
Abstract
Somatic cells are reprogrammed to induced pluripotent stem cells (iPSCs) by overexpression of a combination of defined transcription factors. We generated iPSCs from mouse embryonic fibroblasts (with Oct4-GFP reporter) by transfection of pCX-OSK-2A (Oct4, Sox2, and Klf4) and pCX-cMyc vectors. We could generate partially reprogrammed cells (XiPS-7), which maintained more than 20 passages in a partially reprogrammed state; the cells expressed Nanog but were Oct4-GFP negative. When the cells were transferred to serum-free medium (with serum replacement and basic fibroblast growth factor), the XiPS-7 cells converted to Oct4-GFP-positive iPSCs (XiPS-7c, fully reprogrammed cells) with ESC-like properties. During the conversion of XiPS-7 to XiPS-7c, we found several clusters of slowly reprogrammed genes, which were activated at later stages of reprogramming. Our results suggest that partial reprogrammed cells can be induced to full reprogramming status by serum-free medium, in which stem cell maintenance- and gamete generation-related genes were upregulated. These long-term expandable partially reprogrammed cells can be used to verify the mechanism of reprogramming.
Introduction
Y
Some pluripotency markers are essential for complete reprogramming and maintenance of the ground state of pluripotency. Expression of endogenous Nanog is a crucial event during reprogramming into the pluripotent ground state [19]. Nanog has the capacity to overcome multiple barriers to reprogramming in mouse cells [20]. NANOG also enhances molecular reprogramming in human somatic cells [21]. In this study, we found that Nanog is expressed in partially reprogrammed cells, which self-renewed for more than 20 passages in vitro. These cells were converted into fully reprogrammed iPSCs with mESC-like properties in serum-free medium [with serum replacement (SR) and basic fibroblast growth factor (bFGF)]. In addition, global gene expression profiles and gene ontology (GO) revealed that the genes associated with partial reprogramming were related to stem cell maintenance, survival, and germ cell development.
Materials and Methods
Cell culture
We used MEFs as somatic cells for reprogramming. MEFs were derived from OG2/Rosa26 heterozygous double transgenic 13.5-day postcoitum (dpc) mouse embryos, which were generated by crossing the Rosa26 (carrying neo/lacZ transgene) strain with the OG2 transgenic strain (carrying GFP under the control of the Oct4 promoter, Oct4-GFP) over several generations [22,23]. Animal handling was in accordance with the animal protection guidelines of Konkuk University and Korean animal protection laws. MEFs were maintained in fibroblast medium: high-glucose Dulbecco's modified Eagle's medium (DMEM; Gibco BRL) containing 10% fetal bovine serum (FBS; HyClone) and 0.5% penicillin/streptomycin (Invitrogen). Mouse ESCs and iPSCs were grown on MEF feeder cells that had been inactivated with 0.01 mg/mL mitomycin C in standard mouse ESC culture medium: DMEM supplemented with 15% FBS, 0.5% penicillin/streptomycin, nonessential amino acids (NEAA; Gibco BRL), 0.1 mM 2-mercaptoethanol, and 1,000 U/mL leukemia inhibitory factor (LIF) (ESGRO; Chemicon). XiPS-7 cells were reprogrammed on inactivated MEFs in KOSR-based medium: DMEM/F12 (Gibco BRL) containing 20% knockout SR (Gibco BRL), 2 mM glutamine, NEAA, and 5 ng/mL bFGF.
Generation of iPSCs
pCX-OKS-2A [Oct4 (O), Klf4 (K), and Sox2 (S), each separated by a different 2A sequence] and pCX-cMyc, were purchased from Addgene. The plasmids were mixed with 3 μg pCX-OKS-2A and 1 μg pCX-cMyc. MEFs were seeded at 1×105 cells/well in six-well plates (day 0). Plasmids were introduced with 1.2 μL of Xfect™ transfection reagent (Clontech) according to the manufacturer's instructions (Fig. 1A). From day 4, the transfected MEFs were cultured in mouse ESC culture medium containing LIF. On day 9, the cells were harvested with trypsin and plated on 100-mm dishes with MEF feeder cells. On days 25–28, Oct4-GFP-positive or -negative colonies were picked for expansion and maintained in mouse ESC culture medium.
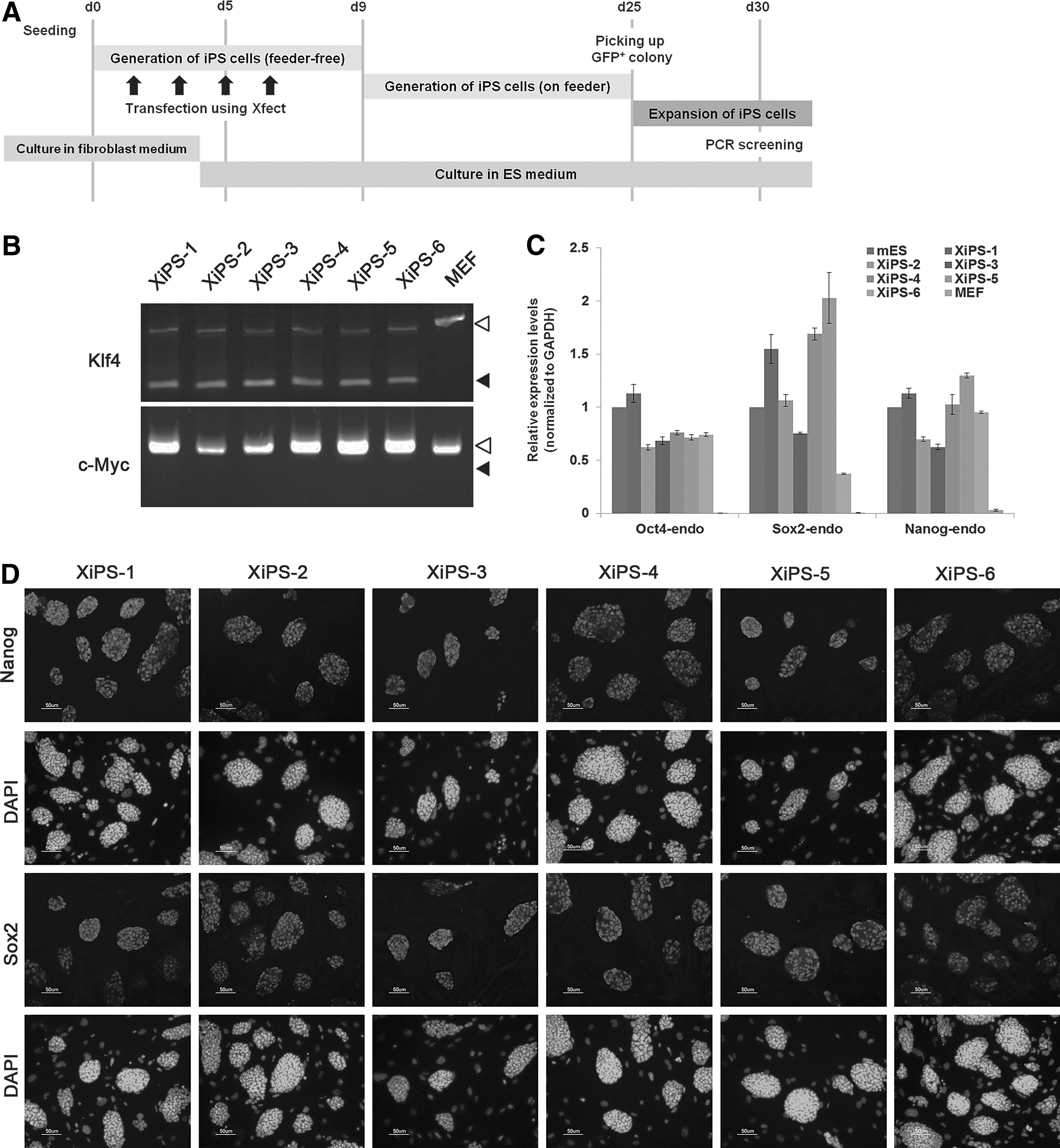
Generation of induced pluripotent stem (iPS) cells using pCX-OSK (Oct4, Sox2, and Klf4) and pCX-cMyc.
