
Abstract
Creatine is a nitrogenous organic acid known to function in adenosine triphosphate (ATP) metabolism. Recent evidence indicates that creatine regulates the differentiation of mesenchymal stem cells (MSCs) in processes such as osteogenesis and myogenesis. In this study, we show that creatine also has a negative regulatory effect on fat cell formation. Creatine inhibits the accumulation of cytoplasmic triglycerides in a dose-dependent manner irrespective of the adipogenic cell models used, including a C3H10T1/2 MSC line, 3T3-L1 preadipocytes, and primary human MSCs. Consistently, a dramatic reduction in mRNA expression of adipogenic transcription factors, peroxisome proliferator-activated receptor γ (PPARγ) and CCAAT/enhancer-binding protein α (C/EBPα), glucose transporters, 1 and 4 (Glut1, Glut4), and adipocyte markers, aP2 and adipsin, was observed in the presence of creatine. Creatine appears to exert its inhibitory effects on adipogenesis during early differentiation, but not late differentiation, or proliferation stages through inhibition of the PI3K-Akt-PPARγ signaling pathway. In an in vivo model, administration of creatine into mice resulted in body mass increase without fat accumulation. In summary, our results indicate that creatine downregulates adipogenesis through inhibition of phosphatidylinositol 3-kinase (PI3K) activation and imply the potent therapeutic value of creatine in treating obesity and obesity-related metabolic disorders.
Introduction
O
Obesity results from hyperplasia (increase in adipocyte number) and hypertrophy (increase in adipocyte volume) of adipocytes, which are influenced by both genetic and environmental factors [2,3]. In particular, hyperplasia correlates more significantly with disease severity and is marked in seriously obese patients [4]. One of the main reasons for increase in the adipocyte number is generation of new adipocytes from mesenchymal stem cells (MSCs), which can also give rise to osteocytes, chondrocytes, and myocytes [3,5]. Therefore, understanding the factors regulating adipogenic differentiation from MSCs may lead to development of therapeutic targets to treat obesity and obesity-related complications.
Peroxisome proliferator-activated receptor γ (PPARγ) and CCAAT/enhancer-binding protein α (C/EBPα) are genetic factors known to play important roles in adipogenic differentiation, forming a positive feedback loop with each other [6,7]. Expression of these transcription factors is elevated during differentiation and regulates the expression of essential downstream adipogenic genes [8]. PPARγ is especially considered a master determination factor because PPARγ-deficient cells do not promote adipogenesis even in the presence of C/EBPα [9].
Recent studies show that various transcription factors simultaneously regulate closely related differentiation processes. In many cases, inducers of differentiation of one lineage often repress differentiation into other cell types [10]. For example, neurogenic factors largely inhibit gliogenesis and vice versa [11]. This relationship is also observed between osteogenesis and adipogenesis. For example, the key adipogenic transcription factor PPARγ strongly inhibits osteoblastogenesis [12].
Creatine is a nitrogenous organic acid synthesized by muscle, the kidney, and the brain, and is also taken in from food [13]. Creatine is mainly known for its function as a spatial and temporal energy buffer [14]. During a period of low energy need, a phosphate group from adenosine triphosphate (ATP) is transferred to creatine by creatine kinase, forming phosphocreatine, the high-energy phosphorylated form of creatine. The large pool of phosphocreatine serves as an energy store to regenerate ATP from adenosine diphosphate (ADP).
Creatine supplements were first consumed by athletes to increase muscle mass, power, and performance [15] and are now increasingly used as important therapeutic agents for patients with muscle diseases as well as other neurodegenerative and neuromuscular diseases, such as amyotrophic lateral sclerosis and Huntington's disease, among others [16,17]. The action of creatine as a myogenic factor in precursor cells has been implicated in muscle mass increase [18 –20]. Creatine was also found to have stimulatory effects on osteoblast proliferation, differentiation, and bone mineralization [21]. Both muscle and bone precursor cells share a common origin, MSCs, which also produce adipocytes as well as muscle and bone cells [3,5]. As mentioned above, adipogenesis and osteogenesis are very often inversely regulated by the same determinants [10,12]. Therefore, these observations raise the question about whether creatine also has a regulatory effect on fat cell formation. However, the potential role of creatine in adipogenesis has not yet been addressed.
In this study, we show that creatine plays an inhibitory role in adipogenic differentiation irrespective of model cell type. These results, combined with previous findings, imply that creatine has pleiotrophic functions in the regulation of MSC differentiation. This suggests the potential of creatine in preventive and therapeutic applications to simultaneously treat multiple diseases, including obesity and osteoporosis as well as neurodegenerative and neuromuscular diseases.
