
Abstract
During storage and transportation of collected cord blood units (CBUs) to the bank prior to their processing and cryopreservation, it is imperative to preserve the functional capacities of a relatively small amount of cells of interest (stem and progenitor cells) which are critical for graft potency. To improve CBU storage efficiency, we conceived an approach based on the following two principles: (1) to provide a better nutritive and biochemical environment to stem and progenitor cells in CB and (2) to prevent the hyperoxygenation of these cells transferred from a low- (1.1%–4% O2 in the CB) to a high-oxygen (20%–21% O2 in atmosphere) concentration. Our hypothesis is confirmed by the functional assessment of stem cell (hematopoietic reconstitution capacity in immunodeficient mice—scid repopulating cell assay) and committed progenitor activities (capacity of in vitro colony formation and of ex vivo expansion) after the storage period with our medium (HP02) in gas-impermeable bags. This storage procedure maintains the full functional capacity of a CBU graft for 3 days with respect to day 0. Further, using this procedure, a graft stored 3 days at +4°C exhibits better functional capacities than one currently used in routine storage (CBUs stored at +4°C for 1 day in gas-permeable bags and without medium). We provided the proof of principle of our approach, developed a clinical-scale kit and performed a preclinical assay demonstrating the feasibility and efficiency of our CBU preservation protocol through all steps of preparation (volume reduction, freezing, and thawing).
Introduction
T
The functional preservation of the relatively small amount of cells of interest in collected CBU, during the period of transportation to the bank and until its processing and cryopreservation, is very important for the graft potency. Nowadays the general practice is to process the CBU within 1–2 days (depending on the country) after the collection, thus limiting the CBU banking to maternity wards located near the CB bank. These time lapses were justified mainly on the basis of the analysis of total and CD34+ viable cell content in CBU without exploring their capacity of the in vivo engraftment [8 –10]. Recently, we investigated the total and CD34+ cell number as well as the number of functional committed progenitors (colony-forming cells [CFCs]) 3 days after collection [11] and found that the storage of CBU at +4°C significantly decreased all three cell categories, specially CFCs. Consistent with a recent study [12], these results suggest that delaying cryopreservation should be minimized to avoid the significant losses in cell potency.
To improve the storage efficiency, that is, functional preservation of stem and committed progenitor cells in CBU, we conceived a novel approach based on two principles: (1) to provide a better nutritive and biochemical environment to stem and progenitor cells and (2) to prevent the hyper-oxygenation of these cells transferred from an environment with a low- (1.1%–4% O2 in the CB) [13] to a high-oxygen (20%–21% O2 in atmospheric air) concentration, that is, respecting the “oxygen stem cell paradigm” [14]. In this study, by employing a functional in vivo repopulation assay to detect hematopoietic stem cell activity (scid repopulating cells [SRCs]) and in vitro colony-forming assay to detect hematopoietic committed progenitors, we demonstrated not only the proof of principle for this approach, but also developed a clinical-scale kit and performed a preclinical assay demonstrating the feasibility and efficiency of our CBU preservation protocol.
Materials and Methods
Cord blood units
For small-volume experiments (Fig. 1A), CBUs rejected for technical, logistical, and organizational reasons were used within 1 day after collection. The large-volume experiments were performed on normal-volume CBUs (adding the medium in the laboratory or in maternity ward within 6 h after collection) (Fig. 1B, C). These CBUs were contraindicated for banking (medical reasons, such as early placental rupture, meconium in amniotic liquid, genetic factors, etc., or mother's absence for late serological and other tests mandatory for banking). In all cases, informed consents (ie, consenting the use of CB for scientific research) were obtained.

Experimental approaches.
Medium
Medium HP02, a clinical-grade product closely related to the medium HP01 (HP01©Macopharma) conceived for the ex vivo expansion cultures [15 –18], was used in these experiments. The exact formula is an industrial secret.
Collection/storage bags
For small-volume experiments 30-mL polyvinyl chloride (PVC) (Fig. 2A) or ethylene vinyl acetate/ethylene vinyl alcohol/ethylene vinyl alcohol (EEE) bags were used. For clinical-scale (clinical volume) experiments, three different sets of devices were used: (1) the control group consisted in using the standard PVC bags (MSC1202PU; Macopharma) for CB collection, (2) the test groups consisted in the standard PVC bag (MSC1202PU) used in association with an EEE bag (0300020 and 0300030; Macopharma) containing (or not) 50 mL of HP02 medium (Fig. 1B) and (3) a newly designed collection kit that included the collection and conservation kits in a single device and contained 50 mL of HP02 medium (Figs. 1C and 2B). A standard volume (21 mL +8 mL) of anticoagulant (citrate phosphate dextrose) was present in each collection bag and satellite bag, respectively.

CB was collected according to the routine protocol in place within the maternity ward and the HP02 medium was added to the CBU up to 6 h after the collection. The diluted CBUs were subsequently stored in refrigerator at 4°C for 2 or 3 days.
Cell counting
The number of total and mononucleated cells was counted by automatic cell counter (Cell Dyn 3000; Abbott). The viable CD45+/CD34+ cells were analyzed and counted directly by using the TruCount microbead three-color fluorescence kit (7AAD exclusion; Becton Dickinson) by flow cytometer (FACSCalibur; Becton Dickinson).
