
Abstract
Mutation of Neuregulin-3 (Nrg3) results in defective embryonic mammary gland development. Here, we investigate functions of Nrg3 signaling in embryonic mammary morphogenesis. Nrg3 regulates the distribution of epithelial progenitor cells within the presumptive mammary-forming region during early mammary morphogenesis. Basal and suprabasal epithelial cells are significantly smaller within the hypoplastic mammary primordium (MP) that forms in Nrg3 mutants, indicative of failure to acquire mammary epithelial cell (MEC) morphological phenotype. Activation of Erbb4 JM-a CYT-1, an Erbb4 isoform expressed in the developing MP, leads to MEC spreading and migration. Nrg3 promotes the accumulation of epithelial progenitor cells at the MP site in embryo explant cultures. Our results implicate Nrg3 signaling in mediating key events of mammary mesenchyme specification, including mesenchymal condensation, mitosis, and induction of mammary marker expression. Taken together, our results show Nrg3 has a major role in conferring specification of the mammary phenotype to both epithelial and mesenchymal progenitor cells.
Introduction
T
The initial stages of mammary primordium 3 (MP3) formation fail to occur in Nrg3ska , hypomorphic mutants that express reduced Nrg3 levels. Our previous studies of Nrg3ska mutants identified Nrg3 as a key mediator of mammary primordial specification, linking Nrg3 to the establishment of the mammary lineage during embryogenesis [6,7]. Nrg3 is expressed in the dermal mesenchyme underling the site where MP will subsequently form between E10.5 and E11.25 [8]. When MP3 becomes morphologically distinct at ∼E11.5, Nrg3 expression is found within the epithelium and is no longer expressed in the underlying mesenchyme [8]. Expression of Nrg3 throughout the epidermal progenitor cell compartment in transgenic K14-Nrg3 mice led to formation of supernumerary mammary gland formation, confirming a role for Nrg3 as a promoter of mammary cell fate during embryogenesis [7,9]. Dramatic alterations were also observed in postnatal K14-Nrg3 epidermis skin including changes in expression pattern of Tenascin C, an extracellular matrix protein of stem cell niches [9]. The receptor for Tenascin C, integrin β1, and integrin α6, a marker for epithelial progenitor cells, also showed perturbed expression in K14-Nrg3 epidermis, further suggesting a link between Nrg3 signaling and progenitor cell regulation [9]. The cognate receptor of Nrg3, Erbb4 is expressed in dermal mesenchymal tissues of the mammary-forming region between E10.5 and E11.0, before becoming strongly expressed in the surface epithelium of the site where the MP forms and remains expressed in the epithelium once MP3 has formed at E11.5 [8]. Erbb4 is alternatively spliced and exists as four major isoforms: JM-a CYT-1, JM-a CYT-2, JM-b CYT-1, and JM-b CYT-2 [10]. The Erbb4 isoforms regulate distinct aspects of postnatal mammary epithelial cell (MEC) and breast cancer behavior including mammary cell differentiation, migration, proliferation, and apoptosis [11 –13]. In fact, chemoattractant properties of Nrg3 and another Neuregulin member, Neuregulin-1 (Nrg1) have been demonstrated: both chemoattract interneuronal cells, a type of neural progenitor cell that expresses Erbb4 in the developing cerebral cortex/forebrain [14]. Ligands for the Erbb receptor tyrosine kinases, the Neuregulins, are compelling candidates to mediate early mammary morphogenesis due to their expression patterns in relevant MP tissues [8].
Until recently, a lack of suitable reporter models existed for monitoring the early events occurring during MP formation. As a result, much remains to be understood regarding the cellular and molecular mechanisms by which the initial MP is developed from the simple epithelium. It has been clearly established that a very low level of cell proliferation is detected within the mammary primordial epithelium during the initial stages of organ formation from E11 through E14 stages [15,16]. Previous studies have concluded that early mammary morphogenesis is primarily mediated via localized cell movements [15,17]. It is not yet clear which signals mobilize the epithelial progenitors that are thought to locally migrate from along and near the mammary line to form the MP [1]. Cell hypertrophy is also thought to contribute to increase in MP size [16].
MP formation and initial organ growth appear to be primarily elicited through epithelial cell migration. We hypothesized that Nrg3 might facilitate epithelial progenitor movements in the presumptive mammary-forming region as the MP forms. We aimed to study the effects of Nrg3 and Erbb4 signaling on morphogenesis of embryonic mammary epithelium and mesenchyme, and cell migration and spreading of MECs. Here, we show that Nrg3 has a major role in progenitor cell positioning and conferring specification of the mammary phenotype to both epithelial and mesenchymal progenitor cells.
Materials and Methods
Sample collection
All animal work was carried out under UK Home Office project and personal licenses following local ethical approval and in accordance with local and national guidelines. TOPGAL mice (stock No. 4623) were purchased from the Jackson Laboratories (Bar Harbor, ME). s-SHIP-GFP mice were kindly provided by the late Professor Larry Rohrschneider (Fred Hutchinson Cancer Research Center, Seattle, WA).
Expression analyses
Immunohistochemistry and whole-mount immunofluorescence (WM-IF) of MP were performed as described [9,18]. Antibodies used for immunostaining were as follows: green fluorescent protein (GFP) (clone FM264G) from BioLegend UK (London, United Kingdom); phospho-Ser10-Histone H3 (clone 3H10) from Millipore (Hertfordshire, United Kingdom); ERα from Santa Cruz Biotechnology (Dallas, TX); Lef1 (clone C12A5), and Cleaved Caspase-3 (Asp175) from Cell Signaling Technologies (Danvers, MA). Antibody-labeled tissues were three-dimensionally analyzed using Leica TCS SP II confocal scanning microscope as described [18]. Mitotic cells within control and Nrg3ska MP3 were scored by manually counting phospho-histone-positive cells through confocal images of MP3 made every 1.5 μm. X-gal staining was performed as described [6]. Quantitative real-time RT-PCR (qRT-PCR) was performed on freshly extracted total RNA from total MP at E12.5, the microdissected components of the MP [mammary epithelium (ME) and mammary mesenchyme (MM)] and separated epidermis and mesenchyme. qRT-PCR was performed using the following TaqMan Gene Expression Assay probes from Life Technologies (Paisley, United Kingdom): Actb (Mm00607939_s1), Esr1 (Mm00433149_m1), Ar (Mm00442688_m1), Pth1r (Mm00441046_m1), and Wnt10b (Mm00442104_m1). Results were analyzed with the comparative CT (ΔΔCT) method normalized to Actb value and compared to comparator in triplicate. Real-time PCR (RT-PCR) detecting Erbb4 isoforms Erbb4 JM-a/b and CYT-1/2 were performed as described [9].
Morphological analysis of MP
GFP pictures of s-SHIP-GFP-expressing embryos at E11.5 and E12.5 were obtained on EVOS FL Color Imaging System from Life Technologies using either ×2 or ×4 objectives. GFP pictures were represented as black-and-white images to highlight where ME progenitor cells expressing s-SHIP-GFP accumulated on the embryo's surface. Briefly, brightness/contrast and high dynamic range (HDR) toning in the raw images were adjusted using Adobe Photoshop CS5, and threshold was adjusted to display GFP+ region of interest as black dots on a white background. Arrangement of MP along the ML was further assessed by measuring the angle of the line connecting MP1 to MP3 and MP3 to MP5 using ImageJ. To examine mammary cell morphology, cell borders were visualized using CellMask Orange Plasma Membrane Stain from Life Technologies. Briefly, WM-IF was performed on microdissected MP3 with a modified permeabilization step using 1 mg/mL digitonin in phosphate-buffered saline (PBS) since detergents and methanol in general method are not compatible for the dye. The stained tissue was photographed three-dimensionally every 1.5 μm slice, using a confocal microscope equipped with ×40 objective. To measure three-dimensional sphericity and volume of basal and suprabasal MEC, individual cell periphery was manually traced using plasma membrane stained images through each slice using the Contour Surface tool in Imaris software from Bitplane (Zurich, Switzerland).
