
Abstract
The precise mechanisms underlying the differential function and cardiometabolic risk of white adipose tissue (WAT) remain unclear. Visceral adipose tissue (VWAT) and subcutaneous adipose tissue (SCWAT) have different metabolic functions that seem to be ascribed to their different intrinsic expansion capacities. Here we have hypothesized that the WAT characteristics are determined by the resident adipose-derived stem cells (ASCs) found in the different WAT depots. Therefore, our objective has been to investigate adipogenesis in anatomically distinct fat depots. ASCs from five different WAT depots were characterized in both healthy lean and diabetic obese rats, showing significant differences in expression of some of genes governing the stemness and the earlier adipogenic differentiation steps. Notch-target genes [Hes (hairy and enhancer of split) and Hey (hairy/enhancer of split related with YRPW motif) families] were upregulated in ASCs derived from visceral depots. Upon adipogenic differentiation, adipocyte cell markers were downregulated in ASCs from VWAT in comparison to ASCs from SCWAT, revealing a lower adipogenic capacity in ASCs of visceral origin than in those of SCWAT in accordance with the differential activation of Notch signaling. Notch upregulation by its activator phenethyl isothiocyanate attenuated the adipogenic differentiation of ASCs from SCWAT whereas Notch inhibition by N-[N-(3,5-difluorophenacetyl-L-alanyl)]-S-phenylglycine t-butyl ester (DAPT) increased the adipogenic differentiation of ASCs from visceral origin. In conclusion, the differential activation of Notch in ASCs is the origin of the different intrinsic WAT expansion capacities that contribute to the regional variations in WAT homeostasis and to its associated cardiometabolic risk.
Introduction
O
The source and conditioning processes responsible for the metabolic differences between SCWAT and VWAT remain unclear. It has been suggested that the proximity of VWAT to the portal vein may allow VWAT to exert more direct metabolic effects on the liver [12]. Other evidences in accordance with the adipose tissue expandability hypothesis [13] support the concept that VWAT and SCWAT have different intrinsic adipose tissue expansion capacities [14]. Understanding differential regulation of resident stem cell differentiation and adipocyte biogenesis and function in anatomically distinct fat depots may be critical to prevent the metabolic comorbidities of visceral obesity.
For many years WAT has been considered as a passive-energy-reserve organ. However, this concept has changed in recent years. WAT is considered a dynamic endocrine organ playing a central role in homeostasis [15]. WAT is also a source of pluripotent cells, adipose-derived stem cells (ASCs), which have been considered an alternative to bone marrow cells for cell-based therapies and tissue engineering [16,17], and have opened new avenues to investigate the mechanisms involved in WAT function.
The generation of mature fat cells from ASCs, or adipogenesis, consists of two related steps: the determination of ASCs into preadipocytes and the differentiation of preadipocytes into mature fat cells [18]. Most of our knowledge about adipogenesis refers to terminal differentiation because most studies have used cell lines that have a restricted potential to differentiate into other cell types [19], limiting the information about the mechanisms that govern earlier differentiation steps and offering little data on depot-specific aspects of differentiation. The molecular pathways implicated in the commitment of stem cells to the adipocyte lineage are incompletely defined. Notch signaling that plays a critical role in controlling stem cell fate, balancing differentiation and proliferation of progenitor cells [20 –24], seems to be one of them. Notch activation leads to the maintenance of self-renewal potential through the downregulation of tissue-specific transcription factors and its inhibition induces terminal differentiation in proliferating cells [25,26]. Notch signaling has been shown to be involved in adipocyte differentiation from mesenchymal stem cells (MSCs). However, the precise role of Notch signaling remains controversial [27 –36].
Thus, the present study investigates how inherent characteristics of resident ASCs could contribute to the regional variation in fat tissue homeostasis. To achieve this goal, the ASCs from different WAT depots were investigated to identify their stemness and their adipogenic differentiation commitment.
Materials and Methods
Animals
Experiments were carried out in male Zucker diabetic fatty (ZDF; Leprfa/fa) rats, and their lean male littermates (ZLC; Leprfa/+ or Lepr+/+) obtained from Charles River Laboratories, Inc. All procedures followed in the study fulfilled the criteria of the Guide for the Care and Use of Laboratory Animals published by the U.S. National Institutes of Health (NIH Publication No. 85–23, revised 1996) and were approved by the Internal Animal Committee Review Board. For further details, see Supplementary Methods (Animals heading) (Supplementary Data are available online at
Serum determinations
Blood was collected in serum-separation tubes, centrifuged, and stored at −80°C to perform further analysis. Biochemical analyses were determined with a CLIMA MC-15 analyzer (RAL). Insulin levels were determined by enzyme immunoassay (Rat insulin EIA kit A05105; Spibio), according to the manufacturer's instructions.
Isolation and culture of ASCs
WATs [epicardial (ECWAT), epididymal (EDWAT), mesenteric (MEWAT), retroperitoneal (RPWAT), and SCWAT] were excised and processed to obtain ASCs as previously described technique [37] and it is detailed in Supplementary Methods (Isolation and culture of ASCs heading).
Flow cytometric analysis
Cultured ASCs were characterized by flow cytometry. A detailed description of the protocol and antibodies used is available in Supplementary Methods (Flow cytometric analysis heading).
In vitro ASC proliferation assay
Cell proliferation was evaluated by using a nonisotopic immunoassay for the quantification of BrdU incorporation into newly synthesized DNA of actively proliferating cells (Calbiochem® BrdU cell proliferation assay; Calbiochem), according to the manufacturer's instructions. ASCs were plated on 96-well culture plates at a density of 5×103 and proliferation was quantified by BrdU uptake 24 h after initial plating.
Gene expression analysis
Total RNA was isolated from ASCs using RNeasy mini isolation kit (Qiagen) according to the manufacturer's protocol. cDNA was synthesized from 1 μg total RNA using High Capacity cDNA Archive kit (Applied Biosystems) and analyzed by real-time polymerase chain reaction (PCR) using TaqMan gene expression assays (Applied Biosystems) and the Applied Biosystems Prism 7900HT Sequence Detection System (Applied Biosystems) according to manufacturer's instructions. Relative gene expression values were calculated by the ΔΔCt method. The raw gene expression values were normalized according to the expression of ribosomal protein, large, P0 (Rplp0). TaqMan gene expression assays used are provided in Supplementary Table S1.
