogenic differentiation. The transcripts of well-known osteogenic markers (ALP, Runx2, and OPN) were analyzed at their previously established peak expression (Fig. 1A), in cells exposed to high CaCl2 (2.5 mM) or calcimimetics R-568 and S-568 (1 μM).
As shown in Figure 3A, both CaCl2 and R-568, but not its enantiomer S-568, enhanced the expression of all osteogenic markers analyzed. Importantly, exposure to the CaSR allosteric inhibitor, Calhex-231, significantly reduced the effect of calcium and R-568 on ALP, Runx2, and OPN expression. This hypothesis was supported by data from the analysis of mineralized matrix deposition (Fig. 3B). ARS staining revealed a significant increase in mineralization when the cells were exposed to high calcium concentration (2.5 mM) or to R-568 (1 μM) for 14 days, as compared with exposure to the standard calcium concentration (1.8 mM) present in basal OM. The increase of ARS staining induced by calcium or R-568 was suppressed by Calhex-231. The mineral content in hAFMSCs was identified as apatite by FT-IR spectroscopic analysis (Supplementary Fig. S1; Supplementary Data are available online at
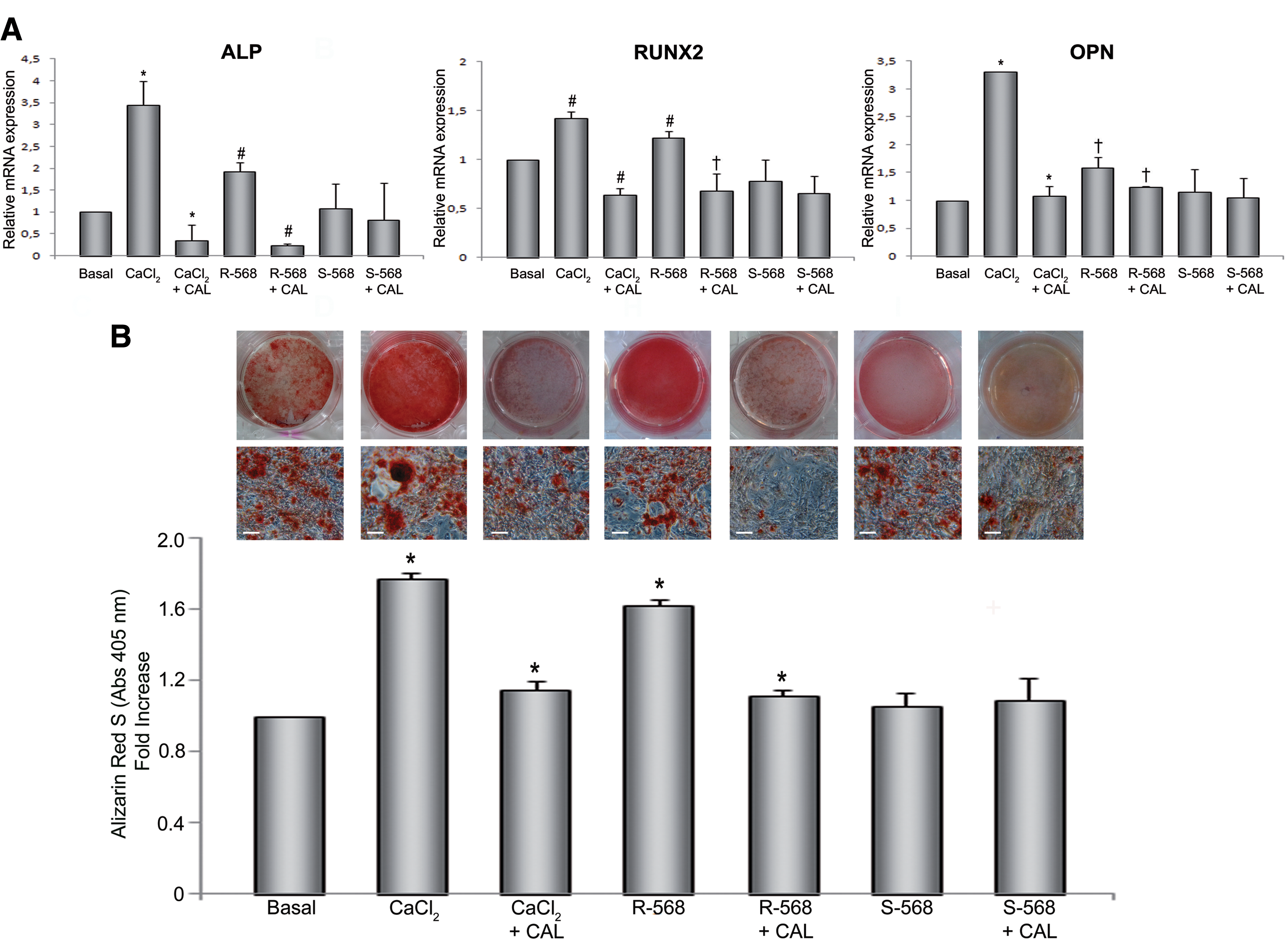
Effect of calcium and calcimimetics on hAFMSC osteogenic differentiation.
