
Abstract
According to the endosymbiotic hypothesis, the precursor of mitochondria invaded the precursor of eukaryotic cells, a process that began roughly 2 billion years ago. Since then, the majority of the genetic material translocated from the mitochondria to the nucleus, where now almost all mitochondrial proteins are expressed. Only a tiny amount of DNA remained in the mitochondria, known as mitochondrial DNA (mtDNA). In this study, we report that the transfer of mtDNA fragments to the nucleus of pluripotent stem cells is still ongoing. We show by in situ hybridization and agarose DNA two-dimensional gel technique that induced pluripotent stem (iPS) cells contain high levels of mtDNA in the nucleus. We found that a large proportion of the accumulated mtDNA sequences appear to be extrachromosomal. Accumulation of mtDNA in the nucleus is present not only in the iPS cells, but also in embryonic stem (ES) cells. However upon differentiation, the level of mtDNA in the nuclei of iPS and ES cells is substantially reduced. This reversible accumulation of mtDNA in the nucleus supports the notion that the nuclear copy number of mtDNA sequences may provide a novel mechanism by which chromosomal DNA is dynamically regulated in pluripotent stem cells.
Introduction
N
Although the method of reprogramming somatic cells to induced pluripotent stem (iPS) cells by using the SKOM factors (ie, Sox2, Klf4, Oct3/4, c-Myc) is very convenient, most somatic cells expressing these factors fail to complete reprogramming and remain as precursors of stem cells, which often undergo apoptosis, senescence or cell cycle arrest [18 –20]. Many current reprogramming methods have low efficiencies [18,19,21 –23]. Although various reasons for the low reprogramming efficiency have been discussed [20,21], one possibility is that frequent nuclear DNA damage during reprogramming lowers the reprogramming efficiency [20,21,24 –41]. The effect of the oncogenes c-Myc and Klf4 may contribute to this observation [42,43]. Despite these significant changes in the genomic DNA, nuclear trafficking and/or amplification of mtDNA has never been considered as a potential player in the process of reprogramming.
We wanted to test the hypothesis that during reprogramming, fragments of mtDNA migrate to the nucleus and accumulate, which may eventually affect nuclear genomic stability and reprogramming efficiency. In this study, we demonstrate that pluripotent stem cells contain amplified mtDNA sequences in their nuclei, mainly in an extrachromosomal form, and that this accumulation is reversible in pluripotent stem cells subjected to differentiation.
Materials and Methods
Cell lines, generation of iPS cells, differentiation of pluripotent stem cells
Two and three independent mouse iPS and embryonic stem (ES) cell lines were used, respectively. We used mouse embryonic fibroblasts (MEFs) derived from C57BL/10 mice for reprogramming. This iPS cell line was generated by expressing Sox2, Klf4, and Oct3/4 (SKO factors) on a single plasmid, which was then introduced into fibroblasts by lentivirus transduction [44]. Expression of c-Myc was omitted in this cell line. iPS cell colonies were identified essentially as previously described by the Yamanaka laboratory [18,19]. The second iPS cell line was originally generated by expressing all four reprogramming factors Sox2, Klf4, Oct3/4, and c-Myc (SKOM factors) individually on plasmids, which were introduced into MEFs by retrovirus transduction (generously provided by Dr. Rudolf Jaenisch, Whitehead Institute, Cambridge) [45]. This iPS cell line expresses a GFP gene driven by the Oct4-promoter. The MEFs were passaged three times and the iPS cells about 15 times before analysis. The Rosa 26 ES cell line, which was generously provided by Dr. Philippe Soriano at the Mount Sinai School of Medicine, NY, was passaged multiple times (Fig. 3) [46]. A second cell line (MUBES-01201; Cyagen Biosciences), which was passaged 25 times, expressed a red fluorescent protein (Supplementary Fig. S5; Supplementary Data are available online at
Fluorescence in situ hybridization
To visualize mtDNA in mammalian cells, we applied a protocol for fluorescence in situ hybridization (FISH) similar to one described in Caro et al. [17]. In some experiments mitochondria were labeled with the live cell stain MitoTracker Red® (Invitrogen) before fixation of the cells in 4% paraformaldehyde. Cells were permeabilized with 0.2% Triton X-100 for 20 min at room temperature (RT) and RNA was digested using 1 μg/mL RNaseA for 1 h at 37°C. Cells were blocked (prehybridization) with 4% bovine serum albumin (BSA) for 1 h and then dehydrated in 70%, 90%, and 100% ethanol. The hybridization solution consisted of 70% deionized formamide (Sigma), 12 mM Tris-HCl (pH 8), 5 mM KCl, 1 mM MgCl2, 0.001% Triton X-100, and 0.25% acetylated BSA (Sigma). Cells attached to a coverslip were put upside down on a slide having a drop (25 μL) of hybridization liquid containing the labeled DNA probes at an approximate concentration of 2 ng/mL. DNA was denatured by putting the slide on a heat block (80°C) for 5 min followed by cooling down on the bench. The hybridization was continued in a moist chamber at RT overnight. The cells on the coverslips were washed with 70% formamide and 2× SSC four times for 10 min each, and then covered with the mounting medium containing DAPI (Fluoroshield™; Sigma). Cells were analyzed using an epifluorescence microscope (Nikon Eclipse 80i) equipped with a CCD camera (DS-Qi1).
