
Abstract
Neither the roles of Asp-His-His-Cys (DHHC)-containing proteins in embryonic cell fate specification are well defined, nor the underlying mechanisms of their activity are well understood. Here, we compared the embryonic function of zinc finger DHHC-type containing (Zdhhc13) in zebrafish embryos and in an in vitro cell model. Zdhhc13, a critical regulator of bone morphogenetic protein (BMP) signaling, specifically bound to Smad6 to induce its perinuclear accumulation and degradation through a mechanism independent of its palmitoyltransferase activity. We showed that Zdhhc13 played a crucial role during zebrafish embryogenesis in the control of germ layer specification, particularly in ectoderm and mesoderm differentiation homeostasis. Depletion of Zdhhc13 led to the neuralization of ectoderm and dorsalization of mesoderm in zebrafish embryos. Moreover, Zdhhc13 antagonized Smad6 during BMP-dependent signaling and early lineage decisions in our in vitro cell model. Our results extended the cellular role of Zdhhc13, suggesting that it acts as a regulator in BMP signaling, and established that the embryonic function of Zdhhc13 is in lineage specification.
Introduction
E
Increasingly, it is becoming apparent that developmental signals can be fine-tuned by modulating protein stability, localization, and activity through post-translational modifications [10
–12]. Palmitoylation is a key post-translational modification that is mediated by a family of Asp-His-His-Cys (DHHC)-containing palmitoyl acyltransferases (PATs) [13,14]. PATs exist in humans and other mammals, where they differ in their intracellular localization and tissue-specific expression [15]. DHHC proteins play a critical role in several biological processes, and disruptions in their function can result in disease. Therefore, PATs serve as potential medical biomarkers or even therapeutic targets. Zdhhc13, a characterized DHHC family member, is highly conserved from invertebrates to vertebrates (Supplementary Fig. S1; Supplementary Data are available online at
Here, we directly address the embryological role of Zdhhc13 in zebrafish embryos and in an in vitro cell model. In addition to the apparent conservation of their expression patterns during the early differentiation process, we show that the function of Zdhhc13 is conserved in vivo and in our in vitro cell system, highlighting its crucial function in regulating germ layer differentiation. Further analyses indicated that Zdhhc13 antagonized Smad6 during BMP-dependent signaling and specified ectoderm and mesoderm cell lineages during differentiation. To the best of our knowledge, these data provide novel insights into the function of Zdhhc13 and the mechanism of its action. These data provide a framework based on which we could fully understand the roles of the DHHC family of proteins and the disease processes caused by their dysfunction.
Materials and Methods
Plasmid construction
The sequence corresponding to mouse zdhhc13 mRNA 189–211 base pairs was chosen as the RNA interference (RNAi) target and inserted into the BglII/HindIII sites of pSuper, to generate the zdhhc13 RNAi expression vector.
Mouse full-length zdhhc13 cDNA was amplified using 5′-CGC
Zebrafish full-length zdhhc13 cDNA was amplified with 5′-CCG
Zebrafish full-length smad6 was amplified with 5′-GGA
Antibodies
The primary antibodies described in this article include anti-Sox2 (Abcam), anti-Nestin (Millipore), anti-Pax3 (Santa Cruz Biotechnology), anti-α-SMA(Abcam), anti-Sox17 (Santa Cruz Biotechnology), anti-Gata6 (Santa Cruz Biotechnology), anti-Smad1 (Abcam), anti-p-Smad1/5/8 (Cell Signaling Technology), anti-Smad2,3 (Cell Signaling Technology), anti-Smad4 (Cell Signaling Technology), anti-Smad6 (Abcam), anti-GFP (Santa Cruz Biotechnology), anti-HA (Santa Cruz Biotechnology), and β-actin (Sigma) antibodies.
Cell culture and immunofluorescence
P19 cells were cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco) that was supplemented with 10% fetal bovine serum (FBS; Gibco), 100 U/mL penicillin, and 100 μg/mL streptomycin. For stable transfection of zdhhc13 RNAi, P19 cells (40%–50% confluent) were transfected with pSuper-zdhhc13 RNAi with Lipofectamine 2000 (Invitrogen) according to the manufacturer's instructions. After 24 h, and every 48 h thereafter for 2 weeks, the culture medium was replaced with fresh medium containing 500 μg/mL of G418 (Invitrogen). Pools of 25 clones were isolated as stable transfectants with zdhhc13 RNAi in P19 cells, five of them showed a significant reduction of zdhhc13 mRNA in the noninduced state by reverse transcription–polymerase chain reaction (RT-PCR), and clones 9 and 19 were chosen for further studies (Supplementary Fig. S3C). For differentiation, the monolayer P19 cells were cultured in 10% FBS/DMEM/F12 (Gibco) in the presence of 10−6 M retinoic acid (RA) (Sigma) for 2 days, and then, the medium was changed to N2 medium (Gibco) for 6 days before immunostaining and western analysis. zdhhc13 up-regulation might be considered a PSC-related event and independent from RA induction (Supplementary Fig. S3B).
Cells were fixed in 4% paraformaldehyde in phosphate-buffered saline (20 mM NaH2PO4, 0.9% NaCl, pH 7.4), permeabilized with 0.3% Triton X-100, and primary antibodies were labeled overnight at 4°C followed by anti-mouse IgG antibody conjugated to Alexa green 488 (Beyotime) or anti-rabbit IgG antibody conjugated to Alexa red 555 (Beyotime).
Zebrafish maintenance
Zebrafish of AB strains were maintained and bred according to standard procedures [19].
