
Abstract
Bone marrow mesenchymal stem cells (BMMSCs) have been used to treat a variety of autoimmune diseases in clinics. However, the therapeutic effects are largely dependent on the immunomodulatory capacity of culture-expanded BMMSCs. In the present study, we show that aspirin (acetylsalicylic acid, ASA)-treated BMMSCs have significantly improved immunomodulatory function, as indicated by upregulation of regulatory T cells (Tregs) and downregulation of Th17 cells via the 15d-PGJ2/PPARγ/TGF-β1 pathway. Furthermore, the therapeutic effect of ASA-pretreated BMMSCs was confirmed in a dextran sodium sulfate-induced experimental colitis mouse model, in which systemic infusion of ASA-pretreated BMMSCs significantly ameliorated disease activity index and colonic inflammation, along with an increased number of Tregs and decreased number of Th17 cells. Taken together, our results suggest that aspirin treatment is a feasible strategy to promote BMMSC-based immunomodulation.
Introduction
M
Recent studies revealed that FAS-regulated monocyte chemotactic protein-1 (MCP-1) secretion by MSCs could recruit T cells to initiate FAS ligand (FASL)-mediated apoptosis, and apoptotic T cells triggered macrophages to produce high levels of transforming growth factor-β (TGF-β), leading, in turn, to the upregulation of regulatory T cells (Tregs) and, subsequently, immune tolerance [11]. Although it has been accepted that MSCs exert immunosuppressive properties that could be induced by proinflammatory cytokines, such as interferon-γ (IFN-γ) [12], the effects of MSCs on immune-related disorders have been challenged in several in vivo studies [13 –15], especially the claim that MSCs function might be impaired by an inflamed microenvironment [16,17]. This partly results from their poor survival rate and impaired immunoregulatory properties when exposed to an inflamed tissue microenvironment, which hinders the clinical application of MSCs in treating immune-related diseases. To improve the therapeutic efficacy of MSCs, genetic modification has been applied to enhance their immunomodulatory properties [18,19], typically through increasing Tregs, but inhibiting Th17 cells, in the peritoneal cavity and spleen [19]. However, the use of genetically modified MSCs raises a major safety concern.
Aspirin, also known as acetylsalicylic acid (ASA), is a widely used nonsteroidal anti-inflammatory drug (NSAID) that plays important roles in multiple biological pathways, including irreversible inactivation of cyclooxygenase-2 (COX-2) enzyme [20 –22]. Our previous studies found that ASA could significantly promote bone marrow MSC (BMMSC)-based bone regeneration via reducing the local concentrations of IFN-γ and tumor necrosis factor-α (TNF-α) [23]. ASA suppressed maturation and subsequent immunostimulatory function of dendritic cells and determined the fate of naïve T cells to regulatory phenotypes, which suggested its immunoregulatory potential in the context of immune tolerance [24,25]. It was therefore hypothesized that ASA-pretreated BMMSCs would have a higher immunomodulatory ability. In this study, we found that ASA could significantly enhance the immunoregulatory properties of BMMSCs via upregulating Tregs and downregulating Th17 cells through the 15d-PGJ2/PPARγ/TGF-β1 pathway. These results have also been verified in an experimental colitis mouse model. These data lend support to the notion that ASA may offer a feasible and effective therapeutic approach for treating immune-related disorders.
Materials and Methods
Animals
All animal experiments were performed under the institutionally approved protocols for the use of animal research (Capital Medical University # 2012-x-53). Female C57BL/6J mice were purchased from Vital River and housed in a specific pathogen-free animal facility under controlled temperature (25°C) and photoperiods (12:12-h light-dark cycle) and allowed unrestricted access to standard diet and tap water. Mice were allowed to acclimate to these conditions for 7 days before inclusion in experiments.
Antibodies and chemicals
Unconjugated antibody to peroxisome proliferator-activated receptors gamma (PPARγ), anti-rat IgG-Alexa Fluor 488 antibody, and anti-rat IgG-Alexa Fluor 555 antibody were purchased from Cell Signaling Technology. Anti-CD3ɛ, -CD28, -CD4-PerCP, and -CD25-APC antibodies were purchased from BD Biosciences. Recombinant mouse IL-6, unconjugated antibodies to IL-17, unconjugated antibodies to Foxp3, anti-Foxp3-PE, anti-IL-17-PE antibodies, and anti-TGF-β1-neutralizing antibodies were purchased from BioLegend. Unconjugated antibodies to Histone H1 were purchased from BioWorld. ASA, 15d-PGJ2, GW9662, PGE2, rosiglitazone, and anti-β-actin antibody were purchased from Sigma-Aldrich.
Kits
Cell Counting Kit-8 (CCK-8) was purchased from Dojindo Molecular Technologies. CD4+ T Cell isolation kit was purchased from Miltenyi Biotec. Annexin V-PI Apoptosis Detection Kit was purchased from BD Bioscience. Mouse IL-10, IL-17, and TGF-β1 ELISA kits were purchased from Dakewei Biotech. PGE2 and 15d-PGJ2 EIA kits were purchased from Enzo Life Sciences. Total Nitric Oxide and Nitrate/Nitrite Parameter Assay Kit was purchased from R&D Systems.
Isolation of mouse BMMSCs
BMMSCs were isolated as previously described [23]. In brief, bone marrow cells were flushed out from bone cavity of femurs and tibias with 2% heat-inactivated fetal bovine serum (FBS; Equitech-Bio) in phosphate-buffered saline (PBS). Single-cell suspension of all nuclear cells (ANC) was obtained by passing all bone marrow cells through a 70-μm cell strainer (BD Bioscience). Ten to fifteen million cells were seeded onto 10 cm culture dishes (Corning) and initially incubated for 48 h with alpha minimum essential medium (α-MEM; Invitrogen) supplemented with 20% FBS, 2 mM L-glutamine (Invitrogen), 55 μM 2-mercaptoethanol (Invitrogen), 100 U/mL penicillin and 100 μg/mL streptomycin (Invitrogen) at 37°C and 5% CO2 in a humidified environment. To eliminate nonadherent cells, cultures were washed with PBS twice. The primary culture of colony-forming cells was incubated for 16 days. Cells were passaged once they became 70% to 80% confluent. BMMSCs at passage 2 were used in this study.