Integration analysis
Transgene integration was analyzed by polymerase chain reaction (PCR): 50 ng genomic DNA was amplified with Taq polymerase and screened with primer sets for pCX-OKS-2A (primers for Klf4) and pCX-cMyc [24]. Primers are listed in Supplementary Table S1 (Supplementary Data are available online at
Bisulfite DNA sequencing
Genomic DNA was treated with sodium bisulfite to convert all unmethylated cytosine residues into uracil residues using EpiTect Bisulfite Kit (Qiagen) according to the manufacturer's protocol. Briefly, purified genomic DNA (0.5–1 μg) was denatured at 99°C and then incubated at 60°C. Modified DNA (ie, after desulfonation, neutralization, and desalting) was diluted with 20 μL distilled water. Bisulfite PCR amplification was performed with 1–2 μL aliquots of modified DNA for each reaction. PCR amplification of the Oct4 and Nanog promoter regions was performed as described [25]. The PCR products were subcloned into pGEM-T Easy (Promega). The reconstructed plasmids were purified, and individual clones were sequenced (Solgent Corporation). Clones with ≥90% cytosine conversion were accepted, and all possible clonalities were excluded based on criteria in BiQ Analyzer software (Max Planck Society). Primers are listed in Supplementary Table S1.
Alkaline phosphatase staining
Alkaline phosphatase (AP) staining was performed with the AP detection kit (Millipore) according to the manufacturer's instructions. Cells were fixed with 4% paraformaldehyde for 2 min at room temperature, washed twice with PBS, and treated with AP solution containing Fast Red Violet and Naphthol AS-BI phosphate (Millipore) for 15 min at room temperature.
Immunocytochemistry
Cells were fixed with 4% paraformaldehyde for 20 min at room temperature and washed with PBS. Cells were treated with PBS containing 10% normal goat serum and 0.03% Triton X-100 for 45 min at room temperature. Primary antibodies were anti-Nanog (1:250 dilution; Chemicon), anti-Sox2 (1:1,000 dilution; Chemicon), anti-SSEA-1 (1:250 dilution; R&D), anti-β III tubulin (Tuj1; 1:1,000 dilution, Chemicon), anti-Brachyury (1:1,000 dilution; Chemicon), and anti-Sox17 (1:200 dilution; R&D). Fluorescence-labeled (Alexa Fluor 568; Molecular Probes) secondary antibody was used according to the manufacturer's specifications.
In vitro differentiation
iPSCs were transferred into a suspension culture dish after trypsinization and cultured for 3 days in DMEM (10% FBS) without LIF. The embryoid bodies (EBs) were plated on 0.1% gelatin-coated glass chamber slides and cultured in differentiation medium (DMEM supplemented with 20% FBS in the absence of LIF) for 12 days. Differentiated cells were stained for germ layer markers: Tuj1 for ectoderm, Brachyury for mesoderm, and Sox17 for endoderm.
Teratoma formation
The properties associated with teratoma formation were evaluated in trypsin-dissociated iPSCs. We subcutaneously transplanted 1×106 cells suspended in DMEM into the testis of 5-week-old nude mice. After 5 weeks, the tumors were dissected and fixed in 4% PFA, processed through graded ethanol, and embedded in paraffin after sectioning with hematoxylin/eosin.
Chimera formation analysis
XiPS cells were aggregated with denuded postcompacted eight-cell-stage embryos to obtain an aggregate chimera. Eight-cell embryos flushed from 2.5-dpc B6D2F1 female mice were cultured in microdrops of embryo culture medium under mineral oil. After cells were trypsinized for 10 s, clumps of iPSCs (4–10 cells) were selected and transferred into microdrops containing zona-free eight-cell embryos. Morular-stage embryos aggregated with XiPS cells were cultured overnight at 37°C, 55% CO2. The aggregated blastocysts were transferred into one uterine horn of 2.5-dpc pseudopregnant recipients. Animals were maintained and used for experimentation under the guidelines of the Institutional Animal Care and Use Committee of the Max-Planck Institute for Molecular Biomedicine and Konkuk University. The institutional review board specifically approved this study (IACUC090003).
X-gal staining
For whole fetal embryo staining, collected fetuses were rinsed with PBS and fixed in 4% formaldehyde for 1 h at 4°C. Fetuses were rinsed thrice at room temperature in PBS containing 5 mM EGTA, 0.01% deoxycholate, 0.02% NP40, and 2 mM MgCl2. The specimens were washed with PBS and stained in X-gal staining solution: PBS supplemented with 1 mg/mL 5-bromo-4-chloro-3-indolyl-galactosidase (X-gal; Promega), 5 mM K2Fe(CN)6, 5 mM K4Fe(CN)6, and 1 mM MgCl2. Blue staining was visualized by light microscopy.
