
Abstract
Body axes and germ layers evolve at gastrulation, and in mammals are driven by many genes; however, what orchestrates the genetic pathways during gastrulation remains elusive. Previously, we presented evidence that microRNA-17 (miRNA-17) family members, miR-17-5p, miR-20a, miR-93, and miR-106a were differentially expressed in mouse embryos and functioned to control differentiation of the stem cell population. Here, we identify function(s) that these miRNAs have during gastrulation. Fluorescent in situ hybridization miRNA probes reveal that these miRNAs are localized at the mid/posterior primitive streak (ps) in distinct populations of primitive ectoderm, mesendoderm, and mesoderm. Seven different miRNA prediction algorithms are identified in silico bone morphogenic protein receptor 2 (Bmpr2) as a target of these miRNAs. Bmpr2 is a member of the TGFβ pathway and invokes stage-specific changes during gastrulation. Recently, Bmpr2 was shown regulating cytoskeletal dynamics, cell movement, and invasion. Our previous and current data led to a hypothesis by which members of the miR-17 family influence gastrulation by suppressing Bmpr2 expression at the primitive streak. This suppression influences fate decisions of cells by affecting genes downstream of BMPR2 as well as mesoderm invasion through regulation of actin dynamics.
Introduction
P
Although known for almost two decades, miRNA biology, especially during development, is still in its infancy. In fact, how miRNAs regulate mammalian development is constantly being defined. miRNAs have become important factors in regulating cellular homogeneity, as mis-expression of various miRNAs leads to various diseases such as cancer, hypertension, autism, and numerous others (accessible at
The miR-17 family is one of the most conserved families of miRNAs and is thought to have evolved along with vertebrates. It is perhaps best known for its abnormal expression in cancer cells [27,28]. Members of this family are expressed at low levels in mESCs and early embryos, but they increase significantly as gastrulating cells differentiate into mesoderm and mesendoderm [11,12]. One of the targets of this miRNA cluster is the 3′UTR of Stat3, a downstream transcription factor (TF) of the Jak/Stat signaling pathway, which is important in early development [7,29 –31]. Although Stat3 is not necessary for hESC maintenance, it is active in both h- and mESCs and its downregulation is necessary for initiating differentiation, not only in h- and mESCs but also in hematopoietic and neural stem cell systems [32].
Investigations into miRNAs usually begin with an initial in silico search for potential targets. Alternatively, if the target(s) is already known, then it is possible to search in silico to find potential miRNAs that bind to that particular target [11,33,34]. miRWalk (
In a number of elegant studies, it was established that ligands such as BMP2 and BMP4 bind BMPR2 in the presence of a type 1 receptor, resulting in phosphorylation and activation of SMAD1/5, which then associate with SMAD4. This complex then enters the nucleus and binds to specific Bmp ligand-Smad-specific DNA sequences [41
–44]. Two significant developmental genes directly regulated by these Smads are the inhibitor of differentiations genes 1 and 3 (Id1/3), which repress differentiation in stem cells when expressed. Another is the gap junction protein Connexin 43 (CX43), an adhesion/communication protein important for proper embryonic tissue development [45
–47]. A fourth gene that is indirectly downstream of these Smads is the cell–cell adhesion protein E-cadherin. In a seminal study by Perez-Moreno et al. [48], the bHLH transcriptional repressor E12/E47 was clearly shown to act as a suppressor of E-cadherin expression by binding to E-boxes within the E-cadherin promoter [48]. The E12/E47 repressor is negatively regulated by their dimerization with ID proteins, in which case ID1/3 when expressed binds E12/E47, leading to its repression, ultimately resulting in E-cadherin expression [49]. Thus, evidence suggests that spatiotemporal regulation of Smad signaling at gastrulation by miRNAs is clearly important and if disrupted, it leads to abnormal downstream regulation of Bmpr2-specific genes such as Id1/3, Cx43, and E-cadherin. Our hypothesis is that miR-17-5p, -20a, -93, and/or -106a are regulating much of this process. To test this hypothesis, we employed miRNA inhibitors and mimics transfecting them into mESCs (our model system) for development, and a semi-in vivo system using mouse pregastrulation-stage embryos. Interestingly, we found that our original hypothesis was only partially correct. miR-17-5p, -20a, -93, and -106a were found to regulate not only Bmpr2 and its downstream genes, but also a novel actin dynamics pathway controlled by BMPR2 association with
Materials and Methods
Cell culture
E14 ESCs (a kind gift from Dr. Xing Dai University California-Irvine and Dr. William Skarnes, Sanger Institute, Cambridge, UK) were grown in feeder-free conditions with media containing 15% ESC qualified serum replacement (Invitrogen). Differentiation of ESCs was induced by removal of leukemia inhibitory factor (LIF; EMD Millipore Corp.) as followed in Foshay and Gallicano [11].
Embryo procurement
Implanted embryos were obtained as described [88 –92]. Briefly, one female is placed in a cage with one male to allow copulation. Females were checked the next day for vaginal plugs, which represents 0.5 days postcoitum. Implanted embryos were obtained from fertilized dames posthumously at 6.0 days postcoitum by dissection from decidua using Dumont No. 5 forceps. Isolated embryos were then placed in 4.0% paraformaldehyde and processed for either in situ hybridization or immunofluorescence.
For overnight incubation, embryos were dissected from their decidua using Dumont No. 5 forceps and all membranes were removed (Fig. 7D). Embryos were cut at their amnion/ectoplacental cone region to enable inhibitors or mock miRNAs to enter and bathe in the primitive ectoderm. All synthetic miRNAs were prepared in Qiagen's hyperfect transfection reagent before incubation with embryos. Embryos were incubated in 24-well dishes in an incubator set for 5% CO2 at 37°C on a slow rocker overnight in Dulbecco's modified Eagle's medium (Thermo Fisher Scientific Inc.) medium supplemented with 15% serum replacement and 1× pen-strep. All experiments involving animals were approved by the Georgetown University Animal Care and Use Committee under a defined protocol.
Fluorescent in situ hybridization analysis
In situ hybridization probes were obtained from Exiqon, and experiments were performed according to the GEISHA protocol (from the University of Arizona) available on the Exiqon Web site. All reagents and apparatus used were diethylpyrocarbonate treated. Hybridizations were conducted overnight at optimum temperatures between 56°C and 58°C.
miRNA inhibitors, mimics, and mocks
miRNA inhibitors and mimics for mmu-miR 17-5p, mmu-miR 20a, mmu-miR 93, and mmu-miR 106a were purchased from Thermo Scientific Molecular Biology (formerly Dharmacon), in addition to mock miRNA controls (Caenorhabditis elegans miRNAs with no targets present in mice). All miR-17 family mimics and inhibitors were transfected together, with each individual miRNA at a concentration of 100 or 200 nM for a total effective miRNA concentration of 400 or 800 nM. All experiments were done using HiPerFect transfection protocol, transfecting into a cell culture with the HiPerFect transfection reagent (Qiagen) and the appropriate miRNA concentration. After the 6 h transfection period, cell media was changed, LIF was removed, and cells were allowed to differentiate at 37°C with 5% CO2. Cells were considered differentiated for one day 24 h after LIF was removed.
Luciferase assay
A 3′UTR dual luciferase reporter construct with the Bmpr2 UTR was purchased from GeneCopeia. This construct contains the 3′UTR of Bmpr2 linked to the firefly luciferase gene under the control of the SV40 promoter. In the same construct, Renilla luciferase is under the control of the CMV promoter and contains a 3′UTR that does not contain any miRNA target sites. The Renilla luciferase serves as an internal control to measure against the firefly luciferase levels. The reporter plasmid was transfected into HeLa cells with Lipofectin at 4 h before inhibitors, mimics, or mock miRNAs were added using HiPerFect. Cells were allowed to grow for 2 days before being harvested for analysis. Luciferase levels were assessed using the Luc-Pair™ miR Luciferase Assay Kit (GeneCopeia) on a luminometer.
Quantitative reverse transcription-polymerase chain reaction
Cells were cultured as described earlier before being freshly harvested for RNA. RNA was extracted from cell cultures using Qiagen's miRNeasy Mini kit (Qiagen Inc.). DNase1 digest was performed to remove contamination (Invitrogen). RNA was quantified and diluted to equalize total content between samples (2 μg/μL). Samples were run using the QuantiTect one-step SYBR green quantitative reverse transcription-polymerase chain reaction (qRT-PCR) master mix kit (Qiagen, Inc.) on an Ependorff RealPlex Multicycler using cycling as given by the kit. Annealing temperature was adjusted per primer pair for optimum reactions. Primers used were Id1 forward: CCCTGAACGGCGAGATCAGT reverse: AGTGTCTTTCCCAGAGATCCCC; Id3 forward: CTGAGCTCACTCCGGAACTT reverse: TCAGGGTAAGTGAAGAGGGCT; Cx43 forward: GAGTTCCACCACTTTGGCGT reverse: AATGAAGAGCACCGACAGCC. All PCR products span an exon:exon junction. All calculations were done using standard qRT-PCR calculations [51]. For Supplementary Fig. S2 (Supplementary Data are available online at
Western analysis
Western blots were performed using precast 7.5% Tris-HCL polyacrylimide gels and run on the Bio-Rad mini gel system. Proteins were transferred to polyvinylidene fluoride membranes using a Bio-Rad Trans-Blot Turbo transfer system, followed by blocking in a solution of 2% bovine serum albumin (BSA) and 5% milk in phosphate buffered saline-tween 20 (PBS-T). Antibodies were purchased from the following places: Abcam phopho-Smad1/5 Cat. No. 9511s; ID3 Cat. No. ab41834. Cell Signaling, E-cadherin Cat. No. 24E10; STAT3 Cat. No. 9132; Bmpr2 Cat. No. 69793; phospho-cofilin Cat. No. 77G2; CX43 Cat. No. 3512s. Santa Cruz E12/E47 Cat. No. sc-349; SMAD1/5 Cat. No. sc-6031; LIMK Cat. No. sc-30696; actin Cat. No. sc1615. BioCheck ID1 Cat. No. 195-14 and used at a 1:15,000 dilution. Bands were visualized using an enhanced chemiluminescence system (Bio-Rad) and exposure to autoradiography film. Similar exposure times were used for each antibody, and actin blots were used to ensure appropriate normalization. Quantification of bands was performed using Adobe Photoshop CS to calculate the pixel intensity of each band. Pixel intensities for each antibody and actin were expressed as a ratio to generate a normalized value for protein expression.