DNA two-dimensional gel electrophoresis
We purified nuclear and mtDNA as previously described [50]. We used the mild detergent NP-40 to remove mtDNA molecules, which are present in mitochondria attached to the purified nuclei. Nuclear and mtDNA were then analyzed by two-dimensional (2D) agarose gel electrophoresis technique as previously applied [51]. DNA samples were separated on a 0.31% agarose gel for 24 h at 1 V/cm in the first dimension and on a 0.8% agarose gel for 48 h at 1.5 V/cm in the second dimension. Both agarose gels contained 0.5 μg/mL ethidium bromide. The 2D gel membranes were hybridized to a 32P-labeled mtDNA probe containing fragments #1 to #5 (see below).
Sequences of primers used in PCR and in preparing the DNA probes
mtDNA fragment #1 (12S rRNA) (Primer 69 F mouse mtDNA: 5′-CAAAGGTTTGGTCCTGGCCT; primer 790 R mouse mtDNA: 5′-TGTAGCCCATTTCTTCCCA)
mtDNA fragment #2 (16S rRNA) (Primer 2100 F mouse mtDNA: 5′-CTTTAATCAGTGAAATTGACCTTTCAG; primer 2650 R mouse mtDNA: 5′-CGTATATATTTTATTTAGATTTTATTCATAAATTAAG)
mtDNA fragment #3 (Cox1) (Primer 5215 F mouse mtDNA: 5′-CACCTTCGAATTTGCATTCG; primer 5709 R mouse mtDNA: 5′-CTGTTCATCCTGTTCCTGCT)
mtDNA fragment #4 (ATPase 6) (Primer 8032 F mouse mtDNA: 5′-CGCCTAATCAACAACCGTCT; primer 8497 R mouse mtDNA: 5′-TGGTAGCTGTTGGTGGGCTA)
mtDNA fragment #5 (ND5) (Primer 12777 F mouse mtDNA: 5′-CATAGCCTGGCAGACGAACA; primer 13435 R mouse mtDNA: 5′-GAGGTGGATTTTGGGATGGT)
DNA fragment GAPDH (Primer 135961 mouse GAPDH-F: 5′-ATCACGCCACAGCTTTCCAGAG; primer 136700 mouse GAPDH-R: 5′-GCAAAGTGGAGATTGTTGCCATCAA)
To amplify the ∼12 kb mtDNA fragment we used the Takara LA Taq® polymerase and primers 12777 F mouse mtDNA and 8497 R mouse mtDNA (see above for the sequence).
Cy3-labeled PNA mammalian telomere probe (generously provided by Dr. Utz Herbig, Rutgers Biomedical and Health Sciences, Newark).
For the FISH PCR-generated DNA fragments were labeled directly with Alexa Fluor® 488 or Alexa Fluor® 546 using the ULYSIS® nucleic acid labeling kit (Molecular Probes).
The PCR fragments amplified from nuclear MEF and iPS cell genomic DNA were sequenced at the company Macrogen USA Corp. For the sequence alignment we used BLASTN (Basic Local Alignment Search Tool).
Statistical analysis
We repeated each experiment at least three times and we used the Student's t-test to calculate P values.
