
Abstract
Cardiovascular disease remains the major cause of mortality, and cardiac cell therapy has recently emerged as a paradigm for heart repair. The epicardium is a layer of mesothelial cells covering the heart that during development contributes to different cardiovascular lineages, including cardiomyocytes, but which becomes quiescent after birth. We previously revealed that the peptide thymosin beta 4 (Tβ4) can reactivate adult epicardium-derived cells (EPDCs) after myocardial infarction (MI), to proliferate, and differentiate into cardiovascular derivatives. The aim of this study was to provide a lineage characterization of the adult EPDCs relative to the embryonic epicardial lineage and to determine prospective cell fate biases within the activated adult population during cardiovascular repair. Wt1GFPCre/+ mice were primed with Tβ4 and MI induced by ligation of the left anterior descending coronary artery. Adult WT1+ GFP+ EPDCs were fluorescence-activated cell sorted (FACS) at 2, 4, and 7 days after MI. Embryonic WT1+ GFP+ EPDCs were isolated from embryonic hearts (E12.5) by FACS, and sorted cells were characterized by real-time quantitative reverse transcriptase–polymerase chain reaction (RT-qPCR) and immunostaining. Adult WT1+ GFP+ EPDCs were highly heterogeneous, expressing cardiac progenitor and mesenchymal stem markers. Based on the expression of stem cell antigen-1 (Sca-1), CD44, and CD90, we identified different subpopulations of EPDCs of varying cardiovascular potential, according to marker gene profiles, with a molecular phenotype distinct from the source embryonic epicardial cells at E12.5. Thus, adult WT1+ GFP+ cells are a heterogeneous population that when activated can restore an embryonic gene programme, but do not revert entirely to adopt an embryonic phenotype. Potential biases in cardiovascular cell fate suggest that discrete subpopulations of EPDCs might be clinically relevant for regenerative therapy.
Introduction
M
Regenerative medicine has emerged as an alternative strategy to facilitate cardiac repair, which includes stem cell therapy, reprogramming and tissue engineering. With regard to transplantation of stem cells, there has been rapid progression to the clinical trials reported [5 –8]; however, patient benefit has been modest at best. Moreover, a general consensus on the most suitable stem cell source remains to be elucidated, and the mechanisms of action remains to be fully determined. Several studies have recently demonstrated that functional improvements obtained after engraftment of stem cells into the heart can be largely attributable to the secretion of paracrine-acting factors stimulating cardio-protection, angiogenesis, and modulation of inflammation, rather than via direct trans-differentiation into new cardiovascular cells [9 –11]. In support of these studies, recent work has confirmed that exogenous transplanted stem cells can mediate heart repair by acting on endogenous cardiac progenitor cells (CPCs) residing in the heart, by activating them to differentiate into new cardiovascular cells, through the modulation of the local microenvironment and the paracrine release of specific trophic factors [12 –14]. Thus, regenerative medicine is now exploring strategies based on the stimulation and activation of the local CPCs, in parallel with the transplantation of exogenous stem cells, in order to reactivate inherent repair mechanisms of the adult mammalian heart.
Several types of CPCs have been broadly described in the literature and mainly characterized according to the expression of specific stem cell markers, such as stem cell antigen-1 (Sca-1) or c-kit, or based on their in vitro culture properties [15 –17] (extensively reviewed in Bollini et al. [18]). These different CPCs have been described as independent populations, although it is not clear whether they are derived from different cell sources during development or whether they represent cells derived from a common precursor subsequently isolated at different stages of differentiation and maturation. Among all the different CPC sources defined so far, there are two distinct populations, the origin of which can be specifically defined and traced back to embryonic development: notably Isl1+ CPCs [19] and epicardium-derived cells (EPDCs) [20,21].
While the Isl1+ population appears to represent remnants of cardioblasts arising during development and are rarely observed at postnatal stages, the EPDCs are derived from a layer of mesothelial cells covering the surface of the heart, which, during development, contributes to the majority of the major cardiovascular lineages. The epicardium develops initially from an outgrowth of cells called the pro-epicardial organ (PEO), a primordial cluster of epithelial cells located at the inflow region of the forming heart. PEO cells migrate to envelop the muscular heart tube and form the mesothelial epicardium. EPDCs then undergo epithelial-to-mesenchymal (EMT) transformation and migrate into the underlying myocardium, where they differentiate into adventitial fibroblasts, give rise to vascular smooth muscle cells, and contribute to the endothelium of the coronary vasculature [22,23]. EPDCs have also been proposed to contribute to the cardiomyocyte lineage in the developing heart [24,25], albeit this remains a controversial finding [26]. EPDCs, therefore, appear to represent a font of progenitors with potential for cardiac regeneration and heart repair; however, their clinical translation is hampered by the fact that the adult epicardium becomes quiescent soon after birth. We previously revealed that stimulation by the same factors which promote the activation and plasticity of the embryonic EPDCs can restore and sustain re-activation of their postnatal counterparts, as an exemplary paradigm of reinstating embryonic potential in putative therapeutic progenitor cell populations in the adult heart. The peptide thymosin beta 4 (Tβ4) has been identified as a regulator of EPDC activation and subsequent differentiation during coronary vasculature formation throughout embryogenesis, as well as a potent inducer of adult EPDC proliferation and differentiation into smooth muscle, endothelium, and fibroblast lineages, after ex vivo outgrowth from adult heart explants [27]. Translating this paracrine approach into murine preclinical models in vivo, we demonstrated that Tβ4 stimulation supported neovascularization in both the intact and ischemic heart and instructed adult EPDCs to recapitulate their developmental program with reactivation of specific embryonic epicardial genes, such as Wt1, followed by limited differentiation into new functional mature cardiomyocytes, after myocardial infarction (MI) [28,29]. Therefore, the adult epicardium can be considered a source of dormant cardiac progenitor cells, which, upon appropriate paracrine stimulation, can become re-activated, such that adult EPDCs have the potential to contribute to cardiac homeostasis and repair. While progress around understanding how to restore the cardiovascular potential of EPDCs has been recently realized, much remains to be determined regarding a detailed characterization of the adult lineage and most notably, in comparison to the embryonic source.
Here, we present a detailed fluorescence-activated cell sorting (FACS)-based molecular phenotyping of murine adult EPDCs as activated after Tβ4-treatment and MI and draw direct comparisons with the embryonic epicardial lineage. We reveal a unique cell surface marker signature and heterogeneity in cellular composition within the adult epicardium, which is reflected in a range of cardiovascular cell fate potential as determined by surrogate marker gene expression.