Cell survival assay for BMMSCs
The rate of cell proliferation was investigated using trypan blue exclusion and CCK-8 assay. For the trypan blue exclusion assay, after 3×105 BMMSCs were seeded on six-well culture plates (Corning) and incubated overnight, cells were treated with various concentrations of ASA (50–800 μg/mL) for 24 h. The number of viable cells was counted using a hemocytometer.
Alternatively, cell proliferation rate was assayed using CCK-8 (Dojindo). Briefly, 1×104 BMMSCs were seeded on 96-well culture plates (Corning) and treated with ASA for 24 h, as mentioned above. OD value at 450 nm was evaluated using an ELISA Reader (Promega) following the manufacturer's instructions.
T lymphocyte isolation
CD4+ T lymphocytes were isolated from mouse total spleen cells using a magnetic sorter with a mouse CD4+ T Cell isolation kit (Miltenyi Biotec) following the manufacturer's instructions.
Coculture of BMMSCs and spleen (SP) cells
Mouse CD4+ T cells were activated with plate-bound antibody to CD3ɛ at 2 μg/mL and soluble antibody to CD28 at 2 μg/mL (BD Bioscience) in complete 1640 medium (Invitrogen) supplemented with 10% FBS for 48 h. Mouse BMMSCs at passage 2 (2×105) were seeded on 12-well culture plates (Corning) and incubated overnight before they were pretreated with ASA for 24 h. Activated CD4+ T cells were loaded on mouse BMMSCs, and they were cocultured for 72 h. Recombinant mouse IL-6 (50 ng/mL) (BioLegend) was added to induce the production of Th17 cells. Ten μM PGE2 or 10 μM 15d-PGJ2 (Sigma-Aldrich) was added in the coculture system. Subsequently, cells in suspension and in culture supernatant were collected. Cells in suspension were analyzed using flow cytometry (FACSCalibur; BD Bioscience) to examine the number of Treg and Th17 cells. The supernatant was collected to analyze levels of soluble factors following the manufacturer's instructions.
Detection of soluble factors
Concentrations of IL-10, IL-17, and TGF-β1 in coculture supernatants were assayed by ELISA kits (Dakewei Biotec). Concentrations of 15d-PGJ2 and PGE2 were measured by EIA kit (Enzo). Levels of nitric oxide were evaluated by assay for nitrates using modified Griess reagents (R&D Systems). All assays were performed following the manufacturer's instructions.
Flow cytometric analysis
To detect the ratio of Tregs, floating cells were stained with anti-mouse CD4-Percp and CD25-APC (BD Bioscience) for 30 min. After fixation and permeabilization, cells were stained with anti-mouse Foxp3-PE (BioLegend) according to the manufacturer's protocols. To detect the ratio of Th17 cells, cells were stimulated with 5 ng/mL Phorbol 12-myristate 13-acetate (PMA) (eBioscience) and 500 ng/mL ionomycin (eBioscience) for 4 h in the presence of a protein-transport inhibitor (Golgi Stop; BD Bioscience). Cells were then incubated with anti-mouse CD4-Percp (BD Bioscience). After fixation and permeabilization, cells were stained with anti-mouse IL-17-PE (BioLegend). To detect the ratio of apoptotic cells, cells were harvested, and BMMSCs (1×105) were resuspended in 100 μL binding buffer with 5 μL Annexin V-FITC and 5 μL PI solution (BD Bioscience) and incubated for 15 min in the dark. Cell preparations were immediately analyzed using flow cytometric analysis (FACSCalibur; BD Bioscience). These experiments were performed at 4°C in the dark.
Neutralization experiments
Anti-TGF-β1 neutralizing antibody at 1 μg/mL (BioLegend) was added to the coculture system of ASA-pretreated BMMSCs and activated CD4+ T cells.
Inhibition and activation of PPARγ
To inhibit or activate PPARγ, an antagonist or agonist of PPARγ was added into the culture medium, respectively. GW9662 (Sigma-Aldrich), an antagonist of PPARγ, was added at 1 μM 1 h before pretreating BMMSCs with ASA (200 μg/mL). Either rosiglitazone or 15d-PGJ2, agonists of PPARγ (Sigma-Aldrich), was added at 10 μM into the culture medium when pretreating BMMSCs with ASA (200 μg/mL).
Western blot analysis
Total protein was extracted using M-PER mammalian protein extraction reagent (Thermo). Nuclear protein was obtained using NE-PER nuclear and cytoplasmic extraction reagent (Thermo). Fifty to 100 μg of protein were separated on 10% polyacrylamide-SDS gels (Applygen) and transferred to Immobilon™-P membranes (Millipore). After blocking with TBS/5% nonfat dry milk (Applygen) for 1 h, the membrane was incubated with antibodies against mouse PPARγ (Cell Signaling Technology), Histone H1 (BioWorld), and β-actin (Sigma) overnight at 4°C. This was followed by incubation with HRP-conjugated secondary antibodies (Pierce) for 1 h at room temperature. Antibody binding was visualized by an enhanced chemiluminescence kit according to the manufacturer's protocols (Pierce).
Acute colitis mice model
Acute colitis was induced in 8-week-old C57BL/6 mice by administration of 3% (w/v) dextran sodium sulfate (DSS) (molecular mass 36–50 kDa; MP Biochemicals) to drinking water for 10 days [26]. BMMSCs (1×106) pretreated with 200 μg/mL ASA, or untreated BMMSCs, were injected into colitis mice (n=5) via tail vein at day 3 after feeding with DSS water. Mice were injected with PBS as a control group (n=5). Mice were matched by age and body weight for each group. Mice were sacrificed at day 10, and samples were analyzed as previously described [27].