Results and Discussion
The number of mtDNA sequences with nuclear localization, which were visualized by FISH, increase during aging in rat liver and brain [17]. Given that mtDNA fragments may translocate from mitochondria to the nucleus and potentially disrupt specific nuclear genes by insertion mutagenesis, we investigated whether mtDNA fragments also accumulate in the nucleus during reprogramming [1]. We used the same FISH protocol that was used to detect mtDNA insertions in nuclear DNA of brain and liver tissues of rats [17]. The mtDNA probes were amplified by PCR using mtDNA purified from cardiac mouse mitochondria, and labeled by covalently linking fluorophores (see Materials and Methods section). All five mtDNA probes (#1– #5) together were used in the hybridization experiments. General cellular DNA was stained with DAPI, and the mitochondria were visualized using the dye MitoTracker Red (Invitrogen) before fixation of the cells with formaldehyde.
We used mouse embryonic fibroblasts (MEFs) and iPS cells derived from MEF cells expressing the three reprogramming factors Sox2, Klf4, and Oct3/4 (SKO) [19,52]. In fibroblasts, the cytosolic mtDNA signals, which were likely derived from mtDNA within the mitochondria, were hazy most likely because the cells were treated with RNase before hybridization (Fig. 1A and Supplementary Fig. S1). Several discrete signals indicating mtDNA-like sequences were observed in the nucleus, which colocalized with the nuclear DAPI staining and looked also similar as previously described by Caro et al. [17]. The amount of mtDNA in the nucleus was clearly elevated in iPS cells as determined by densitometric scanning of the fluorescence images using the program NIH ImageJ. Similar results were obtained with iPS cells derived from MEFs expressing all four reprogramming factors Sox2, Klf4, Oct3/4, and c-Myc (SKOM) (Supplementary Fig. S2) [19,52]. Although there were only one or two strong focal sites visible in iPS cells instead of several spots in fibroblasts, measurements of the intensities of the nuclear signals derived from the mtDNA-Alexa Fluor 488 stained regions demonstrated that iPS cells (SKO, Fig. 1A) contain ∼4.7-fold (P<0.005) more mtDNA with nuclear localization compared to fibroblasts (Fig. 1B). The nuclear mtDNA signals in both fibroblasts and iPS cells appeared to occupy rather large areas in the nucleus compared to single chromosomal sequences. For comparison, we detected mtDNA and telomeric sequences in the same cells (Fig. 1C). Normal diploid mouse cells contain 40 chromosomes which equals to 80 telomeric signals. Telomeric sequence was detected with a Cy3-labeled telomere probe; mtDNA was visualized using Alexa Fluor 488-labeled mtDNA fragments. Given that the average length of telomeres in laboratory mice is between 50 to 100 kb, the telomere signals (Fig. 1C, red signals) were overall considerably smaller than many of the nuclear mtDNA signals (Fig. 1C, green signals). In addition, we detected the single nuclear GAPDH gene in fibroblasts and iPS cells using Alexa Fluor 488-labeled GAPDH-encoding DNA fragments. The GAPDH/FISH signals had comparable intensities in fibroblasts and iPS cells, but were also considerably smaller than the nuclear mtDNA signals shown in Figure 1 (Supplementary Fig. S3).
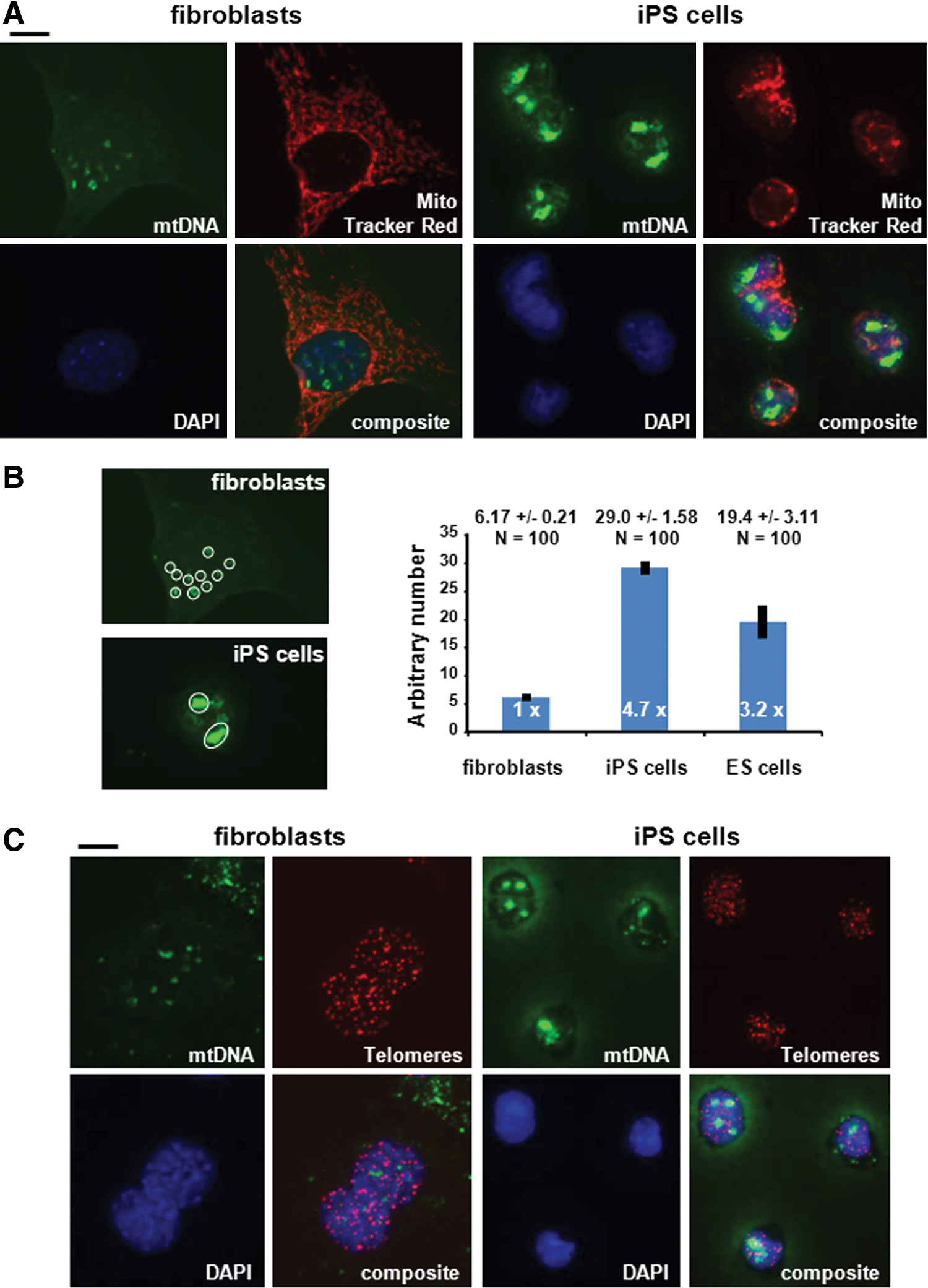
mtDNA sequences accumulate in the nucleus of iPS cells.