Recombinant proteins
Mutagenized Nrg3, referred to as Nrg3mut is described in Rakic et al. [14]. Recombinant protein rNrg3-EGF wt and rNrg3-EGF mut were prepared in S2 Schneider cells as previously described [6]. Nrg1β-EGF (396-HB-050) and Wnt3a (1324-WN-010) were purchased from R&D Systems (Minneapolis, MN).
Ex vivo culture of embryonic flanks
Embryonic flanks from forelimb to hindlimb were carefully bisected from freshly isolated E11.0 Nrg3ska s-SHIP-GFP embryos in D-PBS (Life Technologies) taking care not to damage the epidermis around the presumptive mammary region. By monitoring GFP under fluorescent stereomicroscope, only intact Nrg3ska flanks at a stage in which the MP had not formed yet were used for experiments. Flanks were placed in humidified center-well organ culture dishes, Becton Dickinson (Oxford, United Kingdom) equipped with polycarbonate filter membrane (P9699-100EA; Sigma, St. Louis, MO) and culture medium at 37°C with 5% CO2. Freshly reconstituted EmbryoMax KSOM Embryo Culture media, Merck Millipore (Billerica, MA) containing 75 μg/mL ascorbic acid (Sigma) was used. Two μg/mL rNrg3-EGF or PBS was added to the media. After 24 h incubation, the explants were evaluated for MP3 formation by GFP appearance and were fixed in 4% paraformaldehyde (PFA) in PBS and then subject to phospho-histone staining, which was used to score the number of mitotic cells in the mammary epithelial and mesenchymal tissues.
Cell culture and transfection
HC11 cells were maintained with RPMI1640 (Life Technologies) supplemented with 10% serum, 1% penicillin/streptomycin, 5 μg/mL insulin (19278; Sigma), and 10 ng/mL human recombinant EGF (236-EG-200; R&D Systems). To develop stable cell lines expressing isoform-specific Erbb4, 10 μg of human Erbb4 JM-a CYT-1 and JM-a CYT-2 cDNAs from expression constructs cloned in pcDNA3.1 were introduced into HC11 cells using Amaxa Nucleofector II device and Cell Line Nucleofector kit V with a program T-024 (Lonza, Basel, Switzerland). After nucleofection, cells were maintained with media containing 1 mg/mL G418 (Life Technologies). After 2 weeks, a number of surviving colonies were tested for Erbb4 overexpression. Two independent clones were used for each isoform in this study: JM-a CYT-1 isoform, clone 5 and 6; JM-a CYT-2 isoform, clone 3 and 43.
In vitro chemotactic assay
Polycarbonate membrane transwell inserts (8 μm pore size, 24-well format) were coated with 50 μg/mL rat tail Collagen type I (Becton Dickinson). HC11 cells expressing Erbb4 JM-a CYT-1 and JM-a CYT-2 were treated with serum-free medium for 72 h prior to the transwell migration assay. The serum-starved cells were harvested using HyClone HyQtase cell detachment reagent from Thermo Scientific (Waltham, MA) and resuspended to 4×105 cells/mL with RPMI1640 supplemented with 1% serum, 1% bovine serum albumin, and 20 mM HEPES. Two hundred microliters of cell suspension was added onto the transwell inserts. The inserts were then transferred to 24-well plates filled with medium containing PBS, 2.0 μg/mL rNrg3-EGF wt, 1.0 μg/mL Nrg1β-EGF, or 0.01 μg/mL EGF. After 6 h incubation, cells on the membrane insert were fixed with 4% PFA in PBS followed by 4′,6-diamidino-2-phenylindole (DAPI) staining. The transwell migration was scored by counting the number of DAPI-labeled cells on the membrane at the bottom side.
Cell adhesion staining and spreading assays
0.5×105 HC11 cells expressing Erbb4 JM-a CYT-1 or JM-a CYT-2 per well were placed on fibronectin-coated 12-well plates in culture medium overnight. Cells were incubated with serum-free medium prior to stimulation with growth factor. After 72 h starvation, cells were treated for 15 min with serum-free medium containing PBS, 0.5 μg/mL rNrg3-EGF wt or mut, 0.2 μg/mL Nrg1β-EGF, and 0.05 μg/mL Wnt3a. Cells were immediately fixed with 4% PFA in PBS followed by staining for phospho-Tyr397-focal adhesion kinase (p-FAK) from Cell Signaling Technologies, Alexa555-Phalloidin and DAPI from Life Technologies. p-FAK-labeled cell adhesions were photographed using EVOS FL microscope equipped with ×40 objective. Actin-based cell cytoskeleton and DAPI-labeled nuclei were photographed using ×10 objective. Segmented shape of cell and nucleus were abstracted and automatically measured using CellProfiler cell image analysis software (
Statistical analysis
The data in the graphs are presented as mean and the standard error of the mean. The data were analyzed by two-tailed ANOVA or Student's t-test using GraphPad Prism 6 software. P value<0.0001 is considered as extremely significant (****), P value=0.0001–0.001 as highly significant (***), P value=0.001–0.01 as significant (**), and ≥0.05 as not significant (ns), respectively.
Results
Nrg3ska mutants display aberrant distribution of epithelial progenitor cells along the presumptive mammary-forming region during early mammary gland morphogenesis
We previously found that 55% of Nrg3ska hypomorphic mutants fail to form mammary gland 3 when mice were assessed for the presence or absence of nipples 7–10 days after birth [19]. We backcrossed Nrg3ska hypomorphic mutants and controls (C57Bl6/J) onto the s-SHIP-GFP reporter background, which expresses GFP in a subset of epithelial-lineage progenitor cells during embryonic development, including those present in the mammary-forming region [20]. We were then able to easily examine the formation of the MP between E10.5 and E12.5 since small aggregates of s-SHIP-GFP + cells are readily apparent at, and in the vicinity of, the sites where the MP form in controls (Fig. 1a, b). s-SHIP + epithelial progenitor cells appear less focally distributed along the mammary-forming region of Nrg3ska hypomorphic mutants; more GFP+ cells remain on the dorsal-most aspect of the ventral flank when compared with control embryos in which a much denser ventral accumulation of GFP+ cells is observed at E11.5 (Fig. 1a). GFP+ cells observed along the mammary line fragment that corresponds to the site of MP3 formation appear to be shifted dorsally in Nrg3ska compared with controls (Fig. 1a, b). These observations are further supported when changes in MP3 location were assessed by measuring the angles of the lines that connect MP1 to MP3, and MP3 to MP5 and compared them to those from control embryos and found this is significantly reduced in Nrg3ska embryos (Fig. 1c).