Quantitative real-time PCR
Total RNA was extracted with the MiniRNeasy Kit (Qiagen) according to the manufacturer's instructions. Total RNA (1 μg) was reverse transcribed with SuperScriptIII Reverse Transcriptase (Invitrogen) and Oligo (dt)20-primer (Invitrogen) according to the manufacturer's instructions. Quantitative real-time PCR was performed with SYBR Green quantitative PCR mix (Takara) and a LightCycler5480 instrument (Roche). Duplicate amplifications were performed for each target gene with three wells serving as negative controls. Quantities were estimated by the comparative Ct method normalized to GAPDH and presented as a percentage of biological controls. Primer sequences are described in Supplementary Table S1.
Microarray-based analysis
Total RNA was isolated with the RNeasy Mini Kit (Qiagen) and digested with DNase I (RNase-free DNase; Qiagen) according to the manufacturer's instructions. Total RNA was amplified, biotinylated, and purified using the Ambion Illumina RNA amplification kit (Ambion) according to the manufacturer's instructions. Labeled cRNA samples (750 ng) were hybridized to each MouseRef-8 v2 Expression BeadChip. Signal detection was performed with Amersham Fluorolink Streptavidin-Cy3 (GE Healthcare Bio-Science) according to the bead array manual. Arrays were scanned with an Illumina Bead Array Reader according to the manufacturer's instructions.
Raw data were extracted using the software provided by the manufacturer (Illumina GenomeStudio v2011.1, Gene Expression Module v1.9.0). Array data were filtered by a detection P value <0.05 in at least 50% samples. Selected probe signal was log transformed and normalized by the quantile method. Comparative analysis was performed using LPE test and fold change. False discovery rate was controlled by adjusting the P value with the Benjamini–Hochberg algorithm. Hierarchical clustering was performed using complete linkage and Pearson distance as a measure of similarity.
Results
Generation of fully reprogrammed iPSCs by transfection of plasmid DNAs
To generate iPSCs without viral vectors, we transfected pCX-OKS-2A (Oct4, Klf4, and Sox2) and pCX-cMyc [26] into MEFs using a biodegradable nanoparticle, Xfect. Since MEFs were derived from the Oct4-GFP/Rosa26 double transgenic mouse, GFP was activated on pluripotential reprogramming. The timeline of the reprogramming protocol is depicted in Fig. 1A. Serial transfection (four times) of the vectors into MEFs successfully generated GFP-positive colonies at 25 days after nanoparticle-mediated transfection (Supplementary Fig. S1A). We derived six iPS cell lines, referred to as “XiPS cells,” from Xfect-mediated transfection (lines 1–6). All six XiPS cell lines expressed Oct4-GFP and were positive for AP activity (Supplementary Fig. S1B). To test for transgene integration into the host genome, we genotyped the XiPS cell lines using two primer pairs (Klf4 and c-Myc) specifically designed to amplify regions of the pCX-OKS and pCX-cMyc vectors [24]. All six XiPS cell lines were free of pCX-cMyc but had integrated pCX-OKS (Fig. 1B). Endogenous pluripotency markers Oct4, Sox2, and Nanog were expressed in all XiPS cell lines, comparable to control mESCs (Fig. 1C). Immunocytochemistry also confirmed the expression of Nanog and Sox2 (Fig. 1D).
Oct4-GFP-positive iPSCs (XiPS-1 line) were differentiated in vitro into ectoderm (Tuj1), mesoderm (Brachyury), and endoderm (Sox17) (Fig. 2A). XiPS-1 cells also formed teratomas containing cells of all three germ layers after transplantation into the testis capsule of immunocompromised mice (Fig. 2B). To confirm the developmental potential of XiPS-1 cells, we performed chimera formation analysis by aggregation with zona-free morula embryos. After transfer into the uterus of pseudopregnant mice, the XiPS-1 cells formed chimeric embryos (13.5 dpc) showing germline contribution (Fig. 2C). These results demonstrate that transfection of plasmid DNA using nanoparticles can reprogram MEFs to pluripotent cells with properties similar to those of mouse ESCs.
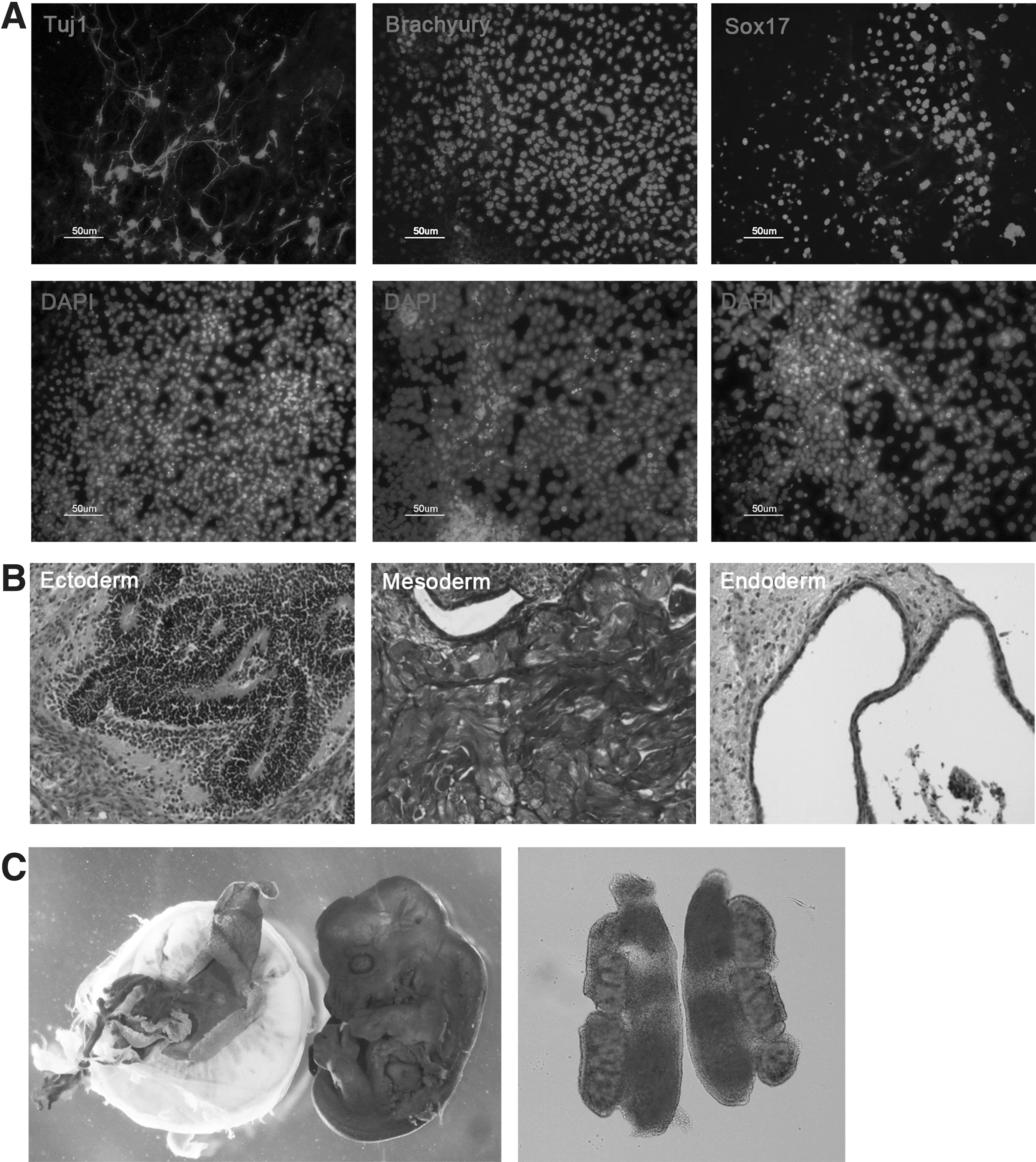
In vitro and in vivo differentiation potential of XiPS-1 cells.
