
Abstract
Germ cells (GCs) are critically important as the vehicle that passes genetic information from one generation to the next. Correct development of these cells is essential and perturbation in their development often leads to reproductive failure and disease. Despite the importance of GCs, little is known about the mechanisms underlying the acquisition and maintenance of the GC character. Using a reprogramming strategy, we demonstrate that overexpression of ectopic transcription factors in embryonic fibroblasts can lead to the generation of chicken induced primordial germ cells (ciPGCs). These ciPGCs express pluripotent markers POU5F1, SSEA1, and the GC defining proteins, CVH and DAZL, closely resembling in vivo sourced PGCs instead of embryonic stem cells. Moreover, CXCR4 expressing ciPGCs were capable of migrating to the embryonic gonad after injection into the vasculature of stage 15 embryos, indicating the acquisition of a GC fate in these cells. Direct availability of ciPGCs in vitro would facilitate the study of GC development as well as provide a potential strategy for the conservation of important genetics of agricultural and endangered birds using somatic cells.
Introduction
T
Despite the utility and strength of PSCs, significant prohibitive limitations exist in mammalian systems. Often times, it would be preferable to directly manipulate PGC cultures instead of PSCs. PGCs are likely more representative of a naive in vivo germline-transmissible cell type than the undifferentiated PSCs, thus their response to a GC toxin or other perturbations may be more representative of how PGCs and eventually GCs behave in the developing embryo and offspring. However, PGC studies are difficult in mammals since mouse PGCs, when cultured in vitro, do not maintain a PGC phenotype and will revert back to a cell type that is equivalent and resembles ESCs [13 –15]. These considerations need to be acknowledged when studying critical signaling or exposure to a toxin and subsequent transfer into an in vivo setting to examine transgenerational effects. In contrast, chicken PGCs are easily isolated from embryos at several time points in GC development and migration, that is, germinal crescent, blood, or nascent gonad [16,17], and do not undergo overt dedifferentiation to an ESC-like cell and they can be propagated as PGCs [18,19]. When transferred into the developing embryo, cultured PGCs migrate to the genital ridge fully undergoing GC development and have been used in avian transgenic studies. However, the early specification and developmental events of PGCs in the chicken are challenging to study, as they happen before the egg is laid. A chicken iPSC GC differentiation system will enable the study of GC development from early specification to functional gamete.
PGCs have significant potential for the development of transgenic animals of unique backgrounds or conservation of valuable genetics, as in the case of endangered birds. Over 1,300 avian species, or 13% of the total population, are threatened by extinction, and an additional 880 species are threatened [20]. Although various conservation measures such as habitat protection and captive reproduction are a solution used to protect and repopulate endangered birds, the overall number of species is still on the decline and some of them are extinct [21]. A PSC to GC approach offers an additional theoretical strategy for the conservation of genetic diversity and repopulation of the endangered avian species and studies using unique adult genotypes [22].
Our previous work demonstrated that quail iPSCs could be generated from somatic cells using human pluripotency transcription factors and revealed that the regulatory mechanisms of pluripotency are conserved across species [23]. Therefore, it may be possible to reprogram chicken somatic cells using similar factors and chicken GC culture conditions to derive chicken GCs [18,24]. In this study, we successfully generated PSCs by overexpression of human transcription factors in chicken somatic cells. The PSCs express typical stem cell markers and are capable of differentiating into all three germ layers. Moreover, these induced chicken PSCs when propagated in a medium used to expand PGCs, expressed important GC genes such as CVH and DAZL. Functional tests demonstrated that these newly reprogrammed chicken induced primordial germ cells (ciPGCs) were capable of migrating to the embryonic gonad after injection into stage 15 chicken embryos, an indication of GC fate. The capability to derive PGCs from somatic cells is the first step toward definitive GC studies in birds and provides a new strategy for the conservation of endangered birds as well as a cell source for the study of toxicology in GC development.
