
Abstract
Atherosclerosis is a major contributor to life-threatening cardiovascular events, the leading cause of death worldwide. Since the mechanisms of atherosclerosis have not been fully understood, currently, there are no effective approaches to regressing atherosclerosis. Therefore, there is a dire need to explore the mechanisms and potential therapeutic strategies to prevent or reverse the progression of atherosclerosis. In recent years, stem cell-based therapies have held promises to various diseases, including atherosclerosis. Unfortunately, the efficacy of stem cell-based therapies for atherosclerosis as reported in the literature has been inconsistent or even conflicting. In this review, we summarize the current literature of stem cell-based therapies for atherosclerosis and discuss possible mechanisms and future directions of these potential therapies.
Introduction
A
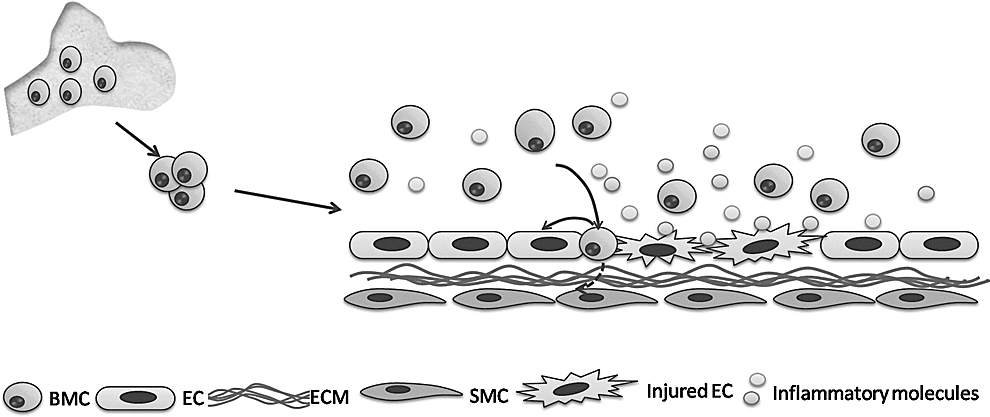
The role of bone marrow-derived stem cells (BMCs) in atherosclerosis. Injured endothelial cells (ECs) would recruitment BMCs to repair them through secretion of inflammatory molecules. Recruited BMCs would directly transdifferentiate to endothelial cells and smooth muscle cells (SMCs), or promote local endothelial cell repair and regulate inflammatory response through the paracrine effect. ECM, extracellular matrix.
Animal Models of Atherosclerosis
Benefited with the advancement of genetic modifying technology, genetically manipulated mouse models become a strong tool to study mechanisms of atherosclerosis. For example, apolipoprotein E-deficient (ApoE−/−) mouse and low-density lipoprotein (LDL) receptor-deficient (LDLR−/−) mouse are the most commonly used models. Given that each animal model of atherosclerosis has its advantages and limitations, it is important to recognize the diversity of animal models to interpret different or potentially conflicting results in different laboratories.
ApoE−/− mouse model
The ApoE−/− mouse model was first developed using homologous recombination in embryonic stem cells (ESCs) in 1992 [36,37], which is perhaps the most widely used model in atherosclerosis. In ApoE−/− mice, fibrous plaques consist of a fibrous cap containing SMCs surrounded by a connective tissue matrix with inflammatory cell accumulation [36,38]. This pathology resembles atherosclerotic lesions found in humans. Unfortunately, this mouse model is not suitable for investigations of plaque rupture or thrombosis because these two critical pathological changes in humans are hardly observed in ApoE−/− mice. Moreover, apoE is present in bone marrow-derived cells (mostly in macrophage) [39 –42], and reconstitution of ApoE−/− mice with bone marrow from wild-type (WT) mice decreases atherosclerosis attributed to the presence of apoE and its effect on plasma lipid levels [40]. Therefore, ApoE−/− mice are appropriate to study the roles of many factors in the development of atherosclerosis, but are not suitable for examining the effects of stem cell-based therapies.
