
Abstract
Somatic cells can be reprogrammed to induced pluripotent stem (iPS) cells by defined sets of transcription factors. We previously described reprogramming of monolayer-cultured adult mouse ciliary body epithelial (CE) cells by Oct4 and Klf4, but not with Oct4 alone. In this study, we report that Oct4 alone is sufficient to reprogram CE cells to iPS cells through sphere formation. Furthermore, we demonstrate that sphere formation induces a partial reprogramming state characterized by expression of retinal progenitor markers, upregulation of reprogramming transcription factors, such as Sall4 and Nanog, demethylation in the promoter regions of pluripotency associated genes, and mesenchymal to epithelial transition. The Oct4-iPS cells maintained normal karyotypes, expressed markers for pluripotent stem cells, and were capable of differentiating into derivatives of all three embryonic germ layers in vivo and in vitro. These findings suggest that sphere formation may render somatic cells more susceptible to reprogramming.
Introduction
T
Sphere formation assays in a serum-free medium were first described as a method to select and expand stem cells [5,6]. Previous studies demonstrated that primary ciliary body epithelial (CE) cells have the ability to form clonal neurospheres, suggesting stem cells-like cells residing in the ciliary body epithelium [7]. CE sphere-derived cells expressed some retinal stem/progenitor cell markers in the culture [7,8]. However, the expression seems to be induced by growth factors in the medium. It remains unclear if these cells are bona fide quiescent retinal stem cells [8,9].
Generation of induced pluripotent stem (iPS) cells through ectopic expression of defined transcription factors holds great potential for regenerative medicine and disease modeling [10]. We previously found that mouse CE-derived cells, grown in monolayer cultures, express Sox2, Klf4, and c-Myc, and that these monolayer CE cells can be reprogrammed into iPS cells with ectopic Oct4 and Klf4 (2F), but not with Oct4 alone [1]. Given that adult neural stem cells, capable of sphere formation, have been previously reprogrammed into iPS cells with Oct4 alone [11,12], and that CE cells have the remarkable plasticity to form spheres displaying certain characteristics of neuroepithelial progenitors [7,8], we attempted to reprogram CE sphere-derived cells with Oct4 alone.
Materials and Methods
Animals
All procedures on mice were performed in accordance with the ARVO Statement for the Use of Animals in Ophthalmic and Vision Research and approved by the Institutional Animal Care and Use Committee at the University of North Carolina at Chapel Hill. Wild-type (WT) CD-1 strain mice, aged 5–6 weeks, were used for the study.
CE spheres and monolayer cultures
Dissection of adult mouse ciliary body was performed as previously described [7]. Dissociated cells were resuspended in a serum-free growth medium (SFM) containing Neurobasal medium, 1× B27 supplement, 1% penicillin/streptomycin, 2 mM
Induction of iPS cells
The procedure of iPS cells generation was essentially as previously described with minor modifications [1]. In brief, all spheres at day 7 were dissociated into single cells with Accutase (Sigma) and plated at 1×105 cells per well of gelatin-coated six-well plates in 1% FBS-containing SFM. CE monolayer cultures at passage 2 (7 days after isolation) were plated the same as above. The next day, for the two-factor transduction, concentrated lentiviruses containing CMV promoter-driven human Oct4 and Klf4 or Sall4 (Cellomics Technology) were added to the cells at a multiplicity of infection of 10 with 4 μg/mL polybrene (Sigma) in 1% FBS-containing SFM, whereas for the one-factor transduction, only lentivirus coding human Oct4 was added. Twenty-four hours post infection, the viral infection mix was exchanged for fresh 1% FBS-containing SFM. The following day, transduced cells were subcultured onto mitomycin C-treated SNL feeder cells (Cell Biolabs) in six-well plates at a split ratio of 1:4 in embryonic stem (ES) cell medium minus leukemia inhibitory factor (LIF) containing Dulbecco's modified Eagle's medium (DMEM) supplemented with 15% FBS, 2 mM
The efficiency of generating iPS cells was calculated by dividing the number of alkaline phosphatase (or Nanog)-positive ES cell-like colonies by the seeded cell number.
Viral vector integration analysis
Genomic DNA was isolated with the Quick-gDNA MiniPrep (Zymo Research) according to the manufacturer's instructions. Genomic polymerase chain reaction (PCR) was performed as previously described [1].