Morpholino and synthetic mRNA injections
For loss-of-function analysis, antisense morpholinos (MO) were designed, purchased from Gene Tools, and injected into 1-cell stage. Two zdhhc13-MOs were used in this work, targeting the translation starting point and the sequence in the 5′ untranslated region, respectively. The sequences were ATCCGTCTCCTTCTGACCAGTCCAT (zdhhc13-ATG-MO) and CTGTACGAGGTTATCCGTGATTTAC (zdhhc13-5′UTR-MO). The control MO (Con MO, ATCCGTgTgCTTgTGACCAcTgCAT) was the five mismatches with regard to zdhhc13-ATG-MO. The defects in morphogenic events were dose dependent. An injection of 8 ng either ATG-MO or 5′UTR-MO per embryo showed the same effects and resulted in abnormal morphogenesis in a majority of embryos; so, we performed subsequent experiments by a microinjection of 8 ng MO.
Mouse zdhhc13 and zdhhc13 ΔDHHC mRNA was in vitro transcribed from linearized pCS2+ constructs using the Sp6 message machine kit (Ambion). Synthetic mRNA was injected into the 1-cell stage either alone or combined with MO.
In situ hybridization and immunolabeling
Whole-mount in situ hybridization was carried out as previously described [20]. Antisense RNA probes were synthesized with digoxigenin RNA labeling kits (Roche).
Procedures for immunolabeling were similar as previously described [21]. Embryos in the shield stage were fixed in 4% paraformaldehyde and embedded in OCT. Cryostat sections were collected at 8 μm in thickness, and primary antibodies were labeled overnight at 4°C followed by incubation with appropriate secondary antibodies conjugated to Alexa green 488 or Alexa red 555.
Quantitative real-time PCR and conventional RT-PCR
Quantitative RT-PCR (qRT-PCR) was performed using iQSybr Green Supermix (Bio-Rad). cDNA levels were determined using relative quantification and normalized to β-actin or ef1α. The protocol was initial denaturation at 95°C for 10 s, and then 95°C for 10 s, 55°C for 15 s, and 72°C for 20 s, repeated for 40 cycles. For every assay, the negative control was without a cDNA template. The 2−ΔΔCt method was used to analyze data. The optimal primers for PCR are as follows: zdhhc13 (forward, 5′-CTGTGCTGTTCTGGCTGGT-3′; reverse, 5′-GCATGTGAATGAGCAGAGG-3′); smad6 (forward, 5′-TTGCAACCCCTACCACTTCA-3′; reverse, 5′-AGTGATGAGGGAGTTGGTGG-3′); sox1 (forward, 5′-CAAGATGGCCCAGGAAAACC-3′; reverse, 5′-ATCCGGGTGTTCCTTCATGT-3′); sox2 (forward, 5′-AACATGATGGAGACGGAGCT-3′; reverse, 5′-TTCTCCTGGGCCATCTTACG-3′); pax3 (forward, 5′-AGAGAACCCGGGCATGTTTA-3′; reverse, 5′-GATGCCATCGATGCTGTGTT-3′); gata2 (forward, 5′-GATGAATGGACAGAACCGGC-3′; reverse, 5′-TTCTTCATGGTCAGTGGCCT-3′); sox17 (forward, 5′-GCACAGCAGAACCCAGATCT-3′; reverse, 5′-CCGGTACTTGTAGTTGGGGT-3′); gata6 (forward, 5′-TTCTACACAAGCGACCACCT-3′; reverse, 5′-CACCAAGAATCCTGTCGCAC-3′); oct4 (forward, 5′-CCTTGCAGCTCAGCCTTAAG-3′; reverse, 5′-GCGATGTGAGTGATCTGCTG-3′); β-actin (forward, 5′-CCTCTATGCCAACACAGTGC-3′; reverse, 5′-CCTGCTTGCTGATCCACATC-3′); and ef1α (forward, 5′-AGCTTCTGCAGACGGTTCAT-3′; reverse, 5′-GACCTCCTTGAGCTCCT-3′).
Immunoprecipitation and immunoblotting
Cell lysate preparation and immunoprecipitation were performed as previously described [22]. Immunoprecipitation was carried out using anti-HA polyclonal antibody (Santa Cruz Biotechnology) or normal rabbit IgG (Thermo). The proteins were then eluted and analyzed by western blotting as previously described [23].
Image and statistical analysis
Images were acquired with a stereomicroscope (Olympus SZX16) for in situ hybridizations and a fluorescence microscope (Olympus IX71) for immunostaining. Images were adjusted for brightness and contrast using Image-Pro Plus 6.0 and Adobe Photoshop software.
Statistical significance was analyzed using GraphPad Prism 5.0 Software and performed with the Student's t-test. All bar graphs are plotted as mean±standard error of the mean.
Results
Zdhhc13 is expressed during zebrafish embryogenesis
The zebrafish genome contained a single gene (zdhhc13) that encoded a Zdhhc13 protein which shared 82.4% amino-acid sequence identity to human ZDHHC13 (Supplementary Fig. S1). Using in situ hybridization and RT-PCR analysis, we examined both maternal and zygotic zdhhc13 expression at different stages of embryonic development (Fig. 1A, B). The zdhhc13 transcripts were ubiquitously distributed in the blastoderm until the shield stage (Fig. 1Aa–c). The transcript levels decreased during gastrulation and increased in the head region during the segmentation period (Fig. 1Ad, e). After 24 hours postfertilization (hpf ), zdhhc13 was weakly expressed in the head region and was almost undetectable in other regions (Fig. 1Af ). These expression patterns suggested that Zdhhc13 was involved in the early processes of zebrafish development.