Materials and Methods
Mouse lines and MI protocol
Wt1GFPCre/+ mice have been previously described [29] and were generated by crossing transgenic Wt1GFPCre/+ male mice with wild-type C57BL/6 females purchased from Charles River. Adult mice were primed with daily intra-peritoneal injections of Tβ4 [12 mg/kg solution in phosphate-buffered saline (PBS) 1×; RegeneRX] for 7 days [29]. For the MI model, Wt1GFPCre/+ mice (n=145) weighing between 25 and 30 g and 8 weeks old were used, as previously reported. Briefly, mice were anesthetized with isofluorane under assisted external ventilation through the insertion of an endotracheal tube and underwent thoracotomy in order to perform a permanent ligation of the left anterior descending artery. Buprenorphine (buprenorphine hydrochloride; Vetergesic) was delivered as a 0.015 mg/mL solution via intra-peritoneal injections at 20 min before the procedure to provide analgesia. After recovery, animals received follow-up intra-peritoneal injections of Tβ4 on alternate days. Two, 4, and 7 days after MI, hearts were assessed using a combination of FACS, immunofluorescence, and real-time reverse transcriptase–polymerase chain reaction (RT-PCR) analysis.
All animal experiments were carried out according to the UK Home Office project licence PPL 30/2987 compliant with the UK Animals (Scientific Procedures) Act 1986 and approved by the University College London Biological Services Ethical Review Process.
WT1+ GFP+ EPDCs isolation
Wt1GFPCre/+ embryos were harvested at embryonic day E12.5 postcoitum and hearts were immediately processed by enzymatic digestion, as previously reported [19]. Briefly, hearts were washed in ice-cold Hank's balanced salt solution (HBSS) buffer (Gibco) and incubated in 0.1% collagenase II (Worthington Biochemical) and 0.25% trypsin-EDTA (Invitrogen) solution for 30 min at 37°C until a single-cell suspension was obtained. WT1+ GFP+ embryonic EPDCs were subsequently isolated via FACS sorting on GFP expression and characterized by FACS. Adult Wt1GFPCre/+ hearts were collected at 2, 4, and 7 days after MI was induced and processed to isolate adult epicardial cells according to [30]. Dissociated cardiac cells were isolated via multiple enzymatic incubations at 37°C in a 1% collagenase IV (Sigma) and 2.5% trypsin (Invitrogen) solution in HBSS. The enzymatic digestion was carried out in order to detach only the activated epicardial and sub-epicardial space, leaving the underlying myocardium intact. The obtained cell solution was subsequently depleted from any hematopoietic contamination using the Lineage Cell Depletion kit (Miltenyi Biotec), following the manufacturer's instructions and further characterized by means of flow cytometry.
Flow cytometry characterization and FACS sorting of embryonic and adult EPDCs
The WT1+ GFP+ embryonic (E12.5) EPDCs and the adult WT1+ GFP+ EPDCs (2, 4, and 7 days after MI) were characterized by FACS using a panel of epicardial, mesenchymal, and cardiac stem cell markers. The EPDCs immunophenotype was assessed using the following antibodies: PerCPCy5.5-conjugated anti-mouse Sca-1 (clone D7; eBioscience); Alexa Fluor 647-conjugated anti-mouse c-kit antibody (clone 2B8; Biolegend); PE-conjugated anti-mouse CD29 (Integrin β1, clone HMβ1-1; Biolegend); APC-Cy7- and Alexa Fluor 647-conjugated anti-mouse CD90.2 (clone 30-H12; Biolegend), APC-conjugated anti-mouse PDGFRβ (clone AP85; eBioscience), PE-Cy7-conjugated anti-mouse CD44 (clone IM7; eBioscience), Alexa Fluor 647-conjugated anti-mouse CD45 (clone 30-F11; Biolegend), APC-conjugated anti-mouse CD184 (CXCR4, clone 2B11; eBioscience), and anti-mouse CD105 primary antibody (R&D System) with Alexa Fluor 647-conjugated anti-rat secondary antibody (Molecular Probes, Life Tech). Compensation was set manually on the analyzer using CompBeads Plus compensation beads (BD Biosciences). Cells were acquired using a Beckam Coulter CyAn analyzer that was equipped with 488 nm blue and 635 nm red diode lasers and running Summit V3.4 software. Data were analyzed using the FlowJo software. Embryonic WT1+ GFP+ EPDCs and subpopulations of adult WT1+ GFP+ EPDCs were isolated by FACS sorting using a Beckam Coulter MoFlo XDP sorter, equipped with UV 355 nm, blue 488 nm, and red 647 nm lasers and running Summit V5.2 software.
Immunofluorescence staining of EPDCs
Hearts were collected at 4 days after MI, fixed in 4% PFA solution for 2 h on ice, and snap frozen in methylbutane and liquid nitrogen for cryosectioning. Eight-micrometer-thick sections were processed for immunofluorescence staining using the following primary antibodies: anti-mouse GFP, WT1, CD29, CD45, Sca-1, c-kit, CD90, CD44 (all Abcam), and PDGFRβ (R&D System). Anti-chicken Alexa Fluor 594-conjugated and anti-rat and anti-rabbit Alexa Fluor 488-conjugated secondary antibodies were used (Molecular Probes, Life Tech). Images were acquired using a Zeiss Apotome miscoscope and a Leica structural illumination DM 6000B microscope with AxioVision and Leica MMAF acquisition software.
Gene expression profile of adult EPDCs
Total RNA was isolated from WT1+ GFP+ embryonic (E12.5) EPDCs, from GFP− epicardial cells, and from WT1+ GFP+ EPDCs FACS sorted at 4 days after MI using the RNeasy Micro kit (Qiagen). cDNA was obtained using the Whole Transcriptome kit (Qiagen), following the manufacturer's instructions and used for real-time quantitative RT-PCR using SYBR Green on an ABI 7900 for the following genes: Wt1, Tbx18, Raldh2, Pdgfrβ, Isl1, Gata4, Flk1, Sma, SM22α, Pecam, and Fapα. Hprt1 was used as an endogenous control. Fold change was determined by applying the 2−ΔΔCT method. Primer sequences are available on request.
Statistical analysis
All values are expressed as mean±standard error. Statistical difference between the considered groups was evaluated by one-way ANOVA multiple-comparison test and Student's t-test, using Graph Pad Prism 6.0 software. A P-value<0.05 was considered significant.