Karyotyping
Karyotyping was performed by KaryoLogic, Inc. (
Bisulfite genomic sequencing
Bisulfite sequencing and DNA methylation analysis was performed as previously described [1]. Briefly, genomic DNA was prepared with Quick-gDNA MiniPrep (Zymo Research) and treated with the EZ DNA Methylation-Lightning Kit (Zymo Research) according to the manufacturer's instructions. Previously published nested primer sets were used to amplify the promoter regions of the genes: Sox2, Nanog, and Oct4 [16,17]. Amplified fragments were cloned using the Zero Blunt TOPO PCR Cloning Kits (Invitrogen). Individual clones (10 or more) were sequenced with the SP6 and T7 primers.
RNA isolation, reverse transcription–polymerase chain reaction, and real-time PCR
Total RNA was isolated from the cells with the Direct-zol RNA MiniPrep (Zymo Research) with DNase I digestion. Total RNA of mouse ES-D3 cell line was purchased from Cell Biolabs. Complementary DNA was synthesized with Superscript III First-Strand Synthesis System (Invitrogen). PCR and Real-time PCR were performed as previously described [1]. Previously published primer sets were used [1,9,18,19]. The mRNA level of target gene was normalized to the housekeeping gene HPRT.
Immunofluorescence staining and alkaline phosphatase staining
Immunofluorescence staining was performed as previously described [1]. Briefly, fixed cells were treated with a blocking solution, followed by an overnight incubation with the primary antibodies at 4°C. After rinsing with phosphate-buffered saline, the cells were incubated with secondary antibodies, rinsed and counter-stained with Hoechst 33342. iPS cells were also stained with an Alkaline Phosphatase Staining Kit (Stemgent) following the manufacturer's instructions.
In vitro differentiation and teratoma formation
In vitro differentiation and teratoma formation were performed as previously described [1].
Statistics
In all real-time PCR experiments, n=3. P-value was calculated using the Student's t-test.
Results/Discussion