Generation of long-term expandable partially reprogrammed cells by plasmid transfection
Although many Oct4-GFP-positive colonies were generated by plasmid-nanoparticle transfection, we also observed mESC-like cells forming relatively flat colonies without Oct4-GFP expression in the same culture dish (Fig. 3A). When the GFP-negative ESC-like cells were picked and cultured under mESC conditions, ESC-like colonies without Oct4-GFP activation were maintained; these were called the XiPS-7 cell line. Although the established XiPS-7 cells did not express Oct4-GFP for approximately 20 passages, they were positive for AP (Fig. 3B). Since they were Oct4-GFP negative, XiPS-7 did not express endogenous Oct4 (Fig. 4B); however, XiPS-7 expressed high levels of Nanog (Fig. 4B, C). Therefore, we regarded XiPS-7 as partially reprogrammed cells that were long-term expandable.
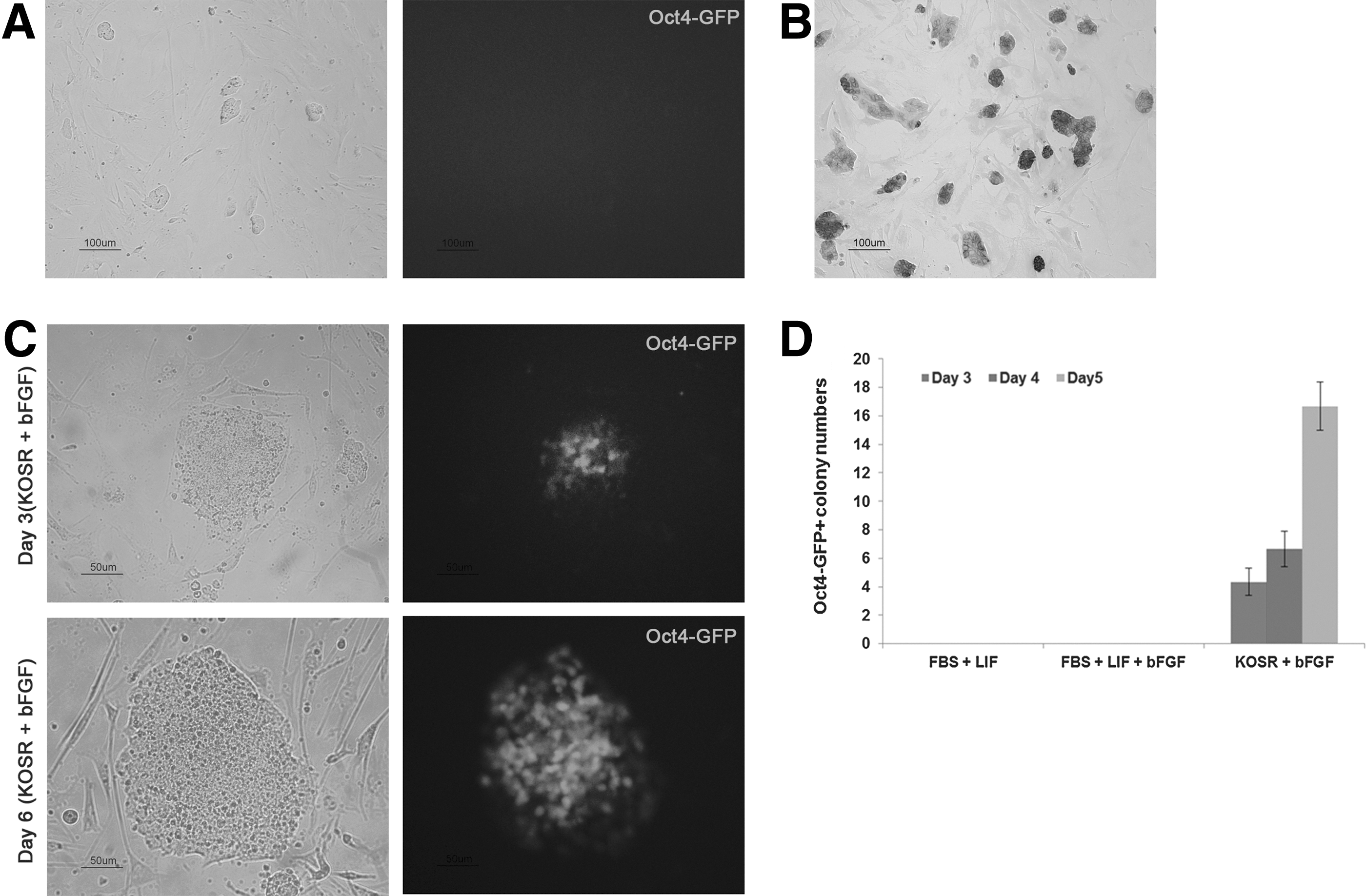
Successful conversion from Oct4-GFP-negative cells to Oct4-GFP-positive cells.