Apoptosis/viability determination
In some experiments (ex vivo expansion of CD34+ cells from stored CBU) the combined test of viability/apoptosis was performed as previously described [19,20] by flow cytometry on the basis of propidium iodide fixation and Annexin V (AnnV) expression.
CFC assay
To detect the committed hematopoietic progenitors, the cytokine-supplemented methylcellulose medium Stem Alpha ID (Stem Alpha SA) was used. The CB containing 150×103 nucleated cells (with addition of a medium volume to reach 0.5 mL) was added to 2.5 mL of Stem Alpha ID medium and mixed, and 1 mL of suspension (50×103 cells) was plated in 35-mm Petri dishes in duplicate (Nunc). After 14 days of incubation at 37°C in water-saturated atmosphere, 20% O2 and 5% CO2, the CFCs (colony-forming unit–granulocyte macrophage [CFU-GM]+burst-forming unit–erythroid [BFU-E]+colony-forming unit–mix [CFU-Mix]) (>50 cells) were counted using an inverted microscope.
SRC assay
Stem cell activity was evaluated by the in vivo assay—transplantation of human cells to immunodeficient NOD-scid IL2rγnull (NSG) mice. The day-0 (D0) or day-3 (D3) CB cells were injected to 7–12-week-old NSG mice (central animal-keeping facility of University of Bordeaux Segalen) conditioned by intraperitoneal injections of 25 mg/kg Busulfan (Busilvex®; Pierre Fabre) at day minus 2 (D−2) and day minus 1 (D−1) before the cell injection time-point (according to the previously established protocol) [21]. In all experiments 1×103 D0 CD34+ cells were injected per mouse as well as the actual number of CD34+ cells in the same CBU fraction (ie, fraction equivalent) after storage, volume reducing, and thawing. In fact at D0 and after each step (storage, volume reduction, and freezing/thawing) CD34+ cells were isolated from total CBU sample and resuspended in the same volume of medium. Then, the same volume fraction as D0 one was injected to each mouse. In this manner, directly and without inducing any calculation error, we studied the impact of each step of procedure on SRC activity (Fig. 2C). Thus, the direct comparison of total SRC activity per CBU was possible. Positive control (mice injected with 20–50×103 CD34+ purified at D0) and negative control (noninjected mice) were included in each experiment (four independent experiments, 8–12 mice/condition/experiment). After 7–8 weeks, the animals were euthanized, their femora were isolated, and the bone marrow was flushed with 1 mL of RPMI 1640 complemented with human serum albumin (HSA; 4% final concentration). Cells were incubated with an fluorescein isothiocyanate-coupled anti-human CD45, phycoerythrin-coupled anti-human CD19, and allophycocyanin-coupled anti-human CD33 antibodies for 20 min at +4°C in the dark.
Washed cells were analyzed on an FACSCalibur cytometer (Becton Dickinson) to detect and quantify human chimerism. This approach, detecting clusters of differentiation (CD) mostly expressed on differentiated hematopoietic, myeloid, and lymphoid cells (CD45, CD33, and CD19, respectively) provides information related to a relatively more mature SRC subpopulation.
To detect the committed progenitors (ie, CFCs) of human origin in murine bone marrow, 30 μL of femoral cell suspension was seeded per 250 μL of methylcellulose (in duplicate) (Stem Alpha-1; Stem Alpha SA) supplemented with 10% human fresh frozen plasma AB (PFC; Aquitaine-Limousin Branch of French Blood Institute) previously sterilized by filtration, 25 ng/mL interleukin-3 (IL-3), 25 ng/mL granulocyte/macrophage colony stimulating factor (GM-CSF), 50 ng/mL stem cell factor (SCF) (all three human recombinant proteins; PeproTech), and 3 U/mL erythropoietin (human recombinant protein; Tebu-Bio) [17]. After 10 days of incubation, the human CFCs (>30 cells) were counted using an inverted microscope. This approach, detecting the committed progenitors (CFCs) in the murine bone marrow after transplantation, provides information related to the relatively less-mature subpopulation of SRCs.
CB volume reduction
Before freezing, the nondiluted (“routine”) CBUs stored for 1 day or the HP02-medium-diluted CBUs stored for 2 days (in some cases) were volume reduced by Sepax (Biosafe) (Fig. 1B, C). This step, performed following the routine procedure [22], results in the elimination of the majority of plasma and red blood cells (RBCs) and reduces the volume (initial CBU volume was 120.8±20.0 mL [range 89.0–173.8 mL]; n=58) to 20 mL of mononuclear cell (MNC)–enriched fraction.
Cryopreservation (freezing/storage/thawing) procedure
The volume-reduced CBU was treated with 5 mL of dimethyl sulfoxide/dextran 40 (55%; 10% final concentration) (Origen Biomedical) with Coolmix device (Biosafe), placed in plastic envelope and in the metal container. These samples were then frozen by automatic control-rate freezer/storage device (Bioarchive; Thermogénésis Corp.). This device allows storing volume-reduced CBUs at −196°C in liquid nitrogen. After at least 10 days of storage, the CBUs were thawed and their functional capacity (number of viable total, mononucleated, and CD34+ cells, CFCs and SRCs) was examined. As recently stated [18], we adopted this pragmatic solution—short period of storage in frozen state on the basis of historical data from our CB bank (n=324) showing no difference in cell loss between short-term (several days) and long-term (up to 12 years) cryopreservation periods. The thawing was performed per routine protocol (immersion of CBU into water bath, 37°C). The cells were then transferred to the tubes (50 mL Falcon, Ref: 734-0448; BD Biosciences) and washed three times in HSA 4% (LFB)/ACD 10% (Acid Citric-Dextrose type A, Ref: 089524; Bioluz) before being counted and tested.