We isolated CE cells from ciliary body of CD1 WT mice, and cultured them as free-floating spheres in SFM (Fig. 1A). In agreement with previous studies [7 –9], our reverse transcription–polymerase chain reaction (RT-PCR) analysis revealed that the expression of retinal progenitor markers such as Pax6, Sox2, Rax, Otx2, Lhx2, Six3, and nestin were induced in the culture in the sphere and monolayer cells, whereas differentiated ciliary epithelial marker Palmdelphin (Palmd) remained stable in the sphere, but decreased in the monolayer, and another ciliary epithelial transcript Tyrosinase (Tyr) decreased progressively in the sphere and monolayer (Fig. 1B). Together, these results suggested that CE cells acquired neural progenitor properties following culturing in mitogen-containing SFM.
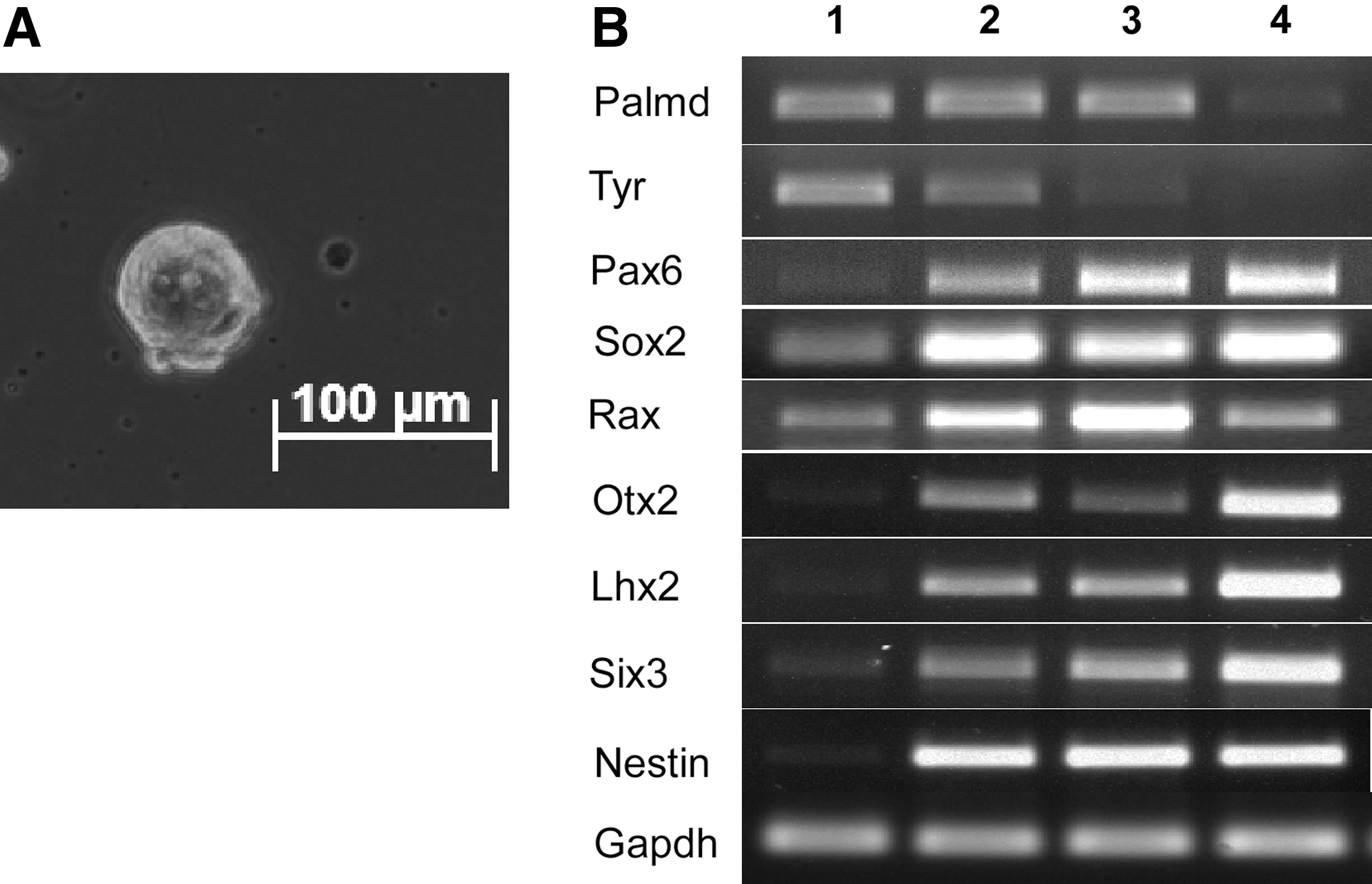
Mouse ciliary body epithelial (CE) cell culture.
Next, we reprogrammed CE sphere-derived cells with ectopic Oct4 and Klf4 (2F), and with Oct4 alone (Fig. 2A). At day 21 after infection, we observed ES cell-like colonies in 2F-transduced CE sphere-derived cells and about 44 alkaline phosphatase-positive colonies (44/100,000) after another 7 days, with a reprogramming efficiency close to the 2F control experiment using CE monolayer culture as described previously [1]. However, with Oct4 alone, no iPS cell colonies emerged, even after another 14 days. As described previously in the 1F experiment using CE monolayer culture, some morphological changes occurred, suggesting a partially reprogrammed state. When switching to serum-free N2B27 medium with dual inhibition (2i) of glycogen synthase kinase-3 and mitogen-activated protein kinase signaling at day 21 post infection, one ES cell-like colony (Fig. 2B, left) appeared at day 28 after infection from 100,000 seeded CE sphere-derived cells, with a reprogramming efficiency of 0.001% at day 35 (1/100,000, three times). In contrast, in the parallel 1F experiment using CE monolayer culture, iPS cell colonies were never observed, even with more initially plated cells (Fig. 2B, right, 0/106, two times).

Generation of induced pluripotent stem (iPS) cells from CE sphere-derived cells with reprogramming factor Oct4.