Confocal microscopy
ESCs and embryos were fixed in 4% fresh paraformaldehyde for 30 min followed by permeabilization for 1 h and blocking in PBS-tween plus 2% BSA overnight at 4°C. After blocking, embryos were incubated in primary antibody at a 1:50 dilution for 4 h or overnight at 4°C. After incubation in primary antibodies, specimens were washed for 4 h in blocking solution, followed by incubation with secondary antibodies in blocking for 3 h at room temperature. Antibodies: (same as above) and anti-GTPRac1 (NewEast Biosciences Inc. Catalog Number: 26903), which has been verified to bind to active Rac1 (14 publications on their Web site), were viewed using an Olympus Fluoview 500 Laser Scanning Microscope (Olympus America Inc.) using 1.4 numerical aperture. Images were acquired and analyzed using the accompanying Fluoview software (version 4.3).
Counting stained cells and filopodia
Using Cell Profiler image analysis software
To count filopodia, unlabeled images were given to three separate viewers, who then hand counted filopodia according to (what are we saying the guidelines are for determining a filopodia?). Filipodia numbers on the outer edges of the cells were then divided by the number of cells along the outer edge of a colony. For each colony, each viewer's values were averaged to determine the accurate count of filopodia/out cell of a colony. These data were then graphed to determine statistical significance.
Statistical analysis
Experimental groups were replicated at least thrice. For display of data, each point on a graph represents a mean±standard error of the mean (error bars) for an experimental group or observation. Student's t-test software at (
Results
miRNA-17-5p, -20a, -93, and -106a target BmpR2 in silico
With the advent of the miRWalk Web site (
Since miRNAs are notorious for having multiple targets, we surmised that Stat3 was most likely not the sole target of miR-17-5p, -20a, -93, and -106a. During our search on miRWalk, we found a number of other targets known to be essential for embryogenesis. One target identified by six of seven algorithms was Bmpr2 (Fig. 1A, B), a membrane receptor that is a member of the Tgfβ superfamily. Delving deeper into the TargetScan algorithm (
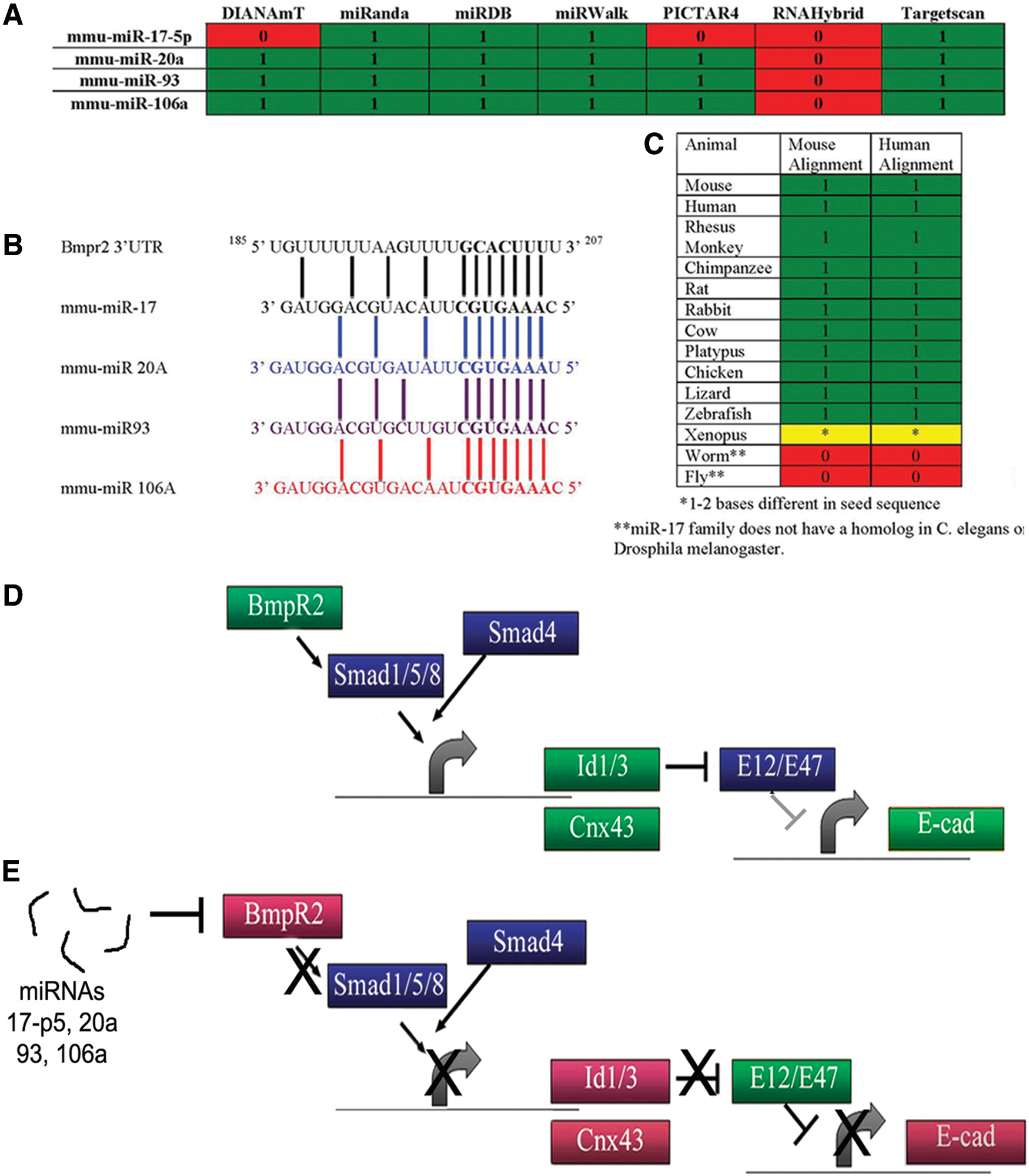
MirWalk.org-generated table predicting Bmpr2 as a target.
Figure 1D shows a schematic of the Bmpr2 pathway compiled from numerous publications [41,42,47,48]. When activated, BMPR2 binds a type 1 receptor, which then phosphorylates SMAD1/5, enabling SMAD4 to bind the complex and enter the nucleus to activate specific genes. Id1/3 genes are directly downstream of BMPR2 as is Cx43. Indirectly, E-cad is also regulated by the Bmpr2 pathways through ID protein regulation/binding to the E-cad repressor E12/E47 [41,42,48,50]. miRNAs 17-5p, 20a, 93, and 106a that target Bmpr2 would also result in downregulation of specific downstream genes (Fig. 1E).
miRNA-17-5p, -20a, -93, and -106a, Bmpr2, and its downstream targets are expressed in a developmentally relevant spaciotemporal pattern in murine embryos
In Foshay and Gallicano [11], our original hypothesis stated that miR-17 family miRNAs were important for ESC differentiation and embryonic development. Evidence supporting that hypothesis included fluorescent in situ hybridization (FISH) data, which identified the expression pattern of specific members of that family in defined regions of the embryo during gastrulation. Figure 2 confirms the miRNA patterns found in Foshay and Gallicano [11]. At embryonic day 7.0 (E7.0), FISH revealed miR-17-5p, -20a, -93, and -106a in both extraembryonic and embryonic regions; however, in embryonic regions, these miRNAs were found exclusively within cells at or near the primitive streak ([11]; Fig. 2A–H). Analyzing FISH-treated embryos at a high magnification revealed a defined pattern of expression (Fig. 2E, F). In embryonic regions, these miRNAs were restricted to the primitive streak, posterior mesoderm, and mesendoderm, while expression in the extraembryonic region was strongest in the extraembryonic visceral endoderm (eve). We found only subtle differences in the expression pattern for all four miRNAs.