Previous analyses of genome sequences of iPS cells did not predict any de novo insertions of mtDNA fragments into nuclear DNA during reprogramming or that extensive amplification of mtDNA sequences within the chromosomal DNA sequence occurs [26,27,39,53]. To investigate whether some of the mtDNA sequences in the nucleus of iPS cells are present in extrachromosomal form, we analyzed fibroblasts and iPS cells using the 2D agarose gel electrophoresis, which allows the separation of the circular form from linear DNA molecules (Fig. 2A, cartoon in left panel) [51]. We isolated nuclei from fibroblasts and iPS cells as previously described [50]. Since crude nuclei still have some mitochondria attached to their surface, we removed the mtDNA, which is present in these mitochondria, from the nuclei by treatment with the mild detergent NP-40 (Fig. 2A, right panel). We analyzed the undigested nuclear DNA by 2D gel electrophoresis and hybridized the membrane to a radioactive-labeled mtDNA probe. The majority of the mtDNA probe is hybridizing to linear nuclear DNA of both fibroblasts and iPS cells (Fig. 2B, left four panels). We propose that these sequences are not extrachromosomal, but are rather part of chromosomal DNA, for example in the form of NUMTs which are mtDNA sequences dispersed throughout chromosomal DNA [1]. In the nuclear DNA of iPS cells, we observed an approximate sixfold increase compared to fibroblasts (determined by phosphorimager) of a DNA species, which does not appear to enter the gel (top left side). It is possible that these molecules are mtDNA circles linked to each other in the form of concatemers as previously described [54,55]. In the iPS gel there is a further arc above the arc of linear DNA fragments, which may also indicate circular mtDNA species of various sizes. As a control, we also analyzed mtDNA derived from the mitochondrial fraction by 2D gel electrophoresis (Fig. 2B). mtDNA has a size of about 16 kb and is mainly present in circular form as a monomer in mitochondria. The full-length circular DNA appears as a homogenous DNA species (visualized as a spot) in the 2D gel, thus mtDNA in mitochondria is mainly present in monomeric, circular form. The weak signal on the arc of linear DNA fragments indicates likely contamination from nuclear DNA. The mtDNA gels also clearly indicate that the nuclear DNA samples are mostly free of full-length mtDNA molecules, which derive from mitochondria attached to the nucleus. As another control, we reprobed the nuclear DNA gels with a probe against the single-copy gene GAPDH. No extrachromosomal GAPDH DNA circles are visible in the iPS gel. To determine further, the size of the highly abundant mtDNA sequences, we amplified short (0.5–0.8 kb) and long (∼12 kb) mtDNA sequences by PCR (Supplementary Fig. S4). Whereas there was an approximately two- to fivefold higher number of short mtDNA fragments in the nuclei of iPS cells compared to fibroblasts, large mtDNA fragments (∼12 kb) were not enriched in the nuclei of iPS cells suggesting that large or even full-length mtDNA molecules are not amplified during reprogramming in the nuclei of iPS cells. We determined the sequences of the short mtDNA fragments (see Supplementary Fig. S4), which were amplified by PCR from MEF and iPS cell nuclear genomic DNA. All sequences of the amplified PCR fragments are identical to mtDNA sequences (Supplementary Sequence). We conclude that in the nuclei of iPS cells a high number of short mtDNA sequences are mainly present as extrachromosomal circles, although we cannot exclude that nuclear mtDNA insertions occur during reprogramming as well.