Clinical progression of the disease was assessed by disease activity index (DAI). The following parameters were used for calculation: (a) weight loss (0 point=none, 1 point=1%–5% weight loss, 2 points=5%–10% weight loss, 3 points=10%–15% weight loss and 4 points=more than 15% weight loss); (b) stool consistency/diarrhea (0 points=normal, 2 points=loose stools, 4 points=watery diarrhea); (c) bleeding (0 points=no bleeding, 2 points=slight bleeding, 4 points=gross bleeding). DAI was calculated as the total of these scores: the sum of weight loss, diarrhea, and bleeding, resulting in a total DAI score ranging from 0 (unaffected) to 12 (severe colitis).
For histological examination, the degree of inflammation and epithelial damage in distal colon was graded with hematoxylin and eosin-stained sections (5 μm). Assessment of histological activity index (HAI) was based on the loss of goblet cells and crypts and infiltration of inflammatory cells [27]. The following parameters were used for calculation of HAI: (a) Epithelial damage (0 point=none, 1 point=minimal loss of goblet cells, 2 points=extensive loss of goblet cells, 3 points=minimal loss of crypts and extensive loss of goblet cells, and 4 points=extensive loss of crypts); (b) Infiltration (0 point=none, 1 point=infiltration around crypt bases, 2 points=infiltration in muscularis mucosa, 3 points=extensive infiltration in muscularis mucosa with edema, and 4 points=infiltration of submucosa). HAI was calculated as the sum of the epithelium and infiltration score, resulting in the total HAI score ranging from 0 (unaffected) to 8 (severe colitis).
Immunofluorescent staining
Colons of acute colitis were harvested, fixed in 4% paraformaldehyde, and embedded with paraffin. Sections were blocked with 5% normal serum and incubated with IL-17 or Foxp3 antibodies (BioLegend) or isotype-matched control overnight at 4°C. Samples were then incubated with Alexa Fluor 555- or 488-conjugated secondary antibodies (Cell Signaling Technology). After sections were counterstained with 4′,6-diamidino-2-phenylindole (DAPI; Vector) and mounted with glycerin, they were observed under a fluorescence microscope (OLYMPUS). Semi-quantification of Foxp3+ or IL-17+ cells was performed by positive fluorescence staining normalized to DAPI-stained nuclei under five random fields of view.
Statistical analysis
Statistical analysis was carried out using SPSS 13.0 software. Results were expressed as mean±standard deviation (SD). Statistical significance of *P<0.05 was determined using independent two-tailed Student's t-test or analysis of a one-way variance (ANOVA).
Results
ASA treatment improved immunomodulatory properties of BMMSCs by upregulating Tregs and downregulating Th17 cells
To choose a suitable concentration of ASA, different doses of ASA (50–800 μg/mL) were used to pretreat BMMSCs for 24 h. Trypan blue exclusion and CCK-8 assay were used to evaluate cell proliferation. Cell ratio of apoptosis was shown in the percentage of Annexin V-positive population in flow cytometery. ASA at low concentration (50–200 μg/mL) had no effect on the proliferation or viability of BMMSCs (Fig. 1A–C), while ASA at 400 and 800 μg/mL could significantly inhibit cell proliferation, and induce cell apoptosis (P<0.05) (Fig. 1A–C). We used 50–200 μg/mL ASA to treat BMMSCs in the following study.

Effects of aspirin (acetylsalicylic acid, ASA) on the proliferation and apoptosis of bone marrow mesenchymal stem cell (BMMSCs). ASA at low concentration (50, 100, or 200 μg/mL) had no effect on the proliferation or viability of BMMSCs, while ASA at 400 and 800 μg/mL could significantly inhibit cell proliferation and induce cell apoptosis, as shown in the trypan blue exclusion assay
We next investigated whether ASA treatment could promote the immunomodulatory properties of BMMSCs. We found that pretreated BMMSCs (50–200 μg/mL ASA in 24 h) could not enhance the ability of BMMSCs to induce the apoptosis of activated T cells (Fig. 2A). However, ASA at 200 μg/mL, but not 50 or 100 μg/mL, significantly promoted BMMSCs-mediated upregulation of Tregs (P<0.01) (Fig. 2B) and downregulation of Th17 cells (P<0.05) (Fig. 2C). BMMSCs pretreated with 200 μg/mL ASA significantly increased the concentration of IL-10 (P<0.01) (Fig. 2D) and decreased the concentration of IL-17 (P<0.05) (Fig. 2E) in the supernatants of BMMSCs/T cell coculture system. These data suggested that 200 μg/mL ASA could improve the immunomodulatory properties of BMMSCs via upregulating Tregs and downregulating Th17 cells in vitro.