Materials and Methods
Cell culture and iPSC derivation
The chicken embryonic fibroblast (CEF) cells used in the transduction were isolated from day-11 Barred Rock (BR) and Black Australorp (BA) embryos and cultured in the fibroblast medium [Dulbecco's modified Eagle's medium (DMEM) high glucose (Hyclone) with 10% fetal bovine serum (FBS; Hyclone), 4 mM L-glutamine (Gibco), and 1× pen/strep (Gibco)] in 5% CO2 at 37°C. One day before transduction, a total of 1.5×105 CEF cells were plated in one well of a 12-well plate. CEFs were transduced utilizing the viPS kit (Thermo Scientific) with lentiviruses containing the human stem cell genes POU5F1, NANOG, SOX2, LIN28, KLF4, and C-MYC at a multiplicity of infection of 10 and in the presence of 1× TransDux (System Biosciences). Twenty-four hours after transduction, the transduced cells were replated at a ratio of 1:10 onto a 1×100 mm plate preseeded with mitomycin C-inactivated mouse embryonic fibroblast (MEF) in the cKSR medium [DMEM/F12 (Gibco) supplemented with 20% knockout serum replacement (KSR; Gibco), 2 mM L-glutamine (Gibco), 0.1 mM nonessential amino acids (Gibco), 1× pen/strep (Gibco), 0.1 mM β-mercaptoethanol (Sigma- Aldrich), and 10 ng/mL basic fibroblast growth factor (bFGF; R&D Systems), the medium was conditioned in MEF for 1 day before use] or the cKO medium [KO-DMEM (Invitrogen) containing 4 ng/mL bFGF, 7.5% defined FBS (Hyclone), 2.5% chicken serum (Sigma), 1× pen/strep (Gibco), 1× GlutaMAX (Gibco), 1× GS nucleoside supplement (Millipore), and 0.1 mM β-mercaptoethanol, with 10% of the KO-DMEM preconditioned in BRL cells (ATCC) for 3 days before use]. The presumed chicken iPSCs were manually isolated by using a pasteurized glass pipette under a stereomicroscope 7 days after transduction and replated onto fresh MEF feeder cells. Cells were passaged every 4 to 5 days by manual isolation or trypsinization.
Alkaline phosphatase and periodic acid Schiff's staining
Alkaline phosphatase (AP) staining was conducted by using the VECTOR Red Alkaline Phosphatase Substrate Kit as per the manufacturer's instructions. Briefly, the culture medium was removed, then the cells were gently rinsed once with the Tris-HCl buffer (pH8.3), and then incubated with the premixed staining solution at room temperature for 20 min. At the end of incubation, the cells were gently rinsed with Tris-HCl twice. Then, cells were overlaid with phosphate-buffered saline (PBS), and cells were imaged on the microscope.
Periodic acid Schiff (PAS) staining was conducted by using the periodic acid solution and Schiff's reagent (Sigma) as per the manufacturer's instructions. Briefly, cells were fixed in a culture dish with 4% paraformaldehyde (PFA) for 5 min and then washed with PBS three times. The periodic acid solution was then added onto the plate and incubated at room temperature for 5 min. Then, the cells were rinsed with PBS three times and the Schiff's reagent was added to the plate and incubated at room temperature for 15 min. The cells were gently rinsed with PBS three times, and cells were imaged on the microscope.
Embryoid body formation and differentiation
Embryoid bodies (EBs) were prepared in cKO medium without bFGF by using an AggreWell plate (Stemcell Technologies). A total of 2.4×106 chicken PSCs at passage 16 were plated in each well of the AggreWell plate, which is equivalent to 2,000 cells per microwell. After 24 h, the aggregates were harvested by gentle pipetting, and then transferred to a Petri dish, and continued to culture for 6 days. EBs were collected for RNA isolation and cDNA synthesis. Polymerase chain reaction (PCR) was performed to detect the differentiation using primers listed in Supplementary Table S1 (Supplementary Data are available online at
DNA, RNA isolation, and PCR
Colonies of chicken pluripotent stem cells (cPSCs) at passage 12 were dissociated from the feeder layer by gentle pipetting. DNA was isolated using the DNeasy Blood & Tissue Kit (Qiagen). RNA isolation was conducted using the RNeasy QIAprep Spin miniprep Kit (Qiagen) following the manufacturer's instructions. cDNA was synthesized using the SuperScript VILO cDNA Synthesis Kit (Invitrogen). Polymerase chain reaction (PCR) and reverse transcription polymerase chain reaction (RT-PCR) were performed using primers listed in Supplementary Table S1. Quantitative PCR (qPCR) was performed using the 7500 Real-Time PCR System and TaqMan certified primers (Life Technologies), and CEF was used as a control cell line and GAPDH as an endogenous control. All the samples were replicated three times.
Immunocytochemistry and flow cytometry
Cells were fixed in 4% PFA for 15 min and blocked in 4% horse serum for 45 min. Cells were then incubated with primary and then secondary antibodies prepared in a blocking solution at room temperature for 60 min, respectively. The primary antibodies used in this study were POU5F1 (Santa Cruz, #sc-9081), SOX2 (R&D Systems, #MAB2018), SSEA1 (Developmental Studies Hybridoma Bank, #MC-480), EMA1 (Developmental Studies Hybridoma Bank, #Supernatant), HuC/D+ (Invitrogen, #A-21271), SOX17 (Santa Cruz, #SC-17355), alpha-smooth muscle actin (aSMA; Abcam, #ab7817-500), SOX1 (R&D Systems, #AF3363), DAZL (gift from James Petitte, NCSU), VASA (DDX4; Abcam, #ab1384-100) and CXCR4 (ECM Bioscience, #CP4211). Secondary antibodies were all from Invitrogen. Imaging for immunocytochemistry was done by using an Ix81 with the Disc-Spinning Unit (Olympus), and flow cytometry analysis was done using a flow cytometer Cyan (Beckman Coulter).
