
Abstract
Temporally distinct populations of neural stem cells (NSCs; embryonic and adult) display the cardinal stem cell properties of self-renewal and multipotentiality; however, their relative frequency and cell kinetics vary through development and into old age. We asked whether changes in NSC behavior could be accounted for by changes in environmental signals over time. We identified a prosurvival signaling cascade that enhances adult-derived NSC survival using cues released from embryonic neurons. Specifically, we demonstrate that stromal-cell-derived factor-1α (SDF-1α) released by embryonic neurons leads to upregulation of neuronal nitric oxide synthase in adult neural precursor cells. The resulting increase in nitric oxide leads to the upregulation of the stem cell factor (SCF) receptor ckit on adult NSCs (ANSCs). SCF released from embryonic neurons results in enhanced NSC survival. Using both in vitro and in vivo assays, we have demonstrated expansion of the size of the NSC pool through this pathway, indicating that ANSCs retain their ability to respond to embryonic-derived cues into adulthood.
Introduction
N
The environment, or “niche,” is highly specified and varies through development and into adulthood [7 –9]. Notably, the pattern of expression of signaling molecules and their receptors varies temporally within the niche. For instance, stem cell factor (SCF) is expressed in the embryonic NSC (ENSC) niche [10] but is not found in the adult NSC (ANSC) niche under baseline conditions [11]. The interplay and balance between the various cell populations that comprise the niche can also dictate cell behavior. In this regard, it has been shown that ablation of NSC progeny leads to activation and recruitment of NSCs into cell cycle [12,13]. Ependymal cells in the NSC niche release Noggin that impacts NSC behavior by promoting increased neuroblast formation and inhibition of glial differentiation [9,14]. Conversely, GABA released from migrating neuroblasts in the adult forebrain has been shown to decrease ANSC proliferation [15]. During development, cardiotrophin-1 (CT-1) released from newborn embryonic neurons feeds back on progenitors to instruct astrocyte differentiation [16]. Hence, cell–cell interactions from niche cells and released factors can regulate NSC behavior and these interactions vary through development and into adulthood.
Herein, we demonstrate that ANSC survival can be enhanced 2-fold by the reintroduction of factors derived from the embryonic brain during development. Specifically, ENSC progeny, the embryonic neurons, provide cues that regulate NSC survival and induce the expression of receptors that enable ANSCs to respond to prosurvival cues. We have identified a prosurvival cascade of events that demonstrate that ANSCs remain responsive to embryonic-derived factors in both in vitro and in vivo assays. The prosurvival cascade is initiated by the release of stromal-cell-derived factor-1α (SDF-1α) from embryonic neurons. The SDF-1α activation results in upregulation of neuronal nitric oxide synthase (nNOS) and nitric oxide (NO) production in ANSCs. Increased NO leads to upregulation of ckit, the receptor for SCFs, and enhanced cell survival in the presence of SCF derived from embryonic neurons. We demonstrated specificity using function-blocking antibodies to SDF-1α or SCF, ckit-knockout mice (KitW/KitWv) or nNOS−/− mice, all of which failed to elicit enhanced ANSC survival. Finally, we demonstrate that intraventricular infusion of embryonic neural precursor conditioned media (CM) or SDF-1α and SCF into adult brains results in increased numbers of stem cells isolated in the neurosphere assay. This expansion in the size of the stem cell pool following the in vivo manipulation is consistent with our in vitro mechanism of enhanced cell survival.
Materials and Methods
Mice
All mice were housed at the University of Toronto animal facilities and maintained in accordance with the Animal Care Committee and Institutional guidelines. Adult male CD1 and C57/BL6 mice (8–10 weeks; Charles River) and pregnant female CD1 mice at gestational age of E14/15 (Charles River) arrived from supplier as needed and all surgical animals were allowed to acclimate to the facility for 1 week prior to use. Adult YFP-expressing mice (Strain No. 5483; Jackson Laboratory), nNOS−/− mice (Strain No. 2986; Jackson Laboratory), KitW (Strain No. 0692; Jackson Laboratory), and KitWv (Strain No. 0049; Jackson Laboratory) were bred in house and both male and female mice were used at 2–4 months of age. Tα1:nlacZ mice was obtained from Dr. Freda Miller (Hospital for Sick Children, Toronto).
Neurosphere assay
NSCs were isolated following dissection of the periventricular region of the embryonic or adult mice as described previously [1,17]. Adult dissections underwent enzymatic treatment prior to mechanical dissociation. Cells derived from adult and embryonic dissections were immediately plated in 24- or 6-well tissue culture plates (NUNC) in 500 μL or 2 mL, respectively, of serum-free media (SFM) supplemented with epidermal growth factor (EGF, 20 ng/mL; Sigma), fibroblast growth factor (FGF, 10 ng/mL; Sigma), and heparin (2,000 ng/mL). Cells were plated at clonal (≤10 cells/μL) or 30–50 cells/μL for collection of CM. A1-only cultures were plated at 1 cell/μL that resulted in a final number of 500 cells per well. Cocultures were plated at a final density of 10 cells/μL. After 7 days in vitro, the numbers and size of individual neurospheres were assayed. Individual neurospheres were passaged by dissociating into single cells and replating in SFM+epidermal growth factor, fibroblast growth factor, and heparin (EFH) conditions to assay for the numbers of secondary neurospheres. Neurosphere size was determined using Image J software. Cell survival in primary cultures was assayed using trypan blue exclusion immediately after plating and at 24-h intervals by collection of media from six-well plates initially plated with 20,000 cells per well.
Conditioned media
CM from primary embryonic, passaged embryonic, and primary adult cultures were derived from high-density (30–50 cells/μL) cultures at 48 h after plating. The media were aspirated, filtered through a 0.22-μm filter (Millex), and used immediately or stored at −80°C. CM were supplemented with EFH prior to plating.
Neuron CM were generated from Tα1:nlacZ mice as described previously [16]. Briefly, the cortices of E16/17 embryos were dissected and sorted for LacZ expression using FluoReporter lacZ Flow Cytometry Kit (Invitrogen/Life Technologies). The LacZ-positive cells were plated on six-well plates (NUNC) coated in poly-
Endothelial cell CM were derived from a mouse brain endothelium cell line (ATCC No. CRL-2299). Cells were cultured to 70% confluence in Dulbecco's modified Eagle's medium+10% fetal bovine serum at which time the media were removed and replaced with SFM+EFH. Following two additional days of incubation, the media were collected, filtered, and used immediately or stored at −80°C.
Enzyme-linked immunosorbent assay
To determine concentrations of SCF and SDF-1α in CM, enzyme-linked immunosorbent assay (ELISA) kits (RayBiotech) were used following the manufacturer's instructions. Plates were read on a fluorometric spectrometer (VERSAmax) at 450 nm. The optical density of each sample was compared with the standard curve for each molecule.