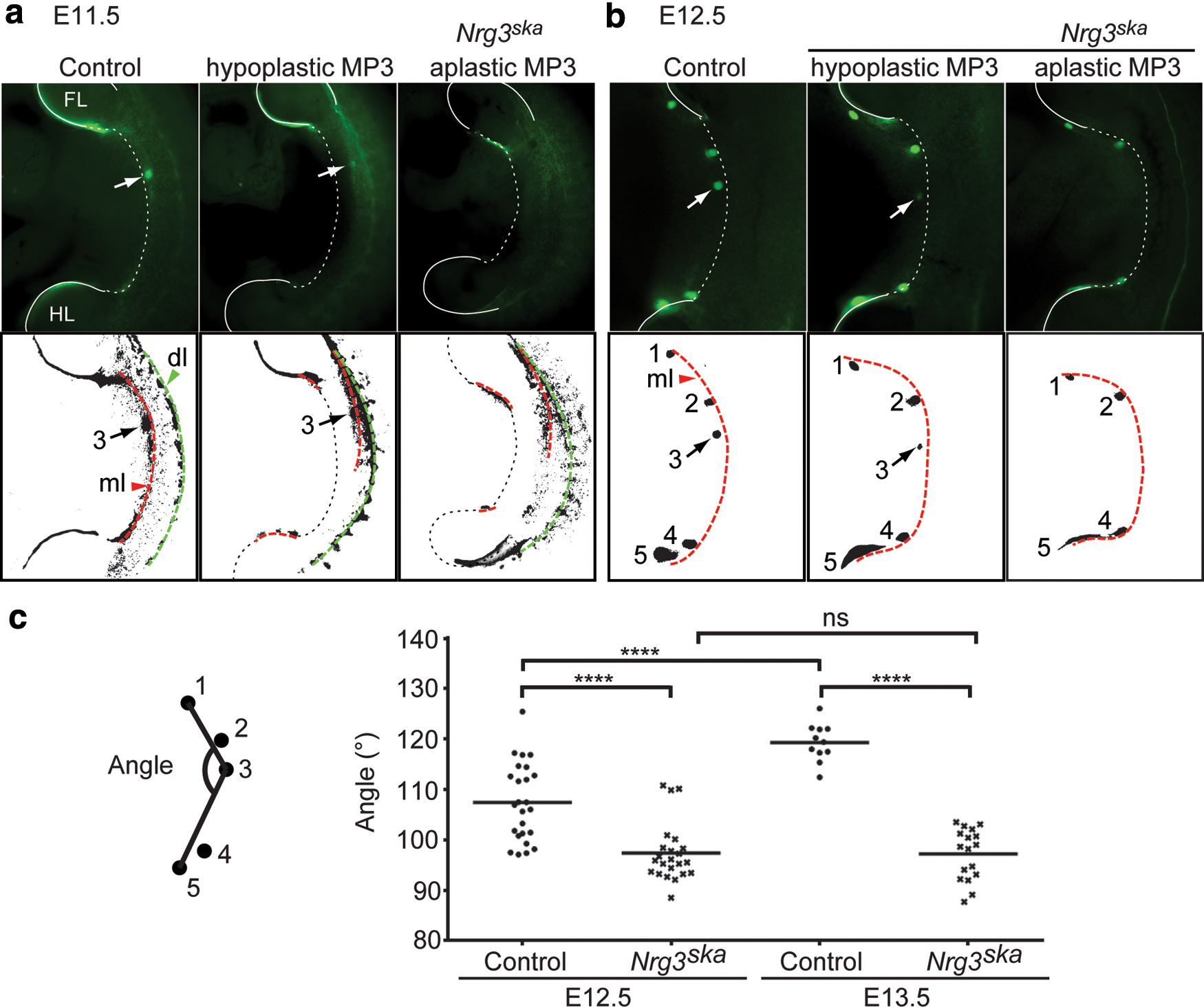
Nrg3ska
mutants display aberrant distribution of mammary epithelial progenitor cells during early mammary primordial morphogenesis. s-SHIP-GFP-expressing cells are shown visualized in whole-embryos. GFP+ cells relevant to embryonic mammary gland development are indicated in black in the digitized images shown below the GFP images. The mammary line fragments and dorsal line are also visible with s-SHIP-GFP
+ cells and indicated by red and green dotted lines, respectively.
Fewer s-SHIP-GFP
+ cells are observed within the nascent Nrg3ska
MP3 when compared with control embryos at E12.5 (Fig. 1b). We restricted phenotypic analysis to MP3 since MP3 exhibits the most striking morphological defects in Nrg3ska
embryos. We found when E12.5-stage embryos are assessed for s-SHIP-GFP
+ cell accumulation at the presumptive site of MP3, 9% of Nrg3ska
mutants fail to show any s-SHIP-GFP
+ cell accumulation and were scored as aplastic; 37% form hypoplastic MP3 with substantial reduction in accumulation of s-SHIP-GFP
+ cells and are greatly reduced in size, and the remaining 57% form a relatively-normal sized organ and were scored as MP3+ (Supplementary Table S1; Supplementary Data are available online at
The s-SHIP-GFP reporter shows a similar expression pattern when compared to the TOPGAL-F reporter mouse [21] in both control and Nrg3ska mutants between E10.5 and E12.5 (Supplementary Fig. S1a). In Nrg3ska mutants, fewer s-SHIP-GFP + and TOPGAL+ cells accumulate along the mammary line fragment that spans the site of mammary MP3 formation. When profiled by qRT-PCR, Wnt10b, a mammary line marker [22], is reduced in the epithelium isolated from hypoplastic Nrg3ska MP3 compared with control MP3 (Supplementary Fig. S1b). Wnt10b expression is higher in the surface epithelium of the epidermis from the region dorsal to MP3 of Nrg3ska embryos compared with control embryos, which is consistent with Wnt10b-positive epithelial progenitor cells remaining dispersed in the mammary-forming region of Nrg3ska embryos near MP3. These results suggest Nrg3 regulates s-SHIP + and TOPGAL+ progenitor cells recruitment to form MP3.
Maturation to achieve MEC phenotype fails in hypoplastic MP3 that form in Nrg3ska mutants
We used plasma membrane staining to assess epithelial cell morphology (Fig. 2a). Nrg3ska hypoplastic MP3 populations are composed of smaller epithelial cells when compared with control cells, which occupy larger volumes, but no significant change in cell sphericity is detected (Fig. 2b). Basally located cells show striking reductions in volume and suprabasal cells also show a significant reduction in cell volume in Nrg3ska MP3 when compared with controls. Changes in sphericity are detected between basal and suprabasal cells, suggesting that morphological differentiation of the MECs is initiated, but does not mature to a full mammary phenotype, in Nrg3ska MP3. We observed no changes in the number of apoptotic cells in hypoplastic Nrg3ska and control MP3 epithelium when assessed by cleaved caspase-3 labeling (Supplementary Fig. S2). Nuclear staining shows that epithelial multilayering is observed in MP3 of Nrg3ska hypomorphic mutants (Fig. 2). We found no significant change in the number of mitotic cells observed in hypoplastic Nrg3ska and control MP3 epithelium as assessed by phospho-histone H3 staining (Fig. 3). It is notable that the surface epithelium has similar mitotic rate in Nrg3ska hypomorphic mutants and control embryos (data not shown). Results from these morphometric analyses are consistent with our confocal and histological observations of the morphology of the cells present at the site of hypoplastic Nrg3ska MP3 (Fig. 2 and Supplementary Figs S1 and S2) and indicate Nrg3 mediates transitioning to the mammary epithelial phenotype.