Results
Isolation and immunophenotypic characterization of embryonic E12.5 and adult WT1+ GFP+ EPDCs
To restore the embryonic program in the adult EPDCs via reactivation of the expression of Wt1, we primed transgenic Wt1GFPCre/+ mice with a daily injection of Tβ4, followed by induction of MI as previously described [29]. WT1+ GFP+ EPDCs were subsequently analyzed by FACS, based on the expression of GFP, avoiding contamination from the underlying myocardium. The adult activated WT1+ GFP+ EPDCs were isolated after 2, 4, and 7 days post-MI and Tβ4 priming. FACS analyses revealed that the GFP+ fraction within the total adult-activated epicardial population was 9.31%±2.36% (n=12 hearts), 15.15%±1.51% (n=12 hearts), and 9.23%±2.50% (n=12 hearts), respectively (Fig. 1A, B). Control treatment with PBS (vehicle) before MI revealed that injury alone stimulated 11.0%±2.7% (n=18 hearts) at 4 days post-MI (Supplementary Fig. 1A; Supplementary Data are available online at
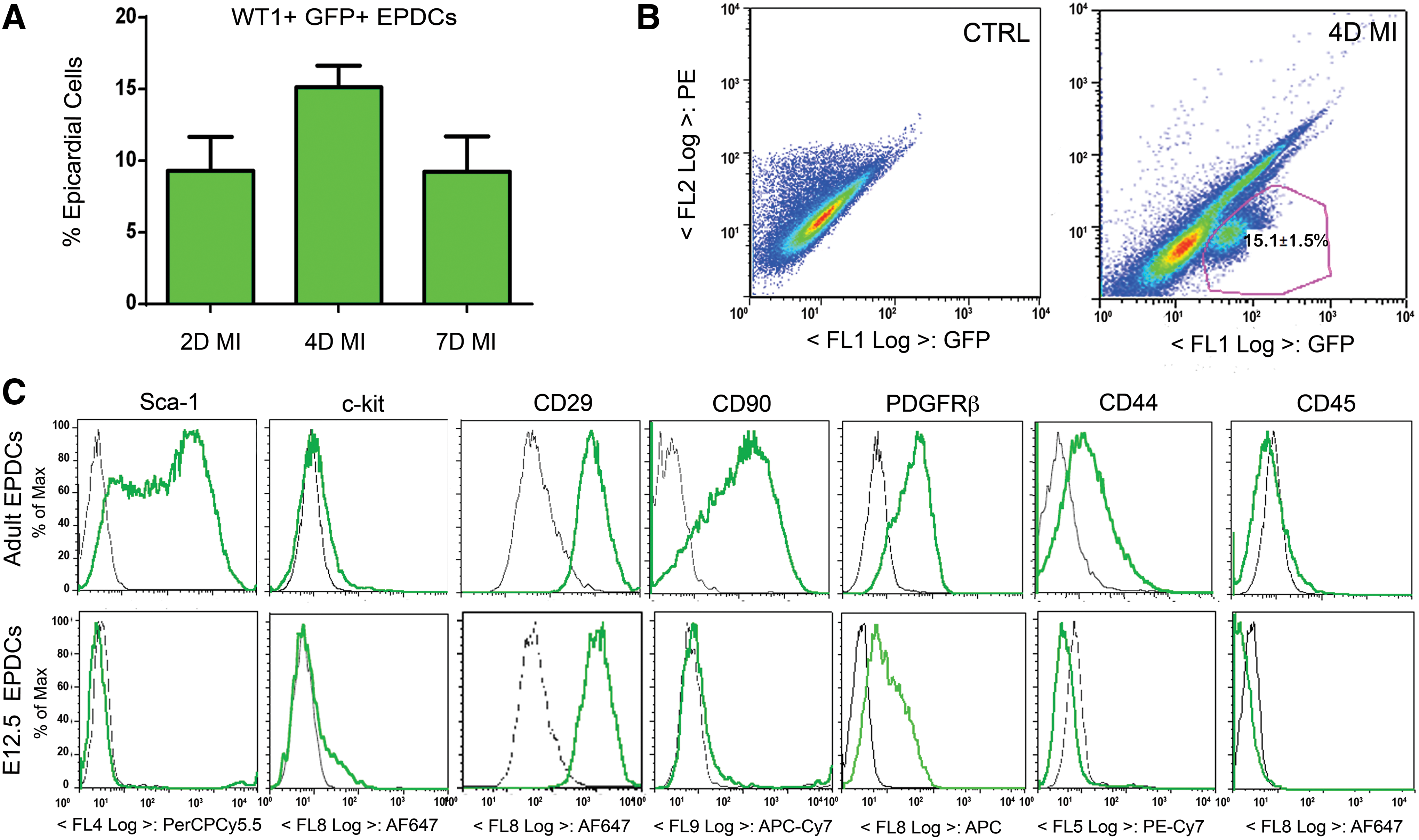
Isolation of adult-reactivated WT1+ GFP+ epicardium-derived cells (EPDCs) after priming with thymosin beta 4 (Tβ4) at day 2, 4, and 7 after myocardial infarction (MI).
The phenotype of the adult-reactivated WT1+ GFP+ EPDCs was then evaluated by FACS using a panel of cardiac and mesenchymal stem cell markers, as well as epicardial cell antigens (Table 1 and in Fig. 1C). A proportion of the adult WT1+ GFP+ EPDCs cells were found to be positive for the expression of the cardiac stem cell marker Sca-1 [31] ranging from 64.0%±2.1% at day 2 post-MI to 54.0%±2.8% at day 4 and 58.0%±4.9% at day 7 post-MI, with no significant differences across the time points analyzed; c-kit antigen [15] was hardly detected in these cells, being expressed in no more than 1.9%±0.2% of the GFP+ EPDCs at 4 days post-MI. A significant proportion of the adult WT1+ GFP+ EPDCs (90%) were of mesenchymal origin, as confirmed by the expression of CD29, with negligible expression of the hematopoietic marker CD45 [32,33]. While the mesenchymal marker CD90 [32] increased from day 2 to 7 post-MI (47.3%±4.6%, 65.5%±4.7%, and 78.1%±3.4%, respectively), the expression of CD44 [33 –35] remained constant over the 7 days after injury at about 20% of the WT1+ GFP+ cells. Chemokine (C-X-C motif) receptor 4 (CXCR4), the receptor for the stromal-derived factor-1 (SDF-1) playing a critical role in stem cell recruitment after MI [36], was expressed at relatively low levels in the adult-reactivated EPDCs, being present in 4.9%±2.2% of the WT1+ GFP+ cells at 2 days after MI and increased to 12.3%±4.2% 7 days after injury. CD105, a marker of mesenchymal stem cells [32], endogenous cardiac progenitors, such as the human cardiospheres [37] and the recently described proepicardial-derived murine cardiac-resident MSC-like stem cells [38], was expressed in 13.2%±3.8% of the adult WT1+ GFP+ EPDCs at 2 days after MI and increased to 22.7%±1.4% a week from injury. The expression of the epicardial marker PDGFRβ [39] increased from 23.0%±12.8% of the WT1+ GFP+ cells at 2 days post-MI to 51.0%±7.5% by day 7. The variable distribution of these markers among the adult-reactivated EPDCs, therefore, revealed a highly heterogeneous phenotype. We identified a consensus signature for the adult-reactivated WT1+ GFP+ EPDCs according to the combination of markers most highly expressed at 4 days after injury, which was Sca-1+/CD90+/CD44+/PDGFRβ+. The presence of EPDCs according to these markers was confirmed in situ by immunostaining in heart sections (n=15 hearts, Fig. 2A–I). Control (PBS-treated; n=18) mice also revealed an equivalent Sca-1+/CD90+/CD44+/PDGFRβ+ cell surface marker profile at day 4 post-MI (Supplementary Fig. 1B), although there was a reduced incidence of both Sca-1 (49.3%±3.1% vs. 54.0%±2.8%) and CD44 (8.7%±2.0% vs. 17.5%±4.8%) between PBS and Tβ4-treated animals (compare plots in Fig. 1C with Supplementary Fig. 1B).