Calcium and R-568 stimulate β-catenin, SLUG, and NFATc1 expression and proper localization in hAFMSCs
We next focused on the expression of other positive regulators of osteogenesis, two mediators of the Wnt pathway, namely β-catenin and Slug, and NFATc1, a key mediator of calcium/calcineurin signaling. Figures 4 –6 show immunofluorescence staining of hAFMSCs grown for 14 days in OM and exposed or not to CaCl2 (2.5 mM) or R-568 (1 μM). Since we did not observe any significant effect with S-568—as in previous experiments—we used only R-568.

Effect of CaCl2, R-568 and Calhex-231 on β-catenin expression and nuclear translocation in hAFMSCs grown for 14 days in OM. Immunofluorescence microscopy images show the expression of β-catenin (green fluorescence) and DAPI positive nuclei (blue fluorescence), and then a merge of the two. In all panels a magnification is shown in the bottom-right corner. Scale bar 100 μm. Images are representative of n=3. Color images available online at
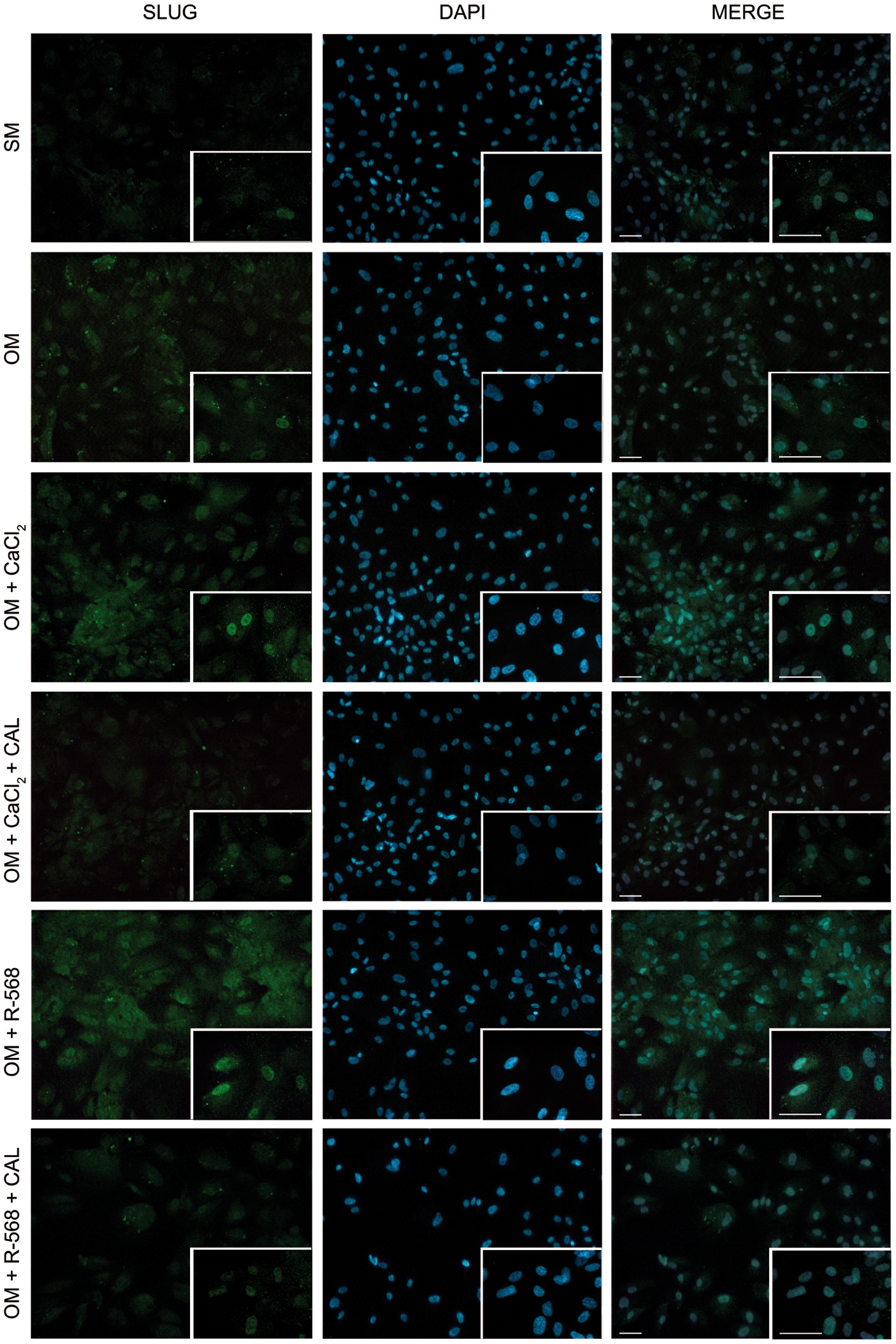