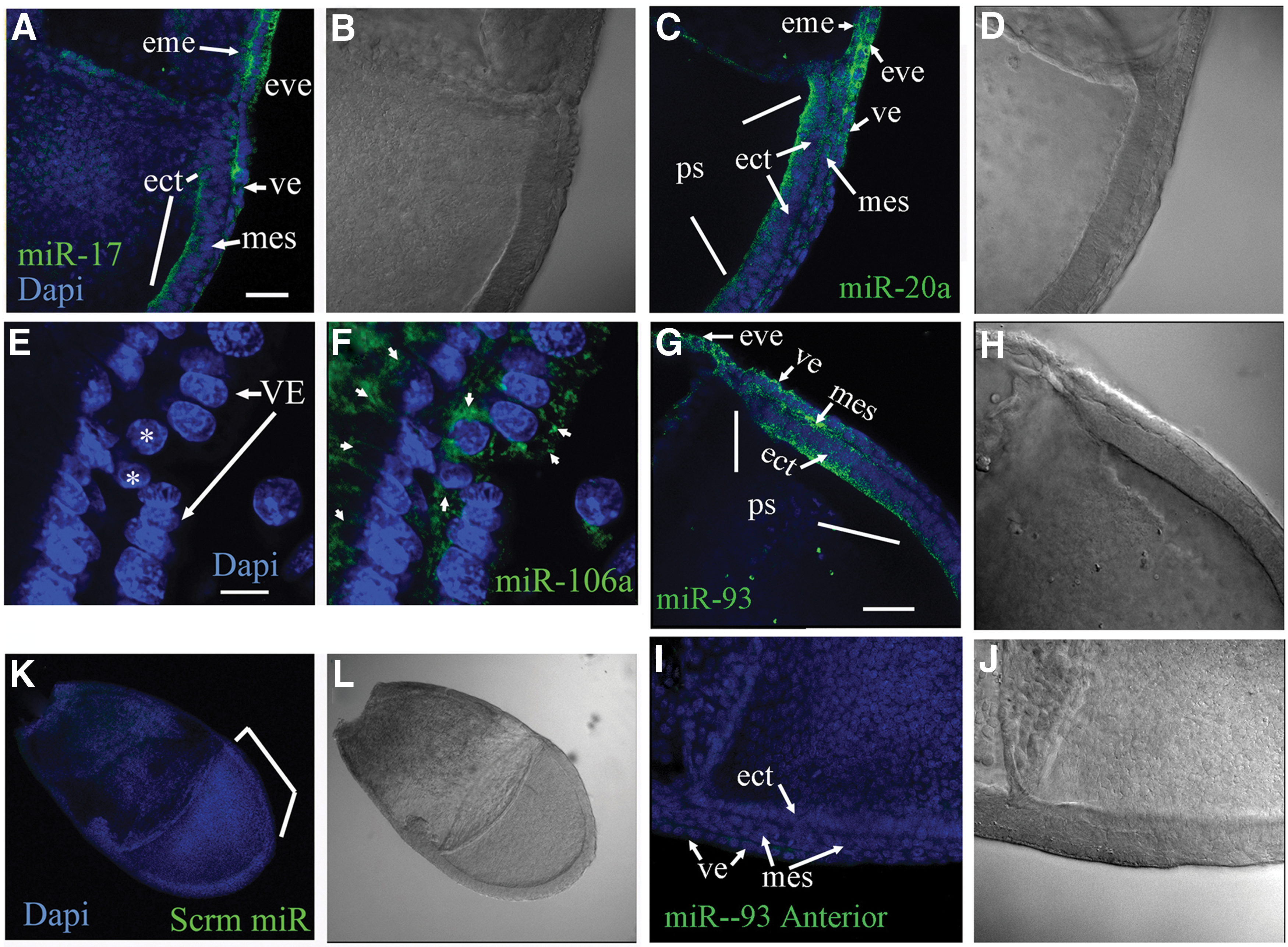
Fluorescent in situ hybridization was used to identify miRNA patterns in gastrulation-stage embryos (E7.0). miRNA probes labeled with fluorescein isothiocyanate to miR-17-5p
To begin analyzing how these miRNAs might affect BMPR2 protein levels, we first had to identify BMPR2 expression in embryos. In vivo, the protein expression pattern of BMPR2 had not previously been known at this early stage of development. We found that BMPR2 levels were distinctly lower near the primitive streak when compared with the anterior regions of the embryo (red fluorescent figures in Fig. 3A and insets and 3D and insets). Since Beppu et al. [36] had previously shown by in situ hybridization that the Bmpr2 gene was expressed throughout the primitive ectoderm, including the streak area, these new data identifying that BMPR2 protein expression is downregulated at the streak provide key evidence that miRNAs are the regulatory mechanism driving suppression of BMPR2 translation. Analyzing embryos in greater detail revealed that expression of downstream targets of BMPR2 signaling could also be directly correlated with expression of miR-17 family members and BMPR2 downregulation (Figs. 1E and 2). Specifically, confocal microscopy of frozen E7.0 embryo sections consistently showed a distinct downregulation of both BMPR2 and BMPR2-regulated genes Cx43 and E-Cad at the primitive streak (Fig. 3B and insets and 3E and inset, respectively). Most interesting, we found ID3 downregulated in primitive ectoderm and mesoderm at the primitive streak but not in the endoderm or primitive ectoderm anterior to the streak (Fig. 4A–D’). This expression pattern is telling, because it revealed that ID3 can promote E-cad expression in anterior regions by binding to the E12/E47 repressor of E-cad. We consistently find E12/E47 expressed in nuclei throughout the embryonic ectoderm (Fig. 4E–H). The lack of ID3 specifically at the streak would enable E12/E47 suppression of E-cad and, ultimately, induction of EMT.
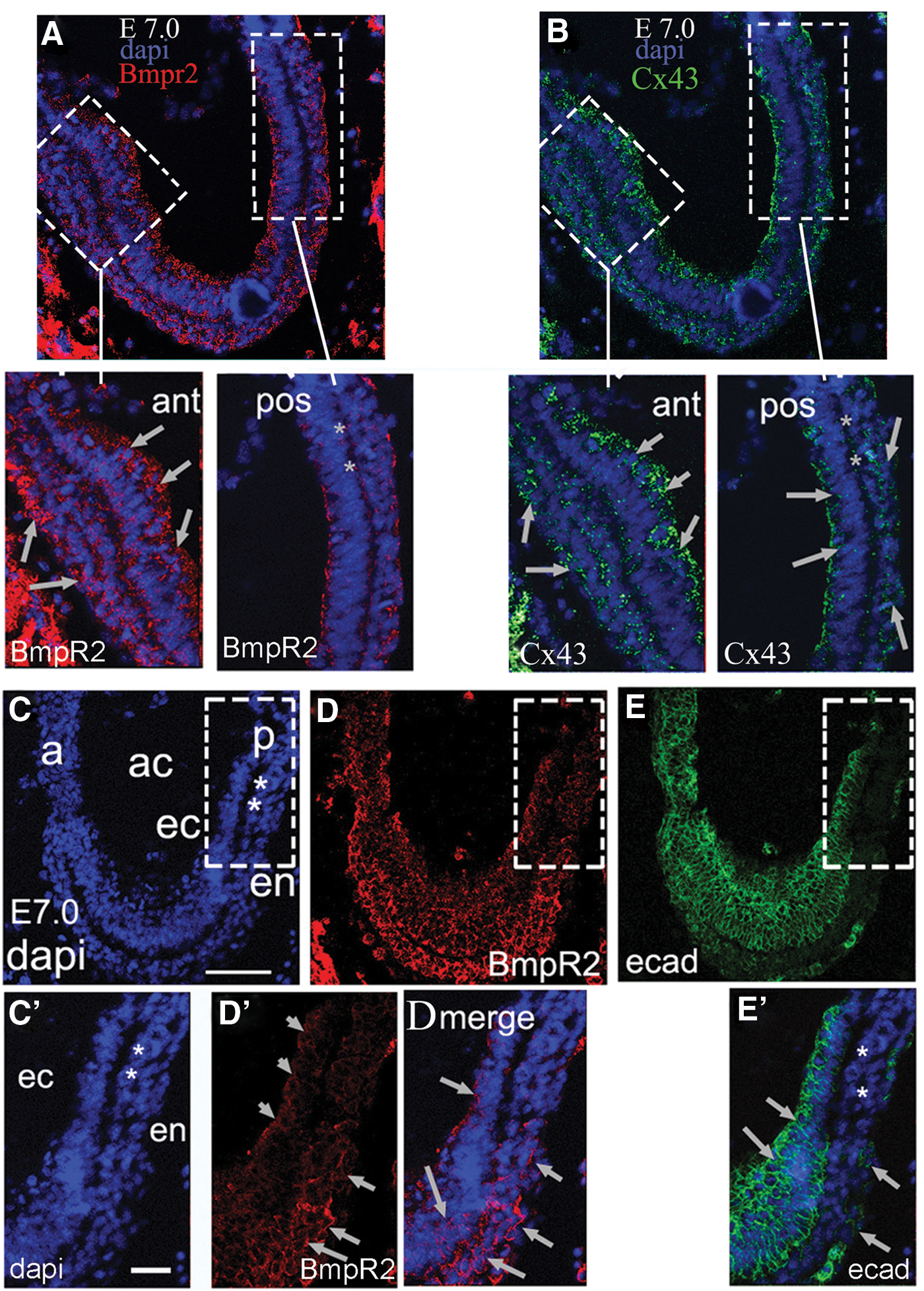
Confocal immunofluorescent images of E7.0 embryos probing for BMPR2 and its downstream targets show loss of BMPR2 at the primitive streak (ps)

ID3 and E12/E47 protein expression was identified using immunofluorescent staining on E7.0 embryos.
miRNA-17-5p, -20a, -93, and -106a targeting of BmpR2 regulates the BmpR2 pathway early in development
Since knockout (KO) mice of Bmpr2 and two of its signaling Smads, SMAD1 and 5, die at or soon after gastrulation, and as Bmpr2 expression corresponds to self-renewal [47], we surmised that spatiotemporal regulation of Bmp-Smad signaling by miRNAs was an important step for regulating aspects of gastrulation. First, we verified the in silico data. In our initial set of experiments, utilizing luciferase constructs linked to the 3′UTR of Bmpr2, we found that these miRNAs, indeed, targeted the Bmpr2 3′UTR. We transfected the construct into HeLa cells because of their reputation for being relatively easy to transfect. We found that mimics of miR-17-5p, -20a, -93, and -106a significantly reduced luciferase expression in every experiment (Fig. 5A). Interestingly, we found that transfection of inhibitors of these miRNAs significantly increased luciferase expression, suggesting that these miRNAs are endogenously present in HeLa cells and may have a function in their physiology. These results are being further pursued in a separate project. We concluded that these miRNAs, indeed, target the 3′UTR of Bmpr2. We should note that we transfected the full complement of four mimics or inhibitors in cells for these experiments, because all of them are expressed at the primitive streak in embryos. We felt this scenario was more physiologically correct than transfecting individual mimics or inhibitors.