Ex vivo expansion of stored CBU CD34+ cells
After routine-storage (D0/PVC), or D2 PVC/HP02 or D3 EEE/HP02 storage, the CD34+ cells were selected using “indirect” CD34+ selection kit (Ref: 130-046-701; Myltenyi Biotech). Purified CD34+ cells were then seeded in expansion cultures (D12 or D14 liquid cultures in medium Macopharma HP01 in the presence of 100 ng/mL SCF, 100 ng/mL fms-related tyrosine kinase 3-ligand [both human recombinant proteins; PeproTech], 10 ng/mL G-CSF [Neupogen®; Amgen], and either 20 ng/mL thrombopoietin [TPO; PeproTech] for D12 cultures [6] or 100 ng/mL megakaryocyte derived growth factor instead of TPO [Amgen] for D14 cultures [16]).
Statistical analysis
To compare experimental conditions the nonparametric paired Wilcoxon test, nonpaired Mann–Whitney test, or t-test for paired samples were used as mentioned in the “Results” section.
Results
Medium “HP02” improves maintenance of functional hematopoietic progenitors (CFCs) in the CB stored at +4°C
If the CB (CBUs arrived in the laboratory <1 day after collection) was stored in miniaturized bags (30 mL) made of the same material as classical routine CB bags (PVC) (Fig. 2A), the addition of HP02 medium (1:1; 10 mL +10 mL) (Fig. 1A) significantly improved the maintenance of CFCs after 3, 8, or 14 days of storage (Fig. 3A). Under these conditions, the addition of HP02 enabled a preservation of ∼80% of these progenitors after 8 days at +4°C. We tested the capacity of CD34+ cells after 2 days of storage with or without medium HP02 to generate ex vivo cultures (our 14-day expansion culture technique [16]), CD34+ cells, and committed progenitors (CFCs). While the expansion of CD34+ cells was similar after 2 days to the values of starting point of experiment, the storage without HP02 medium was deleterious for the cells producing the CFCs in the expansion culture (Fig. 3B). On the contrary, if HP02 medium was present, the capacity of CFC generation in ex vivo cultures was significantly higher and similar to the starting point D0. Thus, these results imply the significance of HP02 medium for the conservation of hematopoietic progenitors inside CB stored at +4°C.
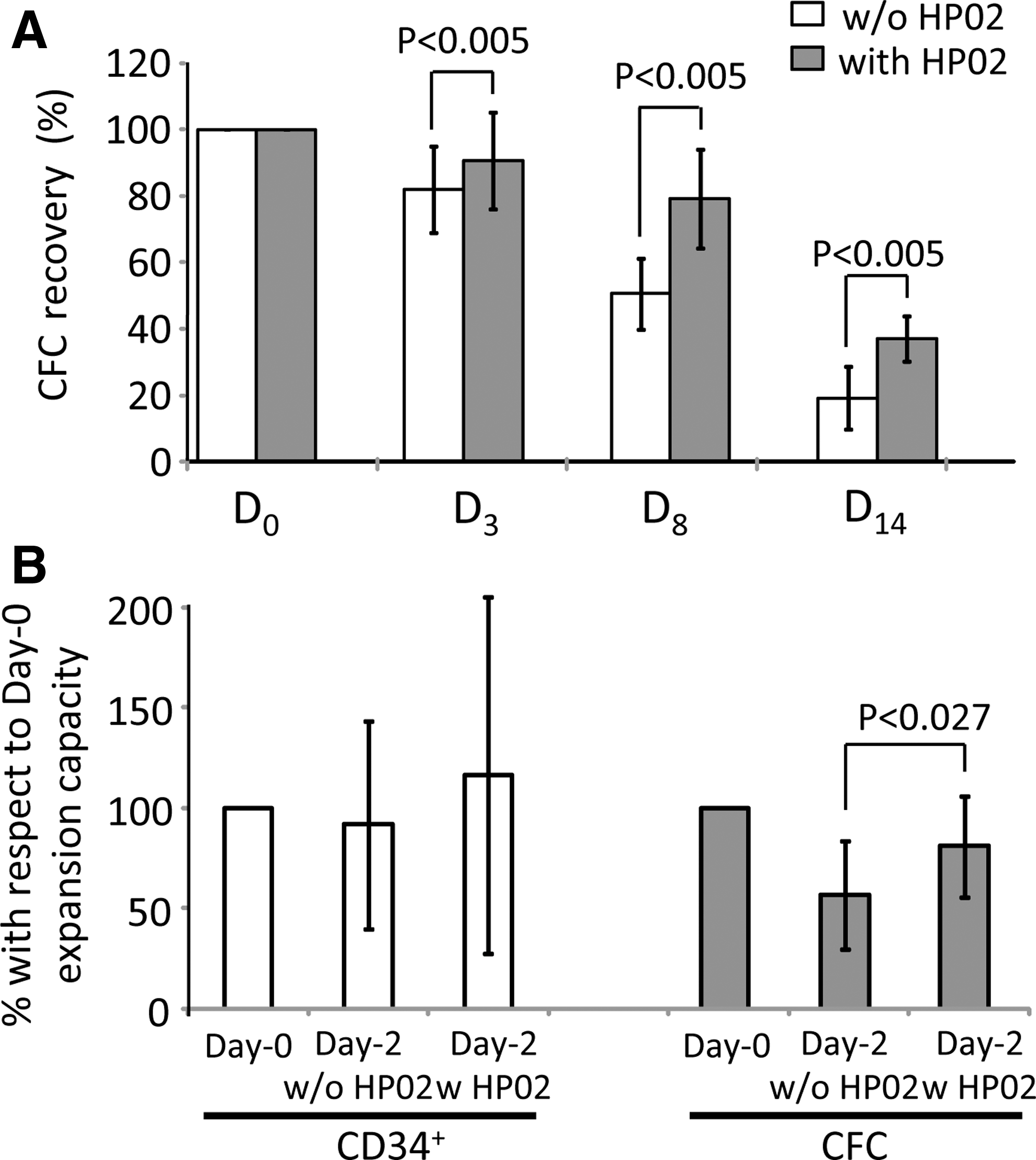
The effects of HP02 medium on the maintenance of functional hematopoietic progenitors in CB stored at +4°C.
Beneficial effect on committed progenitor's maintenance was not induced by a simple dilution of CB but by the properties of HP02 medium, preventing oxygenation/CO2 leak (EEE bags) enhances this effect
Next, we tested a lower proportion medium/CB (1:2, 4 mL of HP02 medium +8 mL of CB) (miniature bags, Fig. 1A). After confirming the beneficial effect, this proportion was judged more interesting for clinical-scale development since it corresponded to medium volume (50 mL) that could be added to real-scale CBU volumes. In fact it fits perfectly with most of CBUs (in range 90–110 mL) and could be added to rare high-volume CBUs (up to 180 mL) since the capacity of bags is up to 300 mL. In this experimental cohort, in addition to testing our hypothesis that the simple dilution was not the reason for the beneficial effect (described in Fig. 3), we wanted to