LDLR−/− mouse model
LDL receptor (LDLR) removes cholesterol-rich intermediate density lipoproteins (IDL) and LDL from plasma and, thereby, regulates lipid levels in plasma [43]. Unlike ApoE−/− mice, lesions formed in LDLR−/−mice are of simple morphology consisting predominantly of lipid-laden macrophages [44]. The advantage of LDLR−/− mice is that the expression of the LDLR on transplanted bone marrow-derived cells does not have an impact on atherosclerosis or plasma lipid levels as it does in ApoE−/− mice [45]. However, a Western-type fat diet (typically containing 21% wt/wt saturated fat) is needed to promote atherosclerotic lesions [46].
Reversa mouse model
Recently, another model, Reversa mouse (LDLR −/− ApoB 100/100 Mttp fl/fl Mx1-Cre +/+) provided the opportunity to study atherosclerotic plaque regression, a hypercholesterolemic mouse model in which high plasma cholesterol levels can be reversed just by flipping a genetic switch. Reversa mouse model has severe hypercholesterolemia as a result of atherogenic LDLR knockout (LDLR−/−) and expression of apolipoproteinB100 (apoB100). However, hypercholesterolemia in this mouse model can be conditionally eliminated by inducing the expression of the Mx1-Cre transgene with intraperitoneal injections of polyinosinic- polycytidylic ribonucleic acid (pI-pC), which inactivates Mttp, the gene for microsomal triglyceride transfer protein, thereby preventing hepatic lipoprotein assembly and secretion [47,48]. This new mouse model will be useful for evaluating the roles of stem cell therapies for atherosclerosis in the context of reversal of hypercholesterolemia and regression of atherosclerosis [49], and exhibiting promising potential to uncover potential pathways involved in the progression of atherosclerosis especially in humans receiving statin therapy.
In conclusion, LDLR−/− mouse is an optimal model for evaluating effects of stem cell-based therapies for atherosclerosis since its plasma cholesterol concentrations are not influenced by the bone marrow-derived source. Moreover, studies focusing on new atherosclerosis models, for example, the Reversa model, may provide insights into stem cell-based therapies for atherosclerosis.
Stem Cell Mobilization
There is accumulating evidence that autologous stem cells could be mobilized from bone marrow to peripheral blood under certain conditions. Hence, enhancing stem cell mobilization has been believed to be a good therapeutic option in cardiovascular diseases. It was found that CXCR4 antagonist AMD3100 could mobilize bone marrow stem cells to peripheral blood, affect plaque regression, and consequently attenuate atherosclerosis [49]. In addition, increasing the number of circulating endothelial progenitor cells (EPCs) by administrations of statin, estrogen, erythropoietin, or granulocyte colony-stimulating factor treatment, physical exercise not only accelerates reendothelialization but also profoundly inhibits neointimal formation after vascular injury [50 –58]. Moreover, promoting smooth muscle progenitor cell (SPC) mobilization by CXCL12 could stabilize the atherosclerotic lesions [59]. Since endogenous stem cell mobilization alone is insufficient to compensate for the reduced stem cell number and function in the context of atherosclerosis, thus exogenous stem cell transplantation is needed.
Types of Stem Cells
There are mainly two broad types of stem cells based on their origin: ESCs and adult stem cells. Adult stem cells include whole bone marrow-derived stem cells (BMCs) and its subpopulations, EPCs, SPCs, and hematopoietic stem cells (HSCs). Adult stem cells, but not ESCs, are commonly used in atherosclerotic studies. In addition to adult stem cells, the efficacy of intravenous administration of induced pluripotent stem (iPS) cells was also investigated [60]. The different studies performed previously are summarized in Table 1. Since few studies directly compared the beneficial effects of different stem cells in atherosclerosis, the optimal cell type for stem cell therapies in atherosclerosis has not been defined.
BMC, bone marrow-derived stem cell; BM-MNCs, bone marrow-derived mononuclear cells; EPC, endothelial progenitor cell; HFD, high fat diet; iPS, induced pluripotent stem; i.v., intravenous; NA, not associated; SMC, smooth muscle cell; SPC, smooth muscle progenitor cell; VEGF, vascular endothelial growth factor; WT, wild type.
Bone marrow stem cells
BMCs are a mixture of many cell populations, including HSCs, MSCs, SPCs, and some inflammatory cells. BMCs are the earliest and widely studied cell type in atherosclerosis, but the therapeutic effects are still controversial.