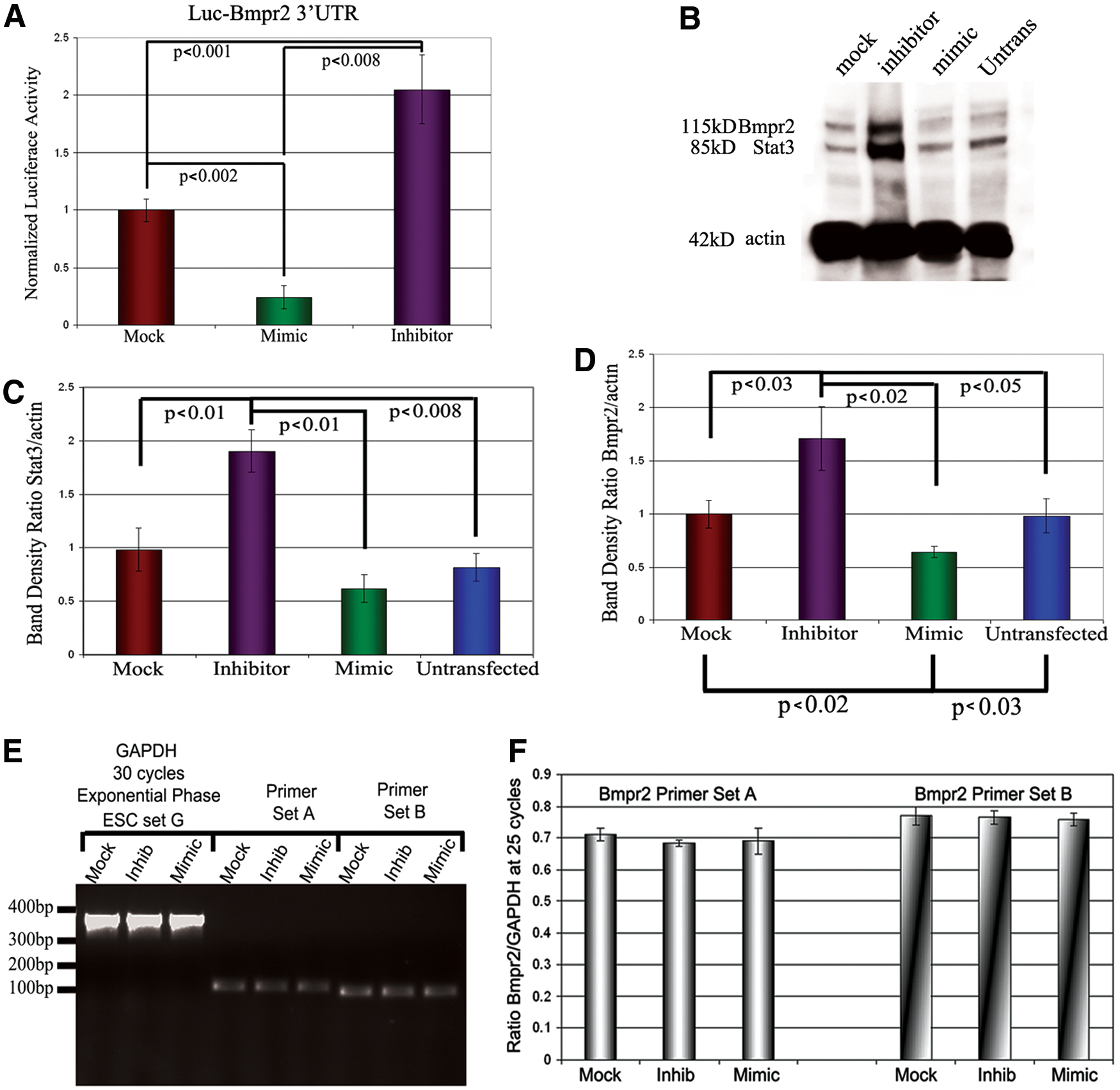
The miRNAs bind to the 3′UTR of Bmpr2 and suppress BMPR2 protein expression.
We then investigated whether these miRNAs affected BMPR2 protein expression levels in ESCs and EBs. After transfecting ESCs with mock miRNA, inhibitors, or mimics to these miRNAs, BMPR2 levels were analyzed in EBs by western blot after 2 days of differentiation. Both BMPR2 and STAT3 levels remained elevated in EBs transfected with inhibitors, while mimics showed decreased levels of expression (Fig. 5B). Quantifying the intensity of the bands showed that expression of both BMPR2 and the control STAT3 was significantly higher in EBs treated with miRNA inhibitors when compared with control and mimic-treated EBs. In contrast to inhibiting miRNAs, mimic-treated EBs showed a significant drop in BMPR2 (Fig. 5C, D).
We wondered whether these exogenous miRNAs were affecting protein levels by degrading the mRNA or by suppressing translation. Semiquantitative RT-PCR results revealed that mimics do not degrade Bmpr2 mRNA even after 2 days of transfection (Fig. 5E, F). Consequently, the data provide evidence that protein levels are reduced at the level of translation. To confirm that the transfected small RNAs were entering ESCs within colonies, we used a fluorescently labeled mock miRNA (Supplementary Fig. S1). A concentration-curve experiment confirmed that transfection efficiency was robust, as a majority of cells present in culture are transfected, especially at a concentration of 200 nM per individual miRNA mimic or inhibitor. We also wanted to know how long these transfected small RNAs were biologically available within EBs. Semiquantitative RT-PCR of the mimics at 4 days after transfection clearly showed that the synthetic mimics for all four miRNAs remained significantly elevated above endogenous levels of miR-17-5p, -20a, -93, and -106a (Supplementary Fig. S2).
To evaluate whether miR-17-5p, -20a, -93, and -106a could influence the expression of genes downstream of BMPR2, we used the highly reliable large-scale mESCs and EB model systems. Undifferentiated mESCs were transfected with miRNA inhibitors or mock miRNA and then allowed to differentiate into EBs. Samples were then obtained for RNA (at day 3 of differentiation) and protein analyses post removal of LIF (Days 2–5 of differentiation). qRT-PCR analyses of Ids 1 and 3, and Cx43 revealed significant differences in gene expression between miRNA inhibitor-treated and mock-treated EBs. Inhibition of miRNAs-17-5p, -20a, -93, and -106a resulted in persistent expression of genes directly downstream of the Bmpr2 pathway that would usually decrease during differentiation as represented by the mock control (Fig. 6A) [51].

miRNAs 17-5p, 20a, 93, and 106a regulate protein expression in ESCs and embryos and Bmpr2 downstream genes.
Protein expression of BMPR2-regulated downstream genes mirrored gene expression data, as inhibition of miR-17-5p, -20a, -93, and -106a resulted in prolonged elevation of IDs 1 and 3, CX43, and E-CAD when compared with mock miRNA (Fig. 6B). Notably, Brachyury expression was markedly lower in inhibitor-treated EBs, suggesting that differentiation was, at least, partially impaired. These results support our hypothesis in that inhibiting these miRNAs enables persistent expression of BMPR2 and, thus, persistent expression of genes downstream of the Bmpr2 pathway. Western analyses using antibodies to phospho-SMAD1/5 helped confirm that these miRNAs were affecting gene expression through the Bmpr2 pathway (Fig. 6C). To verify that phospho-SMAD1/5 remained elevated in EBs treated with the miRNA inhibitors compared with mock-treated EBs, western blot analyses of EBs after 4 days of transfection revealed that phospho-SMAD1/5 was significantly elevated in inhibitor-treated EBs compared with mock-treated EBs. Total SMAD1/5 was only slightly affected (Fig. 6D, E). These data suggest that not only BMPR2 protein levels stay elevated when these miRNAs are inhibited, but also the Bmpr2 pathway remains functionally active.
miRs 17-5p, 20a, 93, and 106a can be inhibited ex vivo in murine embryos
While our in vitro system nicely identified that miR-17-5p, -20a, -93, and -106a can function to regulate the Bmpr2 signaling pathway during differentiation of mESCs, we wanted to test whether these correlations were physiologically relevant within embryos. To accomplish this, embryos were procured at E6.0 from super ovulated, impregnated females. Isolated embryos were dissected at their embryonic-extraembryonic boundary and then transfected and incubated overnight on a slow rocker in mock miRNA or inhibitors to miR-17-5p, -20a, -93, and -106a at 37°C. Embryos were collected the next morning for protein analysis. In each case, when embryos were transfected with miRNA inhibitors, expression of BMPR2 and its corresponding downstream gene remained elevated when compared with embryos transfected with mock miRNA (Fig. 7A–C). The morphology of embryos incubated overnight was sufficient to identify that a visceral endoderm did not appear to form properly in any embryo (n=20) when compared with mock-treated embryos (Fig. 7D–F). These results provide evidence that miR-17-5p, -20a, -93, and -106a function to downregulate Bmpr2 and its downstream targets during development.