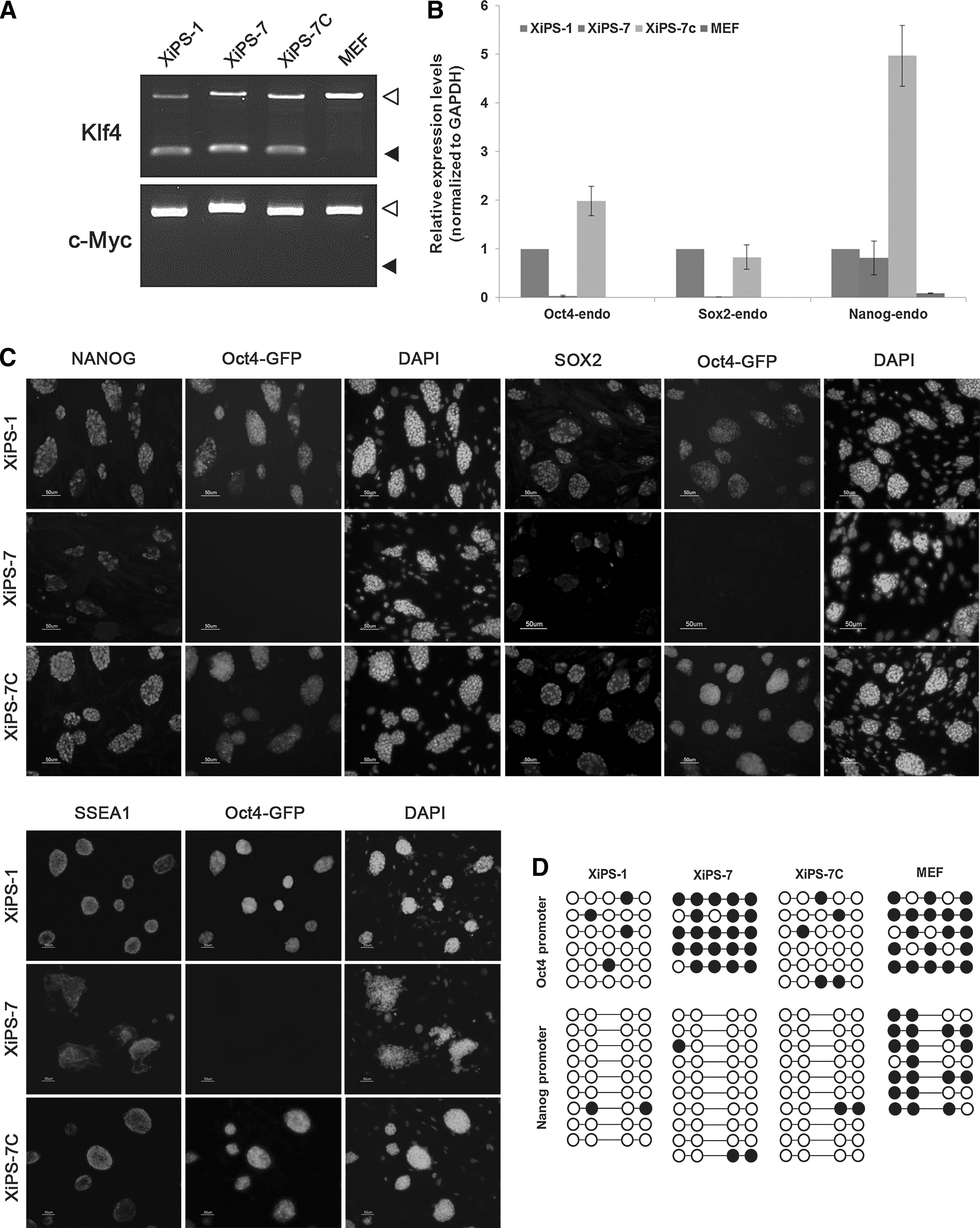
Characterization of XiPS-7 and XiPS-7c cells.
Partially reprogrammed cells (XiPS-7) progress through reprogramming in serum-free defined medium
Typically, mouse iPSCs are generated from somatic cells in conventional ESC culture conditions with FBS and LIF with feeders' support [1,2,27,28]. LIF and BMP signaling pathways are essential for maintaining the pluripotency of mESCs, whereas the bFGF supplementation leads to differentiation [29 –31]. ESCs cultured in FBS and LIF receive differentiation signals from FBS, which were blocked by LIF [32,33]. Therefore, serum-free culture may be suitable for ground-state pluripotency. Thus, iPSC colonies can be obtained at day 8 postinfection when reprogramming occurs in defined SR-based medium with LIF [23,34]. Moreover, iPSCs could also be generated and maintained in mESC medium supplemented with bFGF [35]. Therefore, we examined XiPS-7 cell culture in serum-free medium supplemented with bFGF. To our surprise, at day 3 after culture in SR-based medium, colonies of XiPS-7 cells were converted to dome-like colonies and became Oct4-GFP positive (Fig. 3C). These converted Oct4-GFP-positive (XiPS-7c) cells were picked and expanded. To verify that complete reprogramming of XiPS-7 to XiPS-7c is caused by serum-free bFGF culture conditions, XiPS-7 cells were cultured in three different media: mESC medium with LIF or with both LIF and bFGF, and SR-based medium with bFGF. Oct4-GFP-positive cells were observed in SR-based media but not in mESC medium with LIF or LIF/bFGF (Fig. 3D). Next, we checked whether these results were reproducible in another partially reprogrammed iPS cell line, XiPS-13 cells, which were Oct4-GFP negative but expressed Nanog. XiPS-13 cells also could be converted to dome-like colonies and became Oct4-GFP positive at day 2–4 after culture in SR-based medium (Supplementary Fig. S2A). These results suggest that partially reprogrammed cells progress through reprogramming in serum-free (SR-based) medium.
Partially reprogrammed cells (XiPS-7) express Nanog but not Oct4 and Sox2
We tested for genomic integration of pCX-OKS and pCX-cMyc in XiPS-7 and XiPS-7c cells and found that the XiPS-7 cells were free of pCX-cMyc but had pCX-OKS integration (Fig. 4A). Nevertheless, we speculated that XiPS-7c cells could be in a completely reprogrammed state and were pluripotent unlike XiPS-7 cells, which are in an intermediate state. To determine whether XiPS-7c cells were completely reprogrammed pluripotent, we analyzed transcript levels of endogenous core pluripotency markers Oct4, Sox2, and Nanog (Fig. 4B and Supplementary Fig. S3). Interestingly, Nanog was expressed in XiPS-7 cells in which endogenous Oct4 and Sox2 were not expressed (Fig. 4B). Nanog protein was also detected in XiPS-7 cells but not expressed in MEFs (Supplementary Figs. S4 and S5). A recent report demonstrated that the inner cell mass of Oct4-deficient blastocysts in which both maternal and zygotic Oct4 were silent still expressed Nanog [36], suggesting that Oct4 and Nanog do not need to be co-expressed in certain cells. However, XiPS-7c cells expressed all three core transcription factors, Oct4, Sox2, and Nanog. Gene expression was confirmed by immunocytochemistry. Oct4-GFP-positive XiPS-7c cells stained positive for Nanog, Sox2, and SSEA-1; whereas XiPS-7 cells were positive for Nanog and SSEA-1 but weakly positive for Sox2 (Fig. 4C). Next, the DNA methylation status of the Oct4 and Nanog promoter regions was determined by bisulfite genomic sequencing. The Oct4 promoter regions, which were hypermethylated in XiPS-7, were demethylated after conversion to XiPS-7c (Fig. 4D); however, the Nanog promoter region of all three Nanog-expressing cell types (XiPS-1, XiPS-7c, and XiPS-7) was completely unmethylated (Fig. 4D).