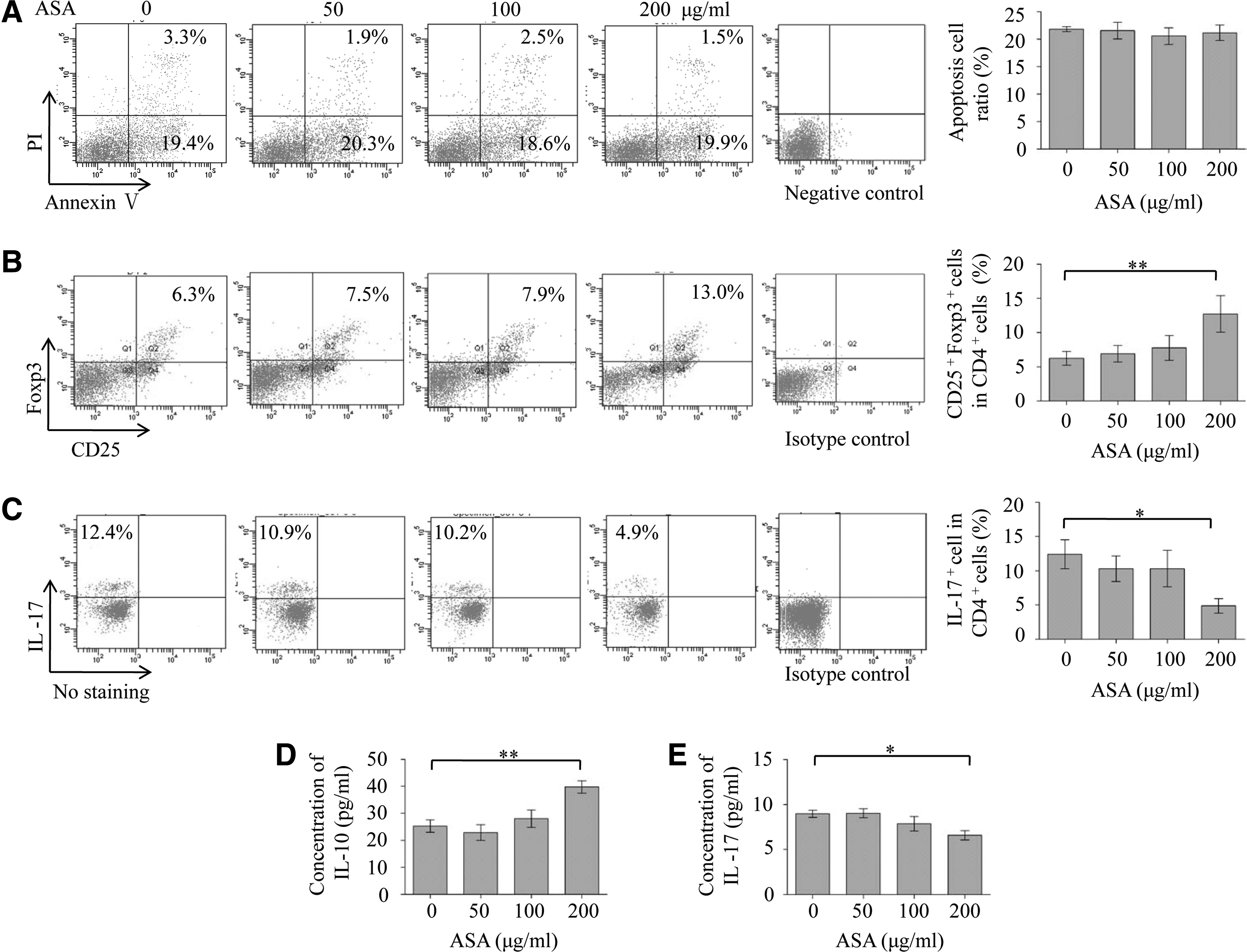
ASA at 200 μg/mL promoted immunomodulatory properties of BMMSCs by upregulating Tregs and downregulating Th17 cells.
ASA promoted immunomodulatory properties of BMMSCs via the 15d-PGJ2/PPARγ/TGF-β1 pathway
To explore the mechanisms of ASA underlying the immunomodulatory properties of BMMSCs, we analyzed expression levels of some soluble molecules associated with the immunomodulation function of BMMSCs, including TGF-β1, nitric oxide, IL-10, PGE2, and 15d-PGJ2, in supernatants of ASA-pretreated BMMSCs. We found that ASA treatment did not influence the production of nitrates (Supplementary Fig. S1A; Supplementary Data are available online at
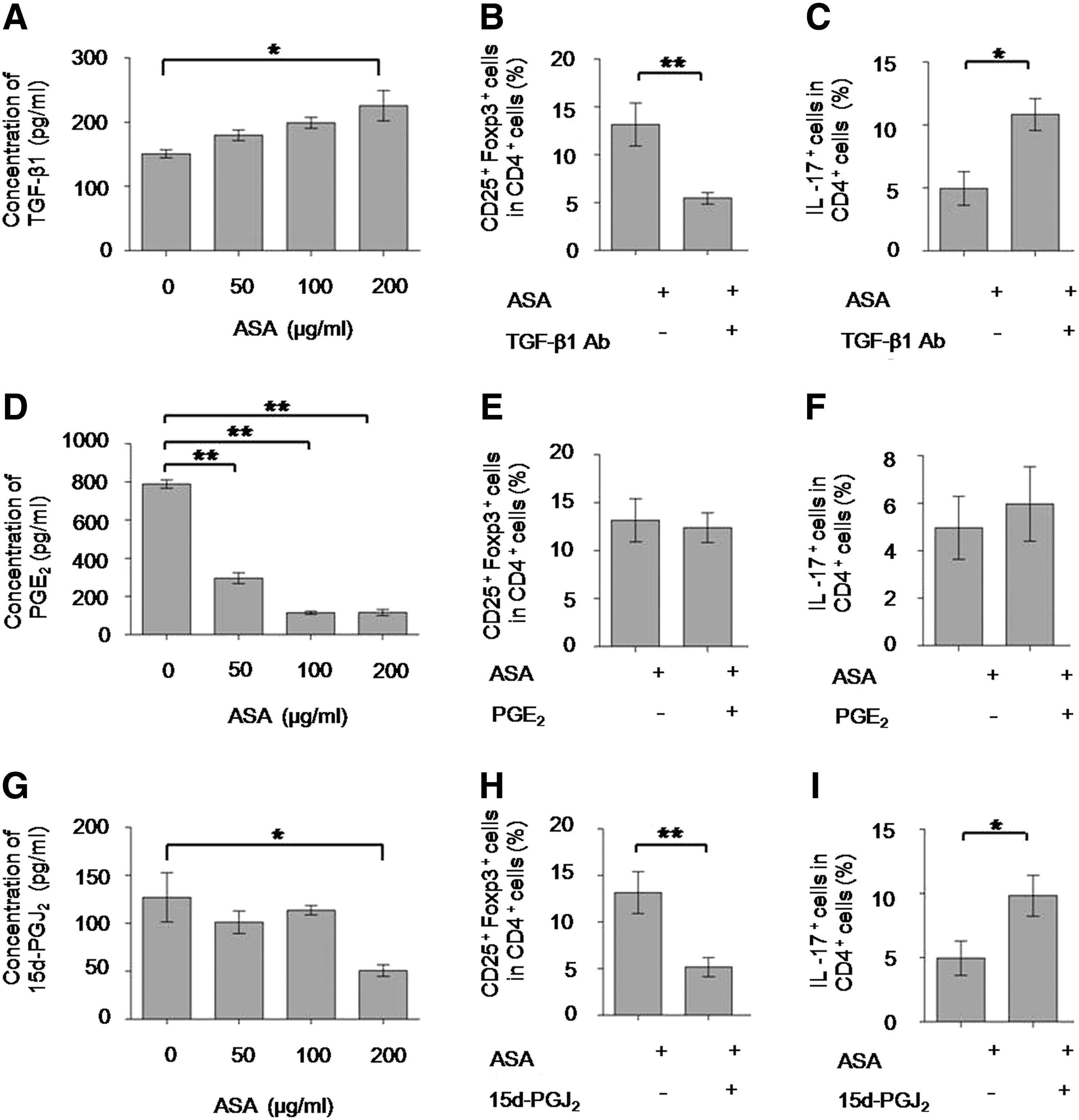
TGF-β1 and 15d-PGJ2 played pivotal roles in the elevated immunomodulatory properties of BMMSCs, as induced by ASA.