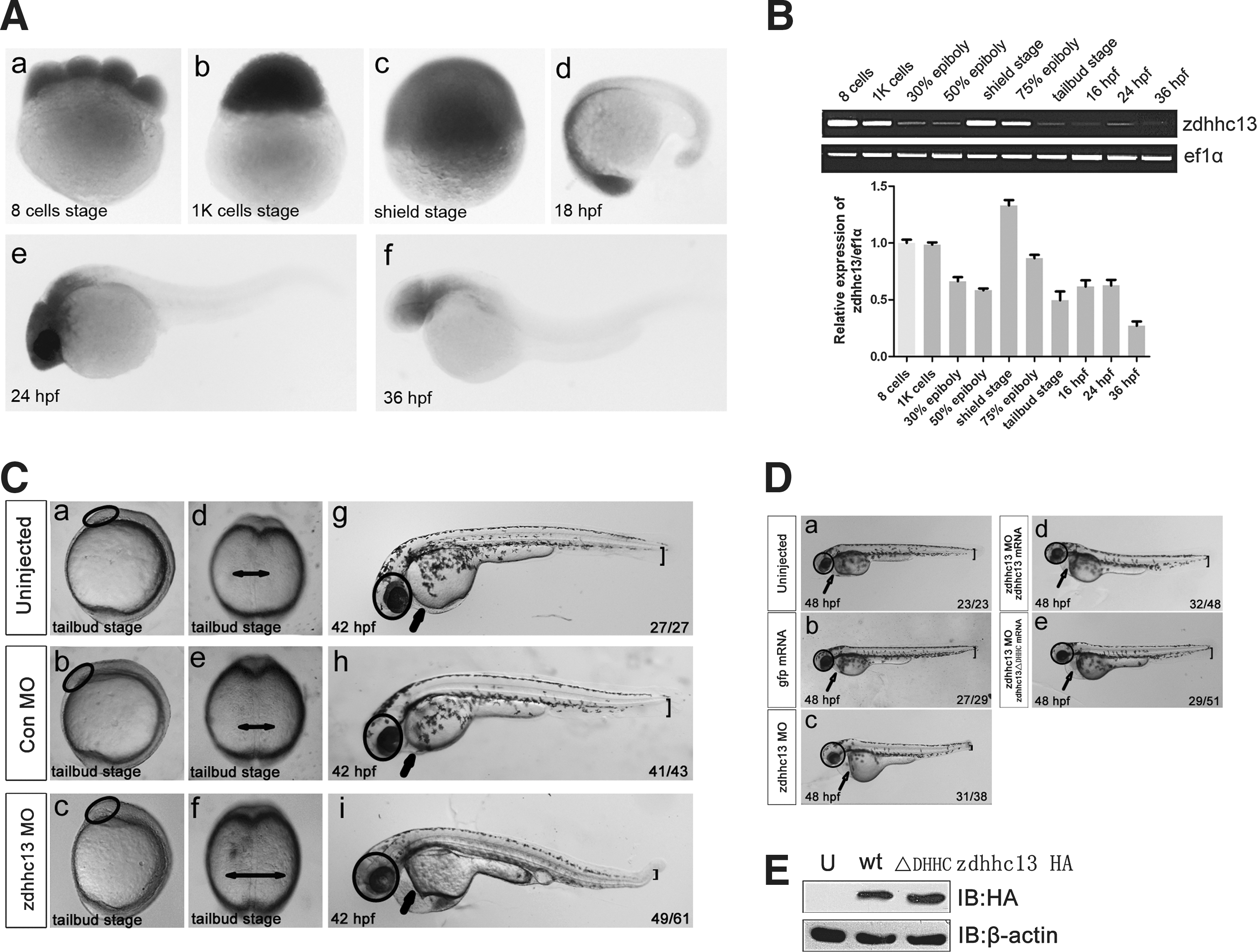
Knockdown of the zinc finger DHHC-type containing gene (zdhhc13) results in morphogenic defects in zebrafish embryos.
Zdhhc13 is necessary for proper ectoderm and mesoderm specification in zebrafish
To explore the function of Zdhhc13, we investigate the phenotypes of zebrafish embryos injected with morpholino antisense oligonucleotides (MO) that specifically attenuated zdhhc13 translation (zdhhc13 MO). A microinjection of zdhhc13 MO into the zebrafish embryos resulted in a slightly larger head and expanded somites at the tailbud stage. Embryos injected with a Con MO developed normally (Fig. 1Ca–f ). By 42 hpf, the morphogenic defects had become more pronounced, including defects in ectodermal differentiation, such as an enlarged brain, and defects in mesoderm-derived tissues, such as a thinner ventral fin, heart failure, and decreased blood flow (Fig. 1Cg–i and Supplementary Movies S1 and S2). In accordance with these observations, the expression patterns of neural ectoderm (pax2 and krox20) and mesoderm markers (myod1) were significantly altered during late somitogenesis, suggesting that the ectoderm had become neuralized and the mesoderm had become dorsalized in the zebrafish morphants (Supplementary Fig. S2). These defects were specifically attributed to the lack of Zdhhc13, because the expression of mouse zdhhc13, which lacks the MO binding site, partially rescued the MO-induced phenotypes (Fig. 1Da–d). Notably, the rescue effect of zdhhc13 might be independent of Zdhhc13 PAT activity, because zdhhc13 ΔDHHC, which encoded a protein lacking the DHHC domain, also rescued the MO-induced phenotype (Fig. 1De).