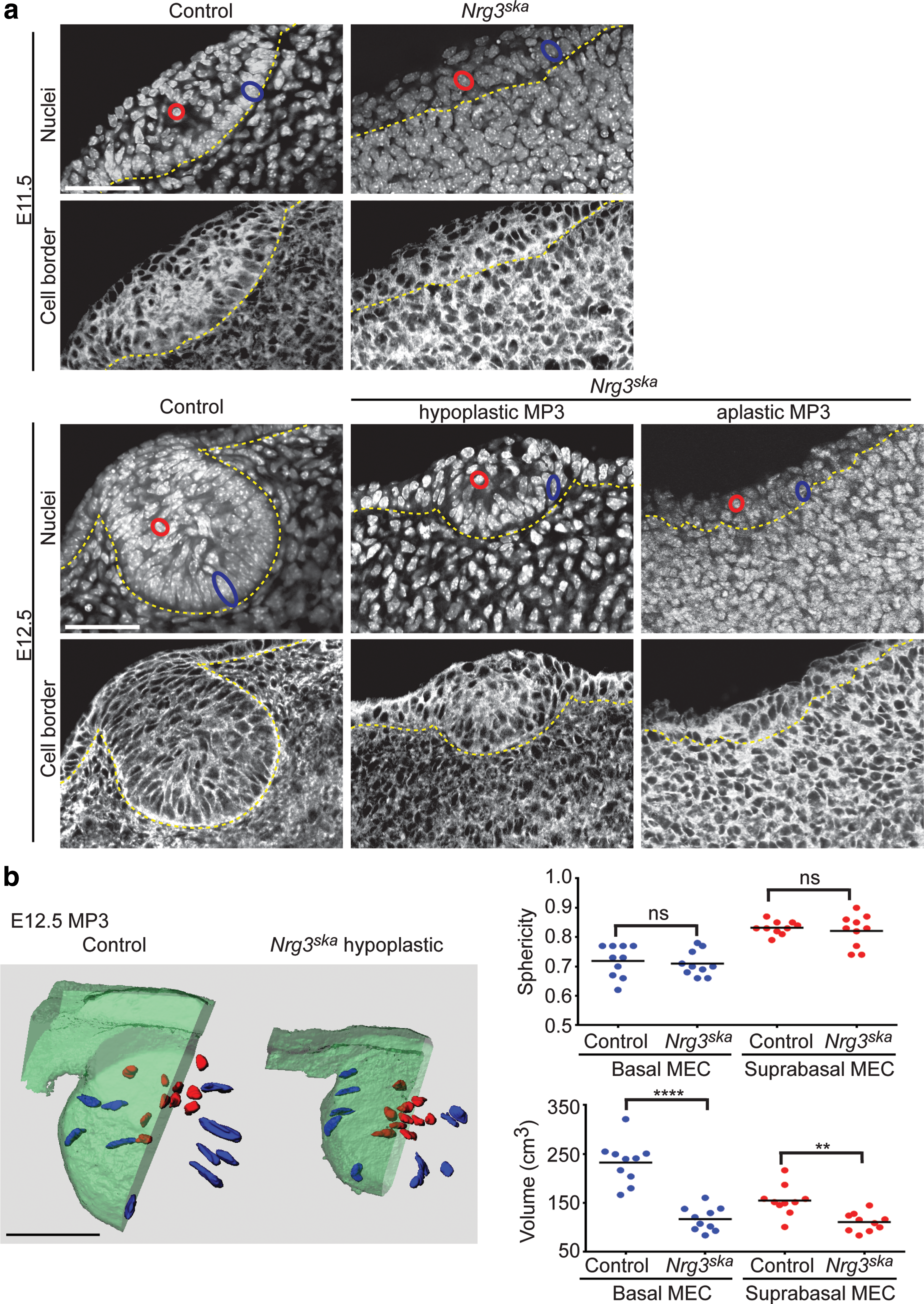
Morphological maturation and transition of surface epithelium to a mammary epithelial cell phenotype does not occur in MP3 of Nrg3ska
mutants. (
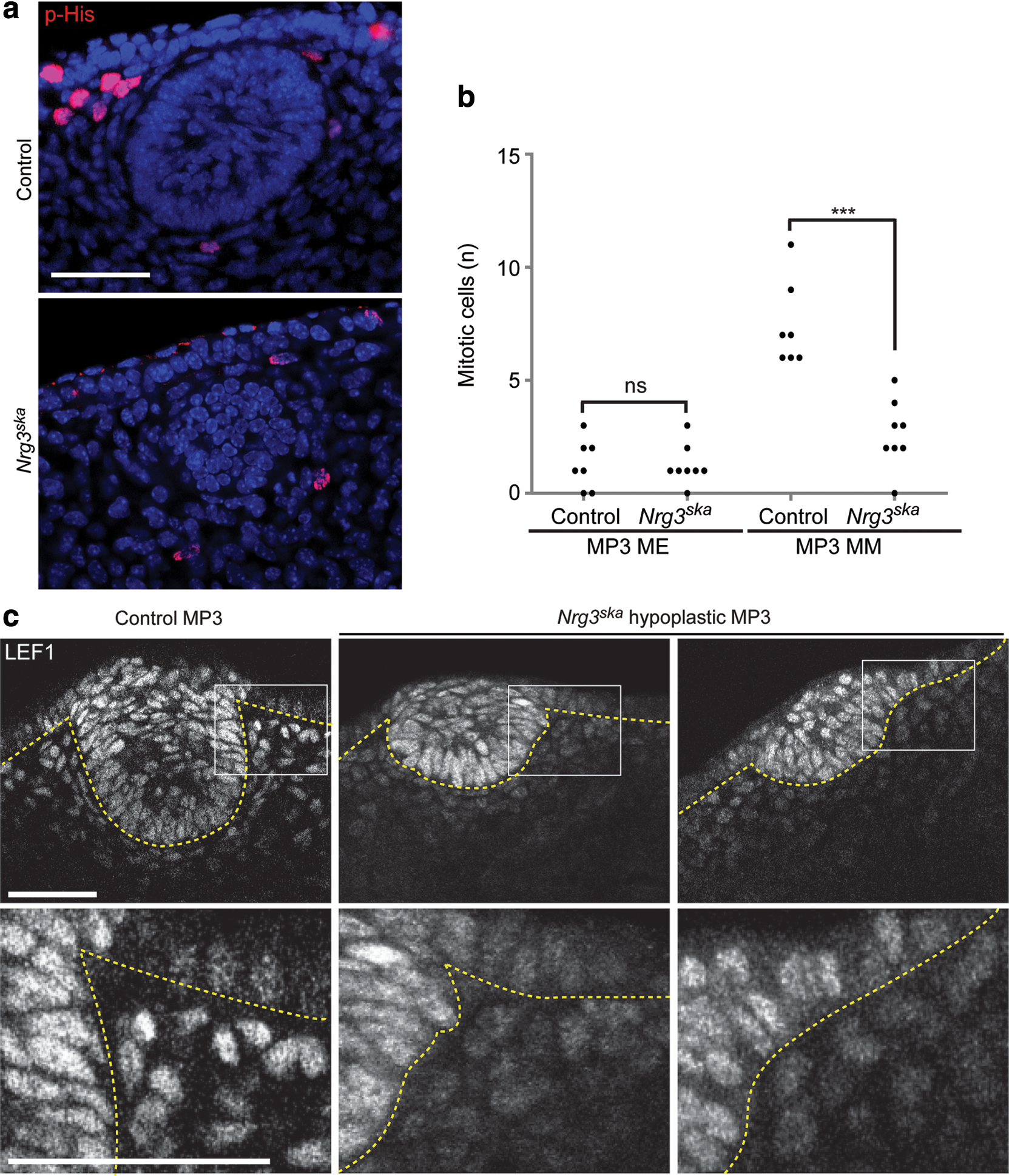
Nrg3ska
mutants display impaired mammary mesenchymal proliferation. (
Nrg3ska mutant MP3 display defective specification of mammary mesenchymal cells
Morphological analyses indicated that defects are also present in mesenchymal cells associated with Nrg3ska MP3. The concentric rings of mesenchymal cells aligned around control MP3 epithelial cells start to appear at E12.5 but are not observed in mesenchymal cells adjacent to Nrg3ska MP3 and these cells do not appear to condense (Figs. 2 –4). Lef1 stains both epithelial and mesenchymal MP3 control cells at E12.5 (Fig. 3c). We found that although Nrg3ska MP3 are comprised of fewer epithelial cells compared with control MP3, Lef1 was expressed in Nrg3ska MP3 epithelial cells. Lef1 expression was observed in mesenchymal cells adjacent to control MP3, but not in the mesenchymal cells associated with Nrg3ska MP3. We also stained MP3 for expression of other mammary mesenchymal markers, including ERα (Fig. 4a). We detected a reduced number of ERα+ mesenchymal cells surrounding Nrg3ska MP3 and lack of their condensation, even in those that are a size similar to control MP3 (denoted MP3+); both are indicative of failed specification of mammary mesenchymal cells (Fig. 4a). Fewer mitotic cells are detected in Nrg3ska mesenchymal cells associated with MP3 at E12.5 when compared with control (Fig. 3a, b). To ascertain whether Nrg3 is required for the Pthrp/Pth1r-mediated specification of mesenchymal cells to a mammary phenotype to occur, we assayed levels of Pthrp and Pth1r, and mammary mesenchymal markers Esr1 and Ar, in RNA isolated from E12.5 mammary primordial tissues. Using qRT-PCR, we found similar levels of Pthrp in epithelial cells from Nrg3ska and control MP3 (data not