Reverse transcription-polymerase chain reaction
RNA was extracted from neurospheres or cells using the protocol established in the Qiagen RNeasy extraction kit (Qiagen). RNA was normalized across samples and cDNA was derived using a SuperScript First-Strand Synthesis System (Invitrogen/Life Technologies). Samples were processed in a GeneAmp PCR System 9700 (Applied Biosystems) at 25°C for 10 min, 42°C for 50 min, 70°C for 15 min, and stored at 4°C until use. Samples were prepared using the manufacturer's specifications. Samples were run at 94°C for 5 min and cycled 35 times at 94°C for 30 s, 60°C for 30 s, and 72°C for 30 s, followed by 72°C for 7 min. Polymerase chain reaction (PCR) samples were run on 2% agarose gels at 100 V for 30 min. Primers for the PCRs are as follows:
ckit (forward): 5′-GTGCAGTCCTGGTTTGGATT-3′
ckit (reverse): 5′-ACAACTCACCCACACGCATA-3′
CXCR4 (forward): 5′-GGTCTGGAGACTATGACT CCA-3′
CXCR4 (reverse): 5′-GTGCTGGAACTGGAACACCA-3′
nNOS (forward): 5′-CCTTAGAGAGTAAGGAAGGGG GCGGG-3′
nNOS (reverse): 5′-GGGCCGATCATTGACGGCGAGA ATGATG-3′
Exogenous addition of SCF, SDF-1α, L-NAME, and Spermine NONOate
SCF (Chemicon/Millipore) was added immediately upon plating of A1 secondary cultures from A1 alone or A1E9 cocultures at a range of concentrations from 1 to 100 ng/mL. SDF-1α (Raybiotech) was added to Al-only cultures at doses from 0.78 to 100 pg/mL. The pan-nitric oxide synthase (NOS) inhibitor L-NAME (Sigma) was added to A1 or A1E9 cultures every 6 h for 48 h to maintain a concentration of 0.1–100 μM. The NO donor (Spermine NONOate; Sigma) was added to A1-only cultures every 90 min for the first 12 h following plating.
Fluorescence-activated cell sorting
Primary adult neurospheres plated at 1 cell/μL and grown in the presence or absence of 2 ng/mL SCF were collected in bulk and mechanically dissociated into single cells. Cells were passed through a 40-μm cell strainer and spun down at 1,500 rpm for 5 min. Supernatant was removed and cells were resuspended in phosphate-buffered saline (PBS) at a final concentration of 1×106 cells/mL. Cells were kept in ice and stained for 1 h at 1 μg/mL with a PE-conjugated antibody to ckit (SC-53147; Santa Cruz Biotechnologies). Cells were washed twice with cold PBS. Samples were sorted using an FACSAria (BD). Gates were set based on the negative control (unstained cells). Cells expressing high or low levels of ckit were sorted into separate tubes and spun at 1,500 rpm for 5 min. Pellets were resuspended and cells were plated at a final density of 1 cell/μL in SFM+EFH only and allowed to grow for 7 days at 37°C.
In vivo infusions
Adult 10-week-old CD1 or C57/BL6 male mice were anesthetized with isofluorane and secured in a stereotaxic apparatus. Cannula were implanted at 0.2 mm anterior and 0.8 mm lateral to Bregma. Mini-osmotic pumps (1007D; Alzet) were placed subcutaneously over the shoulder of the mouse. Pumps contained embryonic CM, adult CM, artificial cerebral spinal fluid (CSF) (aCSF; sham), SCF (20 ng/mL intrapump concentration), low-dose SDF+SCF (100 pg/mL SDF and 40 ng/mL SCF), or high-dose SDF+SCF (200 pg/mL SDF and 80 ng/mL SCF). Pumps infused solutions at a rate of 0.5 μL/h for 5 or 7 days. At 5 or 7 days, mice were cervically dislocated and the periventricular region in each hemisphere was dissected and cultured in the neurosphere assay as described. Control mice were sham operated.
Statistics
All data are represented as average±SEM or average fold increase from control±SEM. Statistical differences between groups were assessed by two-tailed Student's t-test or one-way ANOVA with Bonferroni post hoc test.
Results
An overwhelming embryonic environment increases the number of ANSCs isolated in vitro
Neurospheres grown at clonal densities are derived from single NSCs; hence, the number of neurospheres indicates the total number of stem cells [17 –19]. The size of individual neurospheres represents the size of the progenitor pool as progenitors comprise the vast majority of cells within the neurosphere [18 –20]. To this end, we looked at the formation of neurospheres over development and into old age. Consistent with others, we observe a decline of neurosphere formation in adulthood, which is even more attenuated in old age (Fig. 1A). However, the lack of neurosphere formation is not conclusively the result of less NSCs in these brains, but rather a reflection of the responsiveness of the NSCs to cues (unpublished observations).
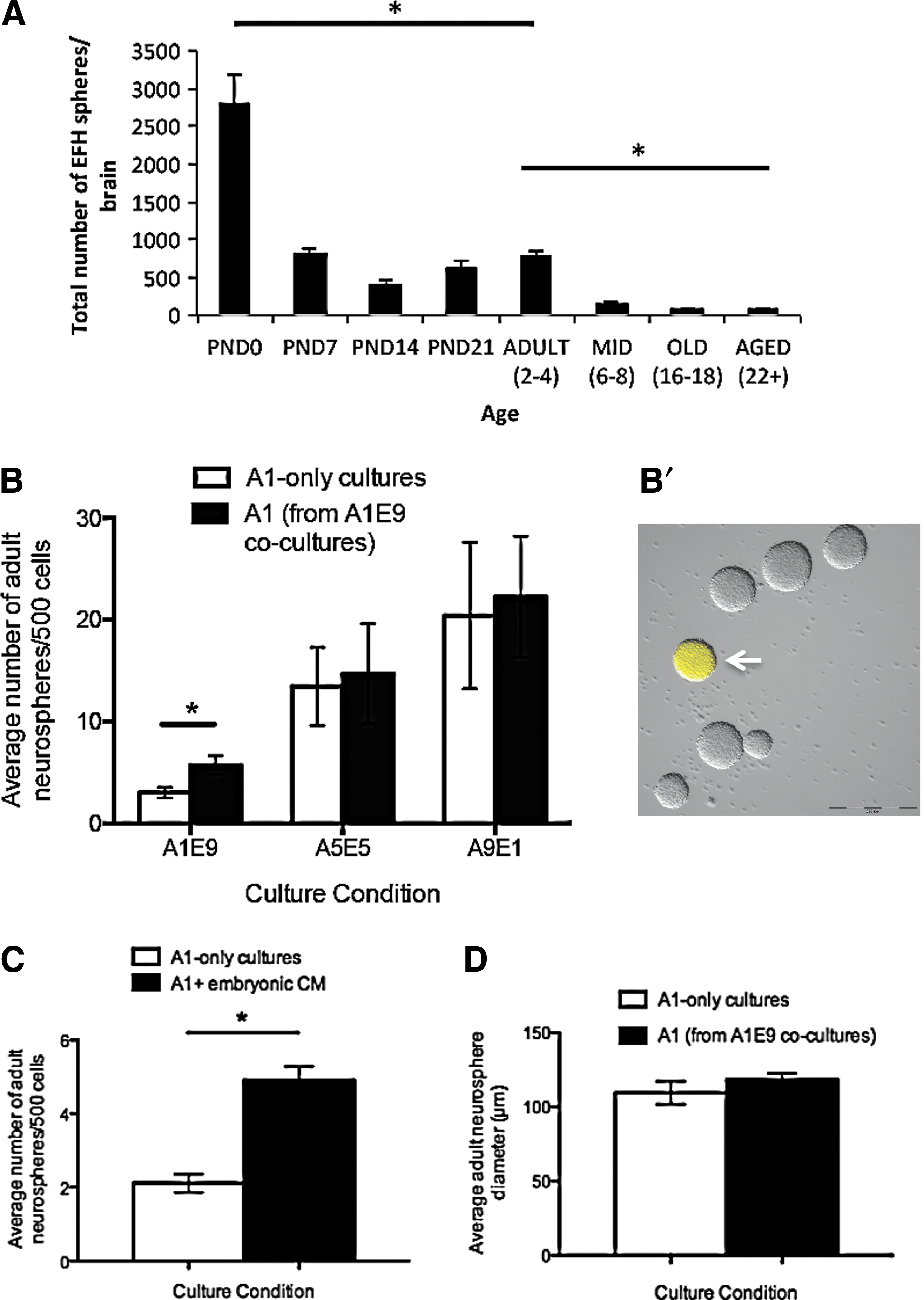
An overwhelming embryonic environment leads to increased numbers of adult neurospheres in vitro.