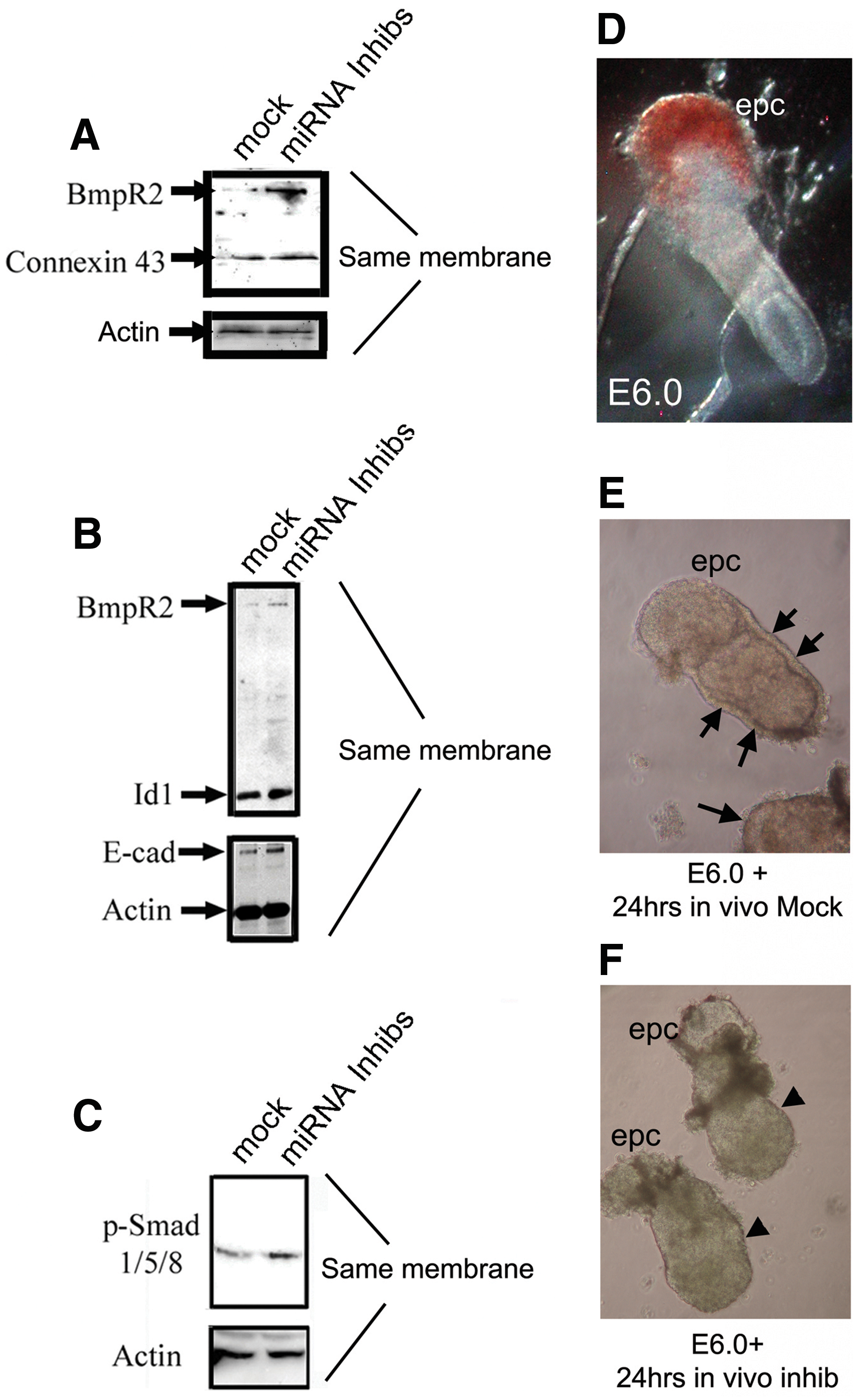
A semi-in vivo system was used to analyze the effects of mock miRNAs and miRNA inhibitors to miR-17-5p, -20a, -93, and -106a on genes downstream of Bmpr2 during gastrulation. Freshly dissected pregastrulation embryos (E6.0) were cultured at 37°C on a rocker in either mock miRNAs or inhibitors.
miR-17-5p, -20a, -93, and -106a affect actin dynamics independent of the canonical Bmpr2 signaling pathway
While differentiation is a key event of gastrulation, cell movement and ingression are essential for proper embryo formation. One key observation we consistently observed when treating ESCs with miRNA mimics was an obvious abundance of filopodia, especially when compared with ESCs treated with miRNA inhibitors. Foletta et al. [52] showed in COS-7 cells that the cytoplasmic tail region of BMPR2 directly associates with LIMK [52]. When liberated from this kinase, BMPR2 phosphorylates cofilin, suppressing its activity to sever actin. However, recently, a novel function for p-cofilin was identified where p-cofilin was shown to activate Rac1 by binding to and activating phospholipase D1 (PLD1) [53 –55]. Both the loss of function to sever actin (because of its phosphorylation by LIMK) and its activation of Rac1 can tilt actin dynamics toward filament elongation, including filopodia/lamellapodia formation [55]. While gastrulation of primitive ectoderm cells results in mesoderm cells acquiring the ability to undergo cell motility, the mechanism of this process has not been elucidated. As such, we speculated that the miR-17 family affects more than just differentiation during gastrulation. We hypothesized that downregulation of Bmpr2 by miR-17-5p, -20a, -93, and -106a creates a situation in which LIMK no longer has BMPR2 as a binding partner. Newly transcribed LIMK then becomes orphaned and is free to phosphorylate cofilin, resulting in Rac1 activation and a change in actin dynamics that can result in actin dynamics that promotes cell movement.
To test this hypothesis, we initially turned to the mESCs model system. First, ESCs were incubated in mock miRNA controls, mimics, or inhibitors to miR-17-5p, -20a, -93, and -106a, followed by removal of LIF to induce differentiation. After 2 days of differentiation, colonies were fixed and subjected to rhodamine-phalloidin (actin stain) and the nuclear dye Dapi, followed by analysis for filopodia formation. In this set of experiments, transfection of inhibitors consistently resulted in EBs with a few filopodia, while miRNA mock- and mimic-treated mESCs consistently resulted in EBs with significantly more filopodia, respectively (Fig. 8A–D). Next, to determine whether p-cofilin could be the mechanism driving actin dynamics during gastrulation, again, we subjected ESCs to mock miRNA controls, mimics, or inhibitors of miR-17-5p, -20a, -93, and -106a, followed by removal of LIF to induce differentiation. After 2 days of differentiation, colonies were fixed and subjected to rhodamine-phalloidin, antibodies directed against p-cofilin, and the nuclear dye Dapi (Fig. 9A–C). Using the computer program Cell Profiler 2.0 that was programmed to count all positive cells within an EB, we found that colonies treated with miRNA mimics contained significantly more cells with p-cofilin than cells treated with inhibitors and mock miRNAs (Fig. 9A–D and Supplementary Fig. S3). Western blot analyses of treated EBs revealed that p-cofilin levels are affected by miR-17-5p, -20a, -93, and -106a, as inhibiting them resulted in a significant decrease in p-cofilin levels when compared with EBs that were subjected to mock miRNAs or mimics (Fig. 9E). Further support for our hypothesis was found in cells undergoing EMT-like differentiation from EBs (Fig. 9F–H). Specific cells emanating from EBs showed highly colocalized staining for LIMK, p-cofilin, and GTP-bound active Rac1, which was identified using an antibody that specifically binds the active form of GTP-bound Rac1 and not GDP-bound Rac1.