Effect of CaCl2, R-568, and Calhex-231 on Slug expression in hAFMSCs grown for 14 days in OM. Expression of Slug is revealed by green fluorescence. Nuclei are stained with DAPI (blue fluorescence). The merge of the two colors is shown in the right-hand column. In all panels a magnification is shown in the bottom-right corner. Scale bar 100 μm. Images are representative of n=3. Color images available online at
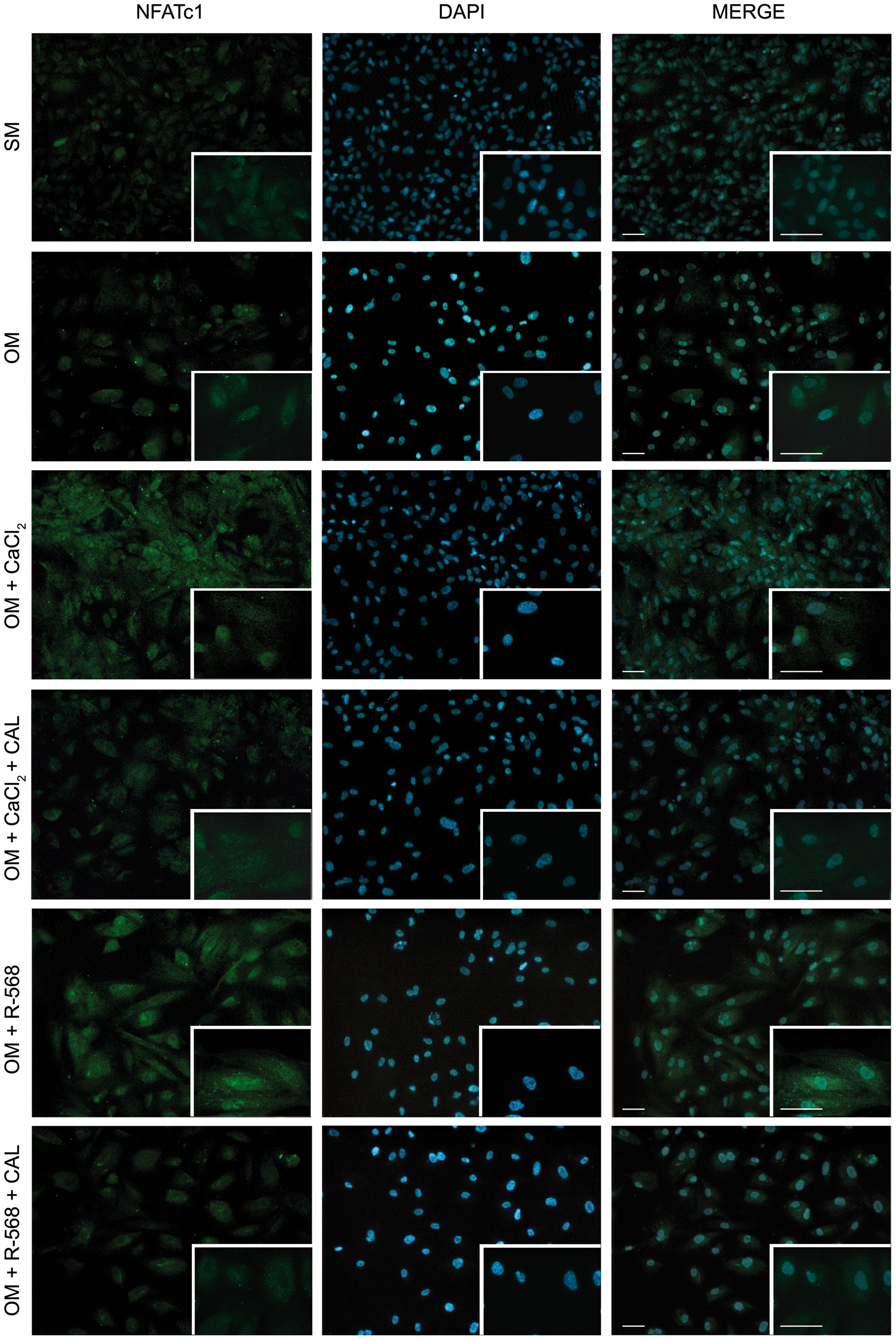
Effect of CaCl2, R-568, and Calhex-231 on nuclear factor of activated T cells c1 (NFATc1) expression and nuclear translocation in hAFMSCs grown for 14 days in OM. NFATc1 (green fluorescence), DAPI-positive nuclei (blue fluorescence). The merge of the two colors is shown in the right-hand column. In all panels a magnification is shown in the bottom-right corner. Scale bar 100 μm. Images are representative of n=3. Color images available online at
An increase of β-catenin (Fig. 4), Slug (Fig. 5), and NFATc1 (Fig. 6) expression was observed in osteogenic differentiated cells and even more when calcium and R-568 were added. Quantification of single cell fluorescence intensity of β-catenin, Slug, and NFATc1 confirmed these observations (Supplementary Fig. S2), suggesting that these transcription factors participate in CaSR agonists-induced hAFMSC osteogenic differentiation. Moreover, fluorescence analysis of nuclear translocation (Supplementary Fig. S3), indicated that both R-568 and CaCl2 induced a translocation of β-catenin (83% and 71%, respectively) and NFATc1 (63% and 75%, respectively) from the cytoplasm to the nucleus, where they act as functional transcription factors to promote osteogenesis. These effects were suppressed by Calhex-231.
Effect of CaSR knockdown during osteogenic differentiation in presence of CaSR agonists
Consistent with the positive effect of CaCl2 and R-568 on markers and molecular regulators of osteogenesis, CaSR expression levels increased after CaCl2 and R-568 exposure during osteogenic differentiation. This was revealed by flow cytometry in both permeabilizing (Fig. 7A) or nonpermeabilizing (Fig. 7B) conditions demonstrating that CaCl2 and R-568 significantly increased both intracellular and membrane CaSR levels, and that this effect was significantly reduced by the allosteric inhibitor Calhex-231 and by the selective inhibitor/antagonist NPS-2143.