shown). Mesenchymal cells surrounding the epithelium from aplastic and hypoplastic Nrg3ska MP3, expressed the transcripts of Esr1, Ar, and Pth1r at significantly reduced levels compared to those from control MP3 and at levels more similar to those found in dermal mesenchymal cells and indicate failed mammary mesenchymal specification (Fig. 4b).
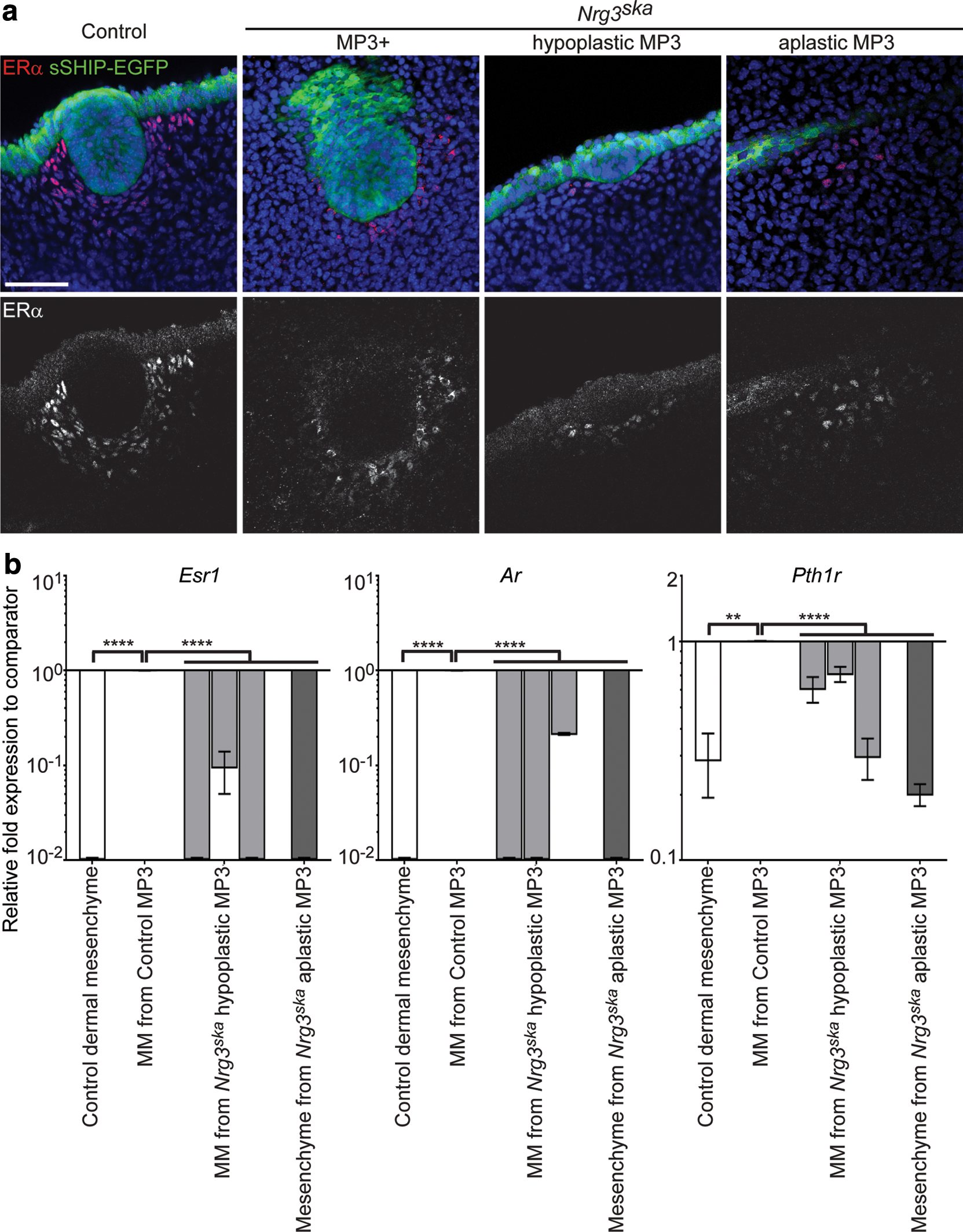
Nrg3ska
mutants display defective specification of mammary mesenchyme.
Rescue of Nrg3ska MP3 cultured with Nrg3 is not achieved through increasing epithelial mitotic activity
When using conventional protocols that include serum-containing media for ex vivo culture of embryonic flanks, we observed a significant amount of epithelial proliferation within MP forming in culture (data not shown). We therefore optimized an ex vivo culture system using serum-free media so that it would mimic in vivo development where very little epithelial proliferation contributes to early stages of MP development (Fig. 5a). Nrg3ska;s-SHIP-GFP ventral flanks were explanted at E11.0, prior to MP3 formation. Following 24 h in culture ex vivo using these serum-free conditions, we assessed each Nrg3ska;s-SHIP-GFP ventral flank for the accumulation of s-SHIP-GFP + cells at the site of MP3. We found that, 50% of the Nrg3ska;s-SHIP-GFP flanks formed MP3, while the remaining 50% were scored as having aplastic MP3. We found that when rNrg3-EGF was added to the culture media, after 24 h in culture ex vivo, 87% of the explanted Nrg3ska;s-SHIP-GFP flanks formed MP3 and only 13% were scored as having aplastic MP3. No difference in the number of mitotic cells was observed in Nrg3ska;s-SHIP-GFP MP3 epithelial cells whether culture media contained rNrg3-EGF or not (Fig. 5b). In contrast, a significant increase in the number of mitotic cells was observed in the mesenchymal cells adjacent to MP3 epithelium when explants were cultured in media containing rNrg3-EGF compared qith control media (Fig. 5b).