Activation of Notch signaling pathway by phenethyl isothiocyanate treatment
ASCs were allowed to reach confluence and pretreated with 5 μM phenethyl isothiocyanate (PEITC; Sigma-Aldrich) prepared in dimethyl sulfoxide (DMSO). An equal volume of DMSO was added to controls. Eight hours or 4 days after treatment, ASCs were induced to differentiate to adipocytes according to the procedure described in Adipogenic differentiation section. After treatment, to assess the response to PEITC, RNA was extracted and the gene expression of the members of Notch pathway was measured. Finally, we evaluated by the gene expression of adipocyte-related markers whether Notch activation via PEITC would induce changes in adipogenic differentiation of ASCs.
Inhibition of Notch signaling pathway by N-[N-(3,5-difluorophenacetyl-L-alanyl)]-S-phenylglycine t-butyl ester treatment
ASCs were allowed to reach confluence and pretreated with 50 μM N-[N-(3,5-difluorophenacetyl-L-alanyl)]-S-phenylglycine t-butyl ester (DAPT, Calbiochem No. 565784; Merk KGaA) for 4 days prior to induction of adipogenic differentiation. ASCs pretreated with DMSO (Sigma-Aldrich) were used as the control group. To assess the response to DAPT treatment, after 4 days, RNA was extracted from treated ASCs, and the expression of the members of Notch pathway was measured. Finally, we evaluated by the gene expression of adipocyte-related markers whether Notch inhibition via DAPT would induce changes in adipogenic differentiation of ASCs.
Adipogenic differentiation
ASCs were seeded at a density of 1.5×104 cells/cm2 and allowed to grow to confluence. Subsequently, cells were allowed to differentiate for 21 days in the adipogenic medium containing 0.5 mM 3-isobutyl-1-methylxanthine, 1 μM dexamethasone, 100 μM indomethacin, and 10 μg/mL insulin. All chemicals were obtained from Sigma-Aldrich. Differentiation was evaluated microscopically by Oil Red O staining and lineage-specific gene expression pattern [Cd36, CCAAT/enhancer binding protein α (C/ebpα), fatty acid binding protein 4 (Fabp4), lipoprotein lipase (Lpl), and peroxisome proliferator activated receptor γ (Pparγ)].
Statistical analyses
All data are presented as mean±SEM. Statistical analyses were conducted using Student's t-test for comparisons of the two groups, and one-way ANOVA with Dunnett's multiple-comparison tests or two-way ANOVA with Bonferroni post-tests analysis. A P<0.05 was considered significant. All statistical analyses were performed using GraphPad Prism software 5 (GraphPad) and Partek Genomics Suite software 6.5 (Partek). Principal component analyses (PCA) were conducted with Partek default settings. PCA procedure was used to represent sample variability, which defines a group of PC, in order to explain the variability existing between the samples.
Results
Metabolic profile
Metabolic profiles of ZDF and ZLC rats are detailed in Supplementary Table S2. ZLC rats showed normal blood glucose levels (92.43±1.99 mg/dL) whereas those of ZDF rats were significantly higher (392.75±11.62 mg/dL, P<0.0001 vs. controls) clearly reflecting their diabetic status. Additionally, insulin levels, total cholesterol, triacylglycerols, urea, and glutamic pyruvic transaminase were significantly higher in ZDF rats than in ZLC animals.
Flow cytometric analysis
Most adherent cells expressed CD29 (over 95%), CD90 (over 93%), and CD44 (over 40%–45%) (Fig. 1) and were negative for hematopoietic marker CD45 (<0.2%). Different fat depots exhibited a differential profile for CD29 and CD90 markers (P=0.0054 and P=0.0042, respectively) while no significant differences between fat depots were observed for CD44 and CD45 markers. It is noteworthy that diabetes was associated with a significant reduction of CD29 marker (P=0.0008) and that the percentage of CD29+ cells in EDWAT and MEWAT differed from SCWAT in the diabetic group.
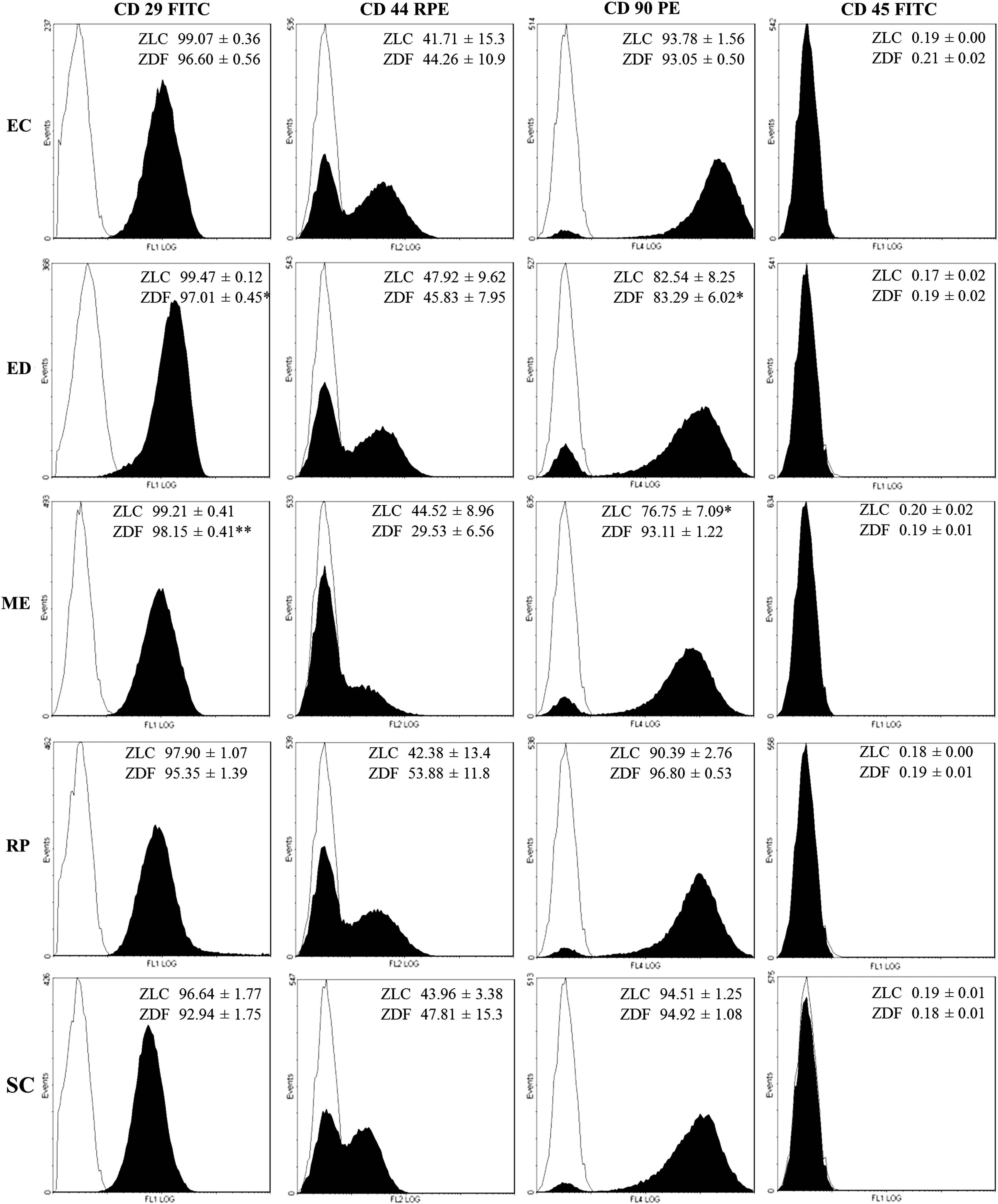
Phenotypic characterization by flow cytometric analysis. Surface markers were analyzed in adipose-derived stem cells (ASCs) after three passages. The data (filled histograms) are the mean±SEM (n=4–5 per group). *P<0.05, **P<0.01 versus subcutaneous adipose tissue (SCWAT) (one-way ANOVA with Dunnett's multiple-comparison tests). Isotype controls were included (open histograms).