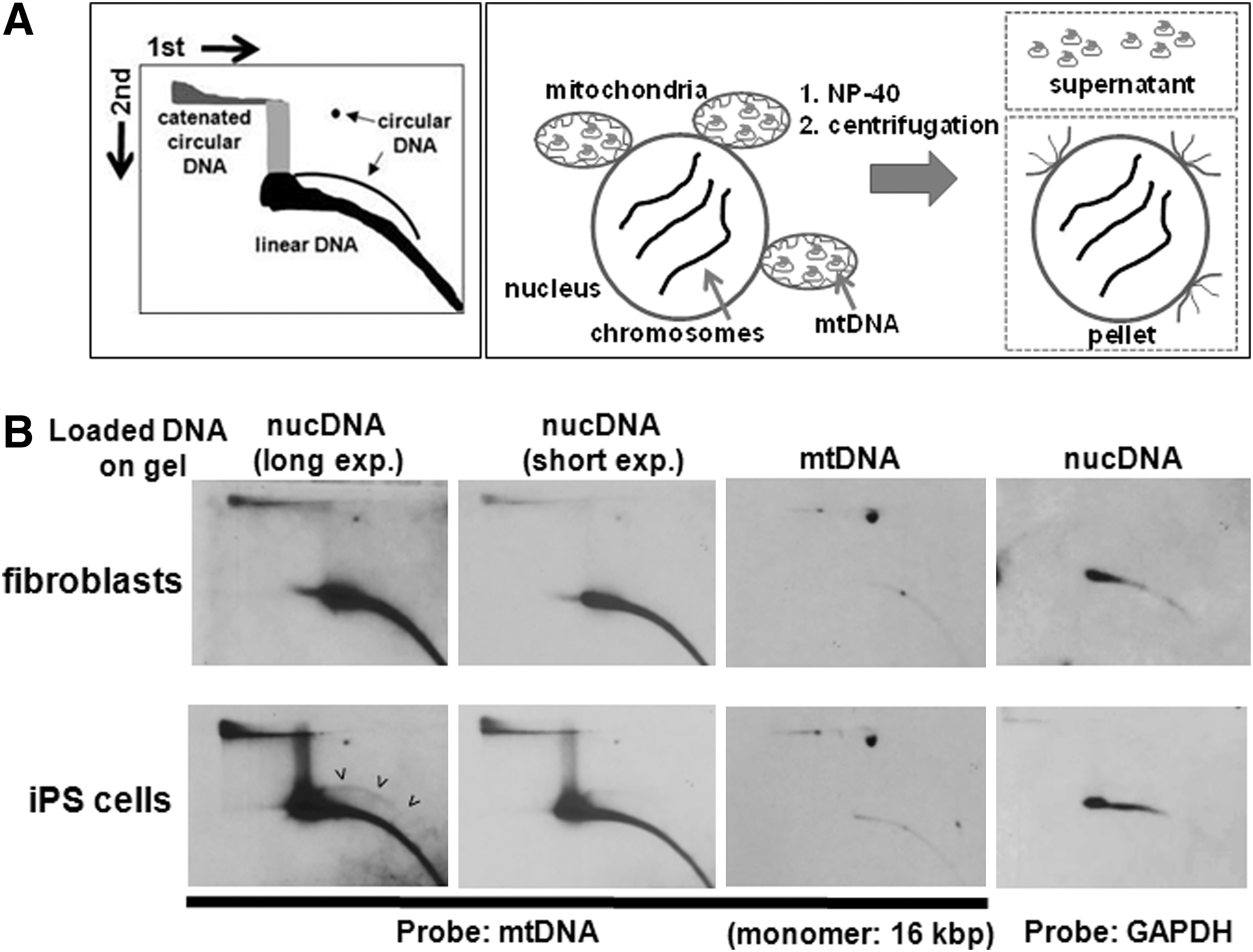
The accumulation of nuclear mtDNA sequences in iPS cells is mainly of extrachromosomal nature.