To gain further insights into the molecular nature of these patterning defects, we analyzed the expression of several early differentiation markers during germ layer specification (Fig. 2). Interestingly, to different degrees, the expression of dorsal ectoderm and mesoderm markers expanded into lateral domains, as indicated by the expanded expression domains of SRY-box containing gene 3 (sox3) and distal-less homeobox 3b (dlx3b; neural ectoderm markers), and chordin (chd; dorsal mesoderm marker) (Fig. 2A, D). In contrast, the ventral markers showed more restricted expression domains in zdhhc13 morphants compared with controls (Fig. 2B, D), including that of forkhead box i1 ( foxi1; non-neural ectoderm marker) and even-skipped-like 1 (eve1; ventral mesoderm progenitor cell marker). Based on these results, we conclude that Zdhhc13 plays a crucial role in the control of germ layer specification. Specifically, Zdhhc13 is required for cell lineage homeostasis of the ectoderm and mesoderm in zebrafish embryos.
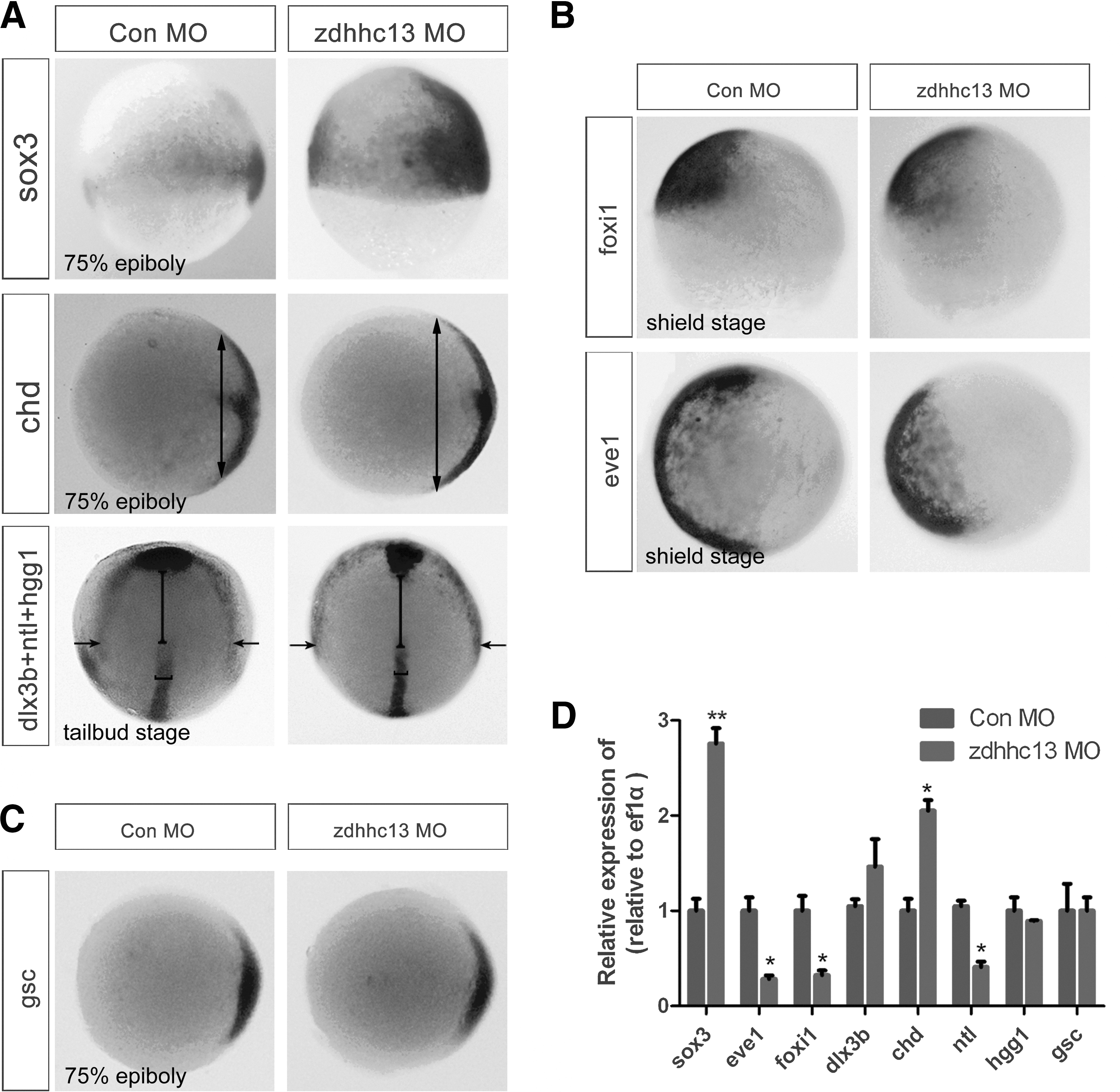
Zdhhc13 is necessary for proper specification of the ectoderm and mesoderm embryonic germ layers.
Zdhhc13 positively modulates BMP signaling
In vertebrate development, BMPs govern the differentiation balance of the germ layers and play a crucial role in fate specification of the ectoderm and mesoderm cell lineages [7,8]. Therefore, we first tested whether zdhhc13 knockdown interfered with the activation of a major mediator of BMP signaling, phosphorylated Smad1/5/8 (p-Smad1/5/8), in zebrafish embryos. This analysis showed that the level of p-Smad1/5/8 was strongly reduced in zdhhc13 morphants compared with controls, suggesting that Zdhhc13 positively modulated Smad-mediated BMP signaling during early lineage decisions (Fig. 3A, B). Attesting to its specificity, Zdhhc13 exhibited little effect on the expression of boz, a direct target of Wnt signaling (Fig. 3C), and hgg1 and gsc, direct targets of Activin/Nodal signaling (Figs. 2C, 2D, and 3C).