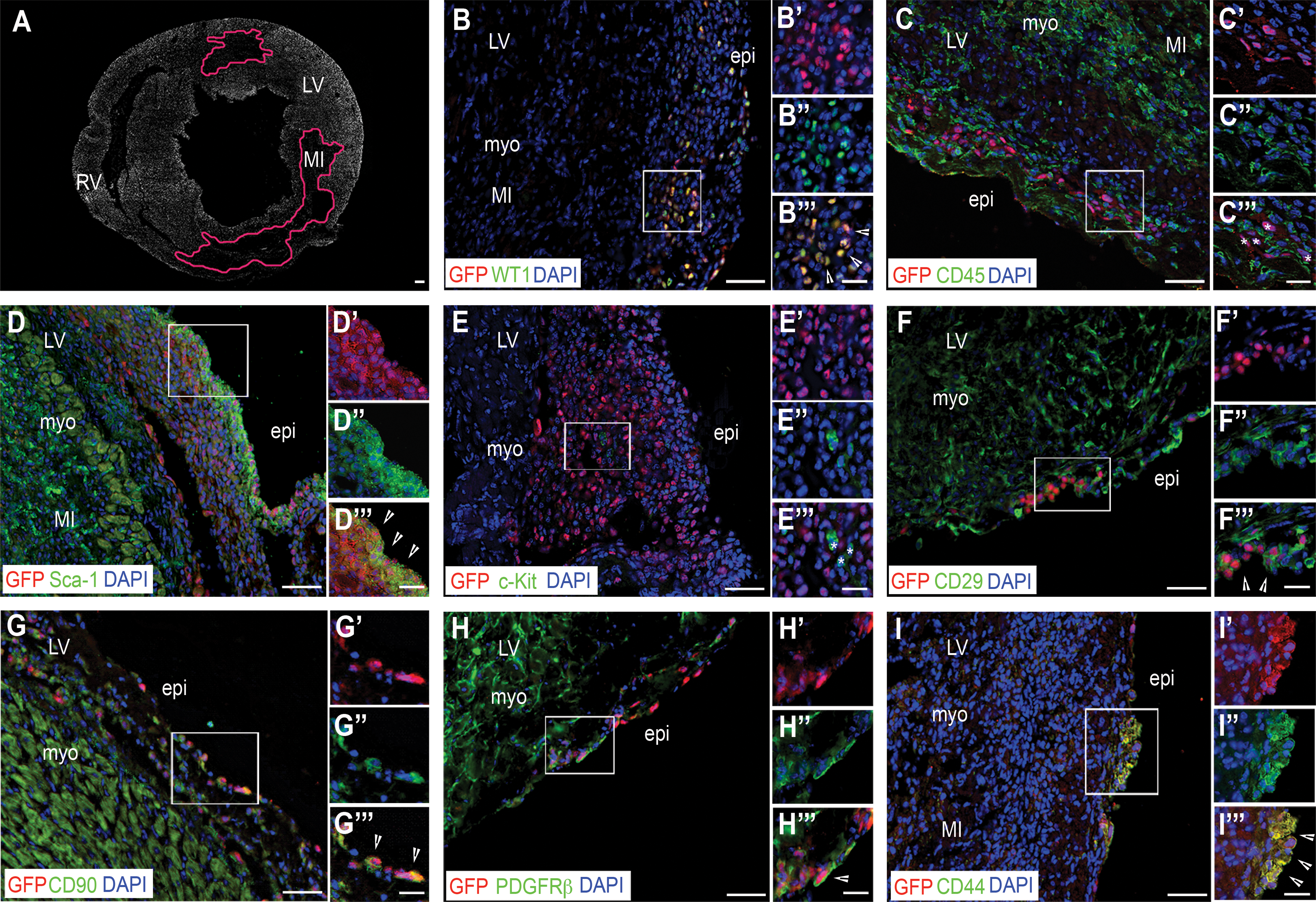
The adult-reactivated WT1+ GFP+ EPDCs reveal a highly heterogeneous phenotype with expression of specific cardiac progenitor and mesenchymal stem cell markers. In
eEPDCS E12.5, WT1+ GFP+ embryonic EPDCs isolated at E12.5; aEPDCs 2D MI, WT1+ GFP+ adult EPDCs isolated 2 days post-MI; aEPDCs 4D MI, WT1+ GFP+ adult EPDCs isolated 4 days post-MI; aEPDCs 7D MI, WT1+ GFP+ adult EPDCs isolated 7 days post-MI; EPDC, epicardium-derived cell; MI, myocardial infarction.
Given that a hallmark of adult-reactivated EPDCs after Tβ4-priming and MI is the re-expression of embryonic epicardial genes, including Wt1, Tbx18, and Raldh2 [29,40], we compared the phenotype of the adult WT1+ GFP+ EPDCs with their embryonic E12.5 WT1+ GFP+ counterparts (Table 1 and Fig. 1C). Embryonic WT1+ GFP+ EPDCs expressed CD29, PDGFRβ, and CXCR4 at similar levels compared with the adult population and were similarly negative for CD45, with equivalent low c-kit expression; however, their expression of Sca-1, CD90 (P<0.0001), CD44, and CD105 (P<0.01) was significantly lower as compared with the adult-reactivated WT1+ population.