In 1995, Linton et al. [40] first transplanted bone marrow from ApoE+/+ mice into ApoE−/− mice that received lethal irradiation. Transplantation of bone marrow cells from WT mice to ApoE−/− mice displayed protection from diet-induced atherosclerosis associated with the apoE secretion by macrophages from BMCs. Additionally, it has also been observed that an accelerated high-fat diet induced atherosclerosis in ApoE+/+ mice reconstituted with ApoE−/− bone marrow [41,42]. Consistently, treating ApoE−/− mice with apoE-encoding vectors could induce plaque regression [61], further supporting the protective role of apoE in atherosclerosis. This initial bone marrow transplantation study has evoked attention to subsequent atherosclerosis studies in understanding potential beneficial effects of stem cell therapies in atherosclerosis.
Subsequent studies demonstrated that periodic treatment of ApoE−/− mice fed saturated fat-enriched diet with BMCs from young nonatherosclerotic ApoE−/− mice or WT mice prevented atherosclerosis progression despite persistent hypercholesterolemia in the recipient mice. In contrast, treatment with BMCs from older ApoE−/− mice was much less effective than the young counterparts [32]. These data indicate that progressive progenitor cell deficits contribute to the development of atherosclerosis and BMCs have atheroprotective properties that would be exhausted with aging and exposure to atherosclerosis. However, when excluding the confounding effect of age of the donor mice as mentioned above, transplantation of BMCs from age-matched ApoE−/− mice resulted in an increased size of atherosclerotic lesions [34]. The injected cells were found predominantly within the lipid core of the lesions and not in the endothelial or subendothelial regions. This might be related to the fact that BMCs are principally mononuclear in origin harboring proinflammatory properties. Together, age is an important factor affecting the result of stem cell therapies.
In addition to age, sex also influences the efficacy of stem cell therapies on atherosclerosis [62]. Nelson et al. provided evidences on the sex-related difference in response to BMC therapy for atherosclerosis. Administration of BMCs derived from WT female donors attenuated plaque formation in only male atherosclerotic ApoE−/− mice. In contrast, no reduction in plaque was observed either in male BMCs administered to males or BMCs from donors of both sex infused into females [63]. These variants might be explained by the fact that endogenous repair and response to cell therapies differ between males and females. Sex-associated differences in endogenous repair, different responses to BMCs at the level of BM and blood (they found female BMCs contained more EPCs), and different hormonal and inflammatory milieu are all responsible to this sex difference in the efficacy of cell therapies.
In the clinic, atherosclerosis is always accompanied by other diseases or lead to systemic vascular diseases such as peripheral arterial disease [64]. The safety of stem cell therapy in patients with widespread atherosclerosis has been tested by Silvestre et al. [33], in which they first examined the BMC-based therapy for atherosclerosis in the setting of hindlimb ischemia. Interestingly, they discovered that transplantation of WT BMCs induced a significant increase in the lesion size in ApoE−/− mice with hindlimb ischemia, but did not the affect lesion size in mice without ischemia. However, ApoE−/−-derived BMCs transplanted into mice with or without ischemia had neither proangiogenic nor proatherogenic effects. Thus, we should not ignore the side effects of stem cell therapies when treating atherosclerosis accompanied by ischemic diseases.
Bone marrow stem cell subpopulations
BMCs are composed of different subpopulations; Tousoulis et al. [65] showed that direct infusion of Lin−/Sca-1+ cells significantly decreased the levels of proinflammatory molecules and oxidative stress parameters, exerting beneficial effects on vascular inflammation and atherosclerotic plaque development. Further studies performed by Song et al. [66] demonstrated that Lin- cells were better than the whole BMCs after comparing different stem cell therapies in atherosclerosis. Although different subpopulations of spleen-derived cells, including CD11b+ (monocyte marker), CD45R+ (B-cell marker), and Sca-1+ (stem cell marker) BMC subpopulations, could all significantly improve endothelium-dependent vasodilation, these cells were not as effective as transfusion of unselected BMCs [67], suggesting that more than one subpopulation of the BMCs are involved in this effect.
In conclusion, considering the complexity of BMC subpopulations, they have both reparative capabilities and inflammatory properties, further studies are needed to determine which BMC subpopulation could contribute to the best effect on atherosclerosis.
Hematopoietic stem cells
Accumulated evidence indicates that HSCs were a potential source of SPCs and EPCs and participate in the progression of atherosclerosis [68]. However, the effect of direct injection of HSCs has not been determined.