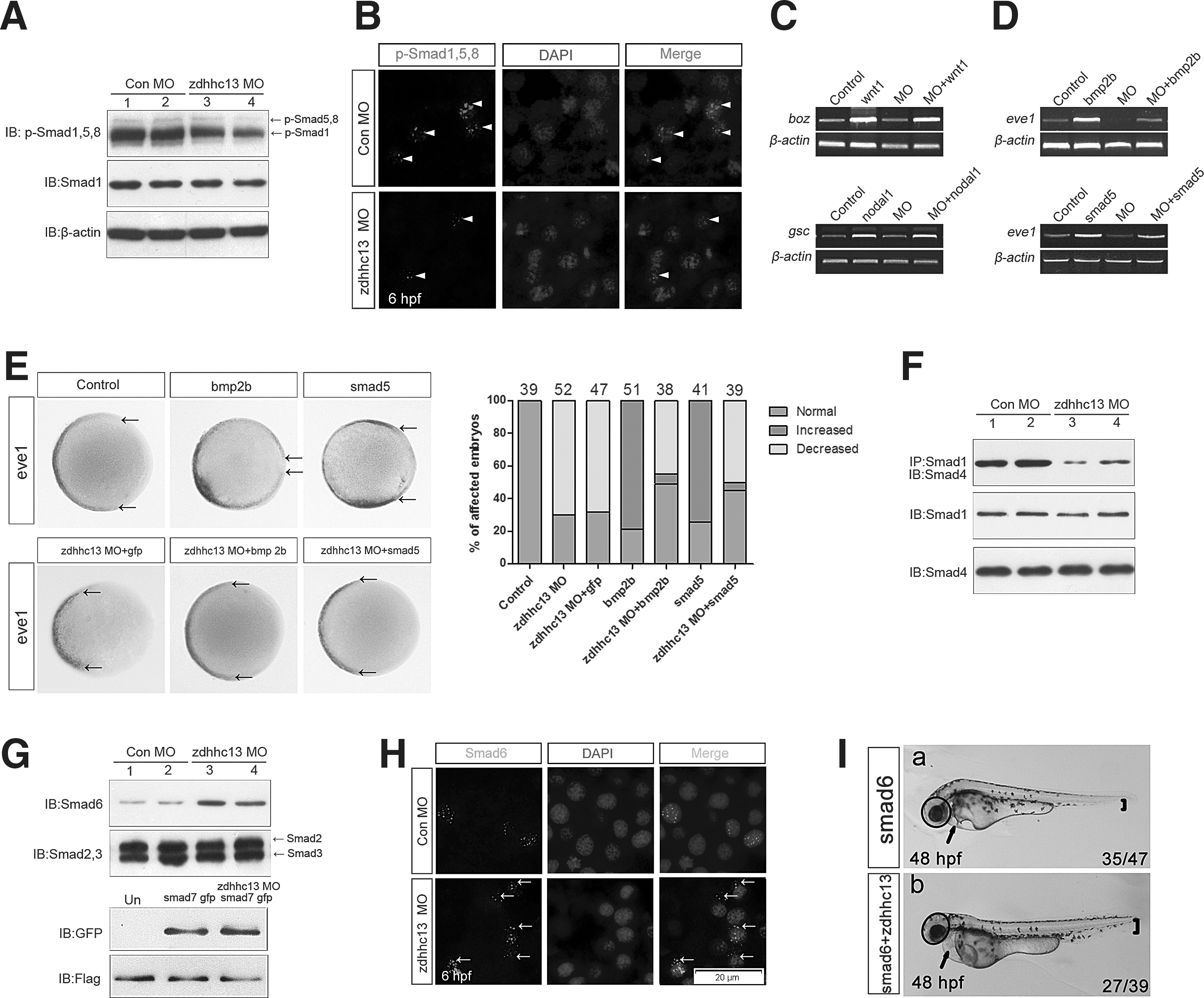
Genetic interactions between zdhhc13 and components of the BMP signaling pathway.
Next, we examined the manner in which Zdhhc13 functionally modulated BMP signaling during early germ-layer specification. bmp2b (25 pg) overexpression activated the BMP pathway and expanded the ventral marker eve1, a target of BMP signaling. A coinjection of zdhhc13 MO and bmp2b mRNA showed a rescue effect at 75% epiboly (Fig. 3D, E). We further tested the functional interaction between Zdhhc13 and Smad5, an intracellular receptor-regulated Smad (R-Smad) essential for BMP signaling in zebrafish embryos. Smad5 mRNA (80 pg)-induced eve1 expression was largely antagonized by a coinjection of zdhhc13 MO (Fig. 3D, E). This specific functional interaction between Zdhhc13 and BMP signaling indicated that Zdhhc13 was likely acted on or downstream of Smad5.
We found that BMP-induced Smad1/5/8-Smad4 complex formation was dramatically inhibited in zdhhc13 morphants (Fig. 3F). Once BMP signaling is activated, phosphorylated R-Smads bind to Smad4 and translocate to the nucleus. Inhibitory Smad proteins bind to Smad1 and interfere with complex formation between Smad1 and Smad4, resulting in the inhibition of BMP signaling [24 –27]. Therefore, we investigated whether Zdhhc13 affected these downstream BMP-signaling components. Surprisingly, we observed a striking increase in the level of Smad6 during germ-layer specification in zdhhc13 morphants compared with control embryos (Fig. 3G). Consistent with these findings, we found a change in the subcellular localization of Smad6 by using immunofluorescent staining (Fig. 3H). We detected relatively high levels of nuclear Smad6 in the control embryos; however, Smad6 was observed in the nucleus as well as in the cytoplasm of zdhhc13 morphants. The functional interaction between Zdhhc13 and Smad6 appeared to be specific, because knockdown of zdhhc13 had no effect on other downstream BMP signaling components (Fig. 3G). To further interrogate the effect of Zdhhc13 on Smad6, we compared zdhhc13 morphants with embryos ectopically expressing Smad6 and found that Smad6-induced embryos had enlarged heads and reduced ventral tissues, similar to Zdhhc13-deficient embryos (Fig. 3Ia). The defects induced by ectopic Smad6 were partially rescued by Zdhhc13 gain of function (Fig. 3Ib). Thus, these observations prompted us to speculate that Zdhhc13 supports BMP signaling by regulating the activity of Smad6 during early fate determination of the germ-layer lineages.