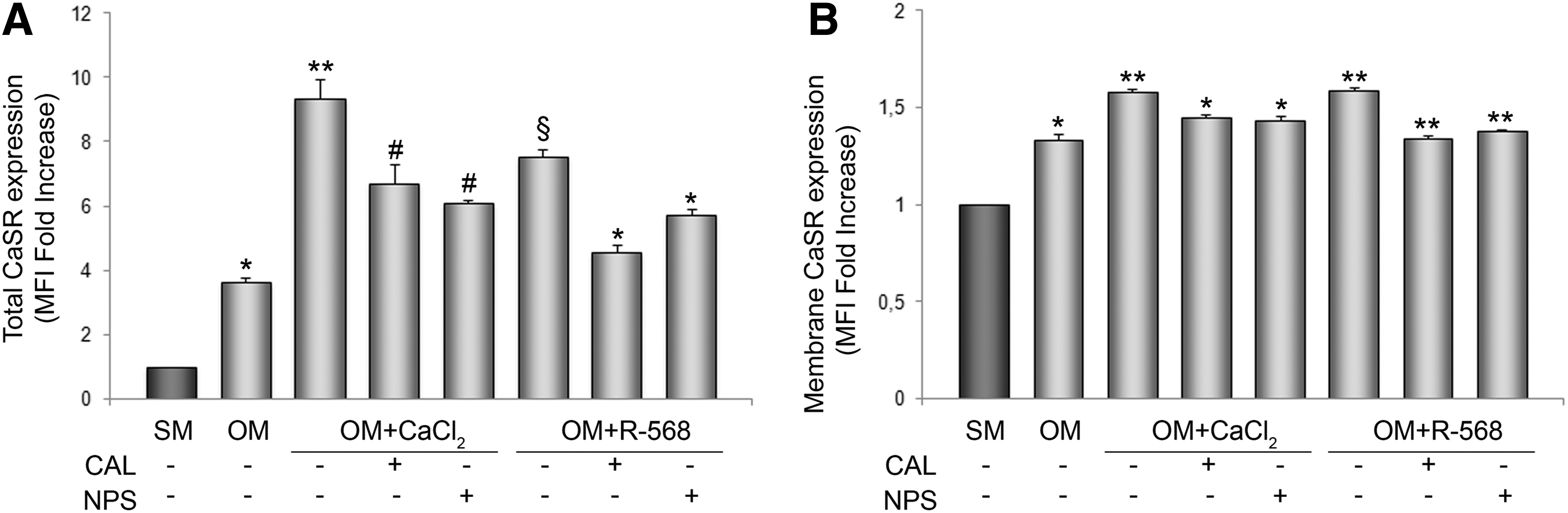
CaSR expression in hAFMSCs evaluated totally and at membrane level. The histogram in
To further investigate the implication of CaSR in supporting calcimimetic induced hAFMSC osteogenic differentiation, we performed gene knockdown experiments with siRNA against CaSR. Cell behavior in terms of proliferation and viability was properly monitored up to 14 days of si-CaSR treatment revealing that the cells were highly suffering if treatment was prolonged after 7 days both in presence and in absence of OM (data not shown). This is not surprising, since depletion is concerning to a critical protein for cell metabolism. On the basis of these observations, we set up the gene knockdown experiments up to 7 days in presence of OM.
The efficiency of gene silencing, monitored at protein level by flow cytometry, revealed that already after 72 h CaSR protein levels were approximately reduced by 50% compared with scrambled siRNA-treated cells, and this was maintained up to 7 days (Fig. 8A). This effect was accompanied by a significant decrease of the expression of the master regulator of osteogenesis, RUNX2, which was reduced to about 30% after si-CaSR treatment (data not shown). The consequence of CaSR knockdown on the positive effect that aforementioned agonists have on β-catenin, SLUG, and NFATc1 was then investigated. RT-PCR analysis revealed that the expression of β-catenin and NFATc1 significantly decreased (Fig. 8B, C), while Slug expression was not affected (data not shown). This evidence together with the effect on RUNX2 expression, suggests that hAFMSCs osteogenic potential and its improvement after CaCl2 and R-568 exposure was reduced after CaSR knockdown, furthering a critical role of CaSR and its agonists on hAFMSC osteogenic differentiation.

Downregulation of CaSR.
Discussion
Accumulating evidence indicates that AF-derived cells collected during pregnancy by routine amniocentesis are a population of multipotent progenitor cells that express both mesenchymal and embryonic stem cells markers and are able to differentiate into specialized cells representative of all three germ layers [1,41]. In particular, their ability to differentiate into bone-producing osteoblasts has been extensively characterized [1]. All these properties and the absence of ethical issues make them signally advantageous for developing new possibilities in regenerative medicine.
In this report we focused on osteogenic potential of these cells. Osteogenesis is a tightly regulated process governed by many factors and mechanisms only known in part. A better understanding of these mechanisms and the identification of possible new molecular modulators is critical for the optimization of cell-based tissue engineering strategies for the treatment of bone tissue defects.