To examine the role of the environment on NSC behavior, we established an in vitro coculture assay that looks at neurosphere formation. We performed primary dissections of adult and embryonic periventricular regions and cocultured the temporally distinct populations at different plating ratios while maintaining the final plating density at 10 cells/μL to ensure the generation of clonally derived neurospheres [17]. Briefly, cells from primary dissections of the adult periventricular region of YFP-expressing transgenic animals (YFP+) and cells from embryonic day 14.5 (E14.5) CD1 germinal zone were plated in standard neurosphere conditions (ie, in the presence of EGF and FGF) in the following groups: (1) low adult (1 cell/μL) and high embryonic (9 cells/μL) (A1E9, adult in embryonic environment), (2) equal ratio (5 cells/μL of each) (A5E5), or (3) high adult (9 cells/μL) and low embryonic (1 cell/μL) (A9E1, embryonic in adult environment). The numbers of adult YFP+ neurospheres that formed in cocultures were counted after 7 days and compared with adult-only controls of the same density (1, 5, or 9 cells/μL; A1, A5, or A9, respectively). We found that coculture of primary adult tissue in an overwhelming embryonic environment resulted in a 1.9-fold increase in the number of adult neurospheres that formed compared with adult tissue plated alone (5.7±0.9 vs. 3.0±0.5 adult neurospheres/500 cells; A1E9 vs. A1, respectively, P<0.05) (Fig. 1B). A similar fold increase was seen using embryonic YFP and adult CD1 mice, revealing that strain differences do not account for the increased numbers of neurospheres observed. These data indicate that ANSCs respond to cues derived from an embryonic environment.
The increase seen in adult neurosphere formation could be mediated by cell–cell contact or be due to soluble factors released in the media. To test these possibilities we cultured primary adult tissue in CM from primary E14.5 embryonic cultures (embryonic CM). We observed a significant increase in the number of adult neurospheres compared with controls grown in standard neurosphere conditions (EGF and FGF only) (4.9±0.4 vs. 2.1±0.2 adult neurospheres/500 cells, embryonic CM vs. control media, respectively; P<0.01) (Fig. 1C). CM from primary adult cultures (adult CM) did not elicit an increase in adult neurosphere formation from primary adult dissections (data not shown), supporting the hypothesis that the factors acting on the ANSCs are derived from the embryonic environment. We compared the size of adult neurospheres that formed in A1E9 cocultures and A1-only controls and found no difference in the average size of neurospheres from each condition (118.4±4.2 μm vs. 109.6±7.8 μm adult neurosphere diameter; A1E9 vs. A1, respectively) (Fig. 1D). Taken together, these findings indicate that the embryonic factors are increasing the size of the ANSC pool but not leading to an expansion of the progenitor population.
Embryonic-derived factors enhance stem cell survival
The increased numbers of neurospheres observed in the presence of embryonic CM could be due to enhanced survival of the ANSCs. Consistent with this idea, when we assayed for viable cells at various times postplating, we found that 67% of the plated, adult-derived cells undergo cell death within 48 h (Fig. 2A). We hypothesized that if embryonic-derived factors were responsible for enhancing adult cell survival, then delayed addition of embryonic cells to adult cultures by 2 days would not result in increased adult neurosphere formation because cell death would already have occurred in vitro. Primary ANSCs were plated for 2 days prior to the addition of primary embryonic cells and the number of adult neurospheres was assayed 9 days after initial plating (7 days after exposure to the embryonic environment). As predicted, we observed no increase in the numbers of adult neurospheres in delayed coculture conditions (3.7±0.7 vs. 3.3±0.9 adult neurospheres/500 cells; delayed addition vs. controls, respectively; P=0.7) (Fig. 2B).
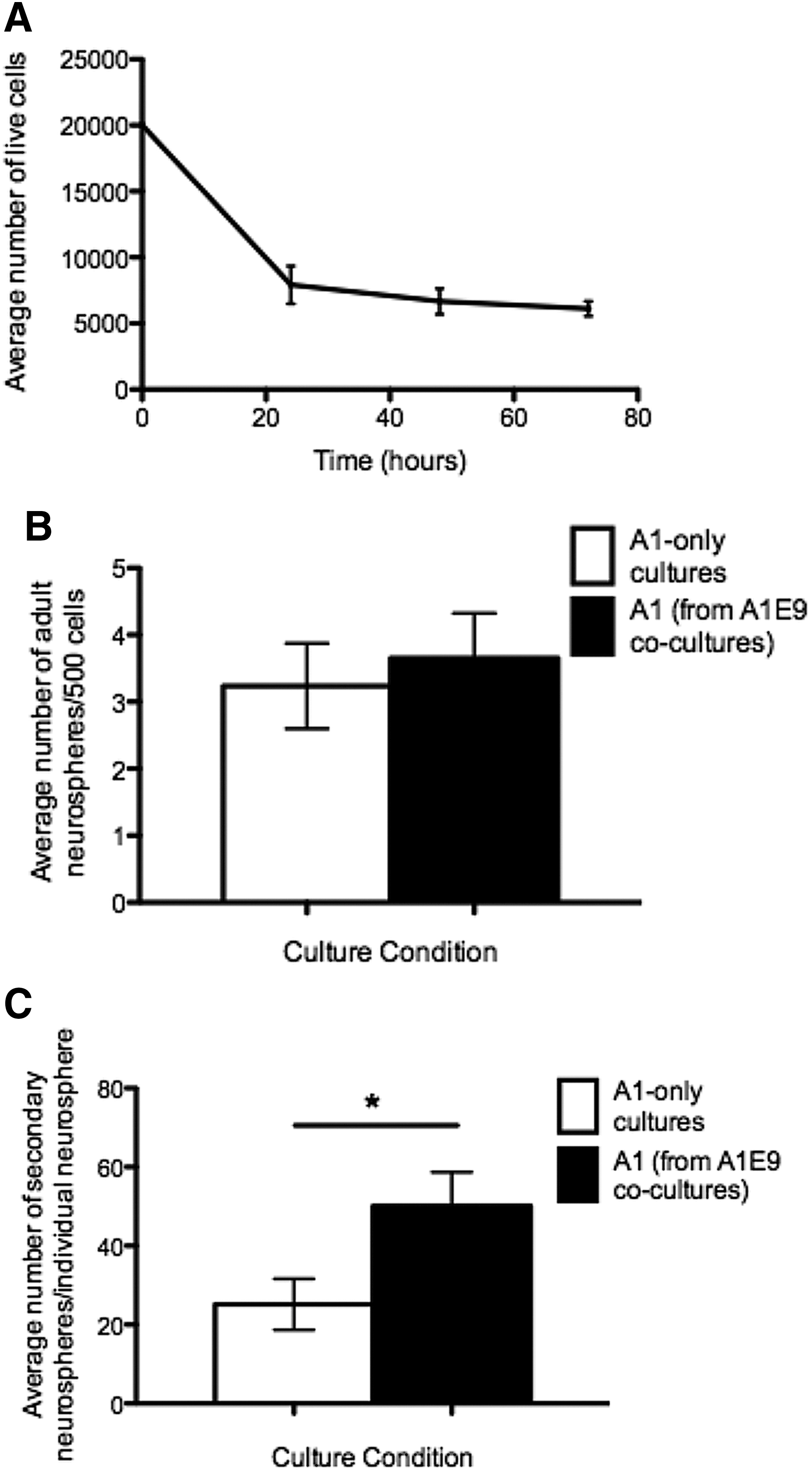
Embryonic factors enhance adult neural stem cell (ANSC) survival without changing the proliferation kinetics.
To test whether the embryonic-derived factor enhanced ANSC survival within developing neurospheres, we took individual adult neurospheres from adult-only cultures and A1E9 cocultures where the embryonic environment was added without a delay and performed single-neurosphere passaging. The number of secondary neurospheres was significantly higher from neurospheres that were exposed to an embryonic environment compared with adult-only controls (50.1±8.6 vs. 25.1±6.5 secondary neurospheres; A1E9 vs. A1, respectively; P<0.05) (Fig. 2C). Taken together, these findings support the hypothesis that the increase in adult neurosphere formation in the presence of an embryonic environment is due to enhanced survival of ANSCs.