Zdhhc13 regulates ectoderm and mesoderm fate in our in vitro cell model
To gain insights into the signal transduction pathways regulated by Zdhhc13 and to demonstrate that zdhhc13 knockdown and its associated BMP signaling defects caused the loss-of-function phenotype, we treated P19 cells with zdhhc13-specific short interfering RNAs (siRNA). RA-induced P19 cell monolayers exhibit embryonic stem cell-like pluripotency and spontaneously differentiate into cells resembling the three germ layers [3,28,29]. During the differentiation of the P19 monolayers, RT-PCR analysis showed that zdhhc13 mRNA was basally expressed in noninduced P19 cells. zdhhc13 expression was significantly increased within the first 2 days of RA induction and decreased during the differentiation stage (Supplementary Fig. S3A). Since no change in zdhhc13 expression was observed in other cell lines after RA treatment (Supplementary Fig. S3B), zdhhc13 up-regulation might be considered a PSC-specific event and could be involved in the differentiation of monolayered P19 cells.
Next, we investigated the role of Zdhhc13 in the differentiation of RA-induced P19 cells and more closely analyzed differentiation markers in control and zdhhc13 siRNA-treated cells by using immunostaining and western blotting. The ectoderm and mesoderm cell lineages were markedly altered in zdhhc13-deficient cells, as shown by the dramatic increase in Sox2 (neuroectodermal marker), Nestin (neural stem cell marker) (Supplementary Fig. S3D and Fig. 4A–C), α-SMA (smooth muscle cell marker), and Pax3 (skeletal muscle progenitor cells marker) (Supplementary Fig. S3D and Fig. 4D–F). However, no difference in the endoderm cell lineage was observed; both Gata6 and Sox17 showed the same expression profile in differentiated zdhhc13 siRNA-treated cells as in control cells (Supplementary Fig. S3D and Fig. 4G–I). The expression of the three germ-layer markers also supported these findings (Supplementary Fig. S3E). Together, these results suggested that Zdhhc13 governs the fate of cells within our in vitro model system, particularly the ectoderm and mesoderm cell lineages. These results are in accordance with those attained for zdhhc13 morphant embryos.
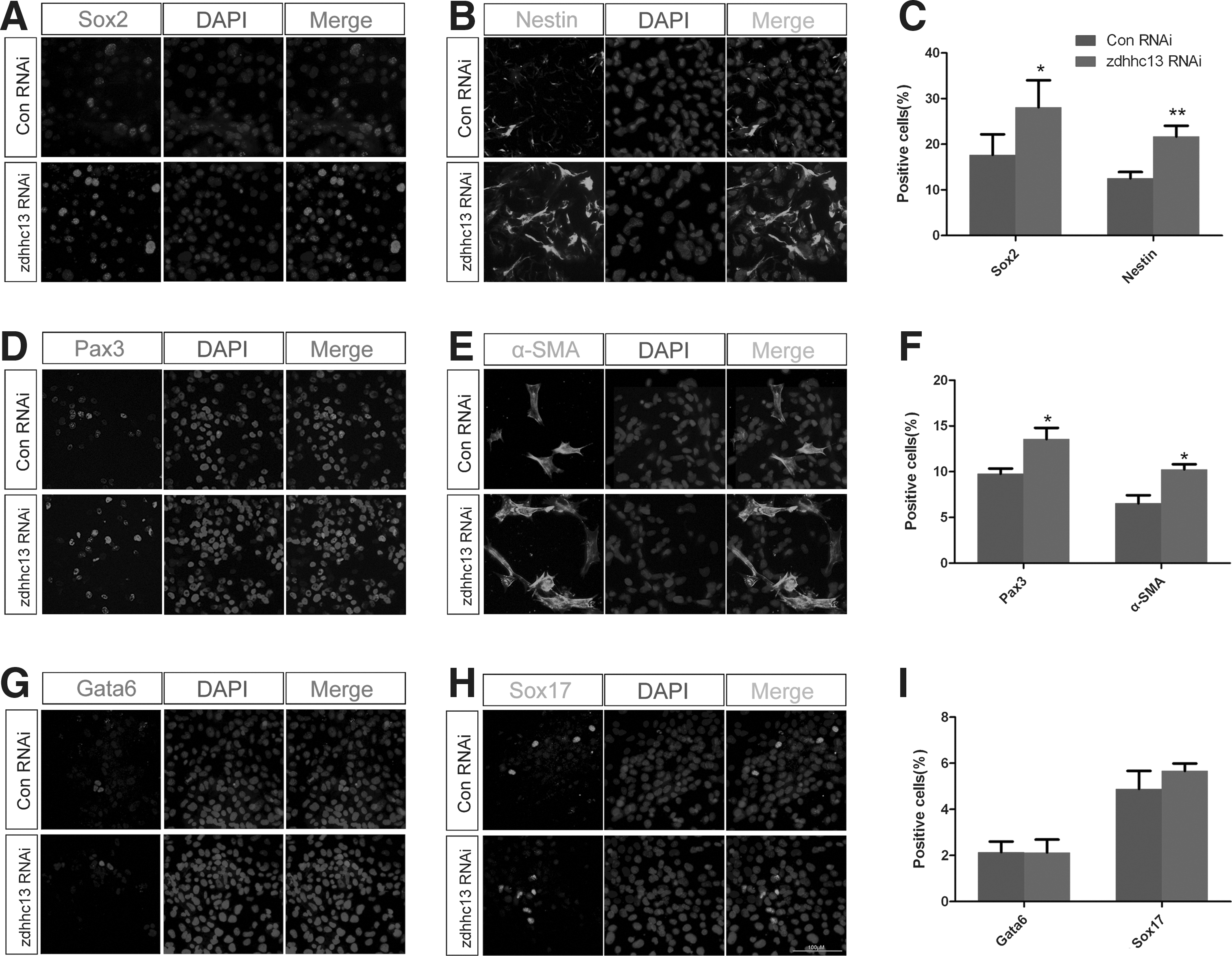
Zdhhc13 regulates fate specification of the ectoderm and mesoderm in vitro.