evaluate whether there was a positive effect in preventing CB oxygenation. In that respect, we added an experimental condition (NaCl instead of HP02) and stored the samples in all three different conditions (CB, CB/HP02, and CB/NaCl) in PVC and EEE bags for 3 days at +4°C (Fig. 4A). These results clearly show (1) an advantage of HP02 medium with respect to NaCl (which does not have any effect) and (2) that a positive effect of EEE bags is visible in this experimental series, both in CB and CB/HP02 conditions.

Specific positive effect of HP02 medium and EEE bags (preventing oxygenation and CO2 leak) on the maintenance of committed progenitors (CFCs) in the course of CB storage at +4°C.
To verify these results in clinical-scale conditions, we stored CB in standard routine PVC or gas-impermeable (EEE) bags of the same size for 3 days. We observed a trend of improved maintenance of CD34+ cells in EEE bags (Fig. 4B). We verified that EEE bags were not gas permeable since dark red blood color (similar to vein blood) was maintained for 3 days while in the standard gas-permeable bag the blood was oxygenated (bright red color similar to arterial blood) (Fig. 4C). Thus, in this experimental context, the impact of preventing the oxygenation (EEE bags) was not very pronounced. Next, we compared the efficiency of CB cell preservation in clinical-scale EEE bags with or without HP02 medium (Fig. 4D). A significantly better preservation of CFCs was obtained with HP02 medium.
Thus, each EEE bag and HP02 medium positively impact CFC and/or viable CD34+ cell maintenance during 3 days of storage at +4°C. These findings allowed us to test our hypothesis by direct comparison of the actual standard routine condition (PVC bags/CB without medium) to the condition that should cumulate the advantages of better nutritive environment and the attenuation of hyper-oxygenation (EEE bags/HP02 medium), in the real-size situation and 3 days of storage. Indeed, the combination of EEE bag with HP02 medium enabled the total preservation of start-point number of viable CD34+ cells and excellent (∼93%) preservation of committed progenitors, which is, in both cases, significantly improved with respect to the PVC bags/CB without medium condition (∼82% vs. ∼77%, respectively) (Fig. 4E).
Positive impact of HP02 medium and EEE bags is maintained after volume reduction (Sepax) and cryopreservation (freezing/thawing procedure): assay using a clinical-scale kit
The HP02 medium (50 mL) was added to the bags with CB that arrived within 6 h after collection to our CB bank. The mixture CB/medium was then stored for 3 days at +4°C and then processed (Sepax volume reducing) and cryopreserved. As controls, the EEE bags with CB without medium were processed and stored in the same way for 3 days. After at least 2 weeks, the CB samples were thawed. At each time point, the recovery of total cells, CD34+ cells, and CFCs was assayed. In the first series of real-scale experiments, the beneficial effect of our procedure with respect to control bags was maintained both after volume-reduction and freezing/thawing procedures. This effect was especially pronounced for CFCs (Fig. 5A).