ASCs derived from different fat depots exhibit different in vitro proliferation rate
Differences in proliferation rates were observed during in vitro culture of ASCs. Cell proliferation was evaluated by the quantification of BrdU uptake. Results showed that fat depot origin significantly affects ASC proliferation (P<0.0001), whereas no differences between ZLC and ZDF groups were observed. ASCs derived from MEWAT and ECWAT showed a lower proliferation than ASCs derived from SCWAT in both ZLC and ZDF groups (Fig. 2).
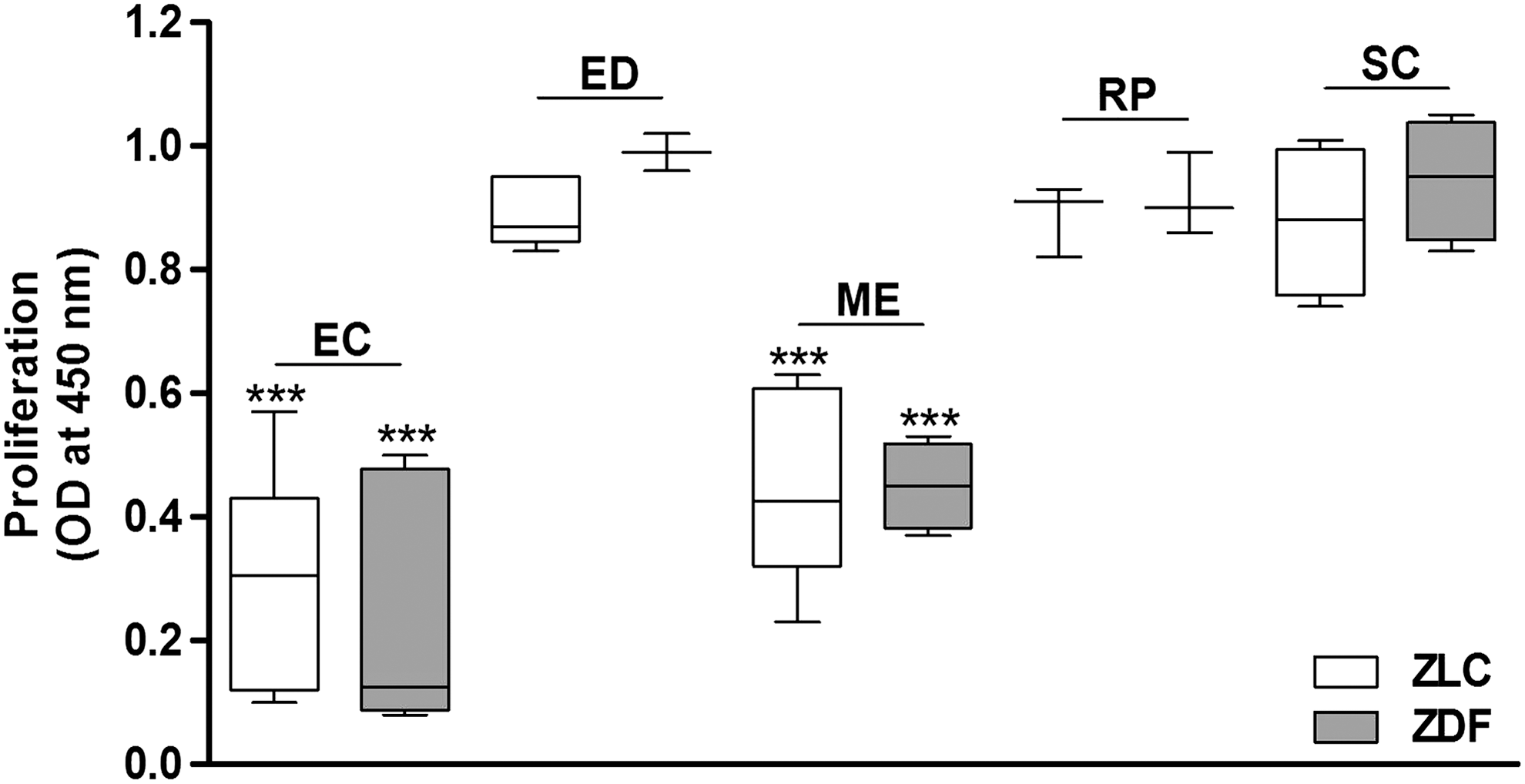
In vitro ASC proliferation. Cell proliferation was evaluated by using a nonisotopic immunoassay for the quantification of BrdU incorporation into newly synthesized DNA of actively proliferating cells and according to the manufacturer's instructions. BrdU uptake was reported as the mean relative OD450. The data are the mean±SEM (n=3–4 per group, in technical duplicates). ***P<0.001 versus SCWAT.
Gene expression profiles and regional variation in fat tissue homeostasis
Expression profiling clearly demonstrated significant differences in gene expression between SCWAT and VWATs in both ZLC and ZDF. PCA showed that the SCWAT transcriptome differed greatly from each other visceral depots in both groups, while ECWAT, EDWAT, MEWAT, and RPWAT depots remained closer (Fig. 3).