We found that the concentration of PGE2 and 15d-PGJ2 was significantly decreased in the presence of 200 μg/mL ASA, as shown in Fig. 3D (P<0.01) and Fig. 3G (P<0.05), respectively. This is consistent with previous reports, which found that ASA could inhibit the production of PGE2 and 15d-PGJ2 [21,22]. Therefore, we added PGE2 and 15d-PGJ2 in the BMMSCs/T cells coculture system. Interestingly, 15d-PGJ2, but not PGE2, could reverse the ASA-mediated elevation of BMMSCs immunomodulatory functions (P<0.05) (Fig. 3E, F, H, I). These data suggested that increasing levels of TGF-β1, or decreasing levels of 15d-PGJ2, induced by 200 μg/mL ASA contributed to BMMSC-based upregulation of Tregs and downregulation of Th17 cells.
Next, we investigated how TGF-β1 and 15d-PGJ2 interacted in ASA-treated BMMSCs. Following ASA treatment, an increased concentration of 15d-PGJ2 significantly blocked the secretion of TGF-β1 by BMMSCs (P<0.05) (Fig. 4A), while anti-TGF-β1 neutralizing antibody had no effect on 15d-PGJ2 production (Fig. 4B).These data indicated that decreasing levels of 15d-PGJ2 contributed to increasing levels of TGF-β1 following ASA treatment, which ultimately mediated BMMSC-based upregulation of Tregs and downregulation of Th17 cells.
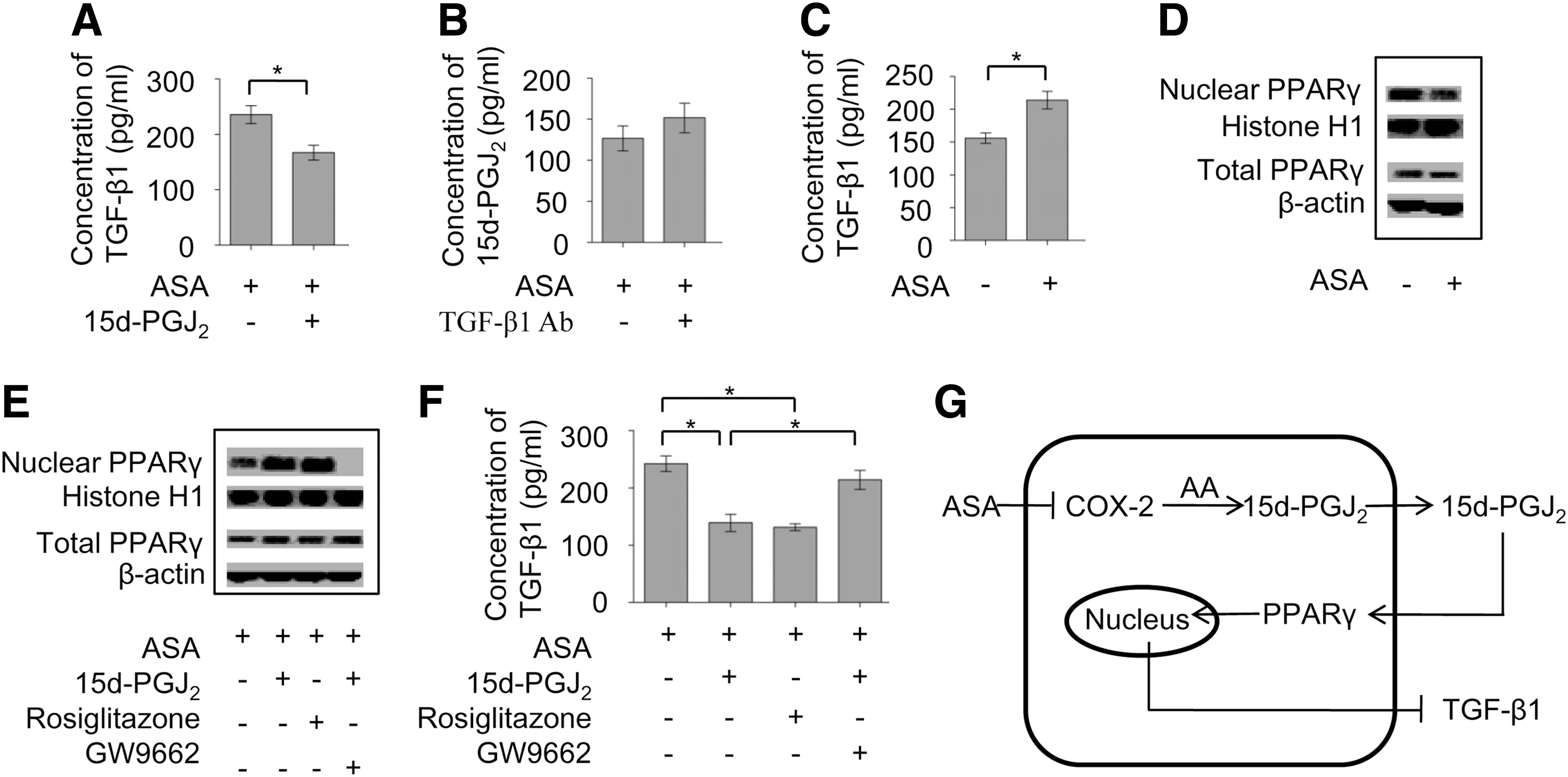
ASA treatment promoted immunomodulatory properties of BMMSCs via the 15d-PGJ2/PPARγ/TGF-β1 pathway.