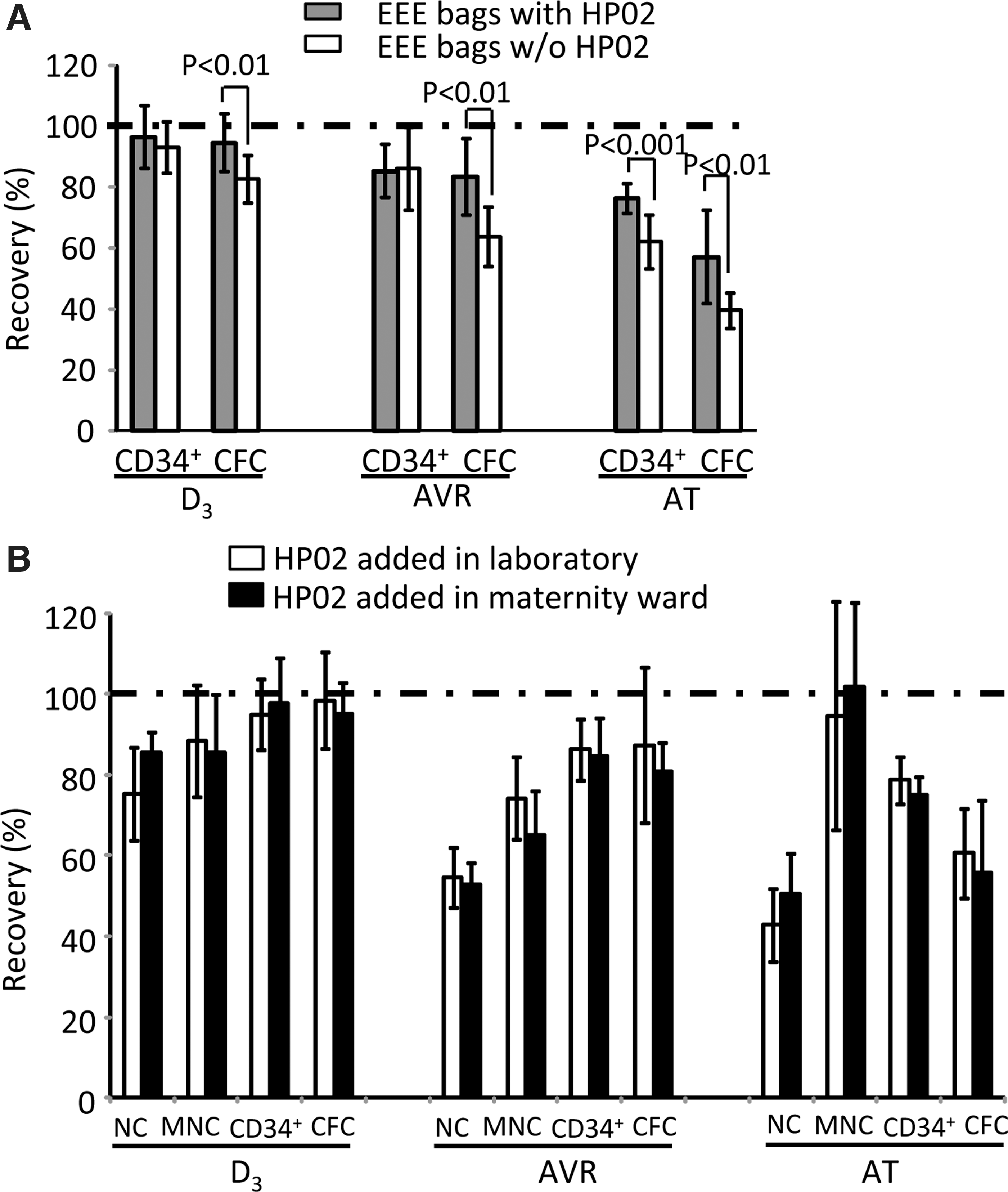
Improved maintenance of hematopoietic progenitors after storage with HP02 medium is preserved after volume reduction and cryopreservation.
To test our procedure in clinical-like collection of CB, a clinical-scale kit was conceived (Fig. 2B). It consists of an EEE bag containing HP02 medium (50 mL) connected with the collection tube, and another EEE bag for CB collection and storage. After collection of CB, medium (50 mL) was transferred to the collection bag (containing the anticoagulant solution) through the collection tube (washing the residual CB from tube into collection bag). The tube was then thermo-sealed and the needles were removed. The samples were then mixed, kept for 1 h at room temperature, and then transferred at +4°C to be transported and stored for 3 days in our CB bank, before being processed and cryopreserved as explained previously. The recovery of total cells, MNCs, CD34+ cells, and CFCs obtained in this “clinical-like collection” was compared with recoveries obtained by the routine (PVC) bags arriving within 6 h after collection in CB bank and then exposed to HP02 medium in EEE bags (Fig. 5B) (the cohort “white bars” from Fig. 5A presented for comparison in Fig. 5B). No significant differences were noted between these two series, confirming the feasibility of this procedure under real clinical conditions.