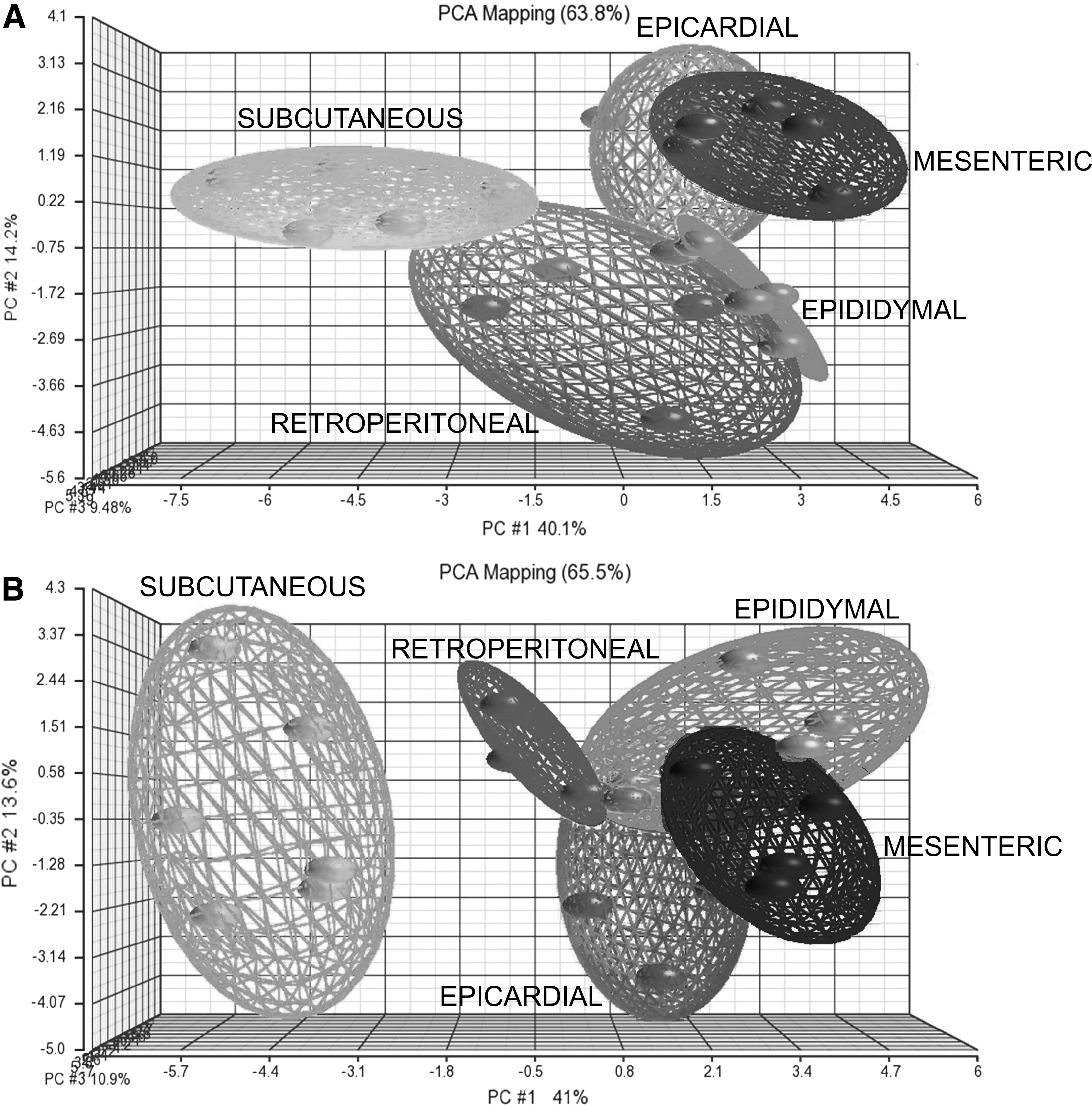
Principal component analysis (PCA). Gene expression analysis was performed using Partek Genomics Suite software 6.5. PCA results indicated a distinct separation between ASCs derived from different adipose tissue depots. The different individuals are visualized as spheres. Closely related individuals are connected through planes.
The gene expression analysis of ASCs for typical panel of surface antigens confirms the mesenchymal stem nature of the cell populations (Table 1). The expression of MSC-specific markers showed significant differences between fat depots for Cd29, Cd73, Cd90, Cd105, and Cd166 in the ZLC group and Cd29, Cd105, and Col1a1 in ZDF.
Presence of stem-cell-specific markers was analyzed in the ASCs derived from different adipose tissues by real-time PCR. Each data was normalized according to the Rplp0 expression level and is presented as gene expression levels relative to reference gene. The data are the mean±SEM (n=4–6, in technical duplicates).
P<0.05, b P<0.01, c P<0.001 versus subcutaneous (one-way ANOVA with Dunnett's multiple-comparison test).
ASCs, adipose-derived stem cells; Col1a1, collagen type α1; Dnmt3b, DNA (cytosine-5-)-methyltransferase 3β; EC, epicardial; ED, epididymal; Fgf2, fibroblast growth factor 2; ME, mesenteric; MSCs, mesenchymal stem cells; PCR, polymerase chain reaction; Podxl, podocalyxin-like protein 1; Pou5f1, POU domain class 5 transcription factor 1; RP, retroperitoneal; SC, subcutaneous; Sox2, SRY (sex determining region Y) box 2; Tert, telomerase reverse transcriptase; ZDF, Zucker diabetic fatty; ZLC, Zucker lean normoglycemic control.
The analysis for genes that define the stemness of ASCs and that maintain their pluripotency and self-renewal characteristics showed a differential expression between fat depots, highlighting that most of the genes analyzed were downregulated in ASCs derived from SCWAT compared with VWATs in both the ZLC and ZDF groups (Table 1). These changes reached significance for fibroblast growth factor 2 (Fgf2), podocalyxin-like protein 1 (Podxl), SRY (sex determining region Y) box 2 (Sox2), and telomerase reverse transcriptase (Tert) in ZLC and DNA (cytosine-5-)-methyltransferase 3β (Dnmt3β), Fgf2, Podxl, POU domain class 5 transcription factor 1 (Pou5f1), and Tert in ZDF rats.