Cell survival factors are released from embryonic neurons
To determine which cells were responsible for releasing the NSC survival factor(s), we first performed coculture experiments with primary cells from different developmental ages to determine when the embryonic environment could elicit the increase in adult neurosphere formation. Embryonic and adult cocultures were established from E12.5 (during neurogenesis), postnatal day 0–3 (PND0–3) (when astrocyte development peaks and oligodendrocyte formation begins), and PND14–16 (peak of oligodendrocyte formation) [21]. The data reveal a significant increase (>2-fold) in the numbers of adult-derived neurospheres in E12.5 and PND0–3 cocultures and no effect in PND14–16 cocultures (Fig. 3A). Based on these observations we hypothesized that the prosurvival factor(s) must be released from embryonic neural precursors, neurons, and/or endothelial cells, as these cells would be found in the cocultures that showed enhanced survival. We sought to determine whether CM from pure populations of neural precursor cells, neuronal cells, or endothelial cells promoted ANSC survival. First, we plated pure populations of embryonic neural precursor cells from passaged primary embryonic neurospheres and collected embryonic precursor CM. A1-only cultures grown in the presence of embryonic precursor CM did not have increased numbers of adult neurospheres compared with adult-only controls (2.1±0.3 vs. 2.5±0.4 adult neurospheres/500 cells; A1-only vs. embryonic precursor CM, respectively; P=0.18) (Fig. 3B). Endothelial cell CM was generated using the cell line derived from mouse brain endothelium [22] and, similarly, the endothelial cell CM did not elicit an increase in the numbers of adult-derived neurospheres. Instead, we observed a loss of neurosphere formation relative to controls (5.8±1.1 vs. 0.25±0 neurospheres/500 cells; A1-only vs. endothelial cell CM, respectively; P<0.01) (Fig. 3B). We propose that this decrease in neurosphere formation could be the result of the difference in the media requirements for endothelial cells versus adult periventricular cultures; thus, endothelial cells from primary embryonic dissections would not be likely to survive in our neurosphere media, making them an unlikely source for the released survival factor. Finally, neuron CM were derived from pure populations of neurons from embryonic cortices of Tα1:nlacZ mice [23]. Embryonic cortices were dissociated and fluorescence-activated cell sorting (FACS) was analyzed to isolate pure population of β-galactosidase-expressing cells (Tα1+ neurons) [16]. The Tα1+ cells were placed in culture for 5 days, and fixed and stained for MAP2 to confirm that >98% of the cells were neurons as previously reported [16]. When primary A1-only cultures were plated in neuron CM, we observed a 1.7-fold increase in the number of adult neurospheres compared with A1-only controls (6.75±0.5 vs. 11.17±0.7 adult neurospheres/500 cells; A1-only vs. neuron CM, respectively; P<0.01) (Fig. 3B). Hence, these findings support the hypothesis that embryonic neurons are the source of the released prosurvival factors.
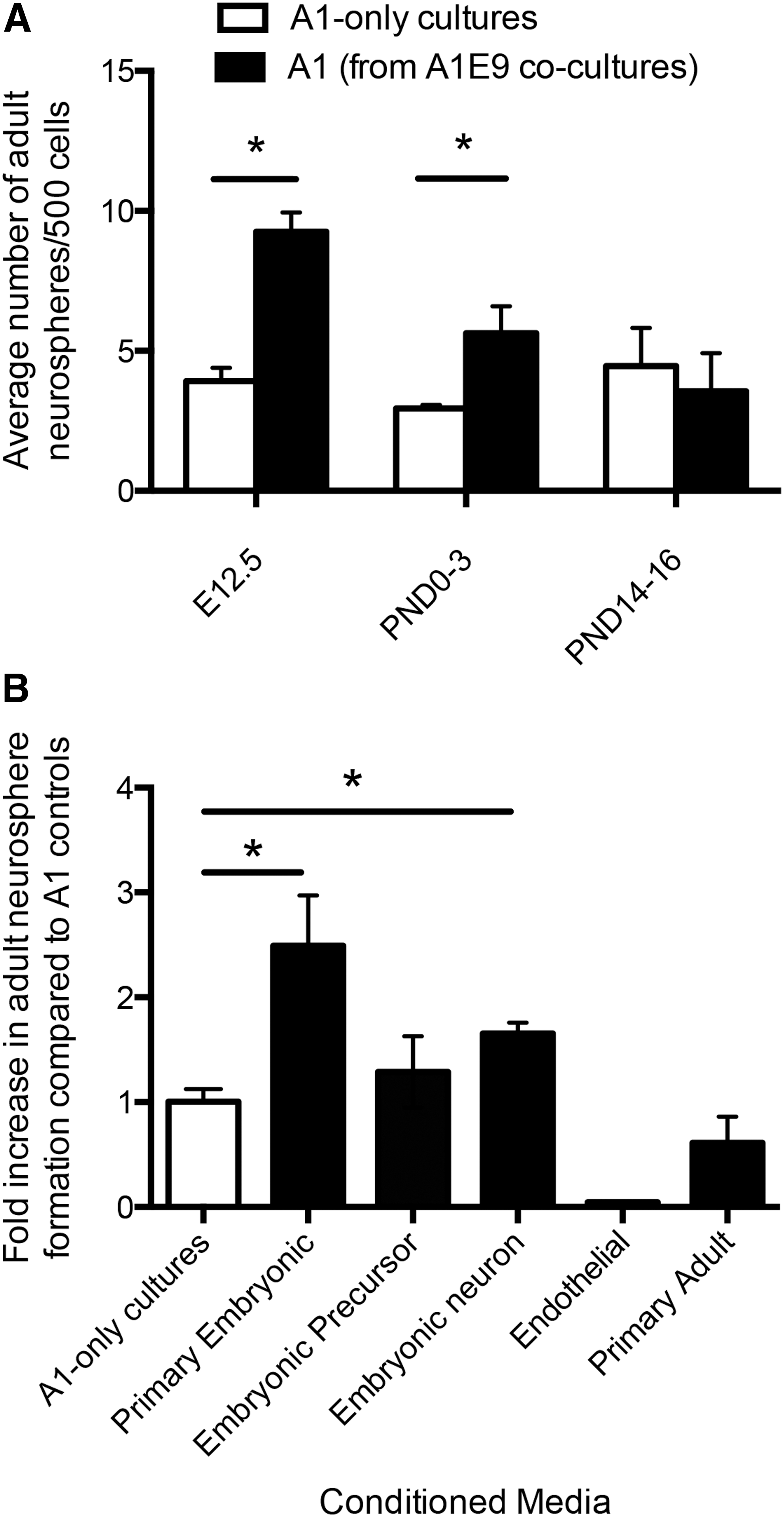
The prosurvival factor is released from embryonic neurons.
The receptor for SCF, ckit, is upregulated in adult neurospheres following exposure to an embryonic environment
SCF is known to be released from neurons and has been shown to promote ANSC survival by acting on its receptor, ckit, which is expressed on neural cells [10,24]. To test the hypothesis that SCF mediates the enhanced survival of ANSCs in the coculture condition, we used an ELISA to detect SCF in neuron CM, embryonic CM, and adult CM from primary cultures (Fig. 4A). The ELISA demonstrated that SCF was present in the neuron CM (116.134±7.114 pg/mL) and embryonic CM (51.354±9.447 pg/mL) and very low levels of SCF were seen in the adult CM (11.824±3.106 pg/mL). Hence, embryonic neurons present in primary dissections of embryonic brain are a main source of SCF.

Stem cell factor (SCF) is released from embryonic neurons and enhances ANSC survival in coculture.