EEE/HP02 conservation procedure enables, after 3 days, the same recovery of viable CD34+ cells and their ex vivo expansion capacity as the actual standard routine procedure after 1 day
Since CFC number is not regularly determined in CBUs at the reception, before and after cryopreservation, the only parameters that we were able to compare with the routine CB series in our bank were total and CD34+ cells. After volume reduction (before cryopreservation), the recovery after 3-day storage with EEE/HP02 procedure was similar to that found under routine conditions after 1 day (Fig. 6A). At the same time, the recovery of total cells was significantly (P<0.001) lower (25%) after 3 days with EEE/HP02 procedure than after 1 day of routine CB bank storage. When we analyzed the daily kinetics of viable MNCs and polymorphonuclear cells (PNCs) in these conditions, it was evident that, until D4, at +4°C, the number of MNCs remained stable, while PNCs rapidly declined (Fig. 6B). Nevertheless, if we compare the recovery of total viable nucleated cells and CD34+ cells, after thawing, it turns out that our procedure enables quite similar recovery of these two cell populations for 3 days as those obtained after 1 day in actual routine banking of CB (Fig. 6C). If the CD34+ cells selected from CBUs were stored in routine storage conditions (D0 PVC bags w/o HP02) or for 3 days in EEE/HP02 condition, their capacity to produce the new CD34+ cells of committed progenitors (CFCs) was comparable (Fig. 6D) (∼80-fold expansion in both cases). Further, the CD34+ cells from CBUs stored for 3 days under EEE/HP02 condition produce significantly less apoptotic cells (AnnV+ IP+) in 12 days of culture than their counterparts from CBUs stored for 1 day under routine conditions (Fig. 6E).
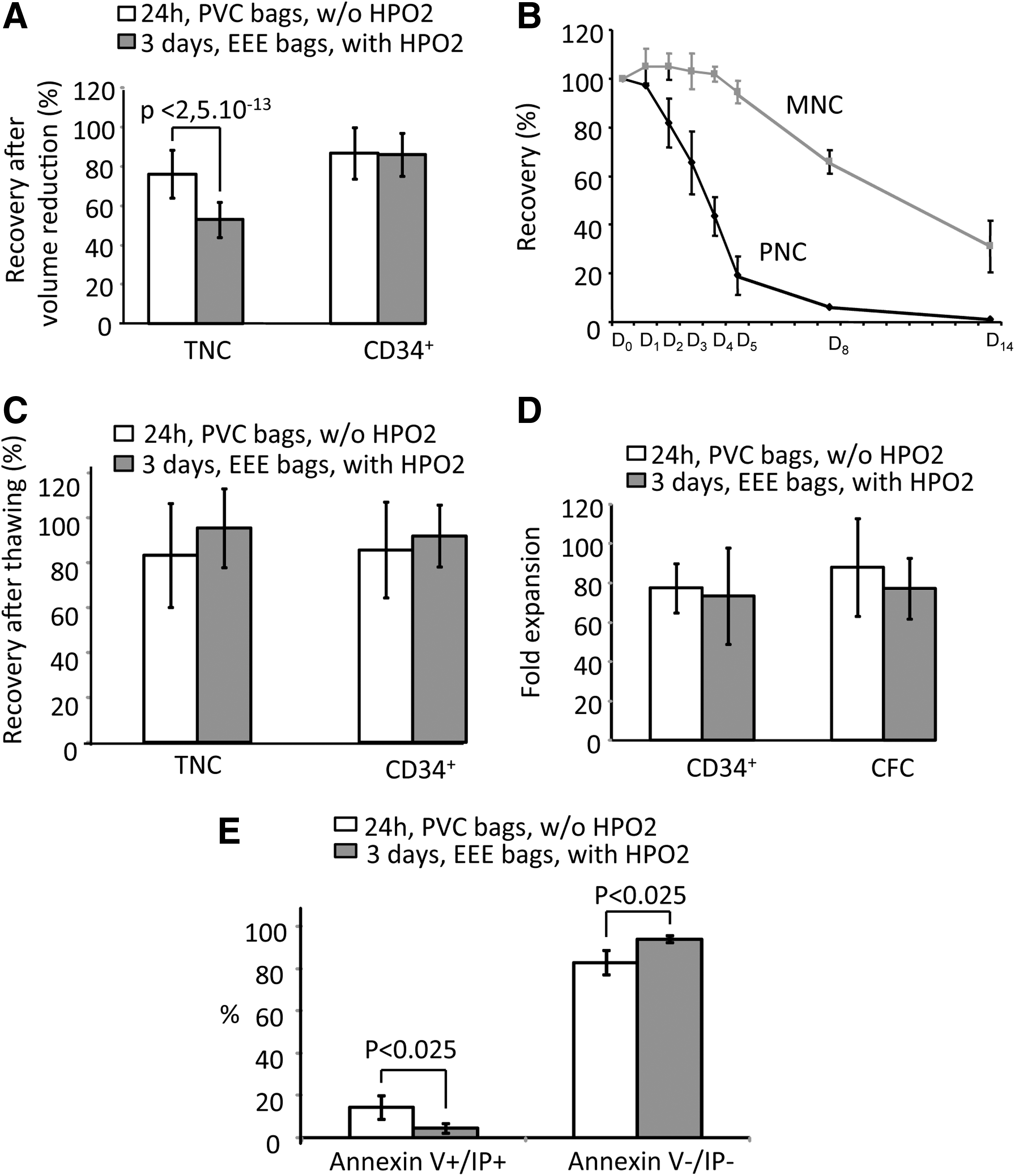
EEE/HP02 CB D3 storage compared with the routine D1 CB storage: the same recovery of CD34+ cells and their comparable ex vivo expansion capacity.