The analysis performed on Notch signaling pathway revealed that ASCs derived from visceral depots were associated with an overall higher expression of the members of this pathway compared with ASCs derived of SCWAT. ASCs derived from ZLC and ZDF rats showed the same pattern (Fig. 4). We observed significant differences between WATs in four Notch receptors. As such there was increase in ASCs from VWATs in the expression of Notch1 (ZLC: P=0.0016; ZDF: P=0.0002), Notch2 (ZLC: P=0.0002; ZDF: P=0.0018), and Notch3 (ZLC: P=0.0006; ZDF: P=0.0017) compared with ASCs from SCWAT. Only Notch4 showed a downregulation in visceral depots versus SCWAT (ZLC: P=0.0241; ZDF: P=0.0375). The analysis of Notch ligands showed a differential expression between WATs and an overexpression of Delta-like (Dll) 3 (ZLC: P=0.0001; ZDF: P=0.0008) and Jagged 1 (Jag1) (ZLC: P<0.0001; ZDF: P=0.0002) in ASCs derived from VWATs compared with SCWAT. However, the expression of Dll1 exhibited a reduction in ASCs derived from visceral depots compared with SCWAT, only reaching significance in the ZLC group (P<0.0001). Expression levels of Dll4 were not detected. Finally, these changes were associated with a widespread upregulation of downstream targets of the Notch signaling in all ASCs derived of VWATs compared with ASCs derived from SCWAT. We observed significant differences between adipose depots for the transcription factors Hes7 (hairy and enhancer of split) (ZLC: P<0.0001; ZDF: P<0.0001), Hey1 (hairy/enhancer of split related with YRPW motif) (ZLC: P<0.0001; ZDF: P=0.0048), Hey2 (ZLC: P=0.0023; ZDF: P=0.0059), and HeyL (ZLC: P<0.0001; ZDF: P=0.0025). Hes1 expression showed significant differences only in the diabetic group (P<0.0001).

Differential gene expression profiles of Notch signaling pathway in ASCs derived from different fat depots. Expression of Notch ligands
Differential adipogenic differentiation capacity of ASCs from different fat depots
Differences in adipogenic differentiation potential between ASCs derived from different fat depots were reflected by Oil Red O staining (Figs. 5A and 6A). The microscopic observations were confirmed by expression levels of the adipogenic-specific markers (Figs. 5B and 6B). After 21 days of differentiation induction, all cell types increased the expression of the adipocyte cell markers. However, ASCs derived from SCWAT showed a higher adipocyte cell marker expression (Supplementary Table S3). This pattern was more accentuated in the ASCs derived from diabetic adipose tissues.
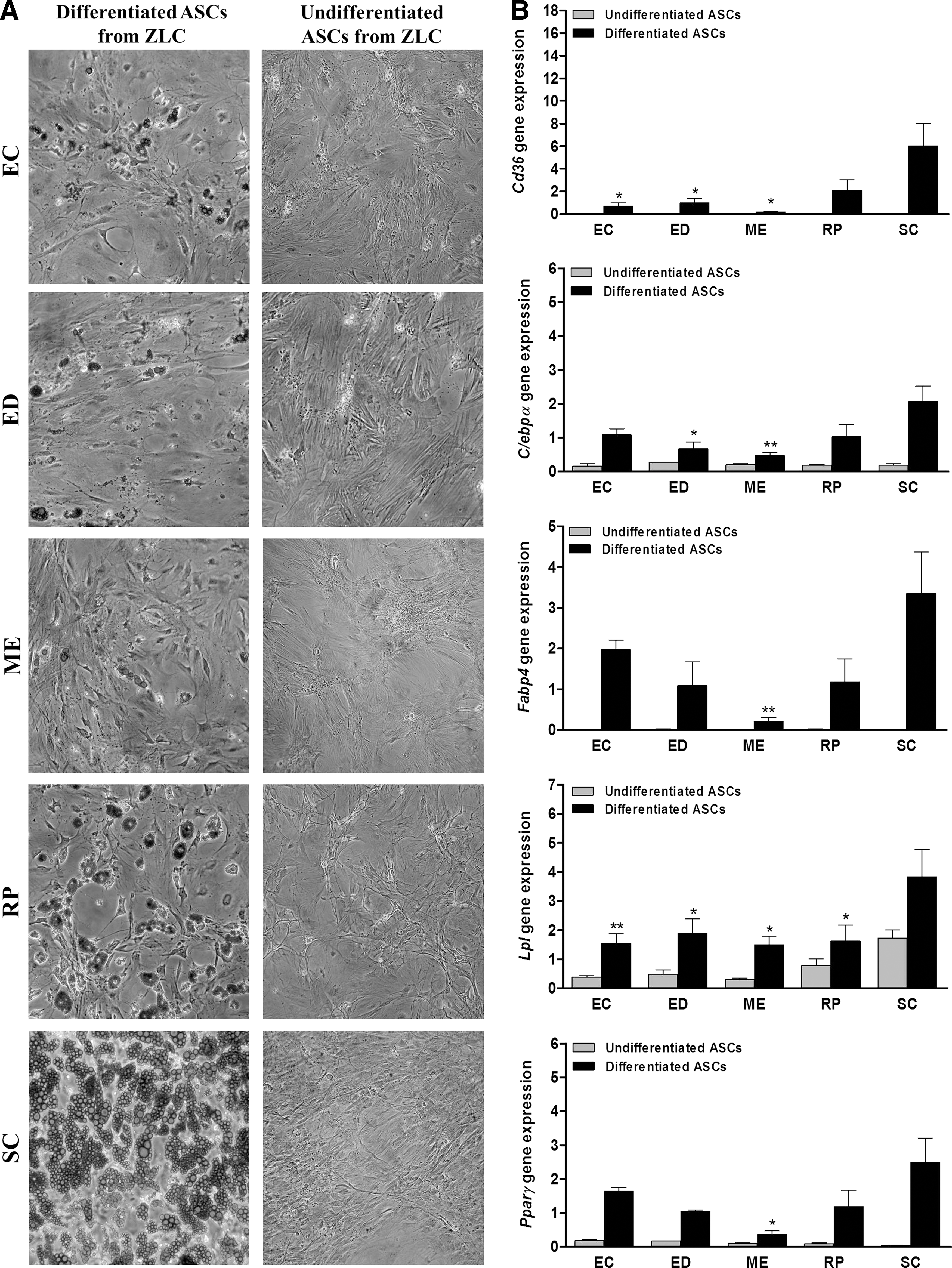
Adipogenic differentiation potential of ASCs from ZLC rats. ASCs from different adipose tissues were induced to differentiate to adipocytes. Adipogenic differentiation potential was evaluated by Oil Red O staining (20× magnification)

Adipogenic differentiation potential of ASCs from ZDF rats. ASCs from different adipose tissues were induced to differentiate to adipocytes. Adipogenic differentiation potential was evaluated by Oil Red O staining (20× magnification)
ASCs derived from MEWAT showed the lower adipogenic potential compared with ASCs derived from SCWAT in ZLC group as well as ZDF group. On the contrary, the adipogenic differentiation capacity of ASCs derived from RPWAT remained closer to SCWAT in both ZLC and ZDF animals.