Cell injection and migration
Before injection, cells were labeled with PKH26 (Sigma) according to the manufacturer's instruction. White leghorn chicken embryos at stage 15 were used as a host for the injection. A 1-cm-diameter window was made on the blunt end above the air cell to expose the embryos. A total of 1×104 cells were loaded into a microglass needle and injected into the vasculature system of each embryo. The window was sealed by applying two layers of parafilm. The injected embryos were incubated for 6 days and then euthanized to isolate the gonads under a stereomicroscope. Images of gonads were captured under an inverted microscope.
Results
Pluripotent chicken stem cell derivation from fibroblasts
To generate PSCs, CEFs isolated from five different BR or five unique BA embryos were transduced with six human reprogramming factors. Putative cPSCs were cultured in the cKSR medium, following a previously developed protocol utilized to generate quail iPSCs [25]. The cPSC colonies emerged as early as day 5 and were manually selected for propagation on day 7 after transduction (Fig. 1A). However, 8 of the 10 lines generated ceased to proliferate by passage 10. The remaining two lines demonstrated high levels of proliferation, but did not express the pluripotent markers POU5F1 or SSEA1 (data not shown) and were not utilized in further studies.
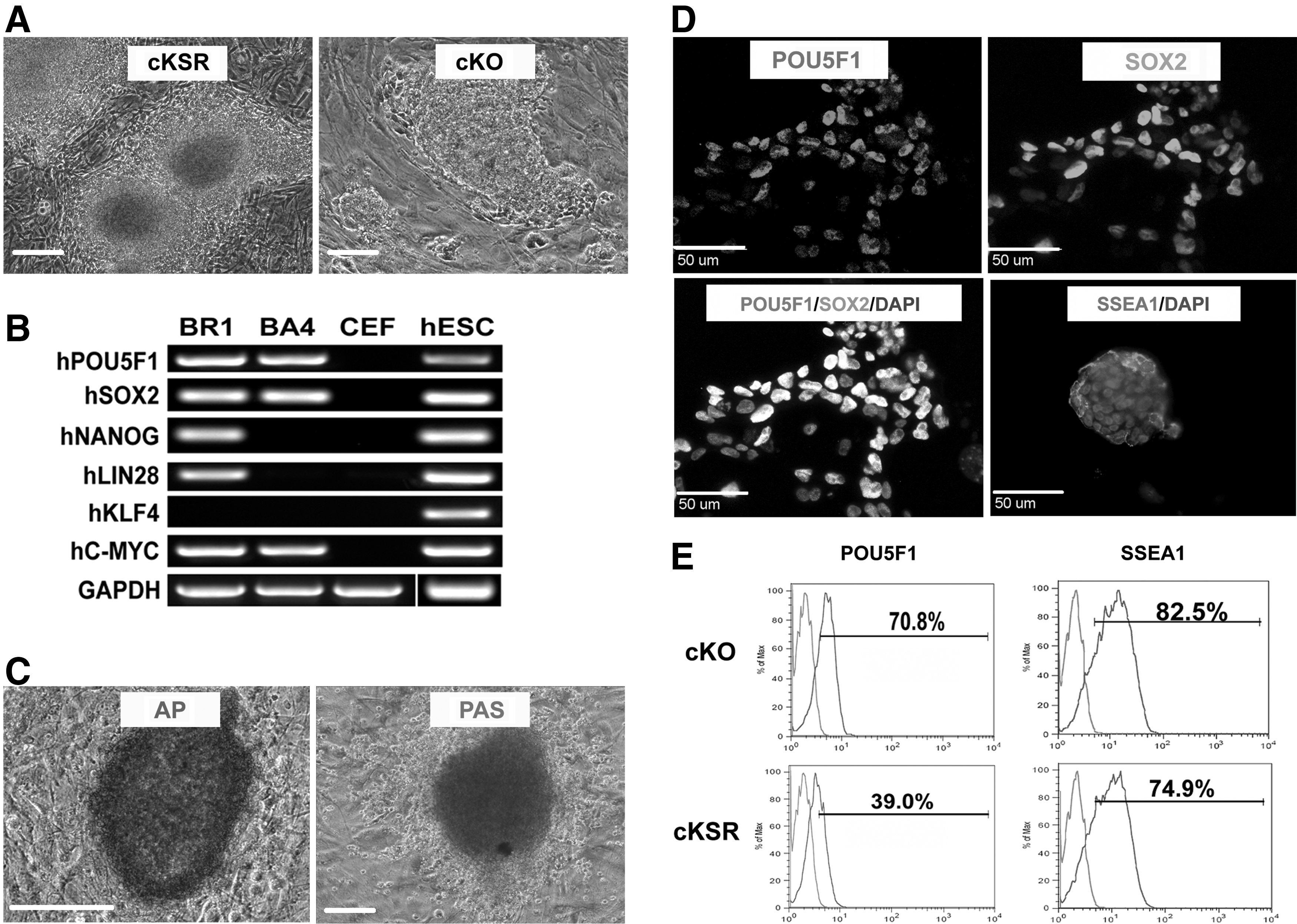
Reprogramming of CEFs into cPSCs requires five factors and the cKO medium for stable propagation. CEFs transduced with human stem cell factors and replated in the cKSR medium and cKO medium formed cPSC colonies in 7 days
We then attempted the reprogramming process utilizing the cKO medium, which has previously been optimized to maintain chicken PGCs in extended culture [18]. Cells were manually isolated from day 7 to 10 after transduction (Fig. 1A). Two lines (BA and BR) of CEFs were transduced with human stem cell factors and both successfully produced cPSCs in this culture system. However, the BA line ceased to grow at passage 10, while the BR line has been successfully propagated up to 40 passages. PCR results revealed that the BA line had only three human genes incorporated (POU5F1, SOX2, and C-MYC), while the BR line had five (hPOU5F1, hSOX2, hNANOG, hLIN28, and hC-MYC) of the six human reprogramming genes successfully incorporated (Fig. 1B). This