Gene expression profile of XiPS-7 and XiPS-7c cells
To compare the molecular signatures of XiPS-7, XiPS-7c, and XiPS-1 cells, we performed gene expression profiles using microarrays (Illumina's MouseRef-8 v2 Expression BeadChip). Counting only those with a fold change (FC) greater than 2, there were 5,807 regulated transcripts (with regard to mESCs). Pearson correlation analysis was used to cluster the cells. The heat map and hierarchical clustering analyses showed that the global gene expression patterns of XiPS-1 and XiPS-7c were similar to mESCs, whereas XiPS-7 clustered closer to MEFs than to mESCs, XiPS-1, and XiPS-7c (Fig. 5A, B). Scatter plot analysis also showed that XiPS-7 was more differentiated from mESCs than from XiPS-7c and XiPS-1 (Fig. 5C). Expression of pluripotency-related genes in XiPS-7 clustered differently from mESC, XiPS-1, and XiPS-7c (except Cd9, Nr5a2, and Nanog) (Fig. 5D and Supplementary Fig. S2B). Since XiPS-7 do not express endogenous Oct4, we checked the expression of Oct4 target genes. As expected, XiPS-7 expressed very low levels of Oct4 target genes (except Nanog) in comparison to completely reprogrammed cells that express Oct4-GFP (XiPS-1 and XiPS-7c) (Fig. 5E). In contrast, Oct4 target genes that are not expressed in ESCs, such as Casp6, Sh3glb1, Tdrd7, and Bmi1, were highly expressed in XiPS-7 and MEF (Fig. 5E).
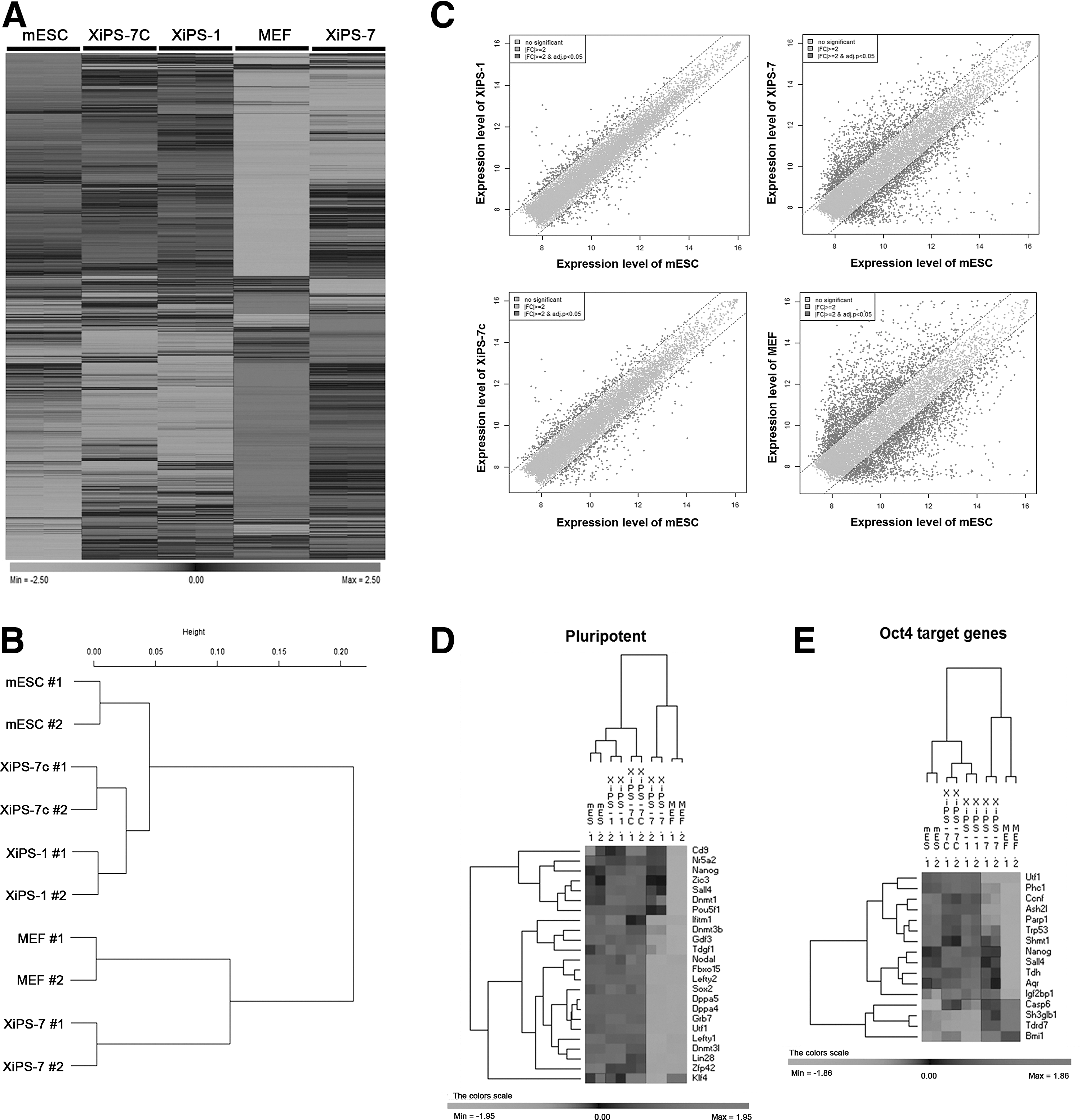
Contrasting global transcriptomes of five cell samples.