Notch activation by PEITC attenuates the adipogenic differentiation of ASCs from SCWAT whereas Notch inhibition by DAPT increases the adipogenic differentiation of ASCs from MEWAT
To further confirm the association between the differential activation of Notch system and the differential adipogenic potential observed in ASCs derived from SCWAT and VWATs, ASCs from SCWAT were treated with PEITC, an activator of Notch signaling. PEITC treatment caused a widespread decrease in the level of the expression of the adipocyte cell markers (Fig. 7A) especially at the 4-day time point. Most of the adipogenic markers only showed significant increase in DMSO-treated control compared with undifferentiated cells. Consistent with our hypothesis, these changes were associated with an induction on mRNA levels of the receptor Notch1 (Fig. 7C) and the downstream gene Hes1, at both time points, and the downstream gene Hes7 at the 8-h time point compared with DMSO-treated control (Fig. 7D). The remaining members of Notch pathway analyzed did not show significant changes. The same treatment was performed on ASCs derived from MEWAT showing resistant to change. No significant changes were observed in the expression of the adipocyte markers nor, consistently, in the Notch pathway components (Supplementary Fig. S1).
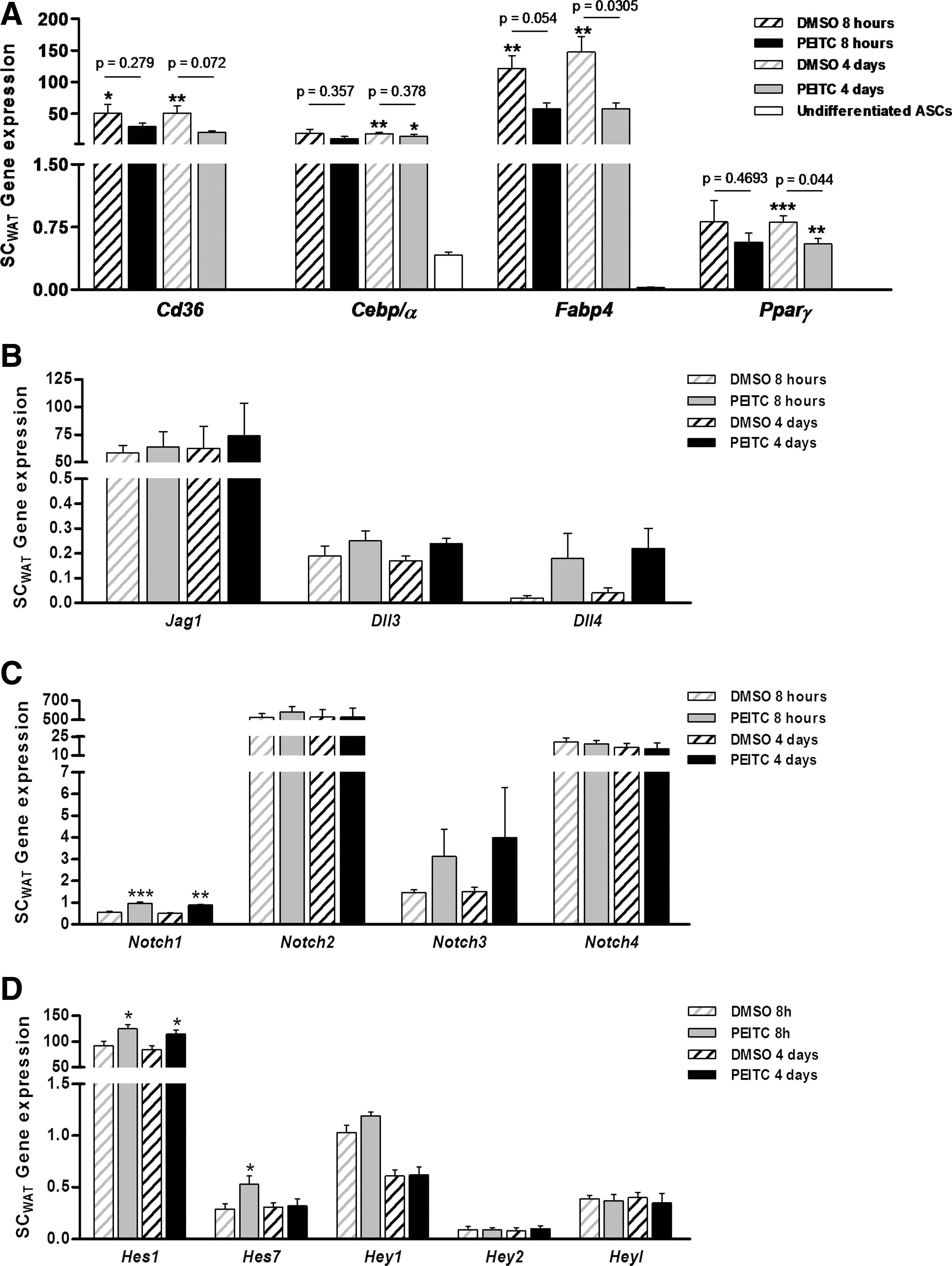
Notch activation in ASCs from SCWAT by phenethyl isothiocyanate (PEITC). ASCs from SCWAT were pretreated with 5 μM PEITC and induced to differentiate to adipocytes. Adipogenic differentiation potential was evaluated by the gene expression of adipocyte molecular markers
On the other hand, ASCs derived from MEWAT were treated with DAPT, an inhibitor of Notch system. DAPT treatment enhanced the adipogenic differentiation capacity of ASCs from MEWAT, showing a marked increase in the mRNA expression of the adipogenic marker compared with undifferentiated ASCs, and significantly different for Cd36 and Cebp/α versus DMSO-treated control (Fig. 8A). These changes were associated to the effect on Notch signaling again. Expression of Dll3 was decreased and Dll4 was increased significantly upon treatment with DAPT (Fig. 8B), possibly to offset the inhibitory effect on Notch system. ASCs treated with DAPT exhibited significant inhibition of Notch1, Notch3, and Notch4 mRNA expression (Fig. 8C) and strong inhibitory effect on transcription levels of all Notch target genes (Fig. 8D). The same treatment was conducted on ASCs derived from SCWAT; however, no change was observed in the adipogenic potential. This lack of response, as expected, was associated with no changes in the Notch signaling pathway (Supplementary Fig. S2).