Identification of subpopulations of adult-activated WT1+ GFP+ EPDCs after Tβ4 priming and injury
Given the heterogeneous profile of the adult-reactivated EPDCS, we further evaluated the presence of different subpopulations of progenitor cells based on the expression of Sca-1, as the most abundant cell surface marker (present in more than 50% of the WT1+ GFP+ EPDCs after MI). Sca-1 also clearly demarcated two sub-populations within the control (PBS-treated, n=12) animals (Supplementary Fig. 1C). Within the Sca-1+ cell fraction, we distinguished four different subpopulations according to the expression of CD90 and CD44. CD90+/CD44+ cells were 10.1%±1.5% (n=9 hearts), 5.3%±0.8% (n=12 hearts), and 5.4%±0.9% (n=9 hearts) of the Sca-1+ fraction at day 2, 4, and 7 post-MI, respectively. CD90+/CD44− cells were 44.7%±5.2% (n=9 hearts), 50.0%±3.2% (n=12 hearts), and 66.2%±4.4% (n=12 hearts) at day 2, 4, and 7 post-MI. CD90−/CD44+ cells were 4.8%±2.1% (n=9 hearts), 3.7%±0.9% (n=12 hearts), and 0.9%±0.2% (n=9 hearts) at day 2, 4, and 7 post-MI. CD90−/CD44− cells were 40.4%±4.1% (n=9 hearts), 37.4%±2.3% (n=12 hearts), and 27.6%±4.4% (n=9 hearts) of the Sca-1+ fraction at day 2, 4, and 7 post-MI. The majority of adult-reactivated WT1+ GFP+ Sca-1+ EPDCs were distributed according to the expression of CD90, with a minor proportion of cells characterized by the expression of CD44 (Fig. 3).
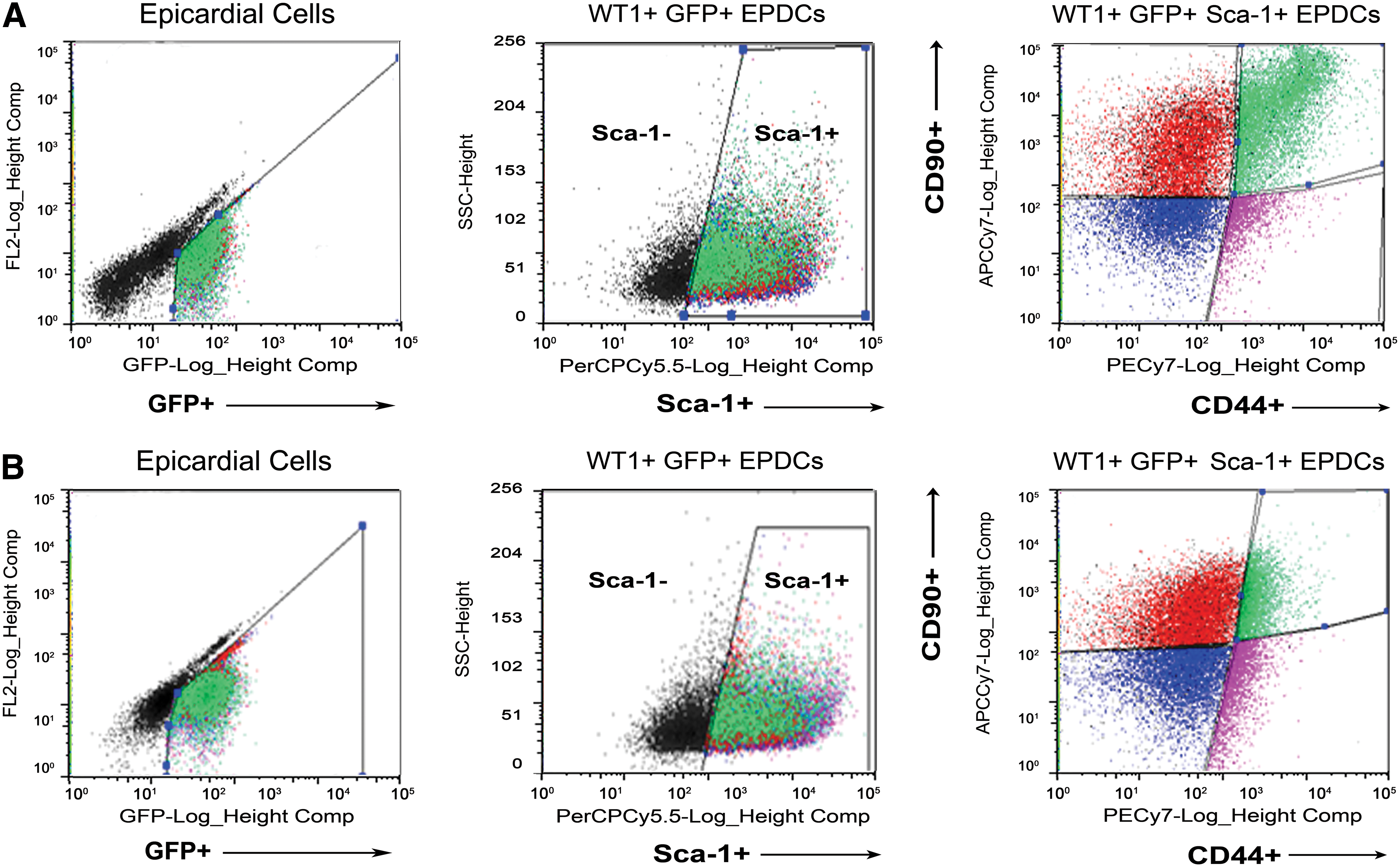
Sca-1 defines the majority of adult-reactivated WT1+ GFP+ EPDCs with cells equally distributed according to the expression of CD90 and a minor proportion of progenitors characterized by the high expression of CD44. WT1+ GFP+ were isolated from the total epicardial cells harvested by enzimatic digestion and subsequently, sorted on Sca-1 expression. Within the WT1+ GFP+ Sca-1+ EPDCs fraction, cells were further discriminated into four subpopulations based on the expression of CD90 and CD44. According to the level of reactivation, within the WT1+ GFP+ Sca-1+ CD90+ CD44+ population, it was possible to discriminate a subfraction of progenitor cells highly expressing CD90 and CD44, represented by the shifted population in green in the dot plot on the right
After Tβ4 priming and injury (Supplementary Fig. 2A), we isolated a sub-fraction of WT1+ GFP+ Sca-1+ cells that were highly positive for CD90 and CD44, here defined as CD90hi CD44hi and represented by the right-shifted population following flow cytometry analysis (Fig. 3A and Supplementary Fig. 2B). Comparing the CD90hi CD44hi subpopulation with the remaining CD90+ and low CD44+ cells within the WT1+ GFP+ Sca-1+ EPDCs, here defined as CD90+ CD44lo (Supplementary Fig. 2B, C), it was evident that the CD90+ CD44lo population constituted a more significant proportion of the total WT1+ GFP+ Sca-1+ population at 40% across days 2–7 post-MI, versus the 10% of CD90hi CD44hi at day 2, which was reduced to 5% by days 4 and 7 (Supplementary Fig. 2C); again supporting the conclusion of a heterogeneous-reactivated adult epicardial lineage.