Formation of MP3 in Nrg3ska
mutants can be restored by rNrg3-EGF treatment ex vivo without increasing epithelial mitosis and is associated with stimulation of mammary mesenchymal mitosis.
MP selectively express cleavable Erbb4 JM-a Isoforms
To further investigate the role of Nrg3 in facilitating the accumulation of epithelial progenitor cells at the site of MP3, we profiled the Erbb4 isoforms expressed during early embryonic mammary gland development by RT-PCR. We found that only the cleavable JM-a isoform is expressed and no JM-b isoform is detected in mid-gestation MP (Fig. 6a). Both CYT-1 and CYT-2 isoforms are expressed (Fig. 6a).
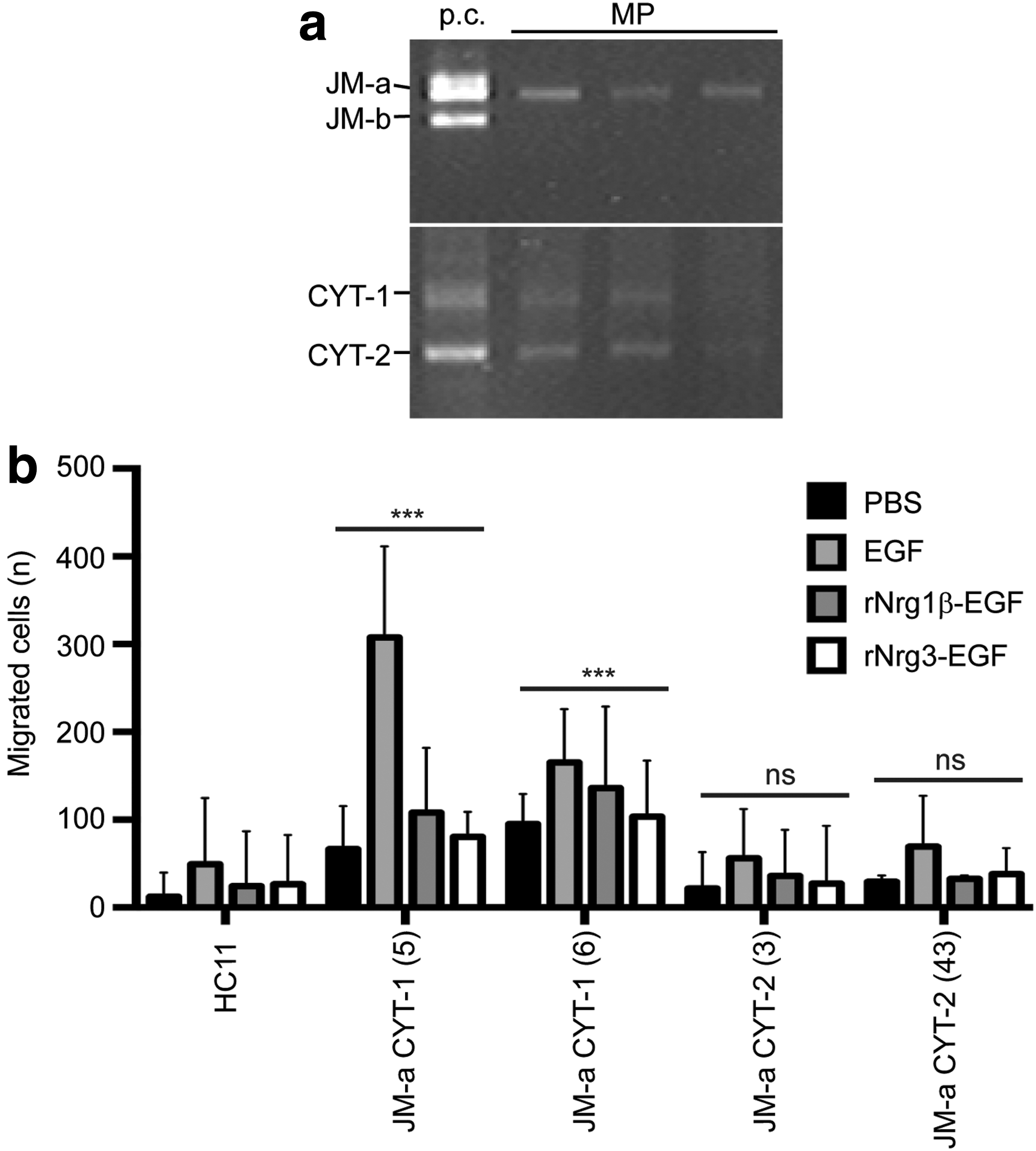
Expression of Erbb4 JM-a CYT-1 isoform enhances migration of HC11 cells.
MECs expressing Erbb4 JM-a CYT-1 isoform are more motile than cells expressing JM-a CYT-2 isoform
We transfected normal mouse mammary epithelial (HC11) cells with either the JM-a CYT-1 or JM-a CYT-2 isoform of ERBB4 to create stable cell lines expressing the isoforms relevant to embryonic mammary development. Using transwell assays, we found that cells expressing JM-a CYT-1 were significantly more migratory than HC11 cells expressing JM-a CYT-2 isoform, which exhibited minimal migratory ability that is comparable to levels detected with untransfected HC11 cells (Fig. 6b). Unstimulated HC11 cells expressing JM-a CYT-1 express activated full-length (180 kD) and cleaved (80 kD) forms of Erbb4 and activated Erbb2 (Supplementary Fig. S3); these cells were slightly more migratory when treated with either rNrg1β-EGF or rNrg3-EGF, ligands for Erbb4 (Fig. 6b). The effects of stimulating cells with rNrg3-EGF wt and rNrg3-EGF mut were evaluated by the levels of Erbb4 and Erbb2 phosphorylation as shown in Supplementary Figure S4. No increase in canonical Wnt signals was observed when HC11 cells expressing either JM-a CYT-1 or JM-a CYT-2 isoform were stimulated with rNrg3-EGF in TOPflash assays (Supplementary Fig. S5). These results indicate that Nrg3 does not increase canonical Wnt signals in MECs expressing either JM-a CYT-1 or JM-a CYT-2 isoform of Erbb4.
Nrg3 promotes spreading of MECs expressing Erbb4 JM-a CYT-1 isoform
To investigate the ability of Nrg3 to modulate MEC morphology, HC11 cells expressing either the JM-a CYT-1 or JM-a CYT-2 Erbb4 isoform were stimulated with rNrg3-EGF. Mammary cells expressing JM-a CYT-2 isoform showed no cell spreading in response to Nrg3 stimulation (data not shown). Mammary cells expressing JM-a CYT-1 isoform responded to both rNrg3-EGF and rNrg1β-EGF stimulation with a modest and significant amount of cell spreading (Fig. 7a, b). Cells expressing JM-a CYT-1 isoform treated with Wnt3A showed no cell spreading response, suggesting that growth factor-mediated cell spreading is specific for Erbb4-ligands in this cell type.