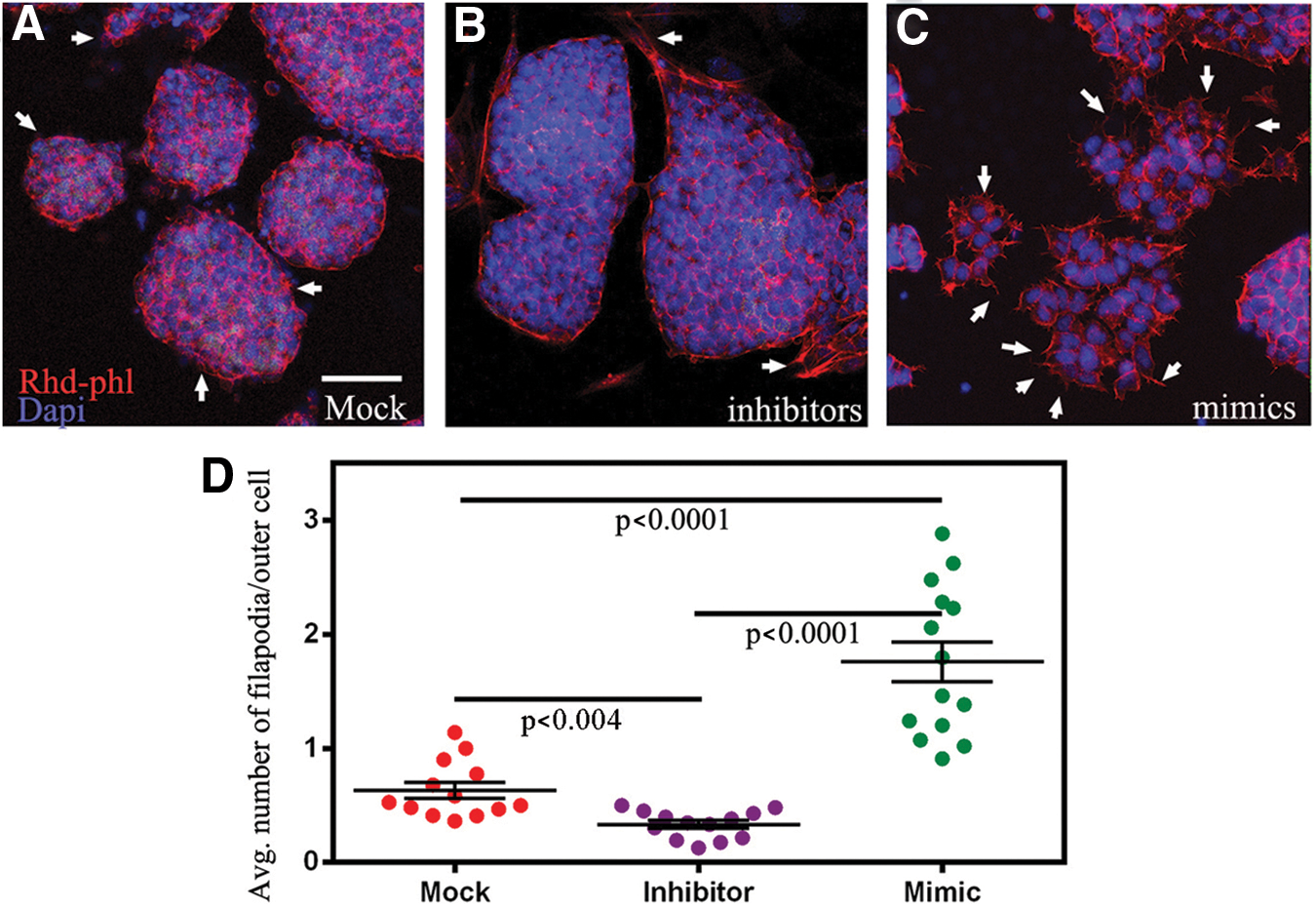
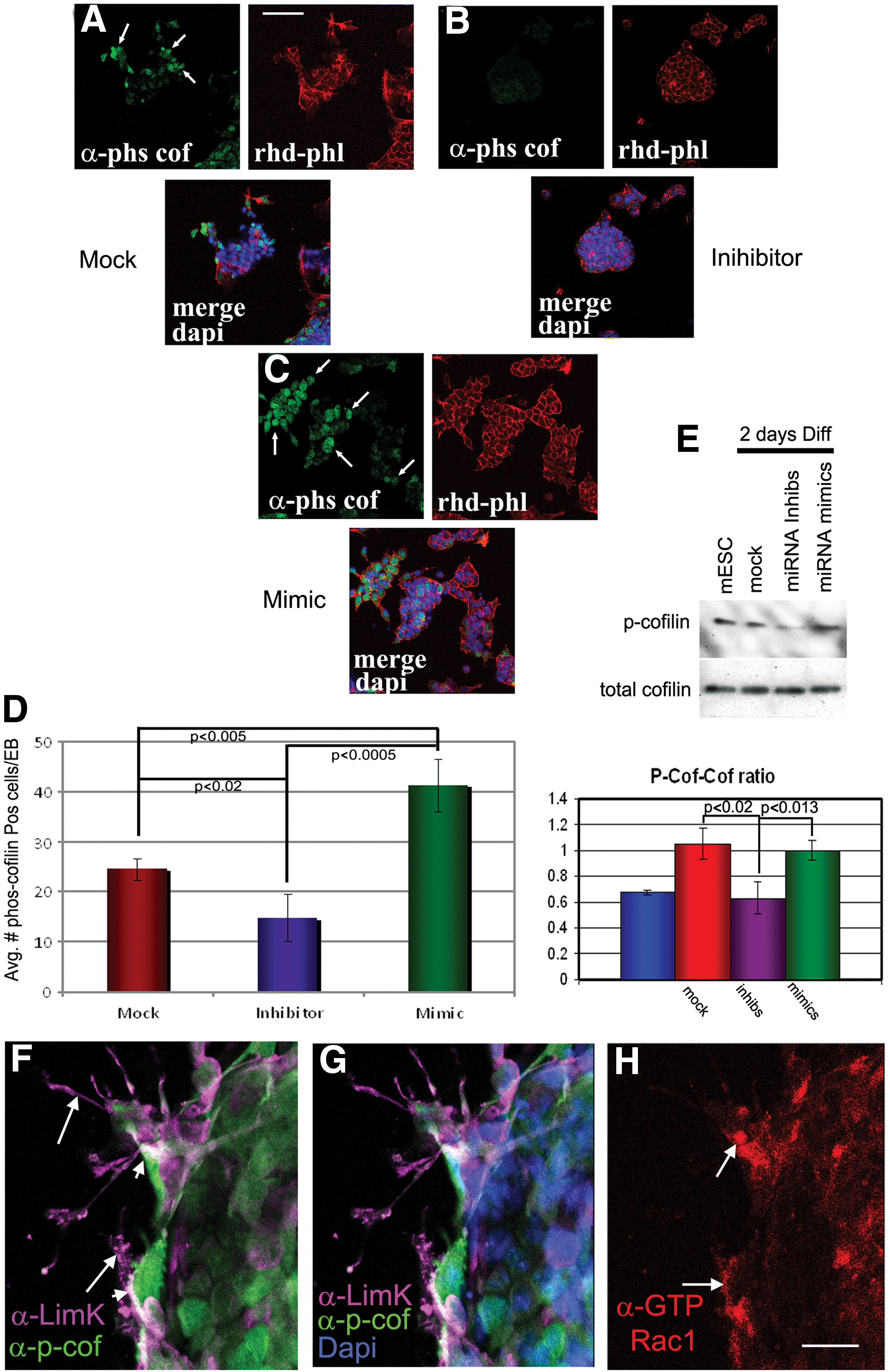
Confocal microscopy of mock, inhibitor, and mimic-transfected ESCs was differentiated into EBs for 2 days and stained with antibodies to p-cofilin and rhodamine-phalloidin
Finally, to determine physiological relevance of our ESC/EB data, we looked in embryos to identify the localization pattern for p-cofilin, LIMK, and GTP-bound active Rac1 (Fig. 10A–K). In primitive ectoderm cells, the cortical actin cytoskeleton defined their columnar morphology (Fig. 10A–C); however, once cells undergo EMT and proceed through the primitive streak, nascent mesoderm cells should begin to migrate, a process heavily reliant on actin dynamics and reorganization of the actin network, as clearly revealed by rhodamine-phalloidin staining (Fig. 10C) [37,56,57]. Antibodies directed against p-cofilin revealed elevated expression of p-cofilin within primitive ectoderm cells bordering the primitive streak as well as nascent, migrating mesoderm cells (Fig. 10D–F). There also was a high correlation of LIMK staining and p-cofilin staining as seen in Fig. 10G and H. High magnification further revealed the staining primarily at the presumptive leading edges of mesoderm cells. In part, these data provided strong evidence that LIMK has access to phosphorylate cofilin in cells at the primitive streak. Delving deeper, we found that when E7.0 embryos were analyzed using the antibody against GTP-bound active Rac1, markedly higher levels of active Rac1 were consistently observed within cells comprising primitive ectoderm, mesoderm/mesendoderm, and visceral endoderm that populate the primitive streak when compared with the anterior side of the embryo (Fig. 10I, J). Higher magnification of the primitive streak region further revealed active Rac1 at the leading edges of mesoderm cells and in some primitive ectoderm that had not yet ingressed. The data show that Rac1 is activated in the same region of cells that are positive for p-cofilin. Thus, when taken together, the ESC/EB confocal, western blot, 3′UTR luciferase data, and semi-in vivo embryo data provide evidence that miRNAs 17-5p, 20a, 93, and 106a can induce actin dynamics by downregulating Bmpr2 and activation of the p-cofilin/Rac1 pathway.