Gene expression profile of adult-activated WT1+ GFP+ EPDCs subpopulations after Tβ4 priming and injury
We subsequently analyzed the potential of the adult-activated WT1+ GFP+ EPDCs isolated 4 days after MI, a stage at which they still retain a cardiac progenitor phenotype [29], by comparing the gene expression profiles for candidate lineage markers with embryonic epicardium at E12.5.
Comparisons were made between the adult WT1+ GFP+ Sca-1+ EPDCs, the Sca-1− sub-population, the remaining adult GFP− epicardial cells, and the embryonic E12.5 WT1+ GFP+ EPDCs (n=17 hearts, Fig. 4). Relative to the embryonic WT1+ GFP+ E12.5 cells, the adult WT1+ GFP+ Sca-1+ EPDCs revealed a significantly elevated expression of embryonic epicardial genes: Wt1 (7.6-fold increase, P<0.0001), Tbx18 (2.4-fold increase; P<0.05), Raldh2 (2.1-fold increase; P<0.05), and Pdgfrβ (16.5-fold increase; P<0.0001); whereas the expression of Tcf21/Epicardin was only elevated in the adult GFP− cells relative to the E12.5 WT1+ GFP+ cells (4.9-fold increase; P<0.001).
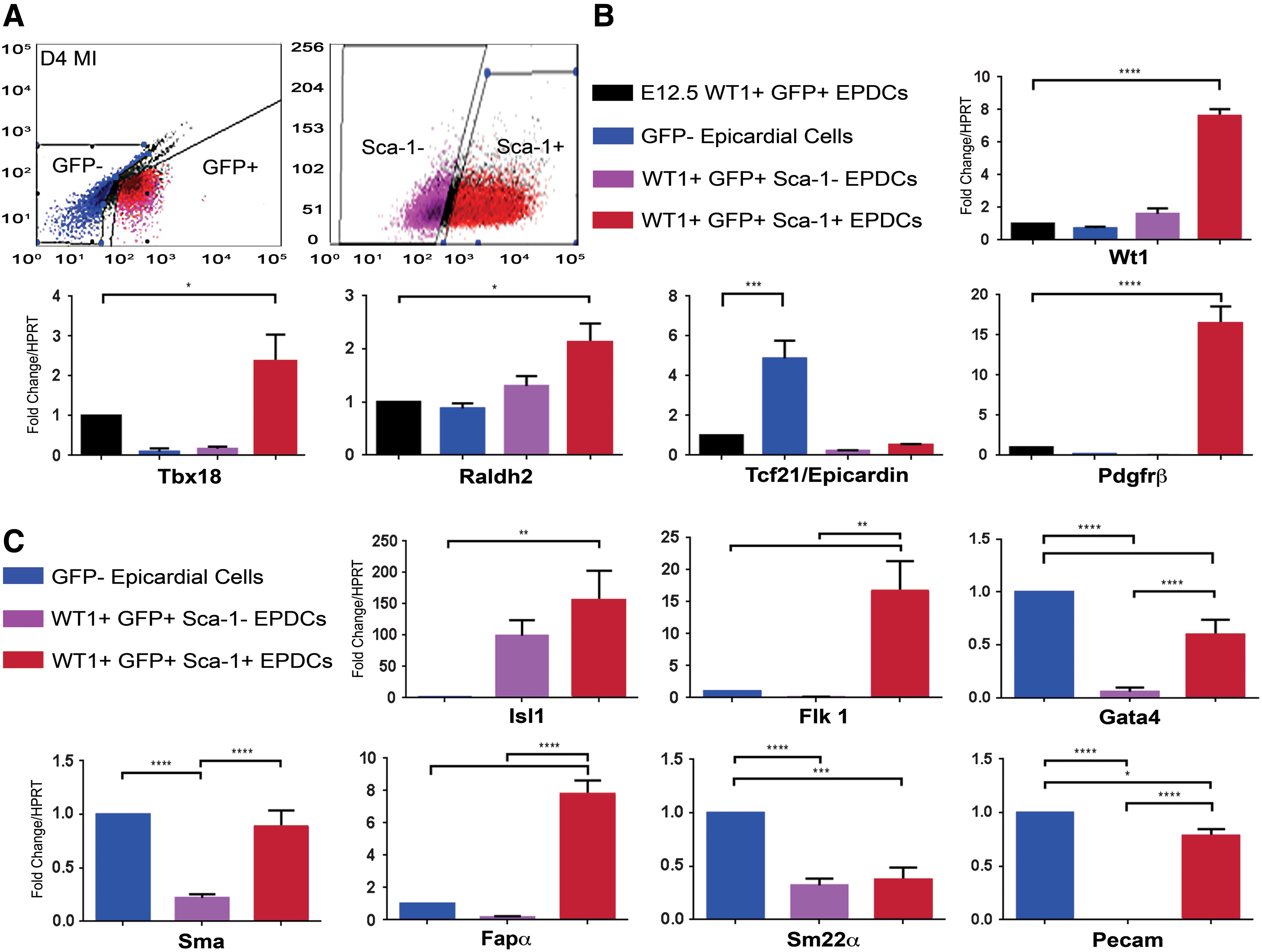
WT1+ GFP+ Sca-1+ progenitors represent the EPDC subpopulation with the strongest capacity to re-activate the developmental programme within the adult epicardium and with evidence of cardiac progenitor potential.
We further investigated the profile of the adult-activated WT1+ GFP+ Sca-1+ EPDCs with regard to the expression of Isl1, as a marker of heart precursors from the secondary heart field during development, and of postnatal cardioblasts [19,41,42], along with Flk1, which defines multipotent cardiovascular progenitors during development that reportedly give rise to cardiomyocyte, endothelial, and vascular smooth muscle lineages [43,44]. Comparing the Sca-1+ and Sca-1− subpopulations of the WT1+ GFP+ EPDCs with the adult GFP− fraction after MI, we observed that Sca-1+ cells expressed Isl1 and Flk1 at higher levels compared with the GFP− population (155- and 16.6-fold increase, respectively; P<0.01). In particular, while Isl1 expression was not significantly different between the Sca-1+ and Sca-1− fractions, Flk1 was differentially up-regulated in Sca-1+ cells (1,762-fold increase; P<0.01). Likewise, Gata4, which is an early cardiac transcription factor that is essential during cardiogenesis [45] and a marker of myocardial and vascular precursors in human and mouse epicardium [41,46], was more up-regulated in the Sca-1+ cells than in the Sca-1− (10-fold increase, P<0.0001), but expressed at relatively lower levels than in the GFP− epicardial cells (0.6- and 0.06-fold, respectively, P<0.0001).