We next examined the expression of the SCF receptor ckit in adult-derived neurospheres using semiquantitative reverse transcription polymerase chain reaction (RT-PCR). Interestingly, we found the complete absence of ckit expression in neurospheres derived from A1-only cultures; however, ckit was expressed in adult neurospheres derived from A1E9 cocultures (Fig. 4B). As previously reported [11], ckit is present in embryonic neurospheres (Fig. 4B). We reasoned that if SCF is mediating the prosurvival effects then cells from dissociated A1-only neurosphere cells (no ckit expression) would not respond to SCF with increased neurosphere formation. Conversely, we predict that A1E9-neurosphere-derived cells would respond to SCF with increased neurosphere formation. Accordingly, we took individual adult-derived neurospheres from each culture condition, and passaged and replated the neurosphere-derived cells with a range of SCF concentrations from 0 to 2 ng/mL. As predicted, SCF had no effect on the A1-only control neurospheres (which do not express ckit); however, the A1E9 adult neurospheres (which express ckit) revealed a significant 1.6-fold increase in neurosphere formation in the presence of 1.0 ng/mL SCF (85.92±31.42 vs. 47.0±14.74 adult neurospheres/500 cells; A1E9 cocultures vs. A1-only, respectively; P<0.05). To confirm that active signaling in the SCF/ckit pathway mediated the enhanced survival seen in coculture, we used a function-blocking antibody to ckit (ACK2). In the presence of ACK2, the increased number of neurospheres in A1E9 cocultures was not observed (Fig. 4C). Finally, we performed coculture experiments using adult cells from mice lacking the ckit receptor (KitW/KitWv double-knockout mice subsequently referred to as ckit-KO). Again, we observed no increase in the number of adult ckit-KO neurospheres in the presence of an overwhelming embryonic environment (3.3±0.5 vs. 3.2±0.3 adult neurospheres/500 cells, K1E9 vs. K1; P=0.59) (Fig. 4D). Thus, the upregulation of ckit on ANSCs is necessary for the enhanced survival in the presence of an embryonic environment. Together these findings reveal that SCF released from neurons mediates the prosurvival effects on ANSCs.
Enhanced NOS activity leads to upregulation of ckit expression
We next sought to determine which factor mediated the upregulation of ckit in adult neurospheres in A1E9 cocultures. NO has been shown to regulate smooth muscle contraction and NOS expression is closely connected to ckit expression in intestinal interstitial cells [25,26]. Moreover, the loss of nNOS expression in intestinal neurons results in decreased expression of ckit [27]. In the developing brain, NOS is strongly expressed in the cortical plate, peaking at E17 and declining steadily until PND2 after which time the expression is lost and the level remains constant into adulthood [28]. Based on these findings, we examined the role of NO in ckit upregulation in neural precursor cells.
We first performed coculture experiments using adult cells derived from nNOS−/− transgenic mice and wild-type embryonic cells. We predicted that if NO was important for ckit upregulation, then the adult-derived cells from nNOS−/− would not respond to SCF since ckit would not be upregulated and no increase in adult-derived neurospheres would be observed in the presence of SCF in vitro. We prepared A1-only and A1E9 cocultures from adult nNOS−/− mice and E14.5 YFP embryos and, as predicted, we observed no difference in the numbers of neurospheres generated in each condition (2.69±0.77 vs. 3.06±0.71 adult neurospheres/500 cells; A1-only vs. A1E9, respectively; P=0.75) (Fig. 5A). Further, PCR revealed no upregulation of ckit expression in adult neurospheres from A1E9 cocultures from adult nNOS−/− mice (Fig. 5A), supporting the hypothesis that nNOS is required for the upregulation of ckit.
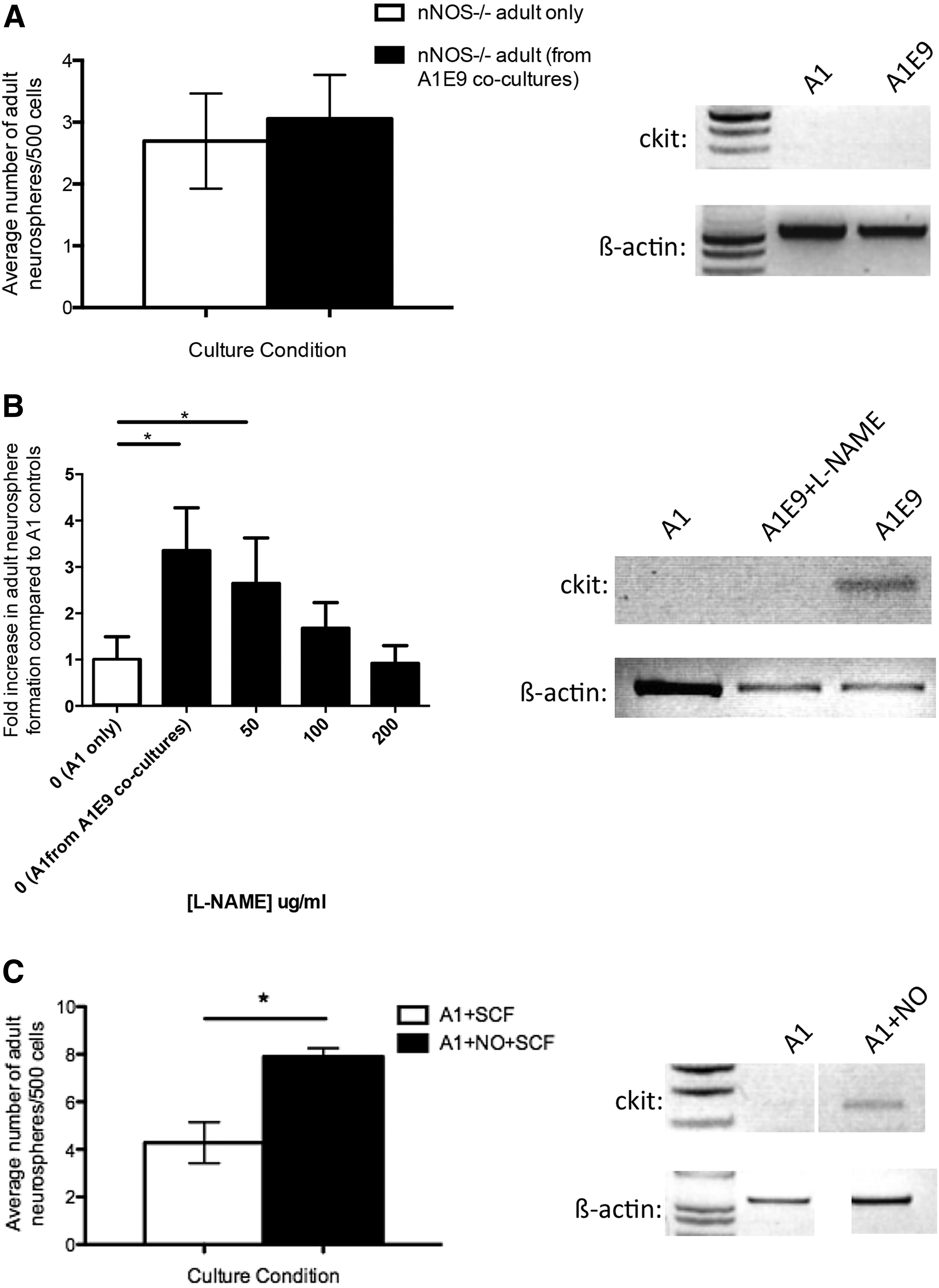
Nitric oxide (NO) results in the upregulation of ckit.