Mesenchymal stem cells
MSCs are nonhematopoietic stromal cells that were first isolated from the bone marrow and subsequently from other adult connective tissues. MSCs are considered as promising candidates for myocardial repair attributed to their unique properties of efficient self-renewing, immune-tolerance, and releases of paracrine molecules and multipotent differentiation potential [69,70]. They have been widely studied in clinical trials, and their beneficial effects on myocardial infarction are convincing. However, few studies investigated the role of MSCs on atherosclerosis. MSCs might exert beneficial effects on atherosclerosis since MSCs have been shown to be highly immunosuppressive [71] and have the ability to differentiate into endothelial cells to enhance endothelial function [72 –75].
Endothelial progenitor cells
Since first isolated from adult peripheral blood in 1997 [12] and later found in bone marrow and other tissues [13,76,77], EPCs have attracted major interest in recent years. EPCs support postnatal angiogenesis and vasculogenesis in experimental models. In addition to directly homing to and differentiating into mature endothelial cells in the damaged endothelium [13,78], EPCs could also produce angiogenic cytokines and growth factors to improve proliferation of existing resident endothelial cells [78 –80]. Unfortunately, the definition of EPCs is still controversial, and isolation and culture methods of EPCs are also not consistent in different laboratories. These issues make difficult to compare the experimental results of EPCs from different laboratories [81,82]. Therefore, it is critical to characterize EPCs and develop a standard procedure to isolate and culture EPCs in the future.
There are two distinct populations of EPCs, that is, early EPCs and late outgrowth endothelial cells (OECs) [82 –85]. Early EPCs constitute a heterogenic population that mainly originates from myeloid hematopoietic cells and shares features with immune cells, particularly monocytes/macrophages [82], and they could indirectly augment angiogenesis in a paracrine manner. In contrast, late OECs always directly incorporate into developing vascular networks, but exert no paracrine angiogenic effects [84]. The different angiogenic properties of early EPCs and late OECs suggest that these cells may have distinct therapeutic roles and that different strategies may be required to optimally harness their respective potential for therapeutic neovascularization. Although there are many differences, including isolation and culture methods of different EPC populations, there is compelling evidence that EPCs could be mobilized to the ischemic site and performed the repair process of injured artery, which could bring beneficial effects on ischemic diseases [29,86].
In clinical studies, the number and activity of cultured EPCs isolated from peripheral blood are much lower in patients with high cardiovascular risk factors compared with those having no cardiovascular risk factors [87,88]. Thus, it is possible that repopulation of EPCs with normal function be beneficial for cardiovascular diseases. The efficacy of direct transplantation of EPCs to atherosclerotic mice is controversial in animal experiments, although EPC capture stent has become available in clinical treatment [89]. Most experiments showed the beneficial effects of EPC transfusion [49,65], since EPCs could provide protection against atherosclerosis by their innate ability to replace dysfunctional ECs [32], improve proinflammatory and oxidative stress markers [65], and enhance atherosclerotic plaque regression [49]. In contrast, George et al. work showed that EPCs isolated from ApoE−/− mice resulted in increases of atherosclerotic lesion size [34]. Zoll et al. reported that EPCs derived from human umbilical cord blood samples had no effects on atherosclerotic plaque progression [90]. Moreover, the findings of Hagensen et al. challenge the prevailing theory that circulating EPCs contribute to atherosclerosis. Their results demonstrated that circulating EPCs rarely contributed to the maintenance and regeneration of plaque endothelium in atherosclerosis [35].
Taken together, EPCs remain to be a promising cell type for atherosclerosis treatment and deserve further studies, although controversies exist. However, potential adverse effects of promoting plaque angiogenesis require consideration when using EPCs to promote therapeutic angiogenesis in ischemic diseases.