EEE/HP02 conservation procedure enables, after 3 days, the D0 level of stem cell activity (SRCs), which is superior to the actual standard routine procedure after 1 day
To get insight into the stem cell maintenance in the course of CB storage, we employed the SRC assay using the NSG mice [6,17,18]. When the human-specific CD45+ cells were analyzed in the bone marrow of NSG mice 7 weeks after injection and similar proportions of positive mice (90.04% vs. 96.5% vs. 94.1%) were detected in D0 CB sample (<6 h), routinely stored CB sample (D1/PVC w/o HP02), and CB stored for 3 days in EEE bags with HP02, respectively, suggesting that the number of SRCs detected this way remains stable until D3 in EEE/HP02 condition (Fig. 7A). On the contrary, the median SRC proliferative capacity (deduced on the basis of the median CD45+-specific human chimerism) declined after routine D1 storage (PVC w/o HP02) (1% vs. 3.2% at D0) but not after D3 EEE/HP02 storage (4.2%) (Fig. 7A). This advantage of EEE/HP02 storage was maintained after volume reduction, but is not statistically significant after freezing/thawing of cells. The capacity of differentiation neither changed after D1 routine storage nor after D3 EEE/HP02 storage as estimated on the basis of the respective proportions of CD33+ and CD19+ cells within CD45+ cell population of human origin (Fig. 7B). Similar results were obtained after volume reduction (“AVR”) by Sepax or after freezing/storage/thawing procedure (“AT”) (Fig. 7A, B). The analysis of committed progenitors derived for 7 weeks from human stem cells engrafted in mouse bone marrow provided insight into an SRC subpopulation relatively more primitive than one estimated on the basis of CD45+ cells. The mean absolute number of human-origin CFCs per femur (Fig. 7C) was twice higher after D3 EEE/HP02 storage than after D1 routine storage in PVC bags without HP02. Also, the proportion of mice negative for human CFCs increased fourfold (from 10% to 40%) after D1 routine storage but was similar to D0 values after D3 EEE/HP02 storage (Fig. 7D). The better conservation of SRCs estimated on the basis of human CFCs in mouse bone marrow was maintained after volume reduction and freezing/storage/thawing procedure (Fig. 7C, D). While the CFC number per femur was significantly decreased after freezing/thawing of D1 standard storage CBUs, it was not significantly different with respect to D0 after D3 EEE/HP02 storage (Fig. 7C). Also, the percentage of mice “positive” for human-origin CFCs was similar after D3 EEE/HP02 to D0, while it was lower after D0 routine storage (Fig. 7D). Collectively, these results demonstrate that the EEE/HP02 storage procedure enables the maintenance of SRCs (our information is related to two distinct subpopulations of SRCs) for 3 days at the D0 level, without impacting their differentiation capacities.

D3 EEE/HP02 storage enables full maintenance of D0 scid repopulating cell potential unlike D1 storage in routine conditions.
Discussion
The results presented in this study clearly demonstrate that our approach that combines the prevention of oxygenation/CO2 leak and optimizes nutritive environment of storage medium increases the preservation of hematopoietic stem cells and committed progenitors in the CB. These conclusions are based on the assessment of stem cell activity and progenitors by the functional approaches: (1) capacity of hematopoietic reconstitution in immunodeficient mice, (2) capacity of ex vivo expansion after storage period, and (3) progenitors' capacity of in vitro colony formation. Application of this storage procedure can maintain a fully functional capacity of CBU graft for 3 days with respect to D0; that is, it yields a graft with better functional capacities over the one nowadays stored for 1 day in gas-permeable bags and without medium.
With regards to the oxygenation and CO2, our approach is based on the “oxygen stem cell paradigm” [14] and the experimental data confirming this concept in hypothermia [23,24]. In fact, combination of low O2 and high CO2 concentration atmosphere successfully maintains very good functional capacities of CD34+ cells mobilized in peripheral blood for 7 days and for those from CB for 10 days [23,24].
Having in mind that the goal of CB storage is not to maintain the RBCs, but hematopoietic progenitors and stem cells, we decided to change completely the actual approach (gas-permeable bags) inherited from blood transfusion practice, aimed to RBC preservation. Thus, in addition to bags preventing oxygenation and CO2 leak from CB, we used a medium not only optimized from nutritional point of view for hematopoietic stem and progenitor cells, but also supplied with a battery of antioxidants, same as in the culture medium HP01 (Macopharma) [6,15 –18]. This strategy resulted in a synergistic effect on preservation of these functional cell populations during the storage period. Further, this advantage is conserved after volume reduction even if the actual centrifugation speed of Sepax device is not adapted to a high-volume/low-hematocrit CBU as one diluted by HP02 medium. Our preliminary data suggest a possibility to further improve the yield of cells after volume reduction applying an adapted centrifugation speed (data not shown, work in progress). Finally, the positive impact of HP02 storage with respect to standard D1 storage is still present after cryopreservation (freezing/storage at −196°C/thawing) for CD34+ cells, CFCs, and SRCs determined on the basis of human-origin CFCs in mouse bone marrow. The only functional stem cell population that seems not to have a better recovery (lack of statistical significance) is SRCs, determined on the basis of human-origin CD45+ cells in mouse bone marrow (relatively less primitive with respect to one determined on the basis of CFCs). It should be stressed that freezing/thawing technique could probably be improved as we recently demonstrated for ex-vivo-expanded CB stem and progenitor cells [6].