Two additional assays were performed to further confirm the role of nNOS in ckit upregulation. L-NAME, an inhibitor of NOS synthesis [29], was added to A1E9 cocultures for the first 48 h in vitro. In the presence of L-NAME, we observed no increase in the numbers of adult neurospheres in A1E9 cultures relative to A1-only cultures and no upregulation of ckit expression (Fig. 5B). Finally, A1-only cultures were exposed to the NO donor (Spermine NONOate) and SCF with the expectation that NO would upregulate ckit and increased neurosphere formation would be observed in the presence of SCF. Indeed, we observed a 1.8-fold increase in the number of neurospheres (4.3±0.9 vs. 7.9±0.4 neurospheres/500 cells; A1+SCF vs. A1+SCF+NONOate, respectively; P=0.055) and an increase in ckit expression (Fig. 5C). Together, these findings support the hypothesis that NO/nNOS is responsible for the upregulation of ckit and subsequent response to SCF that enhances cell survival.
SDF-1α is released by embryonic neurons and results in an increase in nNOS activity
The half-life of NO is very short [30 –34]; hence, the fact that ANSC survival was enhanced using CM (ie, in the absence of cells actively producing NO) led us to hypothesize that another factor in the media was responsible for NOS activation. The chemokine SDF-1 has been shown to upregulate NOS in endothelial cells [35]; it is released from neurons [36,37], and further, the receptor for SDF-1 (CXCR4) is expressed in the periventricular region of the adult brain [38]. SDF-1 has three isoforms (α, β, and γ) of which the α [39] and β [40] isoforms are most highly expressed during development. We performed semiquantitative RT-PCR on neurospheres and confirmed that CXCR4 is present on adult and embryonic neurospheres (derived from A1-only or E9-only cultures) (Fig. 6A). We performed an ELISA on neuron CM, primary embryonic CM, and primary adult CM for SDF-1α and found high levels of expression in neuron CM (15.51±6.96 pg/mL), lower levels in primary embryonic CM (1.52±1.08 pg/mL), and no expression in primary adult CM (Fig. 6B), consistent with the expression patterns of SDF-1α in vivo [39]. Based on the expression of the CXCR4, and the presence of SDF-1α in the CM, we added purified SDF-1α to A1-only cultures over a concentration range of 0 to 12.5 pg/mL, in the presence or absence of SCF (2 ng/mL). We predicted that if SDF-1α activated NOS, then we would observe an increase in ckit expression and subsequent increase in the numbers of neurospheres due to the presence of SCF in the cultures. Indeed, adult cells plated at 1 cell/μL (A1) in SDF-1α and SCF revealed a 2.1-fold increase in the number of neurospheres that formed (10.25±2.5 vs. 4.95±0.83 adult neurospheres/500 cells, 3.625 pg/mL SDF-1α+SCF vs. 3.625 pg/mL SDF-1α-only controls, respectively; P<0.05) (Fig. 6C). We found that the effect was specific to SDF-1α as the β isoform did not increase neurosphere formation (data not shown). Further, semiquantitative RT-PCR on adult neurospheres from A1-only cultures containing SDF-1α revealed that adult neurospheres upregulate nNOS and ckit expression (Fig. 6D). Finally, the addition of a CXCR4 inhibitor (AMD3100) to A1+ embryonic CM cultures completely abolished the increase seen in adult neurosphere formation even at low doses of the inhibitor (4.58±0.6 vs. 3.0±0.5 adult neurospheres/500 cells, A1+ embryonic CM+ 3 μg/mL AMD3100 vs. A1-only+ 3 μg/mL AMD3100; P=0.11; Fig. 6E), indicating that the SDF-1 signaling is necessary to initiate the prosurvival cascade in ANSCs.
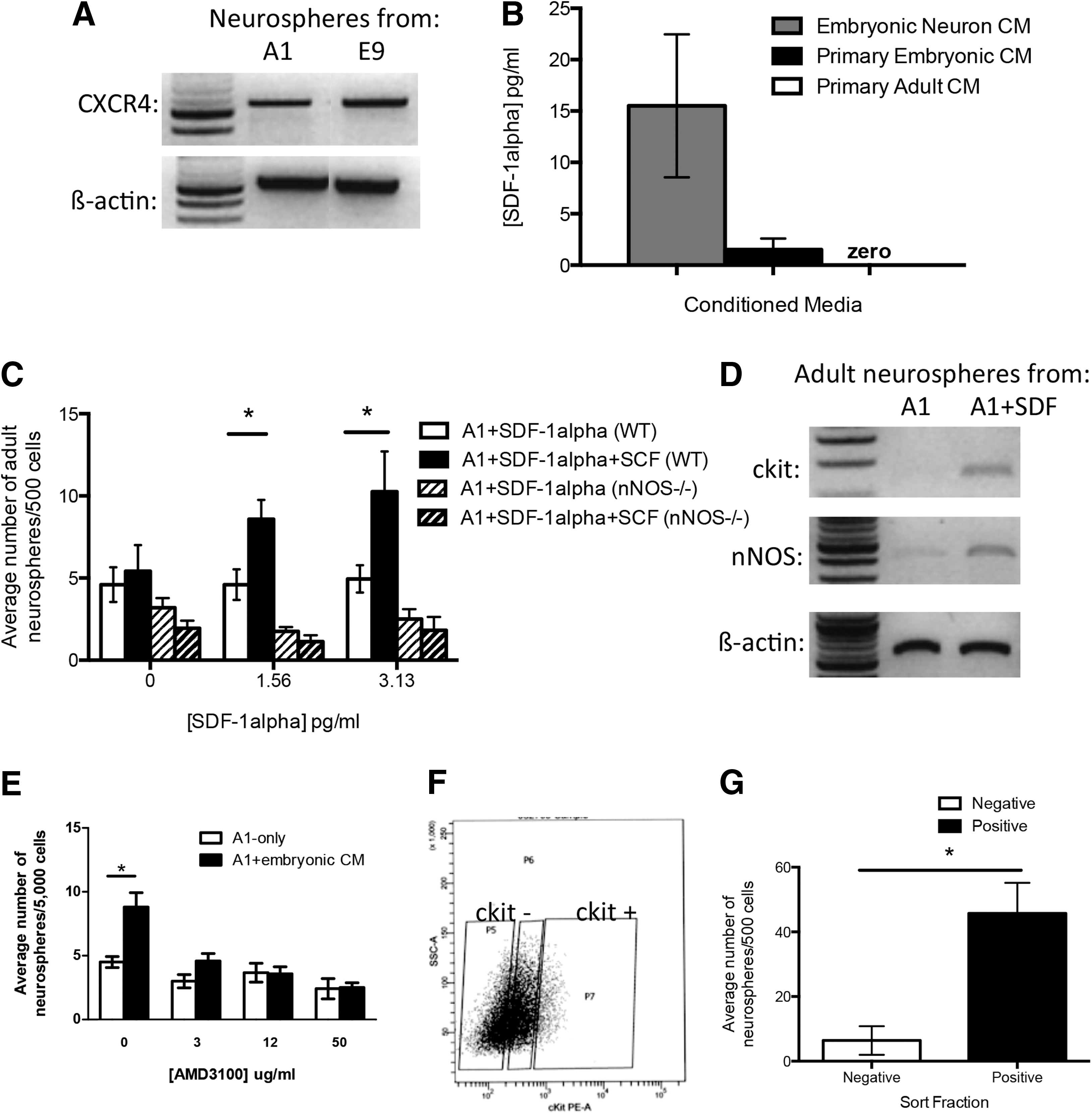
The role of stromal-cell-derived factor-1α (SDF-1α) in the upregulation of ckit.