Like iPS cells, ES cells also exhibit culture-induced copy number variations in chromosomal DNA [53]. Therefore, we investigated whether mtDNA sequences with nuclear localization are present in elevated numbers in two different ES cell lines. We demonstrated that there was a significant increase (3.2-fold; P<0.005) in nuclear mtDNA sequences in ES cells compared to fibroblast cells (Fig. 3 and Supplementary Fig. S5; Fig. 1B for quantification).
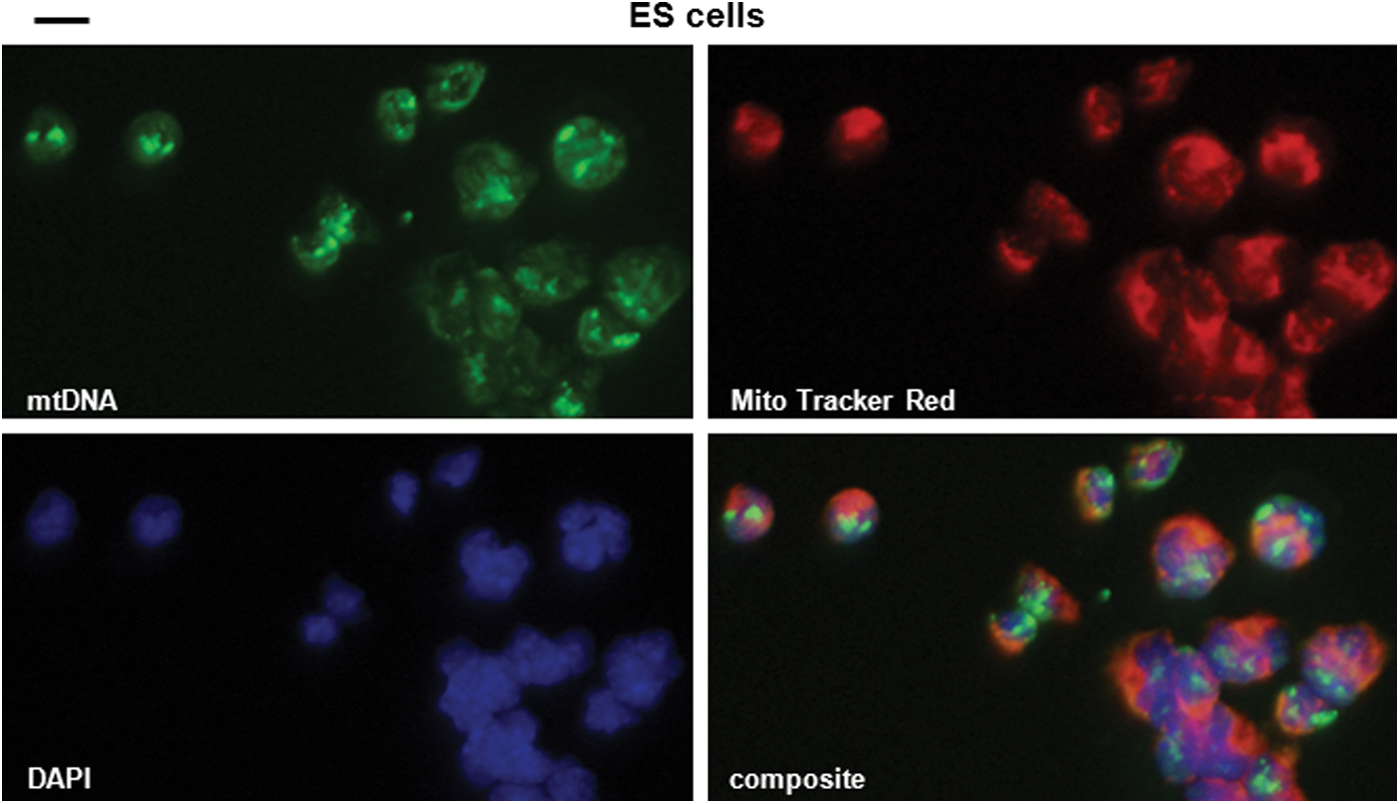
mtDNA sequences accumulate in the nucleus of ES cells. Mouse ES cells (Rosa 26) were processed for in situ hybridization. Nuclear staining for mtDNA superimposes with nuclear DAPI staining. Green: mtDNA (Alexa Fluor 488-labeled mtDNA fragments); blue: DNA (DAPI); red: mitochondria (MitoTracker Red®). Scale bar: 5 μm. Color images available online at
Given that the number of mtDNA sequences in the nucleus increases during reprogramming, we were interested in ascertaining whether these accumulated sequences remain at similar levels when the stem cells initiate differentiation into somatic cells. When we induced differentiation in pluripotent stem cells (iPS and ES cells) to somatic cells using the hanging drop method [47], the accumulated mtDNA species in nuclei of this heterogeneous population of differentiating cells decreased to similar levels as observed in fibroblasts (Fig. 4). Thus, the process of the accumulation of mtDNA sequences in nuclei is reversible.