Nrg3 promotes cell spreading and focal adhesion formation of HC11 cells expressing Erbb4 JM-a CYT-1 isoform.
MECs expressing Erbb4 JM-a CYT-1 isoform form focal adhesions after stimulation with rNrg3-EGF
Under nonstimulating conditions with rNrg3-EGF mut, JM-a CYT-1 expressing HC11 cells have sparse distribution of p-FAK rather than the large focal adhesions at actin bundles that are observed in HC11 cells and HC11 cells expressing JM-a CYT-2 (Fig. 7c). HC11 cells and HC11 cells expressing JM-a CYT-2 show no obvious response to stimulation with rNrg3-EGF wt, whereas large focal adhesions are formed in JM-a CYT-1 expressing HC11 cells after stimulation with rNrg3-EGF wt (Fig. 7c). HC11 cells expressing JM-a CYT-1 isoform show a discernible change in cell morphology after stimulation with rNrg3-EGF (Fig. 7c).
Discussion
Alterations in epithelial progenitor cell distributions in Nrg3 hypomorphic mutants suggest that Nrg3 signals influence the behavior of epithelial progenitor cell populations relevant to embryonic mammary development from E10.5 through E12.5. The failure of a signal that normally leads to s-SHIP-GFP progenitor cell aggregation along the mammary-forming region would explain the striking phenotypes observed uniquely with MP3 in Nrg3ska mice [1]. The results presented here demonstrate the power of using the s-SHIP-GFP reporter model that permits analyses of epithelial progenitor cells through sequential developmental stages in situ using ex vivo embryonic cultures, which had not been previously possible for studies of embryonic mammary morphogenesis. Results from our embryo explant cultures in the s-SHIP-GFP reporter background strongly suggest Nrg3 promotes epithelial progenitor aggregation as MP3 initially forms. We also demonstrate that Nrg3 regulates epithelial progenitor accretion without increasing epithelial mitosis. The limited epithelial proliferation detected cannot account for the observed increases in mammary primordial size that occur between E10.5 and E13.5 and influx of cells from the adjacent surface epithelium accounts for most increases in epithelial cell numbers within the nascent organ [15 –17]. Our results indicate Nrg3 signals mediate the distribution of epithelial progenitor cells along the ventral thoracic flank at the time that they are thought to be locally migrating to form the MP. A number of transcription factors including Gata3 and Tbx3 are required for the formation of all five pairs of MP, while others, such as Gli3 and Lef1, are only required for formation of certain MP, including MP3 [23 –27]. It is not clear how the factors that regulate development of MP3 control epithelial progenitor cell distributions in the mammary-forming region, how these factors interact or whether any directly induce Nrg3 expression.
Using 3D morphometric analyses of MP3, we have quantified hypertophy that occurs in the epithelial progenitor cells within MP3 during maturation of the embryonic mammary epithelium. Our results indicate that Nrg3ska MP3 cells achieve transition from cuboidal to columnar morphology, which normally occurs as the epithelium stratifies and during embryonic mammary development [25]. Further enlargement in size to acquire a mammary cell phenotype fails in Nrg3ska MP3 cells. Defective epithelial hypertrophy is observed in Nrg3ska MP3 and indicates Nrg3 mediates transitioning to the mammary epithelial phenotype. Insufficient Nrg3 levels leads to formation of abnormal MP3 comprised of small aggregates of epithelial progenitor cells that have not fully differentiated to the mammary phenotype. Cell-based studies indicate Nrg3 promotes cell spreading and an ability to modulate cell morphology. Our results are consistent with a potential role for Nrg3 in promoting cell hypertrophy and contributing to initial organ growth via increase in cell area through cell spreading. However, it is not yet clear how Nrg3 elicits the columnar hypertrophy observed in vivo.
Subsequent mammary mesenchymal specification is incomplete in Nrg3ska MP3 as assessed by reduced mammary mesenchymal marker expression. This finding indicates that Nrg3-mediated mammary specification of epithelial progenitors cells also activates downstream effectors required for the transmission of signals from the mammary epithelial to adjacent mesenchymal cells. We also show that Nrg3 signaling increases proliferative potential of mammary mesenchymal precursor cells. Nrg3-mediated specification of the mammary epithelium is required for proliferation of the presumptive mammary mesenchymal precursor cells and their specification to a mammary mesenchymal phenotype to occur.
Threshold levels of Nrg3 are likely to be required to form MP3 with fully specified mammary epithelial tissue capable of inducing Pth1r expression in the adjacent mesenchyme by some unknown mechanism. Sufficient Pth1r levels are required so stimulation of Pth1r by Pthrp leads to induction of mammary mesenchyme marker expression, including AR and ERα [28], which show reduced expression in Nrg3ska MP3. The Pthrp signaling axis is a master regulator of mammary mesenchymal specification that it mediates by activating canonical Wnt signaling in mesenchymal cells [28,29]. Similarities exist between Nrg3ska and Pthrp-deficient MP with respect to their reduction in mammary mesenchymal marker expression and reduced proliferation of mammary mesenchymal cells. The mammary mesenchymal condensation defect observed in Nrg3ska MP3 distinguishes its mesenchymal phenotype from that of Pthrp and Pth1r-deficient MP, which appear to undergo mesenchymal condensation [30]. We did not observe induction of Pth1r expression when mesenchymal tissues from Nrg3ska were cultured and treated with rNrg3 (data not shown) so Nrg3 does not appear to directly induce expression of Pth1r.
Cell-based assays show that Nrg3 does not increase canonical Wnt signals in MECs expressing either JM-a CYT-1 or JM-a CYT-2 Erbb4 isoforms. Changes in expression of TOPGAL+ cells observed within the mammary-forming region of Nrg3ska mice are unlikely to be due to Nrg3 inducing canonical Wnt expression, but are consistent with Nrg3 stimulating migration and local aggregation of TOPGAL+ precursor populations at the site of MP3 but this remains to be formally demonstrated.
MECs expressing the Erbb4 receptor of JM-a CYT-1 and JM-a CYT-2 show that these isoforms elicit distinct effects. CYT-1 and CYT-2, the cytoplasmic variants differ by the presence (CYT-1) or absence (CYT-2) of a short sequence containing a putative binding site for phosphatidylinositol (PI) 3-kinase p85 subunit [31]. In our studies, cells stably transfected with full-length receptor for the JM-a CYT-1 isoform were significantly more migratory than those expressing JM-a CYT-2 isoform. Directional cell migration is regulated by assembly and turnover of integrin-mediated adhesions [32]. Mammalian cells attach to surfaces through focal adhesions, cellular structures characterized by complexes of the transmembrane protein integrin, and intracellular proteins including paxillin and focal adhesion kinase [33]. Since Erbb4 JM-a CYT-1 expression enhances migration ability in HC11 cells, we examined the effect of JM-a CYT-1 expression on cell adhesion formation, which was assessed with immunostaining with p-FAK, a regulator of focal adhesion turnover. Under nonstimulating conditions, JM-a CYT-1 expressing HC11 cells display sparse distribution of p-FAK, rather than the large focal adhesions at actin bundles observed in HC11 cells and HC11 cells expressing JM-a CYT-2 (Fig. 7a). These data are in agreement with the observation that JM-a CYT-1-expressing cells are motile since migrating cells develop small adhesions rather than stabilized focal adhesions. MECs expressing JM-a CYT-1 isoform exhibit distinct responses to Nrg3 stimulation by forming focal adhesions and spreading, when compared to MECs expressing JM-a CYT-2 isoform, which show negligible changes in focal contacts and cell morphology after Nrg3 stimulation. These results suggest that Nrg3 stimulation affects cell adhesion dynamics and morphology, specifically for MEC cells expressing the JM-a CYT-1 isoform.