To further confirm the role of NOS in upregulating ckit in response to SDF-1α, we used adult cultures derived from nNOS−/− mice. We predicted that no increase in neurosphere formation would be observed in A1-only cultures in the presence of SDF-1α and SCF because these animals lack the ability to upregulate nNOS in the presence of SDF-1α. As predicted, we observed no change in the numbers of neurospheres from nNOS−/− mice (Fig. 6C), suggesting that nNOS is required in the ANSCs in order to upregulate ckit and that NO is not provided by other cell types in culture. This is in agreement with our data that demonstrate that only cultures containing embryonic neurons (embryonic CM or neuron CM) were able to elicit an increase in adult neurosphere formation. Hence, SDF-1α acts to upregulate nNOS, leading to NO release, which results in upregulation of ckit expression thereby enabling the cells to respond to SCF and promoting NSC survival.
ckit expression is upregulated on ANSCs
Neurospheres represent a mixed population of stem and progenitor cells. If progenitors, and not stem cells, upregulated ckit, then this would infer that another factor was released by the progenitor cells that would act on the NSCs and promote their survival. To test which cells upregulate ckit, primary adult neurosphere cultures were grown at 1 cell/μL (A1) in the presence of 5 pg/mL SDF-1α, which leads to ckit upregulation in neurospheres (Fig. 6D). The resulting neurospheres were collected, dissociated into single cells, and labeled with a PE-conjugated anti-ckit antibody. The cells were sorted for ckit expression into negative (ckit−) and positive (ckit+) fractions and replated in A1-only conditions. If ckit was upregulated on ANSCs, then the ckit+ fraction would contain the neurosphere-forming cells. However, if ckit was upregulated on progenitors only, then neurosphere formation would be found in the ckit− fraction. We found a significant enrichment in neurosphere formation in the ckit+ fraction (45.7±9.5 vs. 6.4±4.4 adult neurospheres/500 cells, ckit+ vs. ckit−; P<0.05) (Fig. 6F, G). Neurosphere formation was highly enriched within the ckit+ fraction that comprised 4.75%±2.0% of cells and gave rise to 70%±12% of all neurosphere formation. Together, these data indicate that ckit is upregulated on ANSCs.
SDF-1α and SCF enhance ANSC survival in vivo
We asked whether the mechanism delineated in vitro could enhance ANSC survival in vivo. Toward this end, we intraventricularly infused adult mice for 5 days with primary embryonic CM, primary adult CM, or aCSF using osmotic minipumps. Mice were sacrificed and neurosphere cultures were established. Mice receiving aCSF or primary adult CM infusions showed no significant increase in neurosphere numbers compared with naive controls. In contrast, the number of neurospheres generated from mice that received primary embryonic CM was 1.7-fold greater than controls (9.0±1.3 vs. 5.5±0.4 neurospheres/500 cells; primary embryonic CM vs. naive controls, respectively; P<0.01) (Fig. 7A). These data confirm that primary embryonic CM can modify the behavior of ANSCs, both in vitro and in vivo. Next, we asked whether SDF-1α and SCF were sufficient to elicit enhanced survival in vivo. We infused SCF alone or SDF-1α+SCF into adult mice for 5 days (low dose: 100 pg/mL SDF-1α+40 ng/mL SCF or high dose: 200 pg/mL SDF-1α+80 ng/mL SCF). We observed a 1.8- and 1.6-fold increase in the number of neurospheres that formed at low and high doses, respectively [2.8±0.5 neurospheres/500 cells (naive) vs. 4.9±1.2 (low dose, P<0.05) and 4.5±0.7 (high dose, P<0.05)] (Fig. 7B). Hence, SDF-1α and SCF are sufficient to promote ANSC survival in vivo.
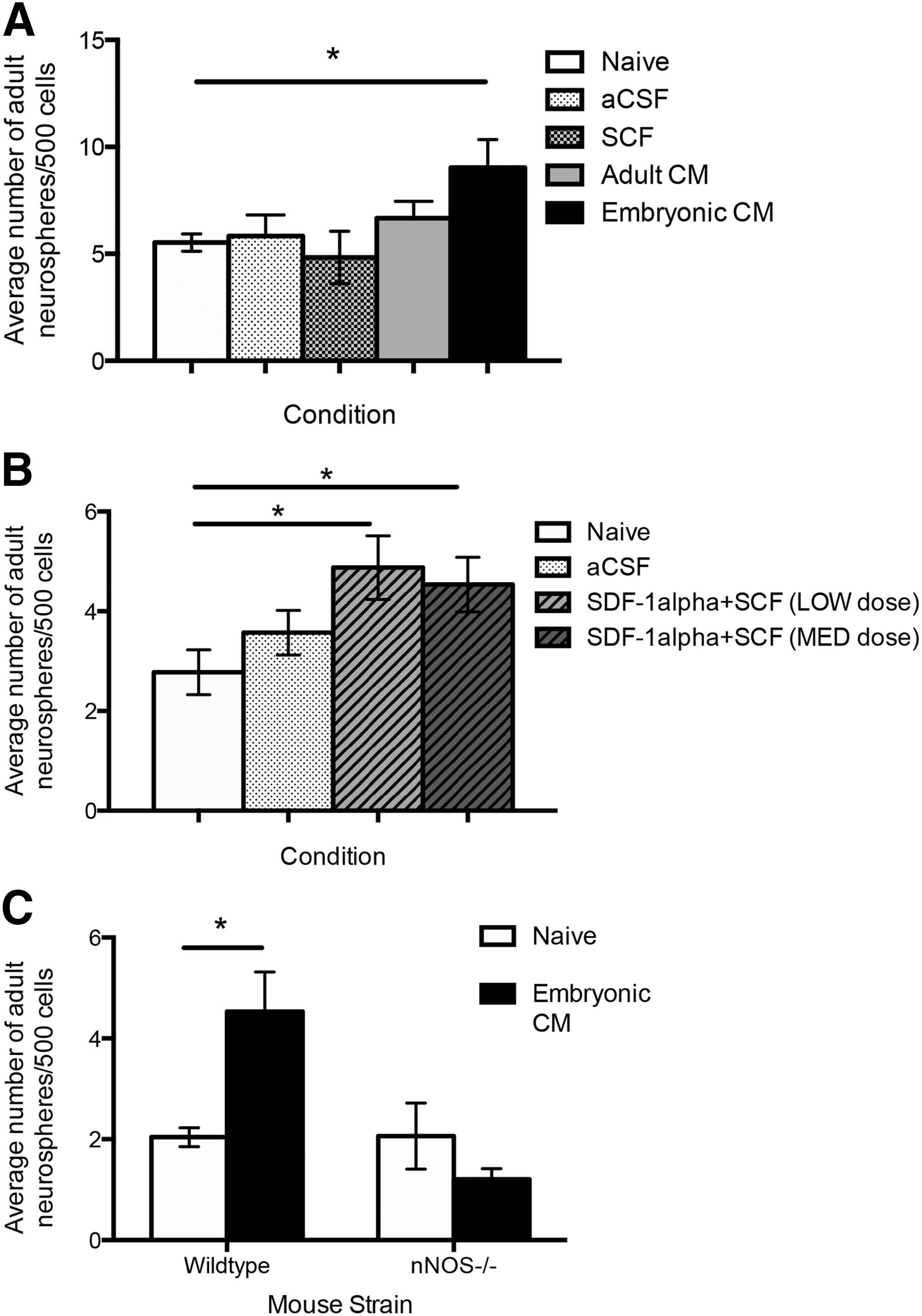
In vivo infusion of SDF-1α and SCF enhanced NSC survival.
Finally, to confirm the role of NO in the prosurvival pathway, nNOS−/− mice were infused with primary embryonic CM for 7 days; the brains were removed and cultured in the neurosphere assay. Naive nNOS−/− control mice generated similar numbers of neurospheres as naive strain-matched (C57/BL6) control mice (2.0±0.2 vs. 2.1±0.7 neurospheres/500 cells, C57/BL6 vs. nNOS−/−, respectively; P=0.97) (Fig. 7C) and primary-embryonic-CM-infused C57/BL6 mice revealed a 2.2-fold increase in neurosphere formation (4.5±0.8 neurospheres/500 cells; P<0.05) as predicted. Further, the numbers of neurospheres from primary-embryonic-CM-infused nNOS−/− mice were not significantly different from controls (1.2±0.2 vs. 2.1±0.7, embryonic-CM-infused nNOS−/− vs. nNOS−/− controls, respectively; P=0.17) (Fig. 7C). Taken together, these findings are consistent with our in vitro model whereby SDF-1α, released by embryonic neurons, can act on ANSCs to enhance nNOS expression and NO release to upregulate ckit expression on ANSCs enabling them to respond to SCF and promoting cell survival (Fig. 8).