GO analysis was conducted to categorize the functions of the differentially expressed genes in partially (XiPS-7) and fully reprogrammed cells (XiPS-7c). We isolated differentially expressed genes [>±3-FC] in XiPS-7 versus XiPS-7c cells. We found that 625 probes were upregulated and 421 probes were downregulated in XiPS-7 versus XiPS-7c cells. We performed the GO term and KEGG pathway annotation using DAVID (
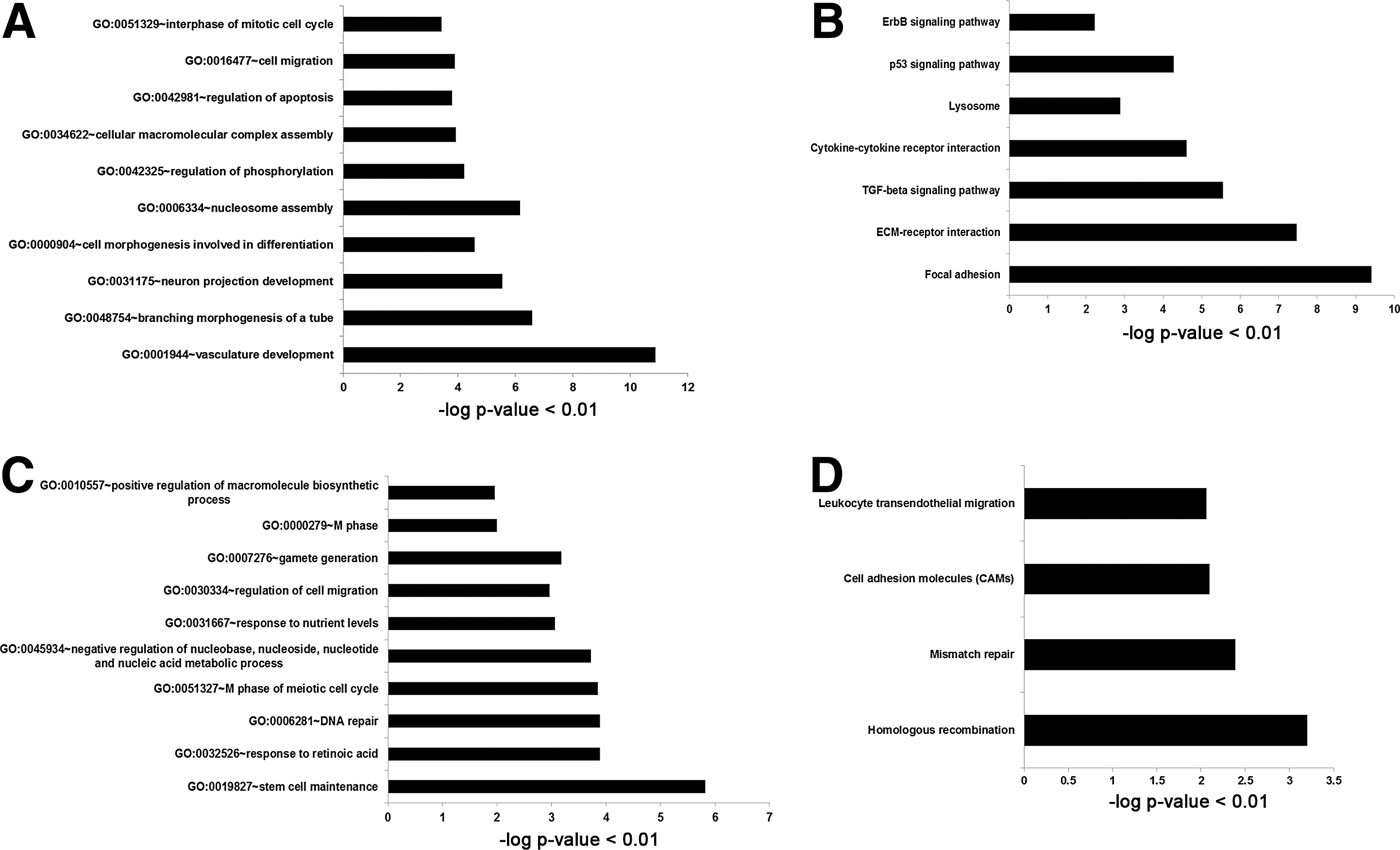
Gene ontology (GO) analysis of three-fold changed genes from XiPS-7 cells to mESCs.
Differentiation potential of XiPS-7 and XiPS-7c cells
Next, we compared the differentiation potential of XiPS-7c and XiPS-7 cells. Although XiPS-7 cells did not efficiently form EBs (data not shown), XiPS-7c cells efficiently formed EBs and differentiated into cells positive for Tuj1 (ectoderm), Brachyury (mesoderm), and Sox17 (endoderm) (Fig. 7A). XiPS-7c cells formed teratoma-containing lineages of all three germ layers (Fig. 7B); mesodermal differentiation was rarely detectable in XiPS-7. Teratoma analysis revealed that XiPS-7 cells preferentially differentiated into endodermal and ectodermal lineages; teratomas contained mostly endodermal (gut-like and secretary epithelium mixed form) and ectodermal (neural rosettes) cell types (Supplementary Fig. S7A, B). Moreover, XiPS-7c cells contributed to germline chimera (13.5 dpc) after morula aggregation and transfer into the uterus of pseudopregnant mice (Fig. 7C), which was not observed in XiPS-7 cells (data not shown). These results suggest that XiPS-7 cells represent partially reprogrammed cells and XiPS-7c represent fully reprogrammed cells.