Divergent cellular responses by distinct Erbb4 isoforms have been previously reported in MECs. A study using inducible mouse models to express CYT-1 and CYT-2 isoforms in postnatal MECs showed that the intercellular domain (ICD) of these two isoforms exert opposing effects on the postnatal mammary epithelium when transgenically expressed in vivo [12]. MECs from transgenic mice induced to express CYT-1 isoform were found to have decreased cell growth, increased lumen formation in 3D culture, increased Stat5a activation, and precocious lactogenic differentiation when compared with noninduced transgenic control postnatal mammary epithelium. Conversely, MECs from mice expressing transgenic CYT-2 isoform showed increased proliferation and no lumen formation in 3D cultures compared with controls. The effect of expressing these isoforms on MEC migration was not explored in this study and it is not clear whether CYT-1 isoform has the ability to mediate migration of both embryonic and postnatal MECs.
To conclude, we present evidence that Nrg3 promotes changes in cell adhesion dynamics and morphology of MECs and have identified a role for Nrg3 in promoting the maturation of the primitive mammary epithelium until it is fully specified to a mammary cell fate. Nrg3 is likely to promote mammary epithelial aggregation through stimulation of the JM-a CYT-1 isoform of Erbb4. We propose a working model (Fig. 8) for the formation of MP3 in which Nrg3/Erbb4 signaling facilitates progressive maturation of epithelial progenitor cells until they achieve a fully specified mammary epithelial phenotype. This is achieved by Nrg3 promoting cell spreading using autocrine signals, which leads to cell hypertrophy, an event that coincides with maturation to acquire the mammary cell phenotype. Nrg3-mediated signals are needed to regulate condensation of mammary mesenchymal precursor cells, presumably by stimulating signal(s), which remain to be identified. Mesenchymal condensation may induce Pth1r expression in mammary mesenchyme precursors, which are then activated and undergo mammary mesenchymal specification. Mammary specification coincides with expression of mammary mesenchymal markers and cell population expansion through increased cell mitosis. In summary, we show Nrg3 signaling is required for both epithelial and mesenchymal mammary specification of embryonic progenitor cells.
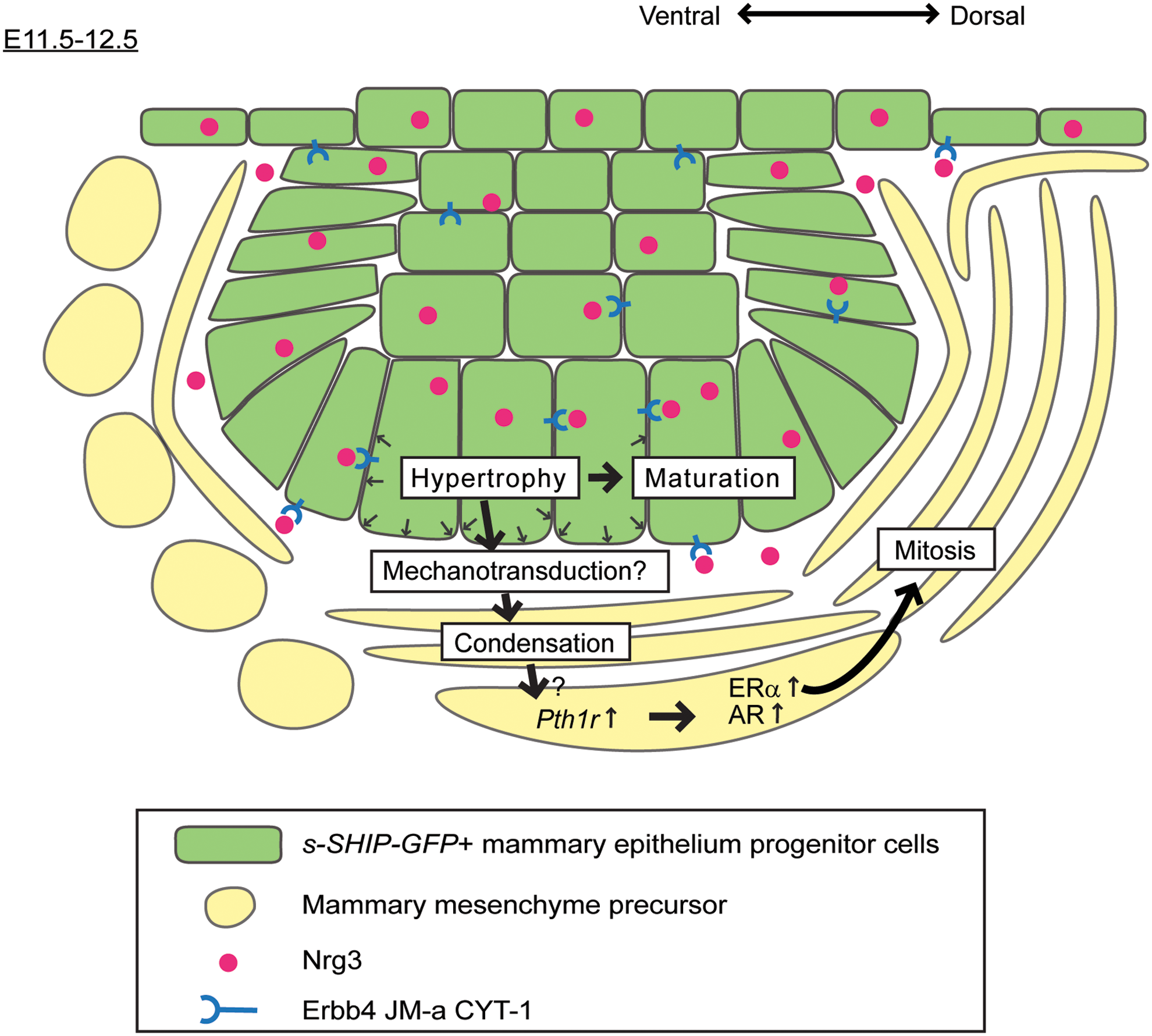
Model for Nrg3/Erbb4 signaling in mammary epithelial and mesenchymal specification. Nrg3 expression within the nascent mammary primordial epithelial cells from E11.5 onward leads to changes in morphology of Erbb4 JM-a CYT-1-expressing mammary progenitor cells, resulting in an increase in cell volume and mammary cell phenotype acquisition. At E12.0, Nrg3-mediated signals are transmitted from the mammary primordial epithelium to adjacent mesenchymal cells, leading to their condensation by mechanisms that remain to be identified. Either Nrg3-mediated condensation or other signals lead to Pth1r expression in adjacent mesenchymal cells. Mesenchymal cells can then respond to activation of Pth1r, which specifies mammary cell fate to mesenchymal progenitors and leads to their differentiation (MM marker expression) and increased mitosis. Color images available online at
Footnotes
Acknowledgments
We thank the Breakthrough Histopathology Facility for their assistance, the late Professor Larry Rohrschneider for s-SHIP-GFP mice, and Professor Klaus Elenius for Erbb4 JM-a, CYT-1/CYT-2 constructs. This research was funded by Breakthrough Breast Cancer. We acknowledge NHS funding to the NIHR Biomedical Research Centre.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.