suggested that additional factors may be needed to fully reprogram the three-factor line. An additional round of transduction of the three-factor BA line generated a five-factor cPSC line that had a similar proliferation competence as the BR line. Morphologically, the established cPSC colonies in the cKO medium were loosely attached to the feeder layer and cells of these colonies demonstrated three-dimensional (3D) structures (Fig. 1A). cPSCs cultured in cKO are highly positive for AP and PAS staining (Fig. 1C). Immunocytochemistry revealed that these cells are positive for POU5F1, SOX2, and SSEA1 (Fig. 1D) indicating pluripotency. Flow cytometry results showed that SSEA1- and POU5F1-positive cells represent 82.5% and 70.8%, respectively, of the whole population (Fig. 1E). To determine if cells cultured in the cKO medium were less prone to spontaneous differentiation than cells in the cKSR medium, we cultured the cKO medium-derived ciPSCs in the cKSR medium. This resulted in a reduction of SSEA1- and POU5F1-positive cells down to 74.9% and 39.0%, respectively (Fig. 1E). Thus, in the following experiments, we used cKO as the preferred culture medium.
cPSCs express endogenous pluripotent genes
Previous reports indicated that ectopic expression of transcription factors resulted in the reactivation of the endogenous pluripotent genes, which was critical for a complete induced pluripotency during cellular reprogramming [26]. Using qPCR for chicken-specific TaqMan primers, chicken PouV, Sox2, Nanog, and Lin28 were highly upregulated in cPSCs, which indicated that the endogenous pluripotent machinery was triggered by the overexpression of the exogenous genes (Fig. 2A). The telomerase reverse transcriptase was also upregulated in cPSCs, indicating the acquisition of immortality in these cells (Fig. 2A). Unexpectedly, the chicken c-Myc was found to be downregulated in cPSCs relative to CEF cells (Fig. 2A) and this finding was confirmed by RT-PCR (Fig. 2B). This c-Myc result may be due to the fact that this was highly expressed in the parent cell line CEFs and not further increased when the exogenous gene was added. The results of RT-PCR also revealed that Klf4 was highly expressed in CEF, comparable to that observed in cPSCs (Fig. 2B). These results indicated that ectopic c-Myc and Klf4 expression might be dispensable in derivation of cPSCs.