All of the three picked 1F colonies could be further expanded for more than 20 passages in serum-free N2B27 medium supplemented with 2i/LIF, displaying proliferation and a morphology characteristic of ES cells (Fig. 3A) and maintaining alkaline phosphatase activity (Fig. 3B). 1F-iPS cells were confirmed to have only Oct4 transgene by genomic PCR (Fig. 3C), suggesting that ectopic Sox2, Klf4, and c-Myc are dispensable for reprogramming CE sphere-derived cells into iPS cells. The established 1F-iPS cell lines were then further characterized. They expressed all tested endogenous pluripotency markers, as detected by RT-PCR (Fig. 3D), whereas the transgene hOct4 was significantly attenuated at late passages (Fig. 3E). With immunofluorescence staining, we confirmed transcription markers such as Oct4, Sox2, and Nanog, and pluripotent cell surface marker SSEA-1 (Fig. 3F). DNA methylation pattern in Oct4 and Nanog promoters in 1F-iPS cells were similar to that in embryonic stem cells (ESCs) (Fig. 3G). In addition, 1F-iPS cells maintained a normal karyotype (Fig. 3H). We next evaluated the differentiation potential of 1F-iPS cells in vitro. 1F-iPS cells effectively formed embryoid bodies (EB) (Fig. 3I). Ten days after EB transfer onto gelatin-coated plates, the attached cells expressed markers of three germ layers, including the ectoderm markers Nefl and Nestin, the mesoderm markers KDR, FLT1, and T (Brachyury), and the endoderm markers Foxa2 and GATA4 as determined by RT-PCR (Fig. 3J). To fully characterize the differentiation potential in vivo, we injected 1F iPS cells into immune-deficient NOD-SCID mice. Histological examinations showed that the generated teratomas contained derivatives of all three embryonic germ layers (Fig. 3K). We confirmed these morphological findings with immunofluorescence staining: neural epithelial-like tissues (ectoderm) were positive for tubulin-β III expression, muscle-like tissues (mesoderm) were positive for α-smooth muscle actin, and gut-like epithelium (endoderm) was positive for E-cadherin expression (Fig. 3L). These observations suggested that the 1F iPS cells were fully reprogrammed into pluripotency.
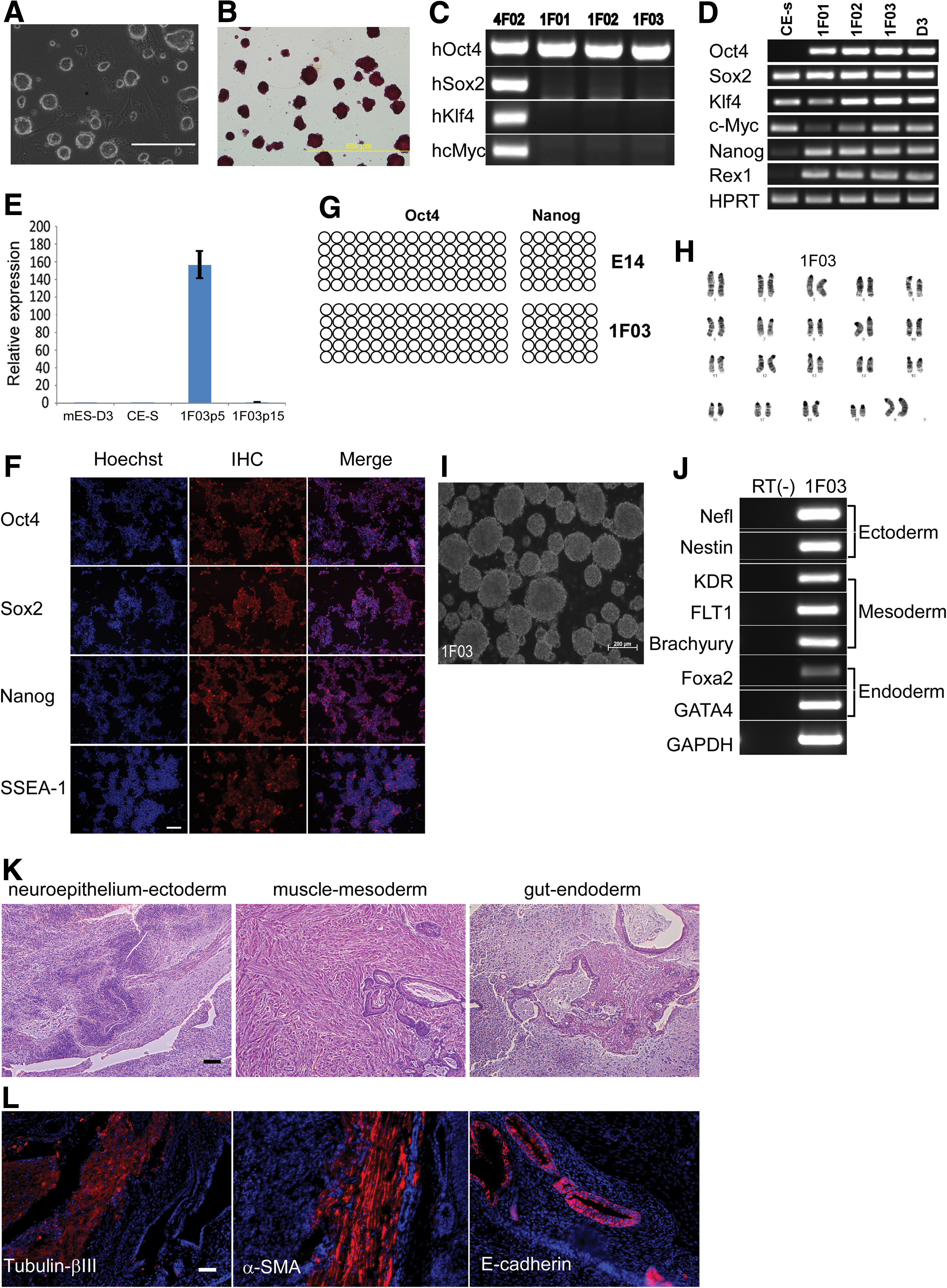
Characterization of 1F iPS cells.