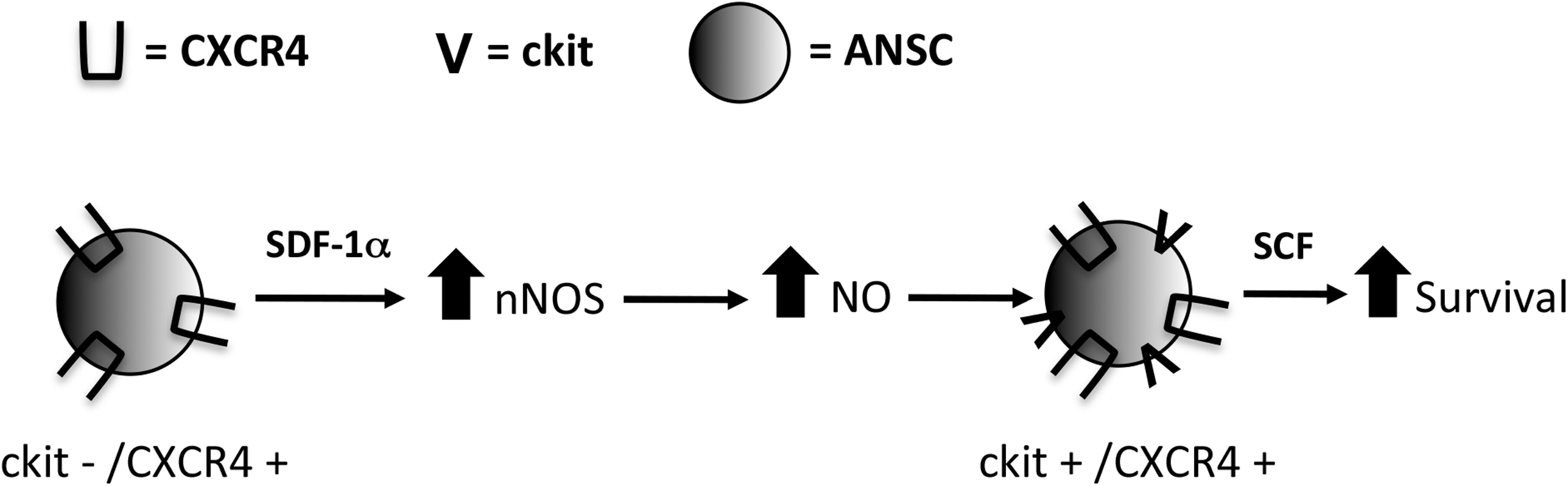
ANSC survival mechanism. Embryonic neurons release SDF-1α and SCF. SDF-1α leads to upregulation of nNOS and enhanced NO production, which results in ckit upregulation on ANSCs. ckit+ ANSCs can then respond to the SCF released from embryonic neurons, which leads to enhanced NSC survival.
Discussion
Herein we describe a novel mechanism of ANSC survival and demonstrate that ANSCs remain competent to respond to cues present in the developing brain. We have shown that embryonic neurons release SDF-1α, which results in increased nNOS activity and the release of NO, leading to the upregulation of ckit on ANSCs. The ckit-expressing ANSCs are then competent to respond to SCF, which is also released from embryonic neurons. We have demonstrated that SDF-1α and SCF can promote NSC survival both in vitro and in vivo. Together, our data reveal the important interplay between the progeny of NSCs and the regulation of NSC behavior.
Cues derived from the differentiated progeny of neural precursor cells have been shown to feed back on the uncommitted neural precursor cells in the developing and adult brain. In the adult brain it is well established that ablating NSC progeny leads to NSC proliferation [12,41] with symmetric divisions [18]. In the developing brain it has been shown that newborn cortical neurons release CT-1, which activates the gp130-JAK-STAT pathway in embryonic cortical precursors inducing astrogenesis [16]. This regulation of embryonic precursors by CT-1 has important implications for the timing and organization of the developing brain. In our study, we have similarly shown that newborn neurons in the developing brain release factors that modify the behavior of NSCs. Neurons are continuously generated in the adult brain and it would be interesting to know whether these newborn neurons also contribute to NSC survival under baseline conditions in the adult brain.
Our data suggest that the survival factors persist in the early postnatal period as coculture with cells from the PND0–3 brain was able to support the enhanced survival of ANSCs. Notably, the prosurvival effects were lost in the PND16–18 brain despite reports that both SCF and ckit are expressed in the PND18 brain [42,43]. These findings could suggest that an inhibitory factor may be present that interferes with the prosurvival effects. One possibility is that oligodendrocytes, which expand during this late postnatal period, are the source of the inhibition. Indeed, myelin has been shown to be inhibitory to NSC proliferation [44 –47]. Hence, a more complex system of pathway activation or inhibition appears to regulate the ANSC niche, which could account for the loss of the effect seen postnatally.
The SCF receptor ckit has been shown to be present in the SE of adult rodents [11,48]; however, its colocalization with ANSCs has not yet been demonstrated. In the adult brain, ckit is expressed from Nestin+ [11] and BrdU+ cells [48], consistent with expression on rapidly dividing NSC progeny. Consistent with this observation, our findings suggest that ANSCs do not express the receptor under baseline conditions; however, ckit can be induced following SDF-1α signaling. Our FACS analysis revealed that the ckit+ fraction, comprising <5% of all sorted cells, contained the majority of neurosphere-forming cells. Most importantly, when naive adult animals are infused with SCF only, there is no increase in neurosphere formation (Fig. 7A), indicating that ANSCs are not responsive to SCF under baseline conditions, likely due to a lack of receptor expression in the stem cell pool.
NO has been shown to have pleiotropic effects in the adult brain. In models of ischemia, NO overproduction results in cell death [49]. NO production has also been shown to regulate neurogenesis and to both promote and inhibit neural precursor cell proliferation [50,51]. Further, recent work has demonstrated that reactive oxygen species have a dramatic impact on the self-renewal and survival of the ANSCs and their progeny [52]. Hence, while the role of NO is diverse and context dependent, the recent work by Le Belle and colleagues is consistent with our findings that NO plays a role in regulating NSC survival.
Both SCF [11] and SDF-1 [36,53,54] have been shown to be upregulated after injury. SDF-1 induces chemotaxis of a variety of cells, including ANSCs, to home to sites of injury [53]. SCF has specifically been shown to be upregulated in the ANSC niche after injury [11]. Interestingly, human NSCs have been shown to express CXCR4 [55 –57] and can respond to injury cues following transplantation with migration to sites of injury following spinal cord injury [55] and hemorrhagic stroke [57]. The role of SDF-1 and CXCR4 in human NPCs suggests a potential role for this mechanism in regenerative strategies. Further, brain injury leads to an expansion of the size of the stem cell pool [18,58], which, based on the findings herein, could be accounted for by the upregulation of SDF-1 and SCF. The ability to regulate NSC survival has implications for the development of novel therapeutics to promote neural regeneration of the injured brain.
Footnotes
Acknowledgments
The authors would like to thank Dr. F. Miller for the generous gift of the Tα1:nlacZ mice and Dr. Dave Piccin for assistance with confocal imaging. N.S. is supported by the CIHR TPRM award. The work was supported by operating grants to C.M.M. from CIHR and Heart and Stroke Foundation of Canada.
Author Disclosure Statement
No competing financial conflicts exist.