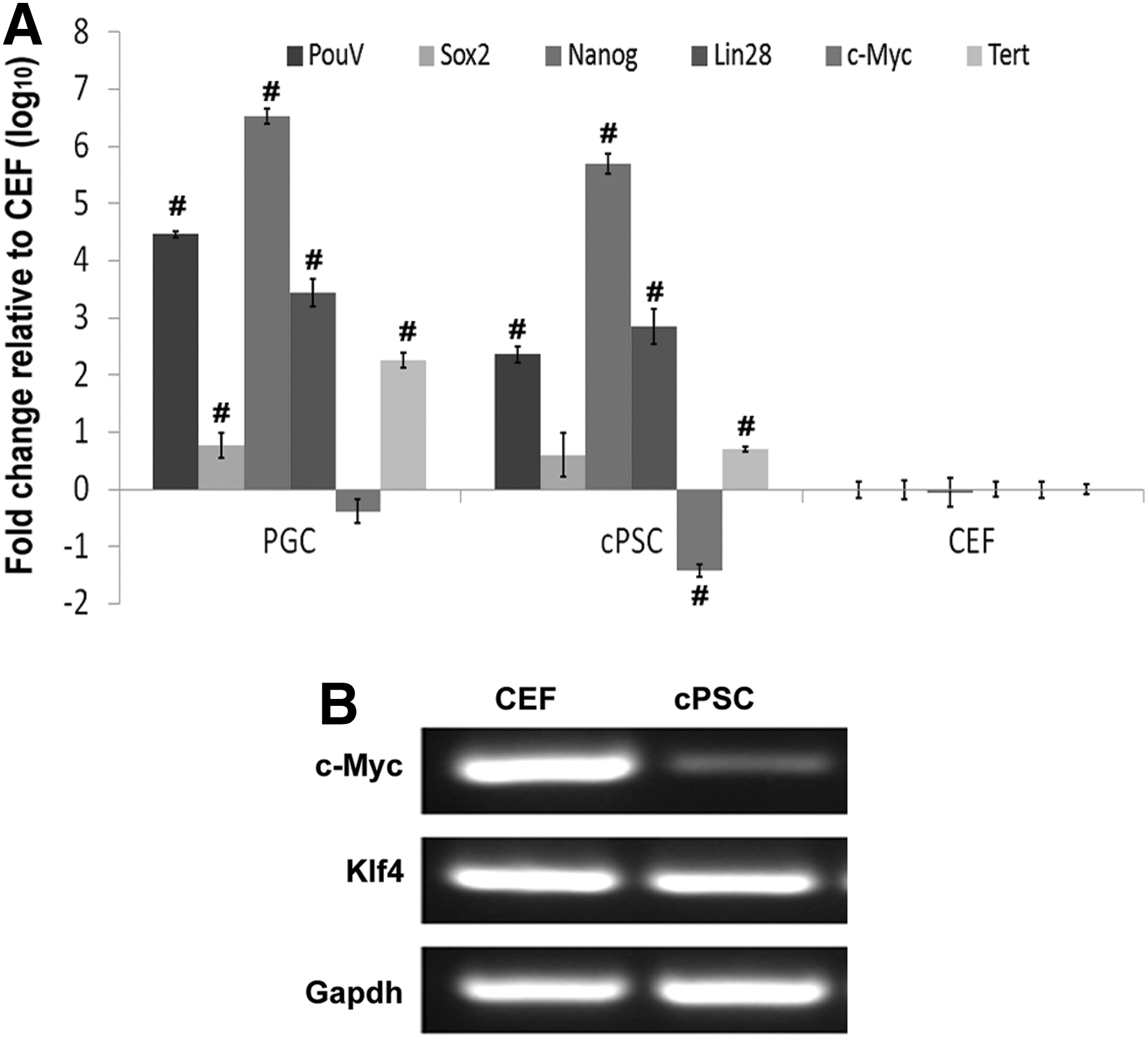
Reactivation of endogenous pluripotent genes in cPSCs. Quantitative PCR showed that the endogenous pluripotent genes PouV, Sox2, Nanog, Lin28, and Tert were highly upregulated, while c-Myc was downregulated in cPSCs. This indicates a significant difference relative to CEFs (P<0.01)
cPSCs express GC-specific markers
cPSCs were derived in a cKO medium that was optimized for GC maintenance. Next, cPSCs were examined for shared morphology and gene expression with cultured PGCs. Quantitative RT-PCR expression of chicken VASA homologue (Cvh) and deleted in azoospermia (Dazl), although lower than in PGCs, were both highly upregulated in cPSCs (Fig. 3A) and c-Kit expression in cPSCs was comparable to PGCs. We then further characterized the expression of GC-related proteins by immunocytochemistry and flow cytometry. cPSCs were positive for EMA1, DAZL, and CVH at the protein level (Fig. 3B), while the parent cell line CEFs were negative for these markers (data not shown). Flow cytometry confirmed that 65% of the cPSCs were positive for EMA1, 92.0% and 93.4% for DAZL and CVH, respectively (Fig. 3C). Moreover, we found that the cells were positive for these GC markers as early as early passage 2 and later at passage 20, which indicated that GC characteristics were well maintained in the current culture system. These results indicated that cPSCs derived in this study acquired a GC fate and are henceforth called ciPGCs.