To better understand why sphere formation makes CE cells amendable to 1F reprogramming, we evaluated mRNA expression of some key pluripotency-associated genes and genes involved in mesenchymal–epithelial transition (MET) by quantitative RT-PCR (Fig. 4A). Neither the CE sphere nor the monolayer culture expresses Oct4 at appreciable levels. Sox2 expression was high, but almost at the same level. Although Prdm14, Esrrb, and Dppa2 expression was increased, the expression levels of Sall4, c-Myc, Lin28, Glis1, Klf4, and Nanog were most significantly upregulated in the sphere compared with the monolayer. Combination of these factors with or without Oct4 has been shown to successfully reprogram mouse embryonic fibroblasts to iPS cells [20
–22]. Recently, Sall4 was identified as one of the two most induced genes (the other Sox2) in the early phase in response to the seven small molecule combination in a chemical reprogramming that efficiently drives iPS cells reprogramming in mouse somatic cells [18]. To assess whether Sall4 could enhance 1F-Oct4 reprogramming, we asked if overexpression of Sall4 in the CE monolayer with Oct4 could generate iPS cells. Surprisingly, we did not observe any ES-like colonies, which we performed twice. This suggests that additional mechanisms are responsible for the reprogramming advantage observed with CE spheres. Since we determined that CE monolayer could be reprogrammed with 2F (Klf4 and Oct4), we propose that upregulation of Klf4, or genes that can replace Klf4 function, can circumvent the requirement of Klf4. Indeed we found that Klf4, and Esrrb, Glis1 and Prdm14, which have been shown to replace Klf4 in reprogramming [21,23,24], were upregulated by CE sphere formation. Whether the upregulation of these genes is responsible for 1F-Oct4 reprogramming of CE sphere-derived cells requires further investigation. It is, however, tempting to speculate that one step of sphere formation might reduce the small molecule number and speed up the reprogramming process in a chemical reprogramming of CE spheres since Sox2 expression is already high or induced by sphere formation (Figs. 1B and 3D). Two de novo DNA methyltransferases, Dnmt3a and Dnmt3b, and the 10–11 Translocation-family demethylases, Tet1 and Tet2, were also expressed higher in the sphere, suggesting