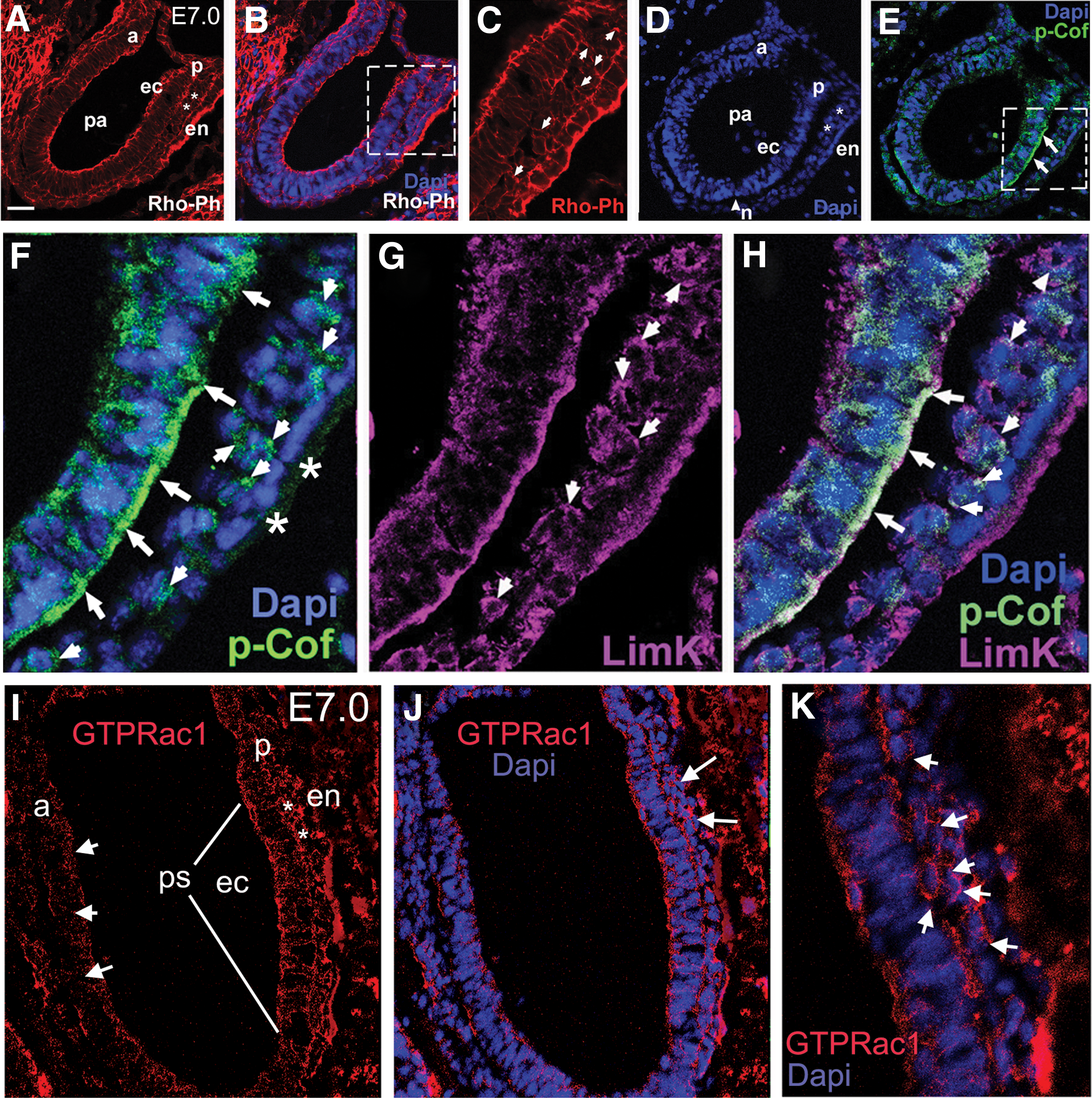
LIMK colocalizes with p-cofilin in cells undergoing gastrulation in E7.0 embryos.
Discussion
In Foshay and Gallicano [11], we used ESCs as a model system to help us define that miR-17-5p, -20a, -93, and -106a targeted Stat3 mRNA to briefly shut down STAT3 expression in cells approaching and proceeding through the primitive streak. This brief shut-down of STAT3 by this set of miRNAs has become one event among many necessary for pluripotent ectoderm cells to “decide” to differentiate [37,38,58 –60]. What makes our work unique is that we show that miRNAs can function to regulate essential pathways during gastrulation, providing the robustness important for development [10].
Here, the finding that a specific member of the TGFβ superfamily, Bmpr2, is a target of miRNAs 17-5p, 20a, 93, and 106a solidifies the importance of miRNAs controlling specific aspects of gastrulation. The data show that translation of BMPR2 protein expression is downregulated by miRNAs 17-5p, 20a, 93, and 106a at gastrulation. The result of BMPR2 downregulation then influences primitive ectoderm cell differentiation and their eventual movement once through the streak. This multimodal regulation appears to be a necessary component for normal development and when disrupted, results in prolonged expression of genes involved in self-renewal and epithelial lineages (ie, primitive ectoderm).
Targeting of Bmpr2 by miRNAs 17-5p, 20a, 93, and 106a suppresses the canonical BMPR2 signaling pathway
The schematic flow chart of the pathway regulated by BMPR2 in Fig. 1D as thoroughly defined by many laboratories enabled us to delve deeply into the affects that miRNAs have on its Bmpr2 target [41,42,48,50]. Applying this information to both ESCs and embryos uncovered the mechanistic role that these miRNAs play in driving development. Overall, two goals were accomplished: (1) we now have a better understanding of how miRNAs function in ESC biology and (2) we were able to apply and understand our miRNA/Bmpr2 findings to a semi in vivo model of embryogenesis.
FISH analysis of E7.0 embryos showed that miRNAs 17-5p, 20a, 93, and 106a localized to the primitive streak in embryonic regions and mixed areas in extraembryonic regions, including the extraembryonic endoderm. Importantly, though, in embryonic regions, expression was persistent in cells as they proceeded through the primitive streak but then seemed to subside as cells moved away and toward their fate. Knowing these miRNA expression patterns led to the hypothesis that they must be regulating aspects of gastrulation.
Directly working with gastrulation-stage embryos is difficult, primarily because of the dearth of tissue and cells to work with; however, the information we attained provided clear evidence that not only BMPR2 is downregulated at the primitive streak, but so are its downstream genes/proteins IDs 1 and 3, CX43 and E-CAD [45,47,48,61]. More importantly, we found that this regulation was through SMAD1/5, providing the evidence that these miRNAs can affect the canonical Bmpr2 pathway.
Although Ids 1 and 3 are expressed in the embryonic regions of the embryo ∼E6.0–6.5 [62], their patterns are different. Id3 mRNA expression is detected throughout the primitive ectoderm, while Id1 appears restricted to the most posterior and anterior regions of the primitive ectoderm [62]. It was elegantly shown in Ying et al. [47] that BMPs induce expression of Ids through BMPR2 (in combination with BMPR1a or 1b). Id gene downregulation is important for suppressing self-renewal and preserving multilineage differentiation. If Ids are not downregulated, ESCs seem to begin a quasi-differentiation pattern, but induction of specific lineages (eg, neural) is lost [47,63]. We clearly show for the first time that ID3 protein expression is downregulated in the streak region and that its expression pattern almost exactly matches FISH expression for all four miRNAs and decreased BMPR2 levels. qRT-PCR confirmed that miRNAs 17-5p, 20a, 93, and 106a regulate Id3 expression through Bmpr2.
Yancey et al. [64] demonstrated that CX43 was highly expressed in embryonic ectoderm cells and downregulated in mesoderm cells that have passed through the primitive streak [64]. Our in vivo data support their findings; however, we also showed that Cx43 loss of expression mirrors BMPR2 downregulation. Analyses of ESCs and subsequent EBs revealed that Cx43 gene expression is affected by miRNAs 17-5p, 20a, 93, and 106a through suppression of BMPR2 expression. Gap junctions, primarily composed of CX43, are present in ESCs [65]. When we subjected ESCs to inhibitors of miR-17-5p, -20a, -93, and -106a, persistent expression of CX43 resulted when compared with ESCs subjected to mock miRNA controls. Since the miRWalk algorithm does not predict Cx43 mRNA as a target for these miRNAs, it can be surmised that one function of these miRNAs is to facilitate downregulation of Cx43 through suppression of BMPR2 translation at the primitive streak, resulting in cellular disassociation and promotion of mesoderm formation.
Unlike Ids 1 and 3 and Cx43, E-cadherin is not a direct downstream gene in the Bmpr2 pathway; instead, it is indirectly regulated by BMPR2 through an intermediate step involving IDs association with the E12/E47 basic helix-loop-helix TF [66,67]. Under normal circumstances, IDs 1 and 3 dimerize with and inhibit E12/E47 DNA binding activity. E12 and E47 are alternative splice products of the E2A gene and usually function in tissue-specific gene regulation when dimerized with a tissue-specific class II bHLH [67 –69]. The Cano laboratory provided strong evidence that E47 associated with the E-boxes of the E-cadherin promoter acting as a suppressor [48]. E47 is expressed in ESCs; however, so is E-cadherin creating a paradox [70 –72]. This paradox is relieved because of the expression of IDs 1 and 3 and their ability to dimerize with and inhibit E12/E47. This suggests that when BMPR2 is expressed (as it is in ESCs and embryonic ectoderm), downstream IDs 1 and 3 bind to and inhibit E12/E47, releasing its suppression of E-cadherin. Since miRs 17-5p, 20a, 93, and 106a target BMPR2 expression at the primitive streak, IDs expression is also lowered, resulting in E12/E47, which is free to function as a suppressor of E-cad, and, thus, promotion of EMT. We, in fact, see this occur in vivo; FISH analyses show miRNAs expressed at the primitive streak, while confocal microscopy show lowered BMPR2 expression and concomitant lowered E-CAD expression at the primitive streak. In vitro, we see this occur in ESCs when we subject them to inhibitors of miR-17-5p, -20a, -93, and -106a. E-CAD protein remains elevated in EBs after removal of LIF.
Targeting of BmpR2 by miRNAs 17-5p, 20a, 93, and 106a suppresses a noncanonical BMPR2 pathway
A very specific observation we consistently noticed and measured when subjecting ESCs to miRNA inhibitors, followed by their differentiation (by removing LIF) was the virtual complete lack of filopodia extensions, especially when compared with miRNA mimic-treated ESCs, which sprouted filopodia from almost every outer cell of every EB. Usually, when ESCs are allowed to differentiate while attached to a surface, filopodia are easily identified within 2–3 days using conventional phase-contrast microscopy or confocal microscopy using rhodamine-phalloidin staining. We confirmed these observations after exacerbating the situation in ESCs by subjecting them to mimics of miR-17-5p, -20a, -93, and -106a. When differentiated for 2 days, significantly more filopodia/cell were observed when compared with EBs subjected to mock or miRNA inhibitors.