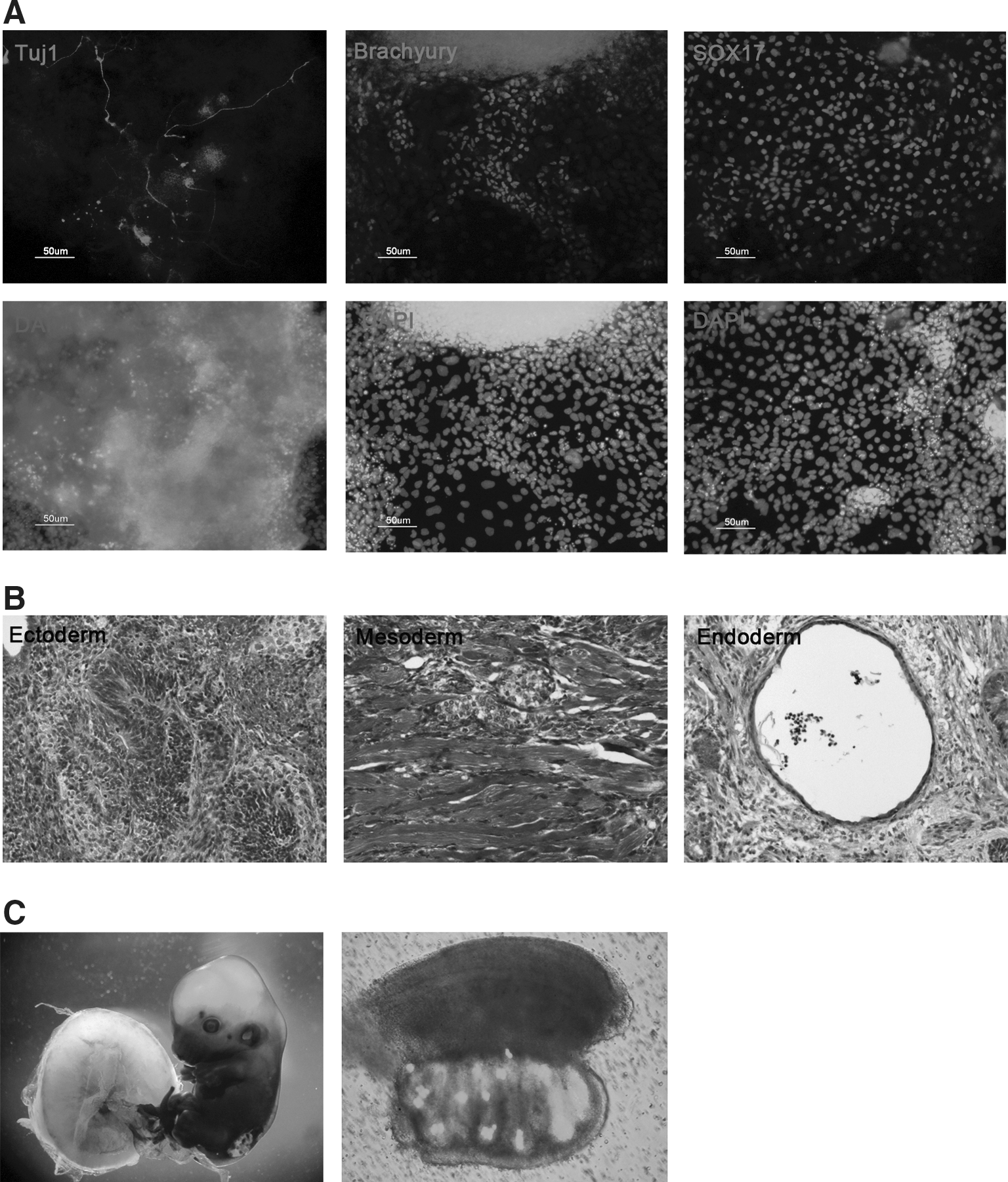
In vitro and in vivo differentiation potential of XiPS-7c cells.
Discussion
Since the discovery of somatic cell reprogramming by defined factors, various studies have focused on the technical development of iPSC establishment and mechanistic characterization of the sequential steps of reprogramming. To study the mechanism of reprogramming, previous studies analyzed intermediate cells before complete reprogramming; however, these intermediate cells are not in a specific stage of reprogramming but comprise a heterogeneous population containing cells in early- to late-stage reprogramming and of various transgenic modifications by random integration of reprogramming genes. In this study, we cultured intermediate cell lines, which were long-term expandable without expression of Oct4-GFP. These partially reprogrammed XiPS-7 cells formed flat colonies and did not express endogenous Oct4 and Sox2. Interestingly, XiPS-7 cells converted to fully reprogrammed iPSCs (XiPS-7c) in serum-free (SR-based) medium with bFGF. Typically, SR and bFGF are used in culture for mouse epiblast stem cells [37], human ESCs [38], and human iPS cell generation [7]; whereas bFGF supplementation leads to differentiation in mESCs [29,31]. However, according to recent studies, bFGF plays a positive role in mouse somatic reprogramming. Mouse iPSC colonies can be obtained when serum-based media is changed to SR-based media, including LIF [23] and derived in serum-based media in the presence of bFGF [35,39]. SR contains abundant vitamin C, which improves reprogramming efficiency when added to culture medium during reprogramming [40,41]. The combination of SR, bFGF, and N2 significantly enhances reprogramming efficiency and kinetics [34]. bFGF is also a crucial factor for the derivation and maintenance of human naïve pluripotent cells (human ESCs and iPSCs) [42]. Therefore, the role of bFGF in mESC differentiation should be reconsidered.
In GO analysis, we found that “stem cell maintenance,” “gamete generation,” and “M phase of meiotic cell cycle” were the most affected genes in XiPS-7 versus XiPS-7c cells. Therefore, we suggest that partially reprogrammed cells could be further reprogrammed to the pluripotent state by upregulation of stem cell maintenance- and gamete generation-related genes (Supplementary Table S4). In KEGG pathway annotation, “focal adhesion” and “ECM-receptor interaction” categorized the most affected genes in XiPS-7 cells. Recently, ECM regulation by bFGF has been found to be crucial for somatic cell reprogramming [39]. We suggest that bFGF could play a role in somatic cell reprogramming through downregulation of extracellular collagens.
In this study, the partially reprogrammed cells did not express core pluripotency genes Oct4 and Sox2, but expressed Nanog. Accordingly, the Nanog promoter region was demethylated, but the Oct4 promoter region was hypermethylated in XiPS-7 cells. These partially reprogrammed cells could maintain the intermediate state for more than 20 passages without expression of Oct4 and Sox2. Oct4 and Sox2 regulate Nanog expression in pluripotent stem cells [43,44]; however, in XiPS-7 cells, Nanog was independent of the Oct4 and Sox2 regulatory network. This phenomenon could be explained by a recent report suggesting that Nanog could be expressed in the inner cell mass of Oct4-knockout embryos [36]. The expression of endogenous Nanog mediates somatic cell reprogramming into the pluripotent state [19], and forced expression of Nanog overcomes the obstacles in somatic cell reprogramming by switching to serum-free medium with LIF, a culture condition that does not support maintenance of mESCs [20]. In this regard, we suggest that Nanog without Oct4 and Sox2 maintains the partially reprogrammed cell state and conversion to fully reprogrammed iPSCs from partially reprogrammed cells by switching from mESC culture conditions to SR-based media with bFGF. The long-term expandable partially reprogrammed cells may provide useful clues to understanding stepwise reprogramming.
Footnotes
Acknowledgments
This work was supported by the Biomedical Technology Development Program and the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science, and Technology (grant no. 20110019489 and 2013R1A1A2011394).
Author Disclosure Statement
The authors have no conflicting financial interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.