An examination of more mature cardiovascular markers was performed to define the adult-activated WT1+ GFP+ EPDCs commitment to fibroblast, smooth muscle, and endothelial lineages. α-smooth muscle actin (Sma) was up-regulated in the adult WT1+ GFP+ Sca-1+ EPDCs compared with Sca-1− counterparts (4-fold increase, P<0.0001) but comparable to levels in the adult GFP− epicardial cells, (0.9-fold, not statistically different). The fibroblast activation protein alpha (Fapα) was more up-regulated in the Sca-1+ cells compared with the Sca-1− WT1+ GFP+ EPDCs and compared with the GFP− epicardial cells (47- and 7.7-fold increase respectively, P<0.0001). The vascular smooth muscle marker Sm22α was down-regulated in the adult WT1+ GFP+ Sca-1+ and Sca-1− EPDC fractions compared with the GFP− cells (0.4- and 0.3-fold, P<0.001 and P<0.0001), as was the endothelial cell marker Pecam, which was significantly less expressed in the two WT1+ GFP+ EPDCs populations (0.8- and 0.0007-fold, respectively, P<0.05 and P<0.0001), although the Sca-1+ fraction showed higher expression than the Sca-1− (approximately a 1,000-fold increase, P<0.0001). These results suggest a myo-fibroblast potential in the Sca-1+ fraction within the adult-activated WT1+ GFP+ EPDCs by virtue of the expression of Sma and Fapα at the expense of coronary vascular potential, given the low expression of Sm22α and Pecam when compared with the GFP− epicardial cell population. Notably, this potential was enhanced in Tβ4-treated animals, as compared with PBS-treated controls (n=12), by which Sma and Fapα expression was significantly elevated in Tβ4-treated GFP+ Sca-1+ and Sca-1− populations (Supplementary Fig. 1D).
Given our identification of a sub-fraction of WT1+ GFP+ Sca-1+ cells, which was highly positive for CD90 and CD44 at 4 days after MI, we further investigated the gene expression profile of these CD90hi CD44hi cells comparing them with the remaining WT1+ GFP+ Sca-1− fraction and calibrated against the adult GFP− epicardial cells (n=17 hearts; Fig. 5). These WT1+ GFP+ Sca-1+ CD90hi CD44hi progenitors showed a significant up-regulation of the specific embryonic epicardial genes Wt1 (10- and 4-fold increase relative to the WT1+ GFP+ Sca-1− and the Sca-1+ CD90+ CD44lo cells, P<0.05 and 26.2-fold more than the GFP− cells, P<0.01), Tbx18 (15- and 12.3-fold increase, respectively, compared to the WT1+ GFP+ Sca-1− and the Sca-1+ CD90+ CD44lo cells and 3.3-fold more than GFP− cells, P<0.01), Raldh2 (3- and 2.3-folds more than WT1+ GFP+ Sca-1− and GFP− cells; P<0.05), and Pdgfrβ (1.7-fold more than GFP− cells; P<0.05), as well as of the cardiac progenitor markers Gata4 (1,900- and 681-fold increase compared with WT1+ GFP+ Sca-1− and the Sca-1+ CD90+ CD44lo cells and 27.7-fold more than GFP− cells, respectively; P<0.01), Flk1 (57- and 67-fold increase when related to WT1+ GFP+ Sca-1− and the Sca-1+ CD90+ CD44lo cells, P<0.01 and 2.8-fold increase compared with GFP− cells, P<0.05) when compared with the other subpopulations of adult-activated cells analyzed here.

After Tβ4 priming and injury, the Sca-1+ CD90hi CD44hi fraction is the most highly activated WT1+ GFP+ EPDC population that retains cardiovascular multipotency via the expression of early cardiac progenitor markers.
These comparative expression data suggest that the population of WT1+ GFP+ Sca-1+ adult-activated EPDCs, further stratified by CD90hi CD44hi cells, is the most highly activated subpopulation of EPDCs after Tβ4-priming and injury, according to the relative expression of embryonic epicardial genes. Moreover, this sub-population appears to retain multipotency by virtue of the expression of early cardiac progenitor markers Isl1, Gata4, and Flk1.
Discussion
Regenerative medicine strategies based on CPC therapy have emerged as a therapeutic opportunity to facilitate heart repair. Much attention has been focused on the identification of the most suitable source of endogenous CPCs to mediate tissue regeneration after ischemic heart disease.
Of the different CPC populations described to date [18], EPDCs have emerged as a source with significant potential arising from their embryonic origin. During development, EPDCs are multipotent, giving rise to all the cardiovascular lineages [21,22,24,25]. However, in contrast, the adult epicardium is effectively dormant, having virtually lost all developmental plasticity in the mature heart [20]. A working hypothesis is that in order to realize the cardiovascular potential of adult EPDCs, it is necessary to reinstate their developmental program, through stimulation with the same molecules that regulate the lineage potential of the epicardium during embryogenesis. While the embryonic epicardium has been well characterized with distinct subsets of cells arising from the PEO, as defined by markers such as WT1, Tbx18, Tcf21, Sema3D, and Scleraxis, [21,25,27,47 –49], virtually nothing is known about the detailed composition of the adult lineage.
Using FACS analyses, we characterized the adult-reactivated WT1+ EPDCs after MI and demonstrated that these cells reveal a highly heterogeneous molecular phenotype. In particular, we focused our analysis on the expression of specific cardiac progenitor and mesenchymal stem cell markers within the adult-activated epicardium, such as Sca-1, c-kit, CD29, CD90, CD44, and CD105. Sca-1 has been widely reported to discriminate different subsets of cardiac resident stem/progenitor cells in the murine heart, with a significant role in cardiac repair [18,50 –52]. While Sca-1 is expressed by 50%–60% of the adult WT1+ EPDCs, another well-characterized cardiac progenitor/stem cell marker, c-kit, appeared to be present at extremely low level (<1% of the population were c-kit+), suggesting that the adult-reactivated EPDCs are a source of Sca-1+ CPCs with c-kit likely identifying a different progenitor compartment within the epicardium [46,48]. The adult WT1+ EPDCs appeared to have undergone EMT after injury, as confirmed by their broad expression for the mesenchymal antigen CD29. The WT1+ progenitors were further delineated as CD90+, CD44+, and CD105+, confirming a mesenchymal-like stem cell phenotype analogous to adult cardiac-resident stem cells of proepicardial origin [38]. While Sca-1, c-kit, CD29, and CD44 expression was equivalent across days 2, 4, and 7 post-MI; CD90, CD105, and additional markers PDGFRβ and CXCR4 revealed an increasing trend in their expression, most notably between days 4 and 7 after injury, suggesting the maturation, within a week after injury, of an activated mesenchymal phenotype, and one that is potentially more responsive in terms of proliferation and migration.