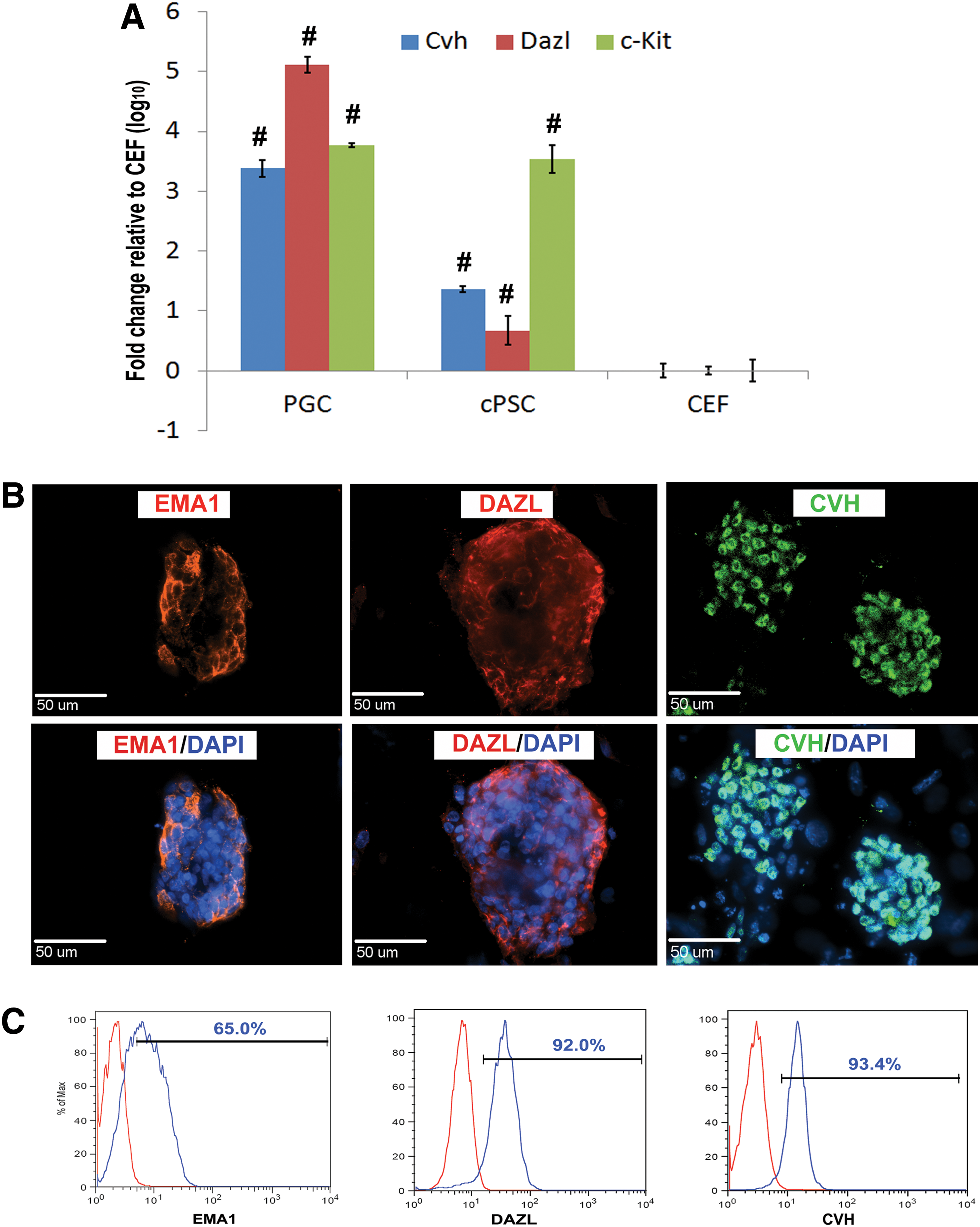
cPSCs express germ cell markers. Quantitative PCR, using PGCs as positive control and CEFs as negative control, revealed that the germ cell-related genes Cvh, Dazl, and c-Kit were upregulated in cPSCs compared to CEFs. This indicates a significant difference relative to CEFs (P<0.01)
ciPGCs are capable of differentiation into all three germ layers in vitro
Chicken PGCs have previously demonstrated a high level of plasticity in vitro and are capable of generating cells from all three germ layers [27,28]. To determine if ciPGCs demonstrate the similar potential, these cells were subjected to in vitro EB differentiation. The recovered aggregates formed compact EBs after suspension culture for 6 days in the differentiation medium (Fig. 4A). RT-PCR analysis showed that the mesoderm (Glut, LPL, and PPAR), ectoderm (TUJ1 and NESTIN), and endoderm (HNF1 and HNF4) genes were highly expressed in ciPGC-derived EBs, but were negative in CEFs (Fig. 4B). Low levels of differentiation lineage marker gene expression were also detected in undifferentiated ciPGCs, which indicated that some cells were undergoing spontaneous differentiation (Fig. 4B). To induce further differentiation, the recovered EBs were replated in Matrigel-coated four-well chamber slides and cultured for an additional 3 days. Results of the immunocytochemistry showed that plated EBs were capable of differentiating into endoderm (SOX17), mesoderm (αSMA), and ectoderm (Hu C/D and SOX1) cell types (Fig. 4C), demonstrating that in vitro ciPGCs possessed developmental plasticity.