Smooth muscle progenitor cells
SMCs are the predominant cell type of vascular walls. There is accumulating evidence that SMC migration, proliferation, and matrix synthesis play essential roles in atherosclerosis development [91]. In the early stage of atherosclerosis, SMCs migrate into the intima, and take up the lipid to form foam cells, which aggravate the stage of atherosclerosis [21]. Conversely, in the complicated progression stage, the fibrous cap composing of SMCs can maintain the stability of plaque, and hence decreases the risk of plaque rupture and life-threatening arterial thrombosis [92,93]. SMC apoptosis accelerates plaque rupture [94]. In this respect, SMCs could serve as a promising target for atherosclerosis treatment. Although it was widely accepted that SMCs in plaques originated from the local vessel wall [91,95,96], its bone marrow origin has been demonstrated by several studies [16,17,97 –99], and SPCs have been discovered in the circulation blood derived from bone marrow [16,100]. When transplanted into ApoE−/− mice or enhanced recruitment with CXCL12, SPCs can exert beneficial effects, limit plaque development, and promote changes in plaque compositions [59,90]. It was assumed that the beneficial role of SPCs resulted from the potential secretion of a number of growth factors that could stimulate collagen synthesis by SMCs in the plaque. Moreover, peripheral blood-derived SPCs decreased in patients with acute coronary syndrome compared with those in patients with stable angina [90], raising the possibility that SPCs might play an important role in plaque stabilization. However, there are a few studies indicating that SPCs could secrete proinflammatory cytokines and promote atherosclerosis [101,102]. Therefore, further studies are required to demonstrate the origin and specific markers of SPCs and their influences on atherosclerosis.
Other cell types
Effects of spleen-derived mononuclear cells and their different subpopulations have also been determined [34,67], in which spleen-derived EPCs are the frequently used cell types [34,67]. These cell types have atheroprotective capability. A more recent study suggested that intravenously administered iPS cell-derived Flk-1+ cells are recruited to the site of vascular injury, thereby enhancing reendothelialization in a vascular injury model [60].
In summary, nowadays, there is no agreement as to which cell type is the best candidate in stem cell therapies for atherosclerosis. The existing evidence supports the notion that stem cells from WT mice have atheroprotective effects, but in ApoE−/− mice, their effects need to be further elucidated. The definition, isolation, and culture methods of stem cells need to be standardized to replicate experimental data from different laboratories.
Methods and Dosages of Stem Cell Delivery
In mice, it is difficult to achieve selective intracoronary injections as in humans; the systemic route is the most commonly used approach. Thus, the distribution of injected cells in mice should be examined. Lots of injected cells might be trapped within the lungs and spleen before reaching the plaque [52,90]. In this case, a single dose of injection is always not sufficient. The dosages of stem cells range from 5×105 to 5×106 according to the nature of administered cells. Accumulating data suggest that the best delivery protocol is intravenous injection of 106 cells/each injection in each 2-week interval for a total of six times [66].
Mechanisms of Stem Cell-Based Therapies
Mechanisms of atherosclerosis have not been fully elucidated. Hyperlipidemia, endothelial injury, and inflammatory response are recognized as critical contributors of this disease. In the treatment of atherosclerosis, it is important to lower plasma cholesterol concentrations, maintain the balance between endothelial injury and repair, and control inflammatory responses.
Lipid metabolism regulation
Hypercholesterolemia is an important risk factor for atherosclerosis. ApoE−/− mice receiving whole bone marrow transplantation from WT mice exhibited decreased plasma cholesterol concentrations, which might be related to the presence of apoE in plasma [40]. However, this effect was not observed in other atherosclerosis models that received stem cell therapies [7,11 –13,16,42,43]. It is not known whether this discrepancy is related to plasma or local apoE concentrations since it was not analyzed in recipient mice.
Direct differentiation into endothelial cells
Rauscher et al. [32] have shown that injected stem cells could directly differentiate into endothelial cells. However, George et al. [34] found that injected cells were predominantly within the lipid core of the plaques, but not in the endothelial or subendothelial regions. In Silvestre's study [33], only occasionally transplanted cells were found within the plaques, making a physical or local contribution of the injected BMCs to plaque growth unlikely. Further studies are required to explore whether stem cells directly differentiate into endothelial cells in stem cell-based therapies for atherosclerosis.
Paracrine effects and inflammatory responses
Paracrine effects and inflammatory responses play crucial roles in stem cell-based therapies for cardiovascular diseases, in which transplanted stem cells can secret a spectrum of cytokines that benefit the resident cells to improve vasodilation or regulate inflammatory and oxidative responses [103 –105]. The data from mouse models of atherosclerosis showed that inflammatory molecules, including IL-6, MCP-1, and vascular adhesion molecules, profoundly changed. Several studies reported that an attenuated lesion size was correlated with decreased expression of inflammatory molecules [32,65,67], and conversely, some studies found that aggravation of atherosclerosis was associated with enhanced expression of inflammatory molecules [34].