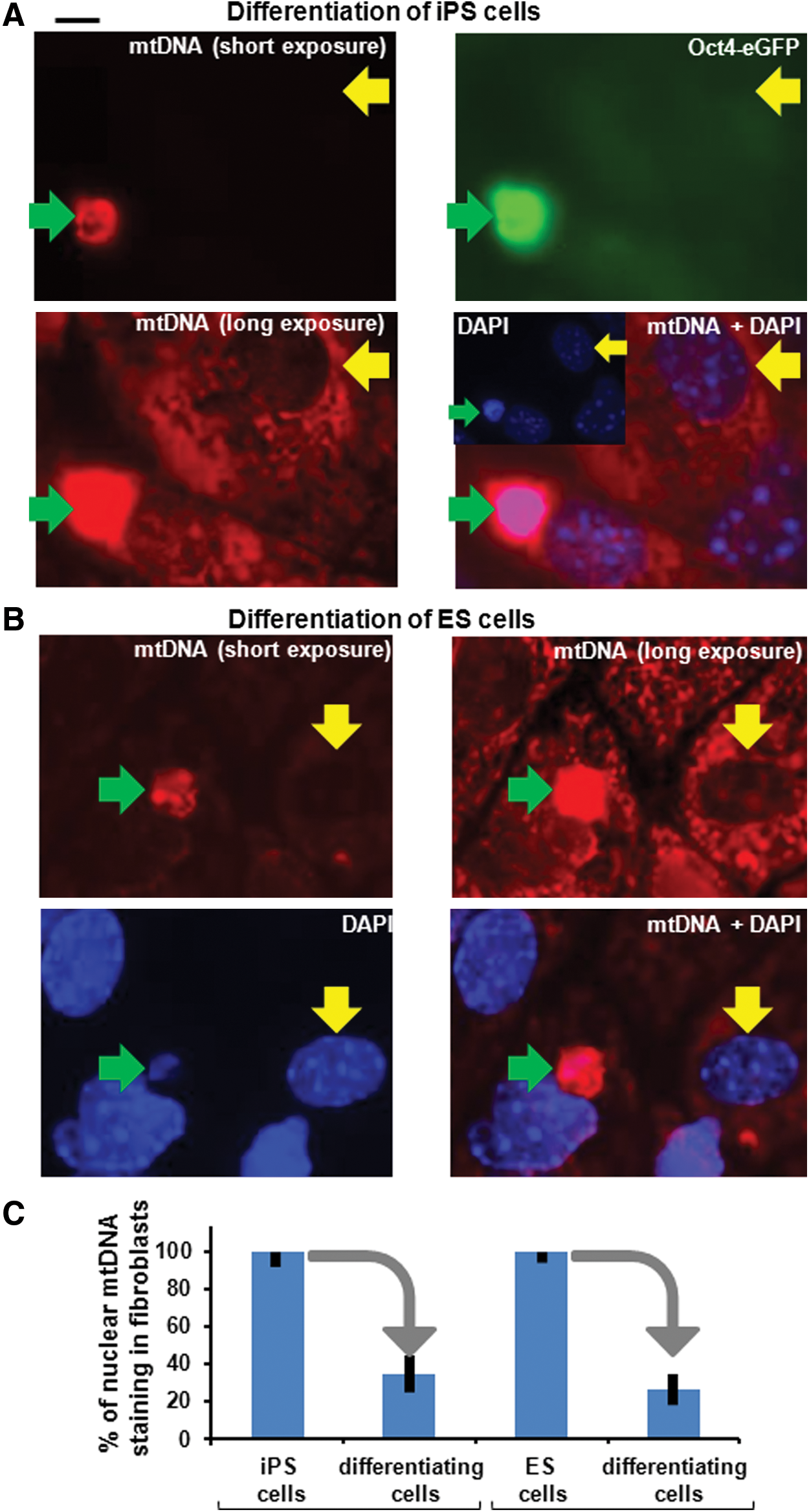
The accumulation of nuclear mtDNA sequences is reversible after inducing differentiation of iPS and ES cells.