iPGCs are capable of differentiation into all three germ layers. iPGCs formed compact EBs after suspension culture for 6 days in the differentiation medium
ciPGCs migrate to the embryonic gonad in vivo
During embryonic development, chicken PGCs migrate through the vasculature system to the gonads at developmental stages 15 to 17 [29]. GC migration has previously been demonstrated to be orchestrated by the SDF-1/CXCR4 chemokine signaling pathway making it important to determine if ciPGCs express CXCR4 [30 –33]. Results of immunocytochemistry showed that CEFs were negative for CXCR4, while both the in vivo-derived PGC-positive control cells and ciPGCs were positive (Fig. 5A). Flow cytometry confirmed that 51% of ciPGCs were positive for CXCR4 (Fig. 5B). To determine the migratory potential of these cells, we labeled the ciPGCs, PGCs, and CEFs with PKH26 and injected them into the vasculature system of stage 15 chicken embryos. All 10 embryos injected with PGCs and 13 out of 18 embryos injected with ciPGCs exhibited migration of exogenous cells to the embryonic gonads. No CEFs were found in the gonads of the injected embryos (nine embryos) (Fig. 5C). Although we also found sporadic PKH26 cells in tissues other than gonads in ciPGC- and PGC-injected embryos, most of them were located in the gonad or the mesentery tissues adjacent to the gonads, which has been documented as the migration route of PGCs before entering the developing gonad [29]. These data demonstrate that the ciPGCs derived in this study acquired a GC-like fate in vitro and are capable of migration to the embryonic gonads.

iPGCs are capable of migrating to the embryonic gonad. Immunocytochemistry showed that CEFs were CXCR4 negative, while both PGCs and iPGCs were positive
Discussion
In this study, we demonstrated for the first time in the avian species, the successful derivation of ciPGCs from a somatic cell line by cellular reprogramming. These cells resemble PGCs in gene expression and protein profiles and are capable of migration to embryonic gonads after injection into stage 15 chicken embryos. This advance represents the first step toward generating germline chimeric individuals derived from somatic cells. As GCs are responsible for passing genetics from one generation to the next, the capability to generate PGCs from somatic cells offers a potentially new strategy for the conservation of endangered birds. This approach also provides scientists a new cell tool to gain insight into the biology of GC development or for PGC studies such as developmental reproductive toxicology using the avian model.
ciPGCs expressed a number of the key GC markers, including CVH, DAZL, and C-KIT, which are GC-specific or highly enriched markers. VASA (or DDX4) is the CVH homologue and is widely conserved and has been demonstrated to be a definitive GC marker in drosophila, xenopus, mice, and human [34 –37]. Similarly, the chicken homologue Cvh has also been found to be specifically expressed in PGCs [19]. In fact, overexpression of Cvh has been demonstrated to drive chicken ESCs to a GC fate [38]. Dazl, a member of the Daz gene family, which encodes RNA-binding proteins, is also specifically expressed in GCs and required for GC development in diverse organisms [39]. Loss of DAZL expression has been closely associated with aberrant gametogenesis and infertility and again the overexpression of this gene in ESCs has lead to the direct reprogramming of ESCs into PGCs, indicating its prominent role in GC development [2,40,41]. c-Kit is a key regulator of PGC development and binding of c-Kit to its ligand activates multiple downstream signaling events (such as MEK/MAPK) and promotes growth and survival of PGCs [42]. In this study, high expression of Cvh, Dazl, and c-Kit in the ciPGCs indicates that the ectopic expression of the transcription reprogramming factors not only induced the pluripotent network in CEFs, as indicated by the upregulation of the endogenous genes, but also triggered the GC-related signaling pathway and resulted in a GC fate in these somatic cells when placed in PGC culture conditions. PGCs possess unique migratory properties in early embryonic development, and the signaling by chemokine receptor CXCR4 and its ligand SDF-1 was reported to be responsible for this migration in mammals and in avians [33]. We found that CXCR4 was expressed in PGCs and ciPGCs, but absent in CEFs. PGCs and ciPGCs injected into stage 15 chicken embryos resulted in significant migration to the embryonic gonads. Chicken ESCs are negative for the GC-associated CXCR4 and are unable to migrate to the putative gonad in a similar manner to PGCs [43]. In total, the cells derived in this study resemble PGCs in gene transcription, protein cell morphology, and in vivo characteristics (Table 1), and thus should be considered ciPGCs.
ciPGCs, chicken induced primordial germ cells; ESC, embryonic stem cell; N/A, not available; PGC, primordial germ cell.