Zdhhc13 interacts with Smad6 and regulates its biological activity
To further elucidate the function of Zdhhc13 and the mechanism by which it regulates BMP signaling, we compared the levels of Smad1 and p-Smad1/5/8 in control and zdhhc13-deficient cells during the differentiation process. Results showed that endogenous Smad1 predominantly accumulated in the cytoplasm and less so in the nuclei of zdhhc13-deficient cells (Supplementary Fig. S4A). p-Smad1/5/8 immunostaining showed that activated Smad1/5/8 was present in control cells, but could hardly be detected in zdhhc13 siRNA-treated cells (Supplementary Fig. S4B). These findings were similar to those of P19 cells that ectopically expressed Smad6 [28].
We examined how Zdhhc13 affected the biological activity of Smad6. First, we focused on the subcellular localization of Zdhhc13 and Smad6 proteins. As shown in Fig. 5Ab, endogenous Smad6 was observed in the nuclei as well as in the cytoplasm. However, cells transfected with zdhhc13 (HA-tagged) constructs revealed a high degree of colocalization in the perinuclear compartment (Fig. 5Ac). This strong colocalization prompted us to question whether Zdhhc13 and Smad6 physically interacted. Using coimmunoprecipitation, we found that Smad6 associated with Zdhhc13, but not with other similar DHHC-containing proteins, Zdhhc8 and 17 (Fig. 6A). We further analyzed the interaction by constructing a series of zdhhc13 deletion mutants. These results demonstrated that a 60–300-residue region located within the ankyrin domains of Zdhhc13 was responsible for its specific interaction with Smad6 (Fig. 6B–D). This interaction with Smad6 was independent of the PAT activity of Zdhhc13, because both wild type and ΔDHHC Zdhhc13 immunoprecipitated Smad6 (Fig. 6B–D). These data were in agreement with our immunofluorescent staining analysis. Strong colocalization was observed between Zdhhc13 ΔDHHC and Smad6 around the nucleus, but weak colocalization was observed between Smad6 and mutant Zdhhc13 lacking residues 60–300 (Fig. 5Ba, Bb, C).

Distribution of Smad6 and Zdhhc13, Zdhhc13 ΔDHHC, or zdhhc13 ΔANK protein in P19 cells. Exogenous Zdhhc13 was primarily disturbed around the nucleus

Identification of Zdhhc13 as a Smad6-interacting protein.
Next, we assessed the protein levels of Smad6 in the in vitro cell system during differentiation. We found that the levels of Smad6 were up-regulated after induction of differentiation, negatively correlating with Zdhhc13 (Fig. 6E). To further investigate whether the accumulation of Smad6 was impaired by Zdhhc13, we assessed the protein levels of Smad6 in zdhhc13-overexpressing cells. Smad6 was cotransfected with different amounts of the zdhhc13 expression construct. Notably, Smad6 protein was down-regulated by zdhhc13 expression in a dose-dependent manner (68% decrease) (Fig. 6F). Overexpression of zdhhc13 did not have an obvious effect on the transcript levels of smad6 (Fig. 6G). Thus, we speculated that Zdhhc13 regulated Smad6 ubiquitination and proteasomal degradation. To test our hypothesis, we examined the ubiquitination of Smad6 after co-expression with zdhhc13. We found that Zdhhc13 induced Smad6 monoubiquitination in a dose-dependent manner (Fig. 6H and Supplementary Fig. S5). As expected, when cells were treated with a proteasome inhibitor (MG132), Smad6 levels were no longer altered by zdhhc13 expression (Fig. 6I).
Taken together, these results suggest that Zdhhc13 supports BMP/Smad signaling during differentiation of the germ layers. Zdhhc13 acts as an adaptor to recruit Smad6 to the perinuclear compartment where Smad6 is readily degraded in a proteasome-dependent fashion, thus regulating its biological activity.
Discussion
Our in vitro and in vivo data provided compelling evidence that Zdhhc13 is an important regulator of embryonic cell fate specification. Here, we investigated the embryonic function of Zdhhc13, a protein that is evolutionarily conserved and specifically expressed during early cell lineage decisions. We found that Smad6 accumulated in zdhhc13-deficient cells and in zebrafish embryos, attenuating BMP-mediated repression of neural induction and causing an imbalance in BMP-induced mesoderm formation. Therefore, our findings establish the role of DHHC proteins during germ-layer specification, contributing to our understanding of the disease processes associated with the dysfunction of DHHC proteins.