We showed here for the first time that human amniotic fluid mesenchymal stem cells (hAFMSCs) express CaSR with a predominant intracellular distribution, and that osteogenic induction is associated with the upregulation of CaSR expression and cell membrane localization, supporting a potential functional role for CaSR in osteogenic differentiation.
This receptor is expressed in cells populating the mature bone, such as osteoblasts, osteoclasts, stromal cells, monocytes, macrophages, and chondrocytes [17,42 –44], and its role in bone biology has been demonstrated [21,22], but the investigation about the possibility to use CaSR as biological marker or therapeutic target is currently limited.
Recent work by our group, and by others, has detected CaSR expression and activity in progenitor cells, including stem cells from ovine amniotic fluid [20], equine umbilical cord matrix [45], and bone marrow hematopoietic stem cells [46].
In the present instance, we identified CaSR in hAFMSCs. Western blot analysis showed a predominant immunoreactive band of ∼160 kDa, which increased by ∼4-fold upon osteogenic differentiation (Fig. 2A). Notably, these monomers undergo dimerization to give the mature active form of the receptor, which is assembled on the cell surface membrane [20] and indeed, using an antibody against an extracellular CaSR domain, we obtained flow cytometric evidence of enhanced CaSR accumulation at the cell surface during osteogenic differentiation (Figs. 2I and 7B). Together with our previous observation with ovine AFMSCs and with data showing CaSR expression and function in human osteoblasts [19], these results suggest that CaSR may play a role during osteogenesis.
To further support this hypothesis, we performed a series of experiments using CaSR agonists and antagonists. We consistently observed that calcimimetic R-568, but not its S enantiomer, stimulated hAFMSC osteogenic differentiation. This effect was abolished by the CaSR allosteric inhibitor, Calhex-231 (Figs. 3 and 7A, B) and selective inhibitor NPS-2143 (Fig. 7A, B). Taken together these data suggest that responsiveness of hAFMSCs to CaSR agonists improve their osteogenic potential and this evidence may have important implications to develop therapeutic interventions against bone injury.
At the molecular level, R-568 and calcium upregulated the expression of CaSR together with classic osteogenic markers such as ALP, Runx2, and OPN. Notably, they also enhanced the expression of other positive regulators of osteogenesis including β-catenin, which is the canonical mediator of the Wnt pathway, and two transcription factors, Slug and NFATc1, which play a critical role in proliferation and differentiation of osteoprogenitors (Figs. 4 –6 and Supplementary Fig. S2). The positive effects of CaSR agonists on osteogenesis improvement were significantly lost when the cells were CaSR depleted by gene silencing approach, confirming the fundamental role of CaSR in supporting osteogenic differentiation. The impact of CaSR on mechanisms of osteogenesis is certainly quite complex and the involvement of this receptor in the different stages of differentiation that lead from the osteoprogenitors to the mature cells require further investigation. However, because among the markers analyzed, Slug appears to be not affected by cell exposure to si-CaSR, preliminary CaSR knockdown experiments lead us to speculate that Slug might participate only in part to CaSR-mediated commitment toward osteogenesis. Otherwise, CaSR downstream signaling may be mediated by Wnt/β-catenin and NFATc1 in a delicate balance that is highly affected by calcium fluctuations. It is in fact to be taken into account that in bone biology Wnt signaling is mainly considered a key regulatory pathway for bone formation, while the calcineurin-NFATc1 axis is mostly associated with bone resorption, NFATc1 being the master transcription factor for osteoclast differentiation [47]. However, at the same time, Wnt signaling may also influence osteoclastogenesis and NFAT-mediated signaling may regulate osteoblast maturation [48].
In conclusion, our present report provides the first evidence that CaSR is expressed in hAFMSCs, where it appears to control a number of pathways related to osteogenic differentiation.
Taken together, our data may shed new light on mechanisms and pathways that could be targeted when designing novel therapeutic approaches to bone diseases.
Footnotes
Acknowledgments
This study was partially supported by a grant to AP from ERA-AMGEN (USA) and to StemTeCh Group from CARICHIETI Foundation. We thank Dr. Romuald Mentaverri for kindly providing us with siRNA sequences.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.