It was reported that Peroxisome proliferator-activated receptor gamma (PPAR-γ or PPARG) could be activated by its endogenous ligand 15d-PGJ2 through binding to PPARγ in cytoplasm, subsequently contributing to nuclear translocation of PPARγ and affecting TGF-β1 gene transcription [28]. We also found that the elevation of TGF-β1 (P<0.05) was accompanied by a decline of PPARγ translocation to nuclear after ASA treatment (Fig. 4C, D), although the levels of total PPARγ remained unchanged (Fig. 4D). To explore the role of PPARγ in regulating TGF-β1 production, an agonist (rosiglitazone or 15d-PGJ2) or an antagonist (GW9662) of PPARγ was used in conjunction with ASA. In the presence of ASA, rosiglitazone or 15d-PGJ2 treatment promoted the translocation of PPARγ to nuclear, while GW9662 treatment blocked PPARγ translocation to nuclear (Fig. 4E). The upregulation of TGF-β1 after ASA treatment could be blocked by rosiglitazone or 15d-PGJ2, whereas this blockage could be reversed by GW9662 pretreatment (P<0.05) (Fig. 4F). Collectively, these data indicated that ASA treatment improved the immunoregulatory properties of BMMSCs via the 15d-PGJ2/PPARγ/TGF-β1 pathway (Fig. 4G). ASA could irreversibly inhibit the activity of COX-2 and thus lead to the downregulation of 15d-PGJ2. Decreasing levels of 15d-PGJ2 could inhibit the nuclear translocation of PPARγ and promote the gene transcription of TGF-β1.
ASA improved the immunotherapeutic effects of BMMSCs in colitis mice
The in vivo immunomodulatory properties of ASA-pretreated BMMSCs were investigated in a DSS-induced experimental colitis mouse model [26,27]. Experimental colitis was induced by oral administration of 3% DSS in drinking water for 10 days, where 1×106 BMMSCs (Group M) or 1×106 BMMSCs treated with 200 μg/mL ASA (Group ASA-M) were injected at day 3, followed by sacrifice of mice at day 10 (Fig. 5A). Naïve mice (Group N) or colitis mice without any treatment (Group D) were established as controls. An overall assessment of DAI was based on the presence of weight loss, diarrhea, and bleeding. Assessment of histological activity index (HAI) of induced colitis was based on the loss of goblet cells and crypts and the infiltration of inflammatory cells [27]. Histologically, experimental colitis is characterized by severe colonic transmural inflammation with increased wall thickness, infiltration of inflammatory cells, degeneration of crypt architecture, and loss of goblet cells (Fig. 6A–group D). Compared with untreated BMMSCs, ASA-treated BMMSCs had better protective effects against tissue damage from DSS-induced experimental colitis, as shown by decreased DAI (P<0.05) (Fig. 5B), alleviation of colonic inflammation, and colitis HAI (P<0.05) (Fig. 6A, B). These findings illustrated that ASA treatment enhanced the therapeutic effects of BMMSCs on DSS-induced experimental colitis.
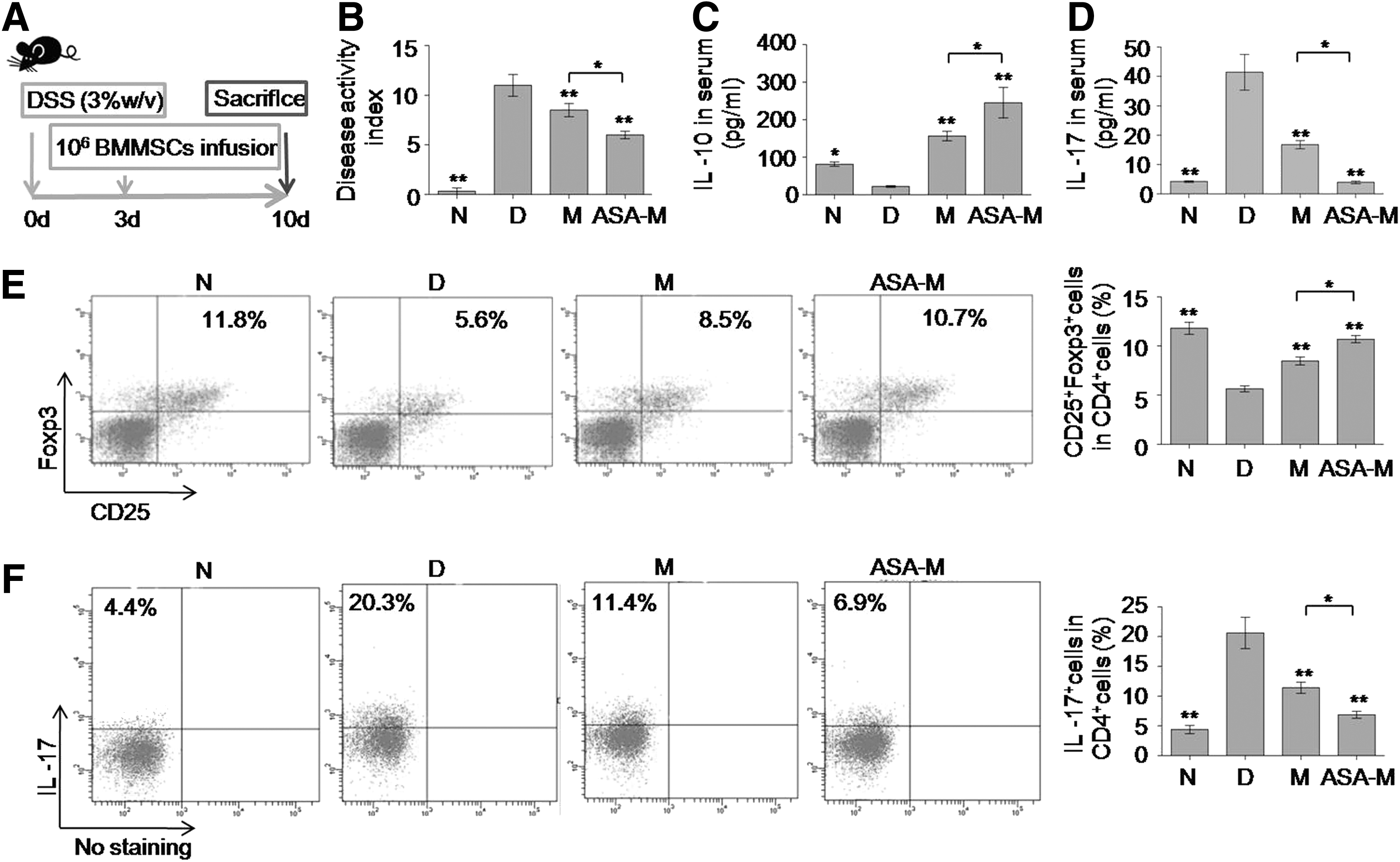
ASA treatment promoted immunomodulatory properties of BMMSCs in vivo.