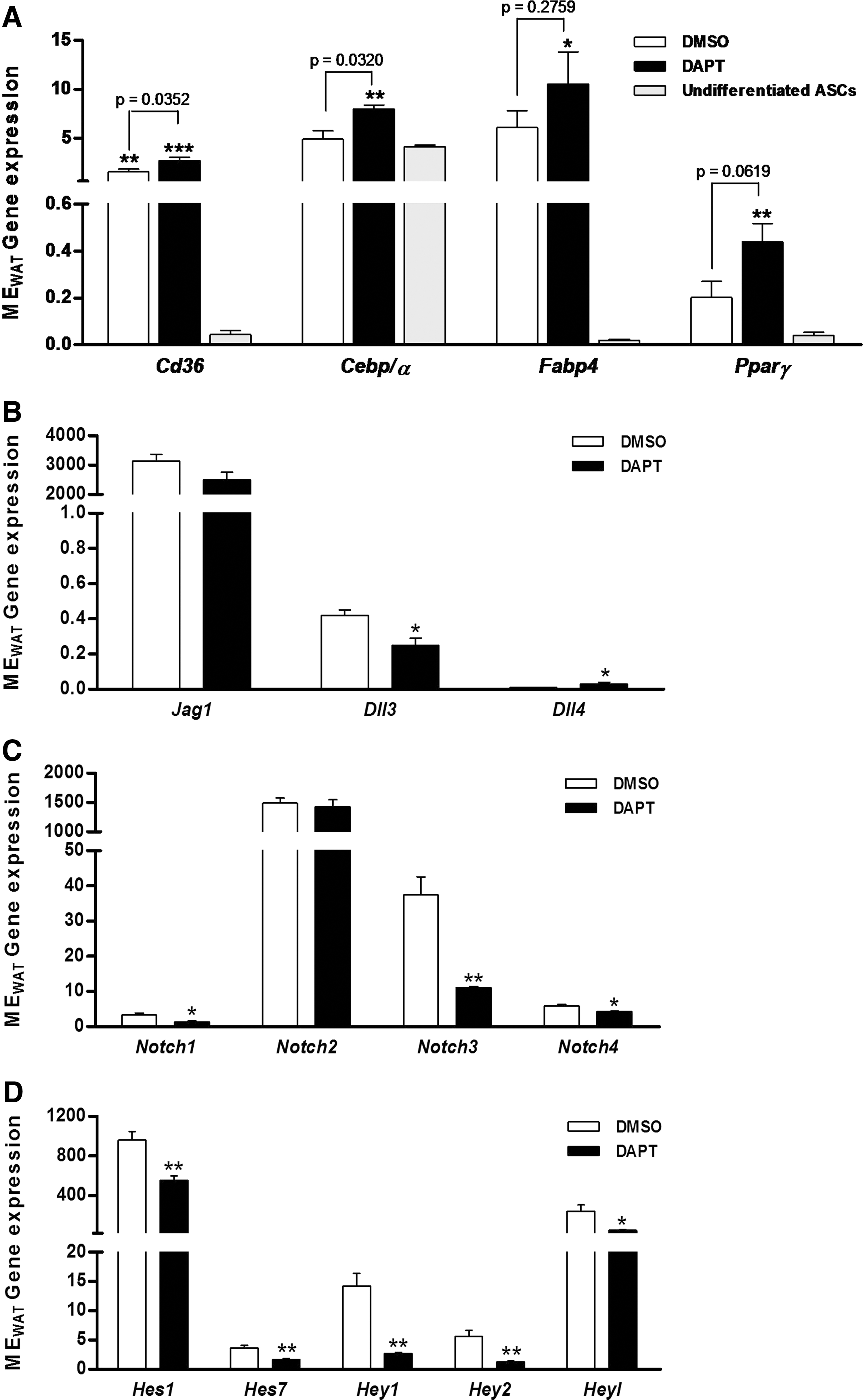
Notch inhibition in ASCs from VWAT by N-[N-(3,5-difluorophenacetyl-L-alanyl)]-S-phenylglycine t-butyl ester (DAPT). ASCs from mesenteric adipose tissues (MEWAT) were pretreated with 50 μM DAPT and induced to differentiate to adipocytes. Adipogenic differentiation potential was evaluated by the gene expression of adipocyte molecular markers
Discussion
Differences between adipose depots are particularly evident when comparing the association between anatomical localization of WAT and cardiometabolic complications [38]. It is not only the total amount of fat but rather its localization and distribution that increase the risk of CVD. Indeed, visceral fat accumulation is closely linked to metabolic complications, whereas the expansion of subcutaneous fat appears to be metabolically protective [7]. The reason for this differential effect remains unclear. It has been suggested that the different metabolic properties between WATs could be ascribed to intrinsic cell-autonomous differences and their different expansion capacities [14]. Several evidences indicate that subcutaneous depots show enhanced ability to generate adipocytes, which may be protective against metabolic dysfunction, while the hypertrophic response characteristic of visceral depots, and subsequently dysfunctionality, may be maladaptive [39]. But the molecular mechanisms to explain this different adipogenic ability remain unknown.
Adipogenesis includes the determination of multipotent stem cells into preadipocytes and the differentiation of preadipocytes into mature fat cells [18]. Although the process of differentiation has been investigated, little knowledge exists on the determination of stem cells to adipogenic lineage. Clearly a large number of questions regarding how adipogenesis is regulated remain to be answered, and understanding this process will be essential to the prevention and treatment of visceral obesity and its associated comorbidities.
Our study proposes that inherent characteristics of ASCs could contribute to the regional variation in fat tissue homeostasis, and have therefore characterized the ASCs derived from different adipose tissue depots. Immunophenotyping and PCR analysis demonstrate that there are substantial differences between ASCs derived from VWAT and SCWAT in control and diabetic animals, above all between MEWAT and SCWAT. In particular, the higher expression of Cd29 and Cd105 in VWATs versus SCWAT is noteworthy in both groups. CD29 is a surface marker present in ASCs as well as committed preadipocytes, but CD105 is a marker only present in ASCs and its downregulation enhances adipogenic capacity of ASCs [40]. Moreover, it has been suggested that downregulation of Cd105 may be a functional requirement for preadipocyte commitment and that the ability to differentiate into adipogenic lineage is restricted to cells not expressing Cd105 [41]. Therefore, we postulate that the differential expression of Cd105 between ASCs derived from VWATs and SCWAT may be a hallmark for the adipogenic ability observed between locations.
Expression profiling revealed significant differences between different depots in expression for some of the main molecules governing the stemness. In general, the expression of these molecules was significantly higher in ASCs derived from VWATs, documenting a greater undifferentiated state in ASCs derived from VWATs compared with ASCs derived from SCWAT. Contrary to expectations, the expression of Pou5f1 was kept lower in ASCs derived from visceral versus subcutaneous fats in the diabetic group. This decrease could be in response to the excess energy availability in order to facilitate preadipocyte commitment and to generate new adipocytes. Among all stemness markers, only FGF2 has been directly related with adipogenesis. It has been recently reported that FGF2 is a regulator of the earliest stages of the adipogenic process [42]. Therefore, in agreement with a negative role for FGF2 in adipogenesis, the observed overexpression of Fgf2 in visceral depots may be a key factor in their differential adipogenic capacities.