In summary, we demonstrate that mtDNA sequences with nuclear localization are present in elevated numbers in both iPS and ES cells, but decreases when the pluripotent stem cells undergo differentiation to somatic cells. Most of the nuclear-amplified mtDNA sequences appear to be present in the extrachromosomal form. We envisage three possible mechanisms by which mtDNA enters into the nucleus and/or amplifies: (1) Fragmented mtDNA may translocate to the nucleus. If the mtDNA fragments enter the nucleus in the linear form they may either insert into the chromosomal DNA, which may occur during the repair of a dsDNA break in chromosomal DNA, or be converted to circular DNA molecules with the help of a DNA ligase (eg, Lig4 of the non-homologous end-joining pathway). (2) Alternatively, a few circles with mtDNA sequences may already be present in the nucleus and those may amplify during reprogramming. This is a likely possibility since in fibroblasts mtDNA sequences are already present in the nucleus (see Figs. 1 and 2), and some of these appear to be extrachromosomal (Fig. 2). Further support for this scenario comes from our observation, if mtDNA is removed from mitochondria using ethidium bromide, the accumulated mtDNA fragments still remain in about the similar amount in the nucleus suggesting that the translocation of mtDNA fragments to the nucleus appears rather not to be a requirement for the amplification of nuclear mtDNA fragments (Supplementary Fig. S6). The extrachromosomal mtDNA circles may be maintained and inherited in the nucleus by break-induced replication mechanisms [56,57]. (3) Another possible scenario is that a few single fibroblasts with already highly amplified mtDNA molecules in the nucleus may be the preferential source for the reprogramming to iPS cells.
The number and variety of extrachromosomal circular DNA in the nucleus increase enormously in tumor tissues and in cells exposed to carcinogens [58 –61]. Overrepresentation of these circular DNAs may disturb chromosomal DNA replication and/or repair of damage in chromosomal DNA. A similar scenario might be linked to the amplification of mtDNA sequences in iPS cells. Expression of the reprogramming factors and oncogenes Klf4 and c-Myc may initiate the amplifications of mtDNA sequences in the nucleus and potentially contribute in this way to an increase in nuclear genomic instability. This is a possibility since deregulated c-Myc expression itself contributes to genomic instability, including the formation of extrachromosomal DNA circles [61].
Conclusions
We demonstrate in this study that pluripotent stem cells contain high numbers of mtDNA sequences in the nucleus, which decrease upon differentiation to somatic cells. This unexpected observation during reprogramming highlights a new feature of pluripotent stem cells, and may represent another level of chromosomal regulation. It is however unclear, whether high levels of mtDNA circles in the nucleus inhibit or improve reprogramming, which will be tested in future experiments.
Footnotes
Acknowledgments
We are grateful to Dr. Rudolf Jaenisch, Whitehead Institute, Cambridge, for providing the Oct4-GFP iPS cells, Dr. Utz Herbig for providing the Cy3-labeled PNA telomere probe, Drs. Yanfei Yang and Junichi Sadoshima for providing GAPDH mouse DNA for labeling, Dr. Lin Yan for providing purified mitochondria derived from mouse hearts. We also thank Dr. Carolyn Suzuki for critical reading of the manuscript and Drs. Sadoshima, Toruner, Tyagi for discussions throughout the project. This work was supported by the NIH (J.S.S., D.F., and E.S.R.), the Muscular Dystrophy Association (D.F.), the Hispanic Center of Excellence (D.F.), and by departmental support (A.S.I. and D.F.).
Author Disclosure Statement
The authors indicate no potential conflict of financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.