As mentioned above, there are two processes required for attenuation of plaques: (1) appropriate inflammation to mobilize the necessary bone marrow progenitors and to trigger digestion of lesions by macrophages; and (2) vessel repair, which includes engraftment of stem cells to renew the injured endothelium [63]. The initial inflammation is a positive signal for the recruitment of endogenous bone marrow stem cells. When reparative cells reach sufficient numbers, the repair process ensues and the inflammation process decreases. On the other hand, once insufficient numbers of reparative cells or cells incapable of repair are recruited, the repair process would stop, and inflammation would increase and become a negative signal to the detriment of the tissue. It is important in stem cell-based therapies to maintain this balance and stimulate it to develop in the right direction.
Taken together, stem cells participate in the progression of atherosclerosis mainly through the secretion of inflammatory factors and their innate inflammatory nature, and regulate the inflammatory processes of atherosclerosis.
Conclusions and Perspectives
Stem cell-based therapies for atherosclerosis have mainly been performed in animal experiments to date, and have not reached a consistent conclusion. This might be, at least partially, attributed to different protocols used, different time points explored, and different analyses of data in different laboratories, hence at least some important problems should be taken into account: (1) proper animal models: there is a need to explore new atherosclerosis models suitable for evaluating stem cell actions; (2) normalizing diverse transplantation protocols, including age, sex, genetic background of donors and recipients, careful and thorough cell-type characterizations, delivery method of transplanted cells, and the best timing of cell infusion; (3) choosing appropriate experimental parameters and endpoints: the study performed by Tousoulis et al. [65] revealed that some inflammatory markers (vascular cell adhesion protein 1 and E-selectin) were increased 7 days post-treatment, but significantly decreased 6 weeks after the initiation of treatment. Exogenous stem cells might induce transient inflammatory responses in recipients; (4) definite tracking of the fate of stem cells: it remains an open question to track the distribution and fate of stem cells in the body. Elucidating this issue would gain new knowledge for mechanisms of stem cell-based therapies; and (5) the species differences between humans and animals: no animal atherosclerosis model could recapitulate the complex development of atherosclerosis in humans. Thus, this alteration should be considered when interpreting results from animals. The problems above should be noted in the future and well-designed experiments need to be done to accurately evaluate influences and mechanisms of stem cell-based therapies for atherosclerosis.
Gene therapies and preconditioning of stem cells are new hopeful therapeutic approaches. With state-of-the-art technologies, it is not just a dream to upregulate or downregulate the expression of some genes or precondition stem cells to exert their beneficial effects to the fullest. Recent studies suggest that modified stem cells with some genes could modulate the properties of stem cells and hence enhance the efficacy of stem cell-based therapies for atherosclerosis. For example, administration of apoptotic bodies or miR-126 limited atherosclerosis and promoted the incorporation of Sca-1+ progenitor cells in different mouse models of atherosclerosis [106]. In addition, eNOS, a vascular protective gene, can enhance EPC action on atherosclerosis [107]. Moreover, endothelium-specific overexpression of SIRT1 can decrease atherosclerosis in ApoE−/− mice through improving endothelial cell survival and function [108]. Thus, modifying stem cells with some genes, which could modulate the properties of stem cells, might provide new hope for enhancing the efficacy of stem cell-based therapy for atherosclerosis.
BMCs reduced plaque burden by 40% in atherosclerotic mice [63], whereas rosuvastatin only led to a 7% reduction in patients with coronary artery disease [109], raising the possibility that the atheroprotective role of stem cells holds promise. However, a considerable amount of work needs to be done before stem cell therapeutics of atherosclerosis can be used widely in clinical patients. If we can make the best use of the advantage of stem cells and bypass their disadvantages, stem cell-based therapies will open new avenues for therapies of atherosclerosis.
Footnotes
Acknowledgment
This work was supported by the National Natural Science Foundation of China (81100092, 31171392, and 31371475). The authors appreciate the editorial assistance of Dr. Hong Lu (University of Kentucky). They also thank Keke Zhu (University of Shanghai for Science and Technology) for preparing ![]() in this review.
in this review.
Author Disclosure Statement
No competing financial interests exist.