Maintenance of the pluripotency of PGCs in vitro has been challenging and little is known about its underlying mechanism. bFGF has been demonstrated to play a key role in this process by activating the MEK/ERK cell signaling pathway and stimulates the proliferation of PGCs [44]. ciPGCs in the present study were derived and maintained in the cKO that contains bFGF. If bFGF is removed from cKO media, ciPGCs lose their 3D colony morphology and are loosely attached to the feeder layer, but then become more adherent with some invading the feeder layer, a phenomenon similar in the culture of chicken ES cells [45] and the conversion of PGCs into EGCs in the mouse [18]. However, the cKSR medium used for human or chicken ESC [46] culture also contains bFGF, but failed to maintain pluripotency in ciPGCs as evidenced by a significant decrease of POU5F1-positive cells after culture in this medium (Fig. 1E). We also tried to culture the derived ciPGCs in the mTeSR medium, which has been successfully used in quail iPSC derivation [25], but the cells differentiated quickly and ceased to grow after one or two passages. Additional factors in the cKO medium such as those secreted by the BRL cells in the conditioned medium (cKO medium) are believed to be critical for the maintenance of avian pluripotent cells [45].
In the current study, we used the same combination of transcriptional factors to reprogram the chicken cells, as we previously used in quail iPSCs [25]. As in mammals, ectopic expression of the master reprogramming transcription regulators could trigger the endogenous pluripotent network [47]. Successfully induced pluripotent chicken cells in the current study confirmed that this process is widely conserved across vertebrates. The tumor-associated gene c-Myc was reported to be essential in regulation of the pluripotency and cell cycle in PSC lines [47,48]. However, our findings showed that although the endogenous genes (eg, PouV) were highly upregulated by the ectopic transcriptional factors, the c-Myc was significantly downregulated in chicken ciPGCs compared to its parent somatic cells. We also found that expression of c-Myc in PGCs was lower than that observed in CEF cells (Fig. 2A). This suggests that the exogenous c-Myc gene may not be required to induce and maintain pluripotency in CEF-generated chicken ciPGCs, and donor CEF c-Myc levels may be sufficient. However, exogenous c-Myc may be required in other cell types that are reprogrammed, perhaps in adult tissue that would have lower endogenous c-Myc expression. Therefore, results may vary depending on the original cell type that is reprogrammed. Another tumor-related gene Klf4 is a widely used reprogramming factor and when overexpressed can change prime state EpiESCs to the naive state [49]. Interestingly, the human KLF4 failed to be incorporated in any of the chicken ciPGC lines in the present study and this result is similar to the quail iPSCs previously isolated [25]. A recent report also revealed that although the exogenous Klf4 constructed in a polycistronic vector was incorporated into the chicken cell genome and expression was detected in the second passage after transduction, it was completely shutdown in the fifth passage and endogenous Klf4 was not upregulated to CEF cell levels [50]. We observed that endogenous chicken Klf4 was highly expressed in chicken ciPGCs as well as in the parent cell line CEFs. These observations indicate that ectopic Klf4 expression might be dispensable during later stages of the cellular reprogramming and induced pluripotency in chicken cells. Taken together, a combination of exogenous POU5F1, SOX2, NANOG, LIN28, and possibly additional C-MYC, seems to be sufficient for ciPGC induction from CEF cells.
For decades, significant effort has been spent seeking and developing practical means for endangered bird conservation with little avail, since the total number of species is still declining [20]. A recent report revealed that functional gametes could be produced from interspecies transplantation of PGCs, suggesting a new strategy for conservation of endangered birds by using a domestic host [51]. However, this process still requires the sacrifice of endangered embryos for the collection of PGCs. Direct derivation of ciPGCs from somatic cells would circumvent the sacrifice of embryos for PGC isolation and facilitate the implementation of this strategy. Given that ciPGCs can be cryopreserved and, therefore, stored and shipped around the world, captive animal breeding programs might use this technology to introduce genetic diversity into isolated populations or preserve valuable genetics from animals that die.
Avian developmental models are used extensively in research. New insight into neural development [52], ovarian cancer [53], and eye defects [54] has occurred because of the ease of manipulating and observing altered phenotypes in developing avian embryos. However, the lack of a gene editing platform in this species has been a hurdle for researchers. Availability of ciPGCs would allow complex genetic modification at the cellular level and thus would significantly advance the use of this model in research. Genetic engineering in chickens would also be of significant interest in the production of transgenic birds that carry specific traits with agricultural or pharmaceutical importance.
In this report, we demonstrated for the first time the generation of ciPGCs from somatic cells utilizing cellular reprogramming by ectopic expression of transcription factors, thereby laying the foundation for future demonstration of germline transmission in chickens. Availability of competent iPGCs offers new strategies for endangered bird conservation and a new cell source for developmental biology and transgenic animal research.
Footnotes
Acknowledgments
The authors would like to thank Julie Nelson at the Center for Tropical and Emerging Global Diseases Flow Cytometry Facility for her assistance in FACS sorting and flow cytometry analysis. This work was supported by the Bill and Melinda Gates Foundation (OPP1027273) and the Guangxi International Collaborative Project (1347004-6).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.