changes in DNA methylation during sphere formation [25]. In agreement with this observation, bisulfite sequencing evaluating the methylation status in the promoter regions of Oct4, Nanog, and Sox2 revealed that the methylation in CE spheres was lower than those in monolayer cells (Fig. 4B), indicating these genes were more transcriptionally activated than monolayer cells. Cdh1 and Epcam, which are beneficial to iPS cell generation [19,26], were higher in the CE sphere. However, the difference could result from downregulation of Cdh1 and Epcam in the monolayer due to the presence of transforming growth factor-β (TGF-β) in serum-containing medium. To rule out this possibility, we grew CE cells in the same serum-containing medium for monolayer culture, but in ultra-low attachment plates. Under such a condition, CE cells formed spheres, and Cdh1 and Epcam were still significantly expressed higher in the spheres (Supplementary Fig. S1; Supplementary Data are available online at
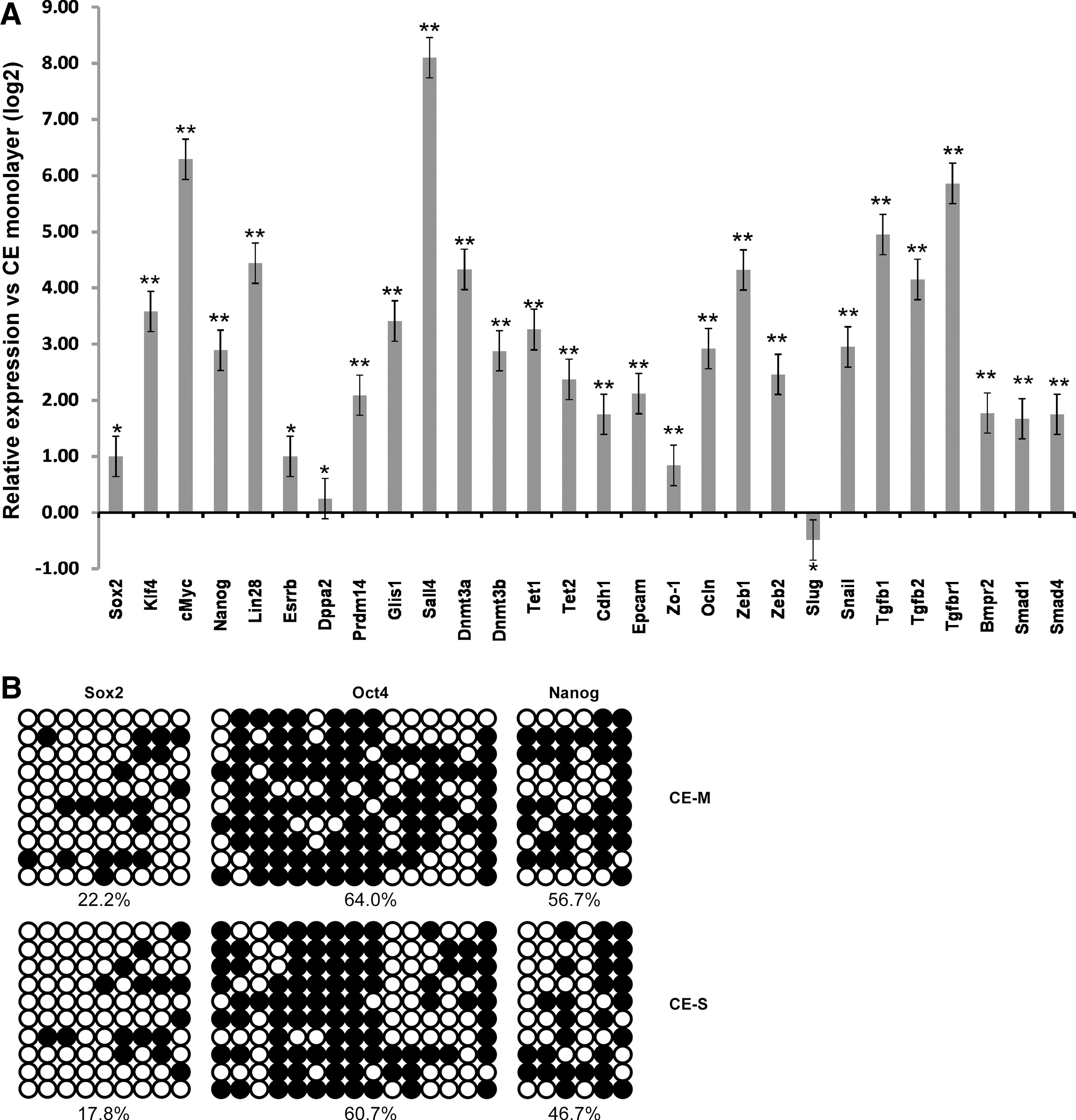
Gene expression in the CE sphere and monolayer.
In this study, we show sphere formation partially reprograms CE cells, and provides an iPS reprogramming advantage. Our observation may be applicable to other cell types such as fibroblasts. Previous studies have shown that forced growth of fibroblasts into spheres caused MET and induced stem cell/progenitor properties [29,30].
In summary, we demonstrated in this study that through sphere formation, Oct4 alone can reprogram CE sphere-derived cells into pluripotent stem cells. Sphere formation reprograms CE cells to progenitor-like cells with the expression of neural stem cell markers and upregulation of various reprogramming factors compared with the monolayer. These findings suggest a two-step reprogramming scheme, in which the first step of sphere formation induces somatic cells into a partially reprogrammed stage, followed by reduced ectopic input for iPS cell reprogramming.
Footnotes
Acknowledgments
S.H.C. would like to acknowledge Hope for Vision, Research to Prevent Blindness Career Development Award, Foundation Fighting Blindness Career Development Award, and NEI K-08 Career Development Award.
Author Disclosure Statement
Authors declare no conflict of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.