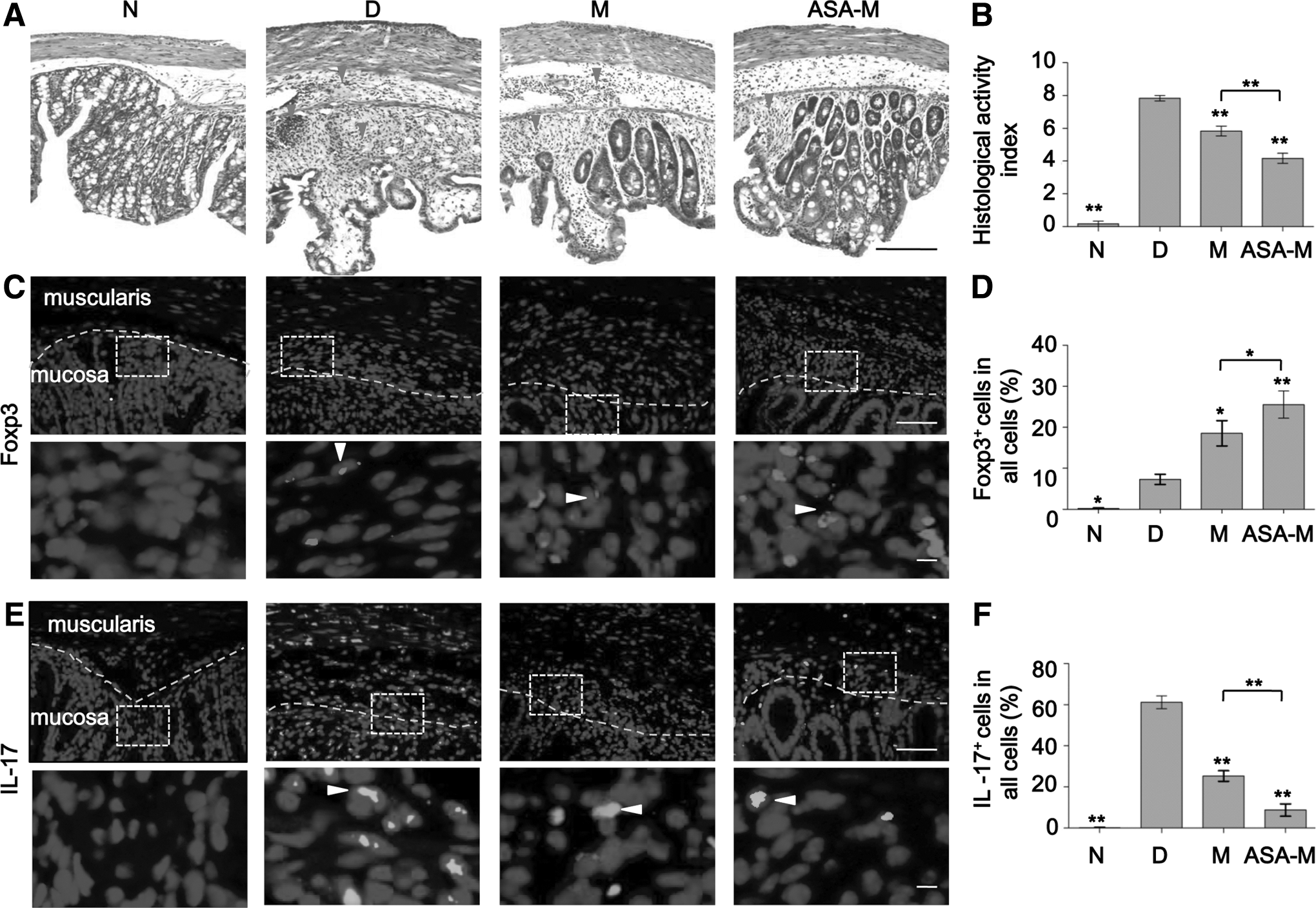
BMMSCs pretreated with ASA (200 μg/mL) showed favorable therapeutic effects in DSS-induced experimental colitis.
We next examined whether ASA treatment could alleviate local and systemic inflammatory cell response in vivo. Compared with untreated BMMSCs, the infusion of ASA-pretreated BMMSCs increased the levels of serum IL-10 (P<0.05) (Fig. 5C) and the ratio of Tregs (P<0.05) (Fig. 5E), while it decreased the levels of serum IL-17 (P<0.05) (Fig. 5D) and the ratio of Th17 cells (P<0.05) (Fig. 5F). Compared with untreated BMMSCs, infusion of ASA-pretreated BMMSCs locally increased the number of Tregs (P<0.05) and decreased the number of Th17 cells (P<0.01), which was demonstrated by the number of Foxp3- and IL-17-positive cells in colonic sites, respectively (Fig. 6C–F). Taken together, these data suggested that ASA-pretreated BMMSCs could alleviate colitis in mice by increasing Tregs and decreasing Th17 cells both in systemic and local colonic sites.