Reactivation of the developmental program in the dormant adult epicardium suggested that the adult lineage might revert entirely to an embryonic phenotype. Here, we reveal that the adult-reactivated WT1+ GFP+ EPDCs are, in fact, molecularly distinct from their embryonic counterparts at E12.5; a stage in development when embryonic EPDCs begin to actively contribute cardiovascular derivatives. The embryonic WT1+ epicardium revealed a significantly reduced expression for markers characterizing the adult WT1+ EPDCs, with the exception of CD29, PDGFRβ, and CXCR4, which were similarly expressed in both populations. This observation suggested that, despite the reactivation of the developmental program via re-expression of the embryonic gene Wt1, the adult EPDCs adopt a different phenotype in response to cardiac injury relative to embryonic EPDCs which contribute to the developing heart. Gene expression profiling revealed that differences in cell surface markers were also reflected at a transcriptional level. Our analysis focused on the Sca-1+ sub-population of WT1+ GFP+ adult progenitors at day 4 after MI. At this stage, EPDCs are still restricted to the epicardium and sub-epicardial space and present with an immature progenitor phenotype, after which they subsequently migrate toward the site of injury within the underlying myocardium and give rise to mature cardiovascular cells [29]. We first calibrated the gene expression fold changes against the embryonic E12.5 WT1+ EPDCs and compared the values obtained among the Sca-1+ and Sca-1− subsets of the adult equivalent versus the remaining GFP− cells populating the sub-epicardial space. Remarkably, the Sca-1+ subset of WT1+ progenitors revealed expression of developmental epicardial genes at levels significantly higher than their E12.5 counterparts, suggesting that the expression of Sca-1 might discriminate a fraction of cardiac progenitors which retained the strongest potential to re-activate the developmental program within the adult epicardium. We isolated the expanded sub-epicardial cell population after MI via enzymatic digestion and compared the myogenic and vascular profiles of the adult WT1+ GFP+ Sca-1+ and WT1+ GFP+ Sca-1− EPDCs against the GFP− epicardial cells after Tβ4 priming and MI. This analysis revealed that the Sca-1+ subpopulation was not only the most active in terms of initiating the developmental epicardial gene program, but also pre-empted cardiovascular lineage commitment, via significantly higher expression of both Isl1 and Flk1, the transcriptional signature of common primordial/multipotent cardiac progenitors, previously described during development [19,42,44]. In addition, the Sca-1+ EPDCs revealed a significant up-regulation of the fibroblast marker Fapα and the smooth muscle actin marker Sma when compared with the WT1+ GFP+ Sca-1− EPDCS and the other GFP− epicardial cells, consistent with a bias toward the (myo-) fibroblast lineage after injury [53,54]. Furthermore, the increased expression of (myo-) fibroblast gene expression was also further augmented by Tβ4 treatment, as compared with injury alone. The early cardiac marker Gata4 was down-regulated compared with the GFP− epicardial cells, suggesting that this latter population might retain a higher myogenic potential. Notably, smooth muscle and endothelial vascular markers Sm22α and Pecam were down-regulated in both subsets stratified by Sca-1 expression within the WT1+ progenitors, confirming that neovascularization, which is augmented after injury by Tβ4 priming, appears to be triggered by a distinct Tβ4-activated subset of epicardial progenitors, with limited involvement of the WT1+ GFP+ EPDCs.
We further investigated the distribution of mesenchymal markers CD90 and CD44 within the Sca-1+ fraction of the adult WT1+ epicardium. Tβ4 priming and reactivation after injury facilitated isolation of a fraction of cells with high levels of CD44 and CD90, identifying a sub-population fated for potential roles in cell–cell interaction, cell adhesion, or migration. This Sca-1+ fraction of WT1+ progenitors retained the most embryonic potential via surrogate expression of the developmental epicardial genes. In addition, the same sub-population revealed expression of the cardiovascular genes Gata4, Isl1, and Flk1, suggesting that it may constitute derivatives of the embryonic Isl1+/Flk1+ population and an equivalent bipotential toward the vascular smooth muscle and cardiomyocyte lineages [44].
Understanding the composition of the adult-activated epicardium alongside the cardiomyogenic or cardiovascular potential of the resident sub-populations provides important insight into the deployment of resident CPCs for cardiac repair. The potential to instigate tissue repair has to be off-set by the fact that injury promoted a (myo-) fibroblast fate within the Sca-1+ sub-population, which could result in increased collagen deposition and scarring. While fibrosis and scar formation is the default wound-healing response in the injured hearts of adult mammals, including humans, this establishes a local environment that is incompatible with cell replacement. Thus, a (myo-) fibroblast fate would need to be modulated to establish more conducive conditions for cardiovascular regeneration. That said, an important balance has to be struck to prevent cardiac rupture at the site of injury and, moreover, to utilize important reciprocal signaling between Sca-1+ epicardium-derived fibroblasts and coronary vascular cells and cardiomyocytes, as is known to occur during developmental stages [55].
It remains to be determined whether the epicardium in human patients offers an analogous potential to contribute toward tissue regeneration after ischemic injury. Studies that realize a therapeutic application of epicardium-derived CPCs will be required to exploit aged animal models, given the prevalence of cardiovascular disease and heart failure in the aging human population, notwithstanding the risk that CPC potential itself may be compromised with age [56]. Moreover, in vivo studies on human patients are essentially limited to functional imaging (eg, via MRI), which has yet to reach cellular resolution for tracking CPCs during ischemic events. Instead, studies that can take advantage of surgical biopsies (eg, after right coronary artery bypass) may pave the way for ex vivo analyses of human primary epicardial CPCs to facilitate a future pathway toward drug discovery.
Footnotes
Acknowledgments
This work was funded by the British Heart Foundation (RG 08/003/25264). The authors are grateful to Ayad Eddaoudi from the UCL Institute of Child Health Flow Core Facility for assistance in flow cytometry analysis and to RegeneRx Biopharmaceuticals for providing clinical-grade Tβ4.
Author Disclosure Statement
The authors have no conflict of interests or any disclosure to declare. No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.