Consistent with recent publications [17,18,25], the present study reveals a discrepancy between functional (in vivo and in vitro assays) and viable CD34+ and MNC counting. From the point of view of experimental hematology, this phenomenon is perfectly understandable, having in mind the functional heterogeneity of CD34+ cell population in which only a minority are hematopoietic stem cells and about a half are committed progenitors. In addition, the viability determination at only one time point could be irrelevant for real estimation of the functional graft capacity that relies on functional stem and progenitor cell content [18,26]. Due to positive correlation between total cell number and transplant outcome established by analysis of big series [27] (even though the same study stressed that the graft progenitor cell content is a better predictor than nucleated cell quantity), the transplantation practice of CB is still operating with total viable cell number of a CBU before cryopreservation [28,29] as the first parameter of the CBU graft richness and potency. In that respect, a CBU stored for 3 days with EEE/HP02 procedure could be incorrectly considered inferior to D1 routinely stored CBU. This difference arises from the loss of granulocytes, which is in fact a beneficial phenomenon, since the absence of granulocytes during freezing/thawing lowers the risk of side effects, specially clamping. Although in multicenter clinical trials more total cells in graft means, in general, more progenitors and stem cells, there is no justification to take granulocytes into account for individual graft richness since these cells do not participate in hematopoietic reconstitution. Indeed, an accumulating number of data urge us to consider the functional parameters and scientific knowledge in transplantation practice [30]. In the case of EEE/HP02 storage procedure, it is obvious that a lower number of total cells before cryopreservation does not represent a pertinent indicator of graft potency, since (1) the number of total cells after cryopreservation/thawing is comparable after D3 EEE/HP02 storage with that of routine D1 storage, (2) ex vivo expansion capacities in these two storage conditions are also comparable (EEE/HP02 procedure offers even an advantage with respect to the proportion and absolute number of CD34+ cells after expansion culture), and (3) the SRC activity is even better maintained with EEE/HP02 procedure.
A number of studies proposed that a D3 routine storage of CBUs at +4°C is not significantly impacting the number of viable total MNCs and CD34+ cells [9,10]. Whatever the level of arbitrarily acceptable loss of CD34+ cells, it is evident that this point could not be properly evaluated without testing at least two functional categories of hematopoietic cells (inside CD34+ population): total number of committed progenitors (CFCs) and stem cell activity in these CBUs. The only study that explores in vivo reconstitution of immunodeficient mice in that context [25] confirms that +4°C storage has a clear advantage over room temperature. The careful analysis of mice reconstituting data in the same study also reveals that the maintenance of hematopoietic reconstituting capacity was lower after D3 storage with respect to D1 storage in routine conditions at +4°C. In line with these results are our data show a significant decrease in committed progenitors after D3 with respect to D1 routine storage [11,12] (Fig. 5A). Thus, improving stem and progenitor cell maintenance during D3 CBU storage appears as an interesting goal.
The clinical-scale procedure presented here could, if employed in practice, enable a rationalization of CB bank network (to lower the cost per CBU) in the countries with shorter mandatory periods, by extending the period necessary for CBU logistics and transportation (with a possibility to include CBUs from overseas territories usually characterized by particular HLA phenotypes), as well as banking of CBUs collected Friday evening and during the weekends. In particular, our revised clinical procedure, presented here, could improve the functional potency of stored CBUs especially in the countries with longer (3 days) mandatory periods.
Footnotes
Acknowledgments
The authors are thankful to Mrs. Elisabeth Doutreloux-Volkmann (EFS Aquitaine-Limousin, Bordeaux) for reading the article and to Mrs. Svetlana Gavrilov, MD, PhD (Sloan-Kettering Institute, NY), for the language corrections. This project was supported by the regular R&D budget of French Blood Institute, Aquitaine-Limousin Branch and the specific project support of Macopharma. The content of this contribution is related to a patent application that was carried out in France on October 10, 2012, with number 1259684 and extended to PCT international on October 9, 2013, with number PCT/FR2013/057150. A subset of results presented here was the subject of oral presentation at the Second Congress “Autour de la Cellule Souche” Bordeaux, France, March 2013.
Author Disclosure Statement
During the course of this project L.R., B.D., and M.P. were employed by Macopharma (division Biotherapy), a private company; J.C., P.D., B.D., M.V., P.B.G., and Z.I. are employed at French Blood Institute (Etablissement Français du Sang), a national public institution (the blood banks, the allogeneic CB banks, and the cell therapy facilities). This contribution is founded by the research funds of French Blood Institute (Aquitaine-Limousin Branch) and Macopharma Company. V.L. declares no potential conflicts of interest.