A previous study reported that zdhhc13 deficiency causes severe systemic phenotypes, including failure to thrive, osteoporosis, multi-organ/system dysfunction, and early death [18]. However, the molecular targets and precise function of Zdhhc13 during embryonic cell fate determination remained unexplored. Systematic knockdown of zdhhc genes has not been performed in vivo, which could identify unexpected physiological roles for DHHC proteins. Using loss-of-function experiments, we showed that Zdhhc13 controls one of the earliest differentiation events, specification of the definitive germ layers. The effects of Zdhhc13 knockdown in zebrafish embryos (Fig. 1C) resembled those observed in vitro in zdhhc13-deficient cells (Fig. 4), reflecting a conserved embryonic function. Indeed, zdhhc13 up-regulation is along with the early specification of ectoderm and mesoderm cell lineage (Fig. 1B and Supplementary Fig. S3A). Therefore, we propose that the increase in zdhhc13 expression in zebrafish embryos and our in vitro cell model during early differentiation stages promoted the transition toward ectoderm and mesoderm cell lineages.
The function of DHHC proteins as PATs has been recently described, but other roles of DHHC proteins are just beginning to be appreciated [30 –32]. Here, we report that Zdhhc13 possesses a novel function which appears to be unique in the PAT family. Since mouse zdhhc13 ΔDHHC partially rescued the MO-induced defects, our data suggest that Zdhhc13 positively regulates ectoderm and mesoderm formation in zebrafish embryos through a mechanism independent of its PAT activity (Fig. 1D). This embryological role was conserved in our in vitro cell model. Importantly, Zdhhc13 is the only PAT that contains a DQHC motif in place of the canonical DHHC, and the zebrafish Zdhhc13 sequence has serine in place of the cysteine in the DHHC motif (DHHS) [33]. Whether these changes in the DHHC motif affect the catalytic activity of Zdhhc13 is a matter of debate, because mutagenesis of these residues in Zdhhc2, 3, and 17 results in a loss of palmitoylation activity in vitro [33 –36]. Zdhhc13 may have evolved to serve a specialized function. The different functions of Zdhhc13 might be systematically connected, and directly or indirectly contribute to the pathogenesis of these diseases. Based on these findings, we propose that the main role of zdhhc13 in preventing these diverse pathologies is independent of its PAT activity.
Here, we uncovered that Zdhhc13 antagonized Smad6 during BMP-dependent signaling and differentiation events in an in vitro cell model and zebrafish embryos. Although the DHHC family of proteins has an important role in BMP pathway activation [37], the precise molecular mechanisms of their activity are not clear. In this study, we found that Zdhhc13 acts as an adaptor and recruits Smad6 to the perinuclear compartment where Smad6 is monoubiquitinated, reducing its mobility and increasing its degradation (Figs. 5Ac and 6H). These data appear to contradict the notion that intracellular trafficking and nuclear import of Smad6 are required for BMP signaling. Ubiquitin-conjugating enzyme E2O (UBE2O), an E2 ubiquitin-conjugating enzyme, potentially mediates Zdhhc13-induced Smad6 degradation in the cytoplasm during ectoderm and mesoderm formation. UBE2O amplifies BMP7 signaling through Smad6 monoubiquitination during adipocyte differentiation [38]. Similar to our observations, previous studies have reported that the E3 ubiquitin-protein ligase SMURF1 interacts with Smad6 to induce its nuclear export and enhance BMP-induced transcription in the nucleus [39]. Therefore, these data support the hypothesis that the subcellular localization of Smad6 might be the major determinant of the differential effects of Zdhhc13 on BMP signaling.
Members of the TGF-β family such as Activin/Nodal and BMPs are of key importance during germ-layer specification [40,41]. Our data indicated that Zdhhc13 specifically amplified BMP/Smad signaling responses; however, Activin/Nodal target genes, sox17 and gsc (Figs. 2C and 4G–I) [42 –44], were not altered by Zdhhc13 in the in vitro cell model or in zebrafish embryos. This biological activity of Smad6 is similar to that in mammals [25], but distinct from that in Xenopus, where Smad6 antagonizes signaling downstream of at least two TGF-β family ligands, BMP and Activin/Nodal [45]. These data indicate that the negative regulatory role of Smad6 in TGF-β signaling has evolved to serve a more specialized function. We speculate that this specific effect on the BMP pathway is related to the biological function of Zdhhc13. In fact, our analysis showed that zdhhc13 was a downstream gene target of BMP signaling (Supplementary Fig. S6A–D). In a positive autoregulatory feedback loop, BMP signaling up-regulated zdhhc13 expression through a Smad1-induced transcriptional response. Thus, any defects in BMP levels would be amplified because of disrupting this loop during embryonic cell fate specification. Since increased BMP-dependent zdhhc13 expression modulated Smad6 protein stability and Smad1-induced gene expression, BMP-induced zdhhc13 expression should be involved in more than just a positive feedback loop in BMP signaling.
In summary, we report that Zdhhc13 possesses a novel function that is independent of its PAT activity. Zdhhc13 acts as an adaptor to recruit Smad6, leading to its reduced mobility and increased degradation (Supplementary Fig. S6E). Our results expanded the cellular function of Zdhhc13, suggesting it is a regulator of BMP signaling, and established the fact that Zdhhc13 plays a critical role in the early specification of the germ layers. It is beneficial to resolve the disturbances in the regulation of DHHC family and to identify the disease processes caused by the dysfunction of DHHC proteins so that DHHC protein can be used as a potential therapeutic target.
Footnotes
Acknowledgments
This work was supported by funding from the National Natural Science Foundation of China (no. 81071057 and 31271289), the Ministry of Education of China (no. 20110131110036), the Natural Science Foundation of Shandong Province (no. 2012GSF11842 and ZR2010HQ022), and Shandong University (no. 2012JC006).
Author Disclosure Statement
The authors indicate no potential conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.