Several factors have been identified that commit or inhibit the conversion of MSCs to the adipocyte lineage [43 –47]. Among these, Notch signaling pathway, which is critical in regulation of self-renewal, proliferation, and differentiation of stem cells [20 –24], is included. Notch system has been shown to play a role in adipocyte differentiation [27 –36]. However, conflicting findings have been reported concerning its role. Several evidences have demonstrated that inhibition of Notch signaling enhances adipogenesis [28 –32], and, on the contrary, other research has shown that its inhibition maintains undifferentiated state of progenitor cells inhibiting its adipocyte differentiation [33 –36]. The analysis of the Notch signaling pathway confirmed a higher activation of Notch signaling in ASCs derived from visceral fat depots compared with ASCs derived from SCWAT. Besides the high expression of Hes1 and Hey1, which are known to inhibit adipogenesis by blocking the expression of key transcription factors, such as Pparγ and C/ebpα [27 –29], we observed a marked increase of expression of Notch target genes Hes7, Hey2, and HeyL in VWATs in comparison to SCWAT, which contributes to explain the differences in their adipogenic capacities [14].
In accordance with the expression profiles, significant differences in adipogenic differentiation potential were observed among ASCs derived from different fat depots. Our data suggest that adipocyte cell determination is associated to the level of Notch signaling activation. The visceral WATs show significantly more Notch activation and significantly lower adipogenic capacity, supporting the results of Roldan et al. [47].
Additionally, recent studies showed that Notch signaling is also important in the regulation of stem cell quiescence, an essential property of this cell population [48]. Notch signaling activation is required to maintain the quiescent state, and its inhibition leads to activation, proliferation, and terminal differentiation of stem cells [49,50]. Notch3, Hes1, Hey1, and HeyL, which are highly expressed in other type of quiescent stem cells [50 –52], showed upregulated in ASCs derived from visceral depots compared with ASCs derived from SCWAT, and consistent with these expression profiles, when the proliferation of ASCs was analyzed by BrdU cell proliferation assay, we observed differences between ASCs derived from different fat depots. Therefore, the upregulation of Notch target genes in ASCs derived from visceral depots might avoid the activation, proliferation, and adipogenic differentiation of these cells, keeping their quiescent state.
Our results suggest that the Notch activation in stem cells controls adipogenesis through Jag1, which showed to be highly upregulated in VWATs with respect to SCWAT. The downregulation of Dll1, no expression of Dll4, and the upregulation of Dll3, which might serve exclusively as a cis-inhibiting ligand attenuating signaling induced by other ligands [53], support our hypothesis. Further studies addressed to confirm this hypothesis will be required.
Collectively, our findings support the association between Notch signaling activation and maintenance of quiescent state inhibiting the adipogenic differentiation in ASCs. Therefore, our data pinpoint to a differential activation of Notch system between ASCs derived from SCWAT and VWATs, which would be responsible for their differential adipogenic potential supporting the concept that VWATs and SCWAT have different intrinsic expansion capacities. Finally, in order to confirm the association between the differential activation of Notch system and the differential adipogenic potential, ASCs were treated with PEITC, a Notch activator described as promising cancer chemopreventive agent to date [54], and ASCs were treated with DAPT, a known inhibitor of γ-secretases [29,55]. The present study reveals, for the first time, that PEITC treatment activates Notch signaling in ASCs from SCWAT through the overexpression of Notch1-Hes1 axis, and that this activation reduces their adipogenic differentiation ability. At the same time, the increase of the adipogenic potential of ASCs of visceral origin, resulting from the suppression of Notch system by DAPT treatment, confirmed the role of Notch signaling pathway in the WAT expansion capacity. The lack of response of ASCs from SCWAT to DAPT and ASCs from MEWAT to PEITC treatment could be the result of the respective inhibition and activation of the Notch system at basal level. Thus, the lack of change in Notch target genes and subsequent adipogenic differentiation allow us to confirm again that the aforementioned changes in the adipogenic potential observed were due to the effect on the Notch signaling pathway.
We observed an expression profile very similar in ZLC and ZDF rats, though ASCs derived from diabetic rats showed a lower activation of the Notch pathway in ASCs derived from VWATs relative to ASCs derived from SCWAT, which would explain the greater adipogenic differentiation capacity associated with ASCs derived from visceral locations in ZDF rats. This greater adipogenic differentiation potential could be in response to the excess energy availability. Therefore, the combination of VWATs expandability through adipocyte hypertrophy and inefficient storing with the severe positive energy balance induced by leptin signal deficiency [56] may result in severe insulin resistance, in spillover of lipid to peripheral tissues,and, consequently, the associated metabolic complications [57]. The higher adipogenic capacity of SCWAT observed in ZDF rats might act as compensatory mechanism, but this is not enough to compensate the inherent lack of VWATs expandability with excess energy availability (Supplementary Fig. S3C).
In summary, the results from this study emphasize the differential role of WAT in accordance to its anatomical localization, and pinpoint to differences in their resident stem cell precursors as responsible for the specific adipose tissue depot development and for the functional differences observed between VWATs and SCWAT. We propose that the adipose tissue expandability hypothesis is based upon the resident stem cells. ASCs are the ground for the pathophysiological differences observed between visceral and subcutaneous fat depots, both in healthy as well as diabetic conditions. The overexpression of factors, such as Cd105 and Fgf2, and Notch target genes might induce a blockade in the progression into an adipogenic phenotype in ASCs from VWATs. In an adipogenic environment, when increased storage requirements are needed and this blockade is maintained, the inability of VWAT to form new adipocytes would lead to the adipocyte hypertrophy and limited storage capacity, leading to ectopic fat deposition and, ultimately, to the cardiometabolic alterations. Taken together, our findings suggest that the Notch signaling pathway is an important regulator of adipogenesis and cardiometabolic risk. Further studies are required to get information on the role of the specific Notch system components. Notch activation may be a biomarker for predicting cardiovascular risk and a new tentative therapeutic target to promote adipose tissue storage capacity.
Footnotes
Acknowledgments
The authors would like to thank Dr. Javier Crespo for his technical help with flow cytometric analysis and Marta Sanchez for her excellent technical assistance. This work was supported by research grants from the Spanish Ministry of Health-Instituto de Salud Carlos III (RETIC RIC and TERCEL) and the Spanish Ministry of Science and Innovation (PNS SAF2013-42962-R). R.F.L. is a Sara Borrell investigator (Spanish Ministry of Science and Innovation-Instituto de Salud Carlos III) and M.T.B. is granted with a fellowship from the Catalan Government (FI-DGR 2012).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.