Discussion
By their immunomodulatory properties, BMMSCs hold great promise in clinical applications for treating immune-related diseases. However, the precise mechanism underlying these effects remains poorly understood. It has been reported that the balance between Treg and Th17 cells plays critical roles in maintaining immune tolerance [11,29]. ASA is a NSAID that has been widely used for a variety of clinical applications to relieve pain, reduce fever, help prevent heart attacks, strokes and blood clot formation, and reduce inflammation. Here, we have identified ASA-pretreated MSCs as a novel therapeutic strategy for treating immune-related disorders. In this study, we reported that ASA treatment can improve the immunomodulatory properties of MSCs in vitro, as shown by the upregulation of Tregs and the downregulation of Th17 cells via the 15d-PGJ2/PPARγ/TGF-β1 pathway. Our data also showed that ASA treatment significantly rescued the disease phenotype of experimental colitis in mice, which is accompanied by the upregulation of Tregs and the downregulation of Th17 cells. Collectively, the present study has demonstrated the feasibility and efficiency of using ASA-pretreated MSCs for treating immune-related disorders in future clinical applications.
In the presence of inflammatory tissue, MSCs have exhibited impaired immunoregulatory properties, prompting efforts to promote their immunoregulatory properties, such as the use of genetically modified cells or systemic infusion of regulatory T cells. However, such strategies remain a continuing safety concern for both practitioners and patients. In contrast, the administration of aspirin avoids the risks associated with the above methods, as aspirin has been used for clinical applications for decades with a known side effect profile. While ASA at high concentration has been demonstrated to have anti-proliferative effect on BMMSCs [30,31], we demonstrated that ASA at low concentration (200 μg/mL) did not induce the apoptosis of BMMSCs. Instead, it significantly promoted the immunoregulatory properties of BMMSCs. Recent publication has showed that mice BMMSCs pretreated with ASA at 50 μg/mL exhibited improved immunomodulatory properties via upregulation of telomerase [32]. The different aspirin dose proposed may be attributed to the different time for ASA pretreatment. Specifically, we pretreated BMMSCs with ASA for 24 h in our study while they pretreated BMMSCs with ASA for 3 days. In addition, we have also examined the effects of different dose of aspirin (0, 50, 100, and 200 μg/mL) on human BMMSCs (hBMMSCs) and human periodontal ligament stem cells (hPDLSCs). Results showed that both 50 μg/mL and at 100 μg/mL aspirin significantly increased TGF-β1 production of hBMMSCs, while only 100 μg/mL aspirin could significantly promote TGF-β1 production of hPDLSCs (data not shown). Thus, MSCs from different species or tissue show different dose response when treated with aspirin.
It is believed that ASA plays important roles in the process of bone metabolism. Human epidemiological studies have revealed that ASA could improve bone mineral density [33]. ASA was also reported as a promising approach to treat osteoporosis via increasing the osteogenesis of BMMSCs by aiming at telomerase activity and inhibiting osteoclast activity [34].We have recently shown that ASA could significantly promote BMMSC-based bone regeneration via reducing the local concentrations of IFN-γ and TNF-α [23]. In terms of the effects of ASA on immunomodulation, ASA suppressed maturation and, subsequently, the immunostimulatory function of dendritic cells, driving the fate of naïve T cells toward their regulatory phenotypes, which suggests its immunoregulatory potential in the context of immune tolerance [24,25]. Here, for the first time, we reported that ASA could improve the immunomodulatory properties of BMMSCs, including upregulation of Tregs and downregulation of Th17 cells. Importantly, ASA treatment could enhance the therapeutic effects of BMMSCs in DSS-induced experimental colitis, as shown by its protection against tissue damage and the alleviation of colonic inflammation.
The secretion of soluble factors, including TGF-β1, IL-10, nitric oxide, PGE2, and 15d-PGJ2, has been shown to play important roles in MSC-associated immunomodulation [12,26,35,36]. In this study, we confirmed that the secretion level of TGF-β1 was critical in ASA-associated BMMSCs immunomodulation. ASA could promote the immunomodulatory properties of BMMSCs via the 15d-PGJ2/PPARγ/TGF-β1 pathway. However, the mechanisms that underlie PPARγ mediation of TGF-β1 repression were complex and unclear. It is reported that activated PPARγ could directly upregulate the levels of PTEN, leading to inhibition of p70 ribosomal S6 kinase-1 (S6K1) and dephosphorylation of zinc finger transcription factor-9 (Zf9), consequently causing TGF-β1 repression [29]. In addition, secretion of TGF-β1 was modulated by various pathways. Except the 15d-PGJ2/PPARγ/TGF-β1 pathway, aspirin may also affect other signaling paths to affect production of TGF-β1 such as NF-κB [26] or COX-2 pathway [37], thus further studies should be done to investigate the detail mechanisms.
In summary, we demonstrate that ASA could promote the immunomodulatory properties of BMMSCs, including the upregulation of Tregs and the downregulation of Th17 cells via 15d-PGJ2/PPARγ/TGF-β1 pathway. Infusion of ASA-pretreated BMMSCs might be a promising therapeutic strategy for treating immune-related disorders.
Footnotes
Acknowledgments
This work was supported by a grant from Beijing Municipal Committee for Science and Technology (Z121100005212004; to S.W.), the National Basic Research Program of China (2010CB944801; to S.W.), the Funding Project for Academic Human Resources Development in Institutions of Higher Learning under the jurisdiction of Beijing Municipality (PHR20090510; to S.W.), the Funding Project to Science Facility in Institutions of Higher Learning under the jurisdiction of Beijing Municipality (PXM 2009-014226-074691; to S.W.), the National Natural Science Foundation of China (81222011 to Y.L.), Science and Technology Activities of Beijing Overseas Students Preferred Foundation (to Y.L.), Beijing Key Laboratory Foundation of Science and Technology Special Work (Z121107002812034 to Z.S.), and the National Natural Science Foundation of China (81300892 to J.X.).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.