Clues to understanding these observations came from an elegant study by Foletta et al. [52], who showed that actin dynamics could be regulated by BMPR2 through its affinity for LIMK. Once liberated from BMPR2, LIMK is free to phosphorylate cofilin, rendering its actin-severing function inactive. We surmised that miRNA downregulation of BMPR2 would result in free-cytoplasmic LIMK, which, in turn, would result in phosphorylation of cofilin tilting actin dynamics toward polymerization and filopodia formation. Our western blot and confocal microscopy results revealed that when BMPR2 is downregulated by mimics to miR-17-5p, -20a, -93, and -106a, LIMK is free to phosphorylate its only known substrate, cofilin [55,56]. The key to these findings, though, was the detection of “Active” Rac1 in mimic treated cells when compared with cells treated with inhibitors. P-cofilin activates Rac1 through its interaction with PLD1. We see this event occurring in ESCs as they begin to differentiate and in embryos from our semi-in vivo system.
That said, it was our in vivo data that convinced us that what we were observing in vitro could actually occur in embryos at gastrulation. P-cofilin protein was found markedly elevated in the primitive streak with a pattern virtually identical to miRNA expression patterns. While LIMK was not elevated in the primitive streak, it did not necessarily need to be. What was important was that it colocalized with p-cofilin positive cells at the primitive streak and in nascent mesoderm cells that had recently undergone the EMT. These data support Foletta et al. [52] with regard to actin dynamics and LIMK; however, again what makes these data unique is that miRNAs are at the beginning of the cascade of events needed for ESCs to acquire the ability to undergo sensing of their environment to make the “decision” to move. Figure 11 illustrates our model of events in cells undergoing gastrulation.
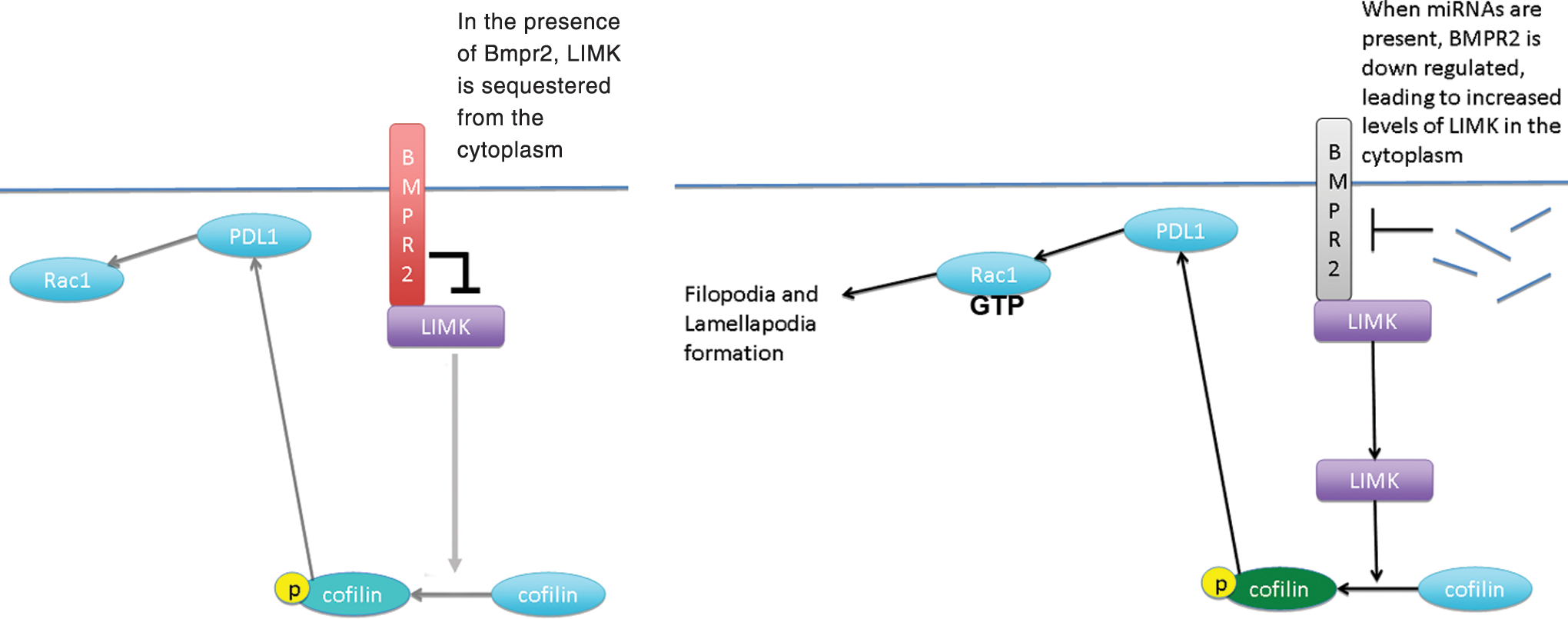
A model representing our hypothesis. Left side in the absence of miRNAs 17-5p, 20a, 93, and 106a; BMPR2 is present in the plasma membrane where it can bind to and sequester LIMK. Right side as miRNAs prevent BMPR2 translation, LIMK is freed into the cytoplasm where it can phosphorylate its only known substrate, cofilin, which, in turn through PLD1, activates Rac1, resulting in filopodia/lamellapodia formation and cell movement. PLD1, phospholipase D1. Color images available online at
A recent article from the Gumbiner laboratory gives further credence to our hypothesis [73]. They demonstrated the ability of the LIMK/cofilin link to decrease cell–cell junction stability in neural crest cells when BMPR2 is bound by BMPs. In this situation, BMPR2 releases LIMK, which can then phosphorylate cofilin, leading to cell–cell junction destabilization and EMT. Applying our data to their model would accomplish the same result; downregulation of BMPR2 by miRNAs would lead to junction instability and EMT at the primitive streak.
Understanding expression of miRNAs 17-5p, 20a, 93, and 106a in development helps define their roles in disease
Mis/over/underexpression of miRNAs has been confirmed in many types of diseases [27,28,74]. Specifically, members of the miR17 cluster have roles in hypertension, neuroblastoma, and B-cell development (accessible at
One of the best examples of how the loss of BMPR2 contributes to the cancer phenotype was from Harold Moses's laboratory, which showed that a dominant-negative Bmpr2 (DN-Bmpr2) promoted migration and invasion of mammary carcinoma cells [81]. EMT was also induced in DN-Bmpr2 expressing cells. Before that study, a number of studies had shown that BMPs were tumor promoters, especially when overexpressed [82 –84]; however, BMPs can bind other type 2 receptors such as activin receptors [42,85,86]. It appears that when BMPs bind BMPR2 receptors, cells lean toward an epithelial fate versus a more mesenchymal fate when BMPR2 is bound by activin receptors [87]. At the primitive streak, shutting down BMPR2 briefly (24–48 h) gives plenty of time to induce downstream effects similar to those shown in Owens et al. [81]. In that work, DN-BmpR2, when induced, promoted significant migration in mammary carcinoma cells within 24 h [81].
In essence, downregulation of BMPR2 promotes migration and characteristic mesenchymal fate in ESCs and embryos as it does in cancer cells. A difference between development and disease in this case is that cancer cells do not “turn off” the suppression of BMPR2 once cells migrate away from the tumor, while mesoderm cells turn off the suppression as they migrate away from the primitive streak because miR-17-5p, -20a, -93, and -106a expression dissipates. It would be interesting to inhibit miRNAs 17-5p, 20a, 93, and 106a in cancers where BMPR2 is suppressed to see whether cancer cells can revert to an epithelial fate.
Conclusions
miRNAs 17-5p, 20a, 93, and 106a can regulate essential pathways involved with gastrulation. We have previously shown that Stat3 is briefly shut down in ESCs and in embryos at the primitive streak by these miRNAs in order to relieve the self-renewal function of STAT3 expression [11]. We now find that Bmp signaling is also abrogated by these miRNAs at the primitive streak by their brief downregulation of BMPR2 expression. This regulation is actually a multimodal mechanism resulting in downregulation of genes involved in self-renewal (Id1/3), adhesion (Cx43, E-cad), and upregulation of actin dynamics (Limk/cofilin pathway).
A very important article by Ventura et al. [19] showed the phenotype of knocking out some or all 14 of the miRNAs in miRNA-17 cluster. Embryonic lethality ensued. Knocking out all 14 resulted in embryos with some differentiation into organs, but embryos were severely runted by E12 with numerous defects. Here, we provide evidence as to why those embryos died.
When mutated, the Bmpr2 pathway results in a number of different types of diseases. Interestingly, overexpression of miRs 17-5p, 20a, 93, and 106a correlates with many of the same diseases, including cancer, hypertension, and vascular disease. A next stage of investigation will be to identify the mechanism(s) regulating these miRNAs to alleviate the disease condition. While EMT is normal at gastrulation, it is a hallmark of diseases such as cancer. The work here is exciting, because it generates many more paths of research that follow for understanding and defining the differences between normal development and disease.
Footnotes
Acknowledgments
The authors thank Dr. Robert Benezra for helpful comments about the ID findings. They also thank Ms. Tammy Gallicano for her help with editing the article.
Author Disclosure Statement
There are no conflicts of interest among any of the authors.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.