Materials and Methods
Cell culture and differentiation
3T3-L1 and C3H10T1/2 cells were obtained from American Type Culture Collection (Manassas, VA), and MSCs from human umbilical cord blood were isolated by MEDIPOST Co., Ltd. (Seoul, Korea) as described [22]. Protocols related to the use of human MSCs (hMSCs) were approved by the Institutional Review Board of MEDIPOST Co., Ltd. All cells were differentiated according to standard protocols. Briefly, at 2 days postconfluence (defined as day 0), 3T3-L1 cells were treated with a combination of 1 μM dexamethasone (a synthetic glucocorticoid agonist), 0.5 mM 1-methyl-3-isobutylxanthine (cAMP phosphodiesterase inhibitor), and 5 μg/mL insulin (DMI; all from Sigma, St. Louis, MO) [23 –25] and 10% fetal bovine serum (FBS; Invitrogen, Carlsbad, CA) for 4 days. Because dexamethasone is known to have antiadipogenic effects when added at later stages of adipocyte maturation [26], differentiation was continued on day 4 by switching DMI media to late differentiation media supplemented with 5 μg/mL insulin and 10% FBS and cultured for another 3 days for full maturation. For C3H10T1/2 or hMSCs, 1 μM troglitazone (a specific PPARγ agonist [27]; Sigma) or 1 μM troglitazone plus 100 μM indomethacin [28] (Sigma) was additionally included, respectively, in both the early and late differentiation media to expedite adipogenesis. For hMSCs, the early and late differentiation stages of hMSCs were extended to 10 and 12 days, respectively.
Cytotoxicity assay
The XTT assay was performed to determine the cytotoxicity of creatine according to the manufacturer's protocol (Biological Industries, Kibbutz Beit Haemek, Israel). Briefly, 3T3-L1 cells or C3H10T1/2 cells were allowed to grow in a 96-well plate in the presence of various concentrations of creatine for 72 h. On the day of the assay, cultures were incubated with 50 μL of reaction solution containing XTT (solution prepared just before use) at 37°C for 2 h. The plate was then gently shaken to evenly distribute the newly formed orange-colored formazan dye in the wells. Absorbances were measured at 495 nm against a background control in the linear range of the absorption curve using a Benchmark microplate reader (Bio-Rad, Hercules, CA).
Oil Red O staining
Maturation of adipocytes was assessed by Oil Red O staining of lipid droplets as previously described [29]. Briefly, cells were washed with phosphate-buffered saline (PBS; Invitrogen) and fixed in 4% (w/v) paraformaldehyde (PFA; Sigma) for 15 min. Then, cells were stained in fresh 0.3% Oil Red O solution (Sigma) for 1 h and washed with 60% isopropanol (Merck, Darmstadt, Germany) and H2O.
Construction of retroviral vector plasmid containing CA Akt, PPARγ, and CA PI3K
To generate the retroviral vector MSIG [31] constitutively active (CA) protein kinase B (Akt) construct, a 1.2 kb BamHI fragment from pMSCV/CA Akt (a kind gift from Dr. Tae Woo Kim, Korea University, Seoul, South Korea) [30] was inserted into the unique BamHI site of the MSIG. MSIG-PPARγ was constructed by inserting a blunted 1.5 kb XbaI/NotI fragment from pcDNA-flag PPARγ (#8895; Addgene, Cambridge, MA) [32] into the BamHI site of MSIG. The CA phosphatidylinositol 3-kinase (PI3K)-expressing vector, pWPI-PI3K p110 CAAX, was generated by inserting a 3.2 kb BamHI fragment from pBabe puro PI3K p110 CAAX (#13339; Addgene) into the lentiviral vector pWPI (#12254; Addgene). All restriction and modifying enzymes were purchased from Takara (Shiga, Japan).
Retroviral vector production and transduction
The method of retroviral vector production has been described previously [33]. Briefly, the retroviral construct was transfected into 293T cells with gag-pol (pCA-gag-pol) and env-expressing vector (VSV-G) using Lipofectamine-PLUS reagent (Invitrogen). Supernatant was collected 48 h after transfection, filtered through a 0.45-μm filter, and frozen at −80°C until used. Concentrated viral stocks were prepared by ultracentrifugation at 25,000 rpm for 90 min at 4°C in an SW28 rotor (Beckman-Coulter, Fullerton, CA). Pellets were resuspended in 50 μL of PBS at 4°C for about 12 h, and virus aliquots stored at −80°C. For transduction, cells were seeded at 1×105 in six-well plates on the previous day. Viral supernatants were added in the presence of polybrene (final concentration 8 μg/mL; Sigma). The viral titer was determined by measuring the percentage of GFP-positive NIH3T3 cells transduced with different dilutions of virus stock.
Western blot
3T3-L1 cells were lysed using the RIPA buffer (Sigma) with a protease and phosphatase inhibitor cocktail (Pierce Biotechnology, Rockford, IL). Equal amounts of protein (20–40 μg, depending on target protein) were resolved in 10% (w/v) sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene fluoride (PVDF) membranes (PALL, Port Washington, NY). The membranes were blocked with TBST [150 mM NaCl, 10 mM Tris/HCl, 0.1% (v/v) Tween 20, pH 8.0] containing 5% (w/v) skim milk and analyzed with the following primary antibodies: anti-PPARγ (1:500), anti-Akt (1:500), anti-phospho-Akt (S473; 1:400), anti-PI3K (1:500), anti-phospho-PI3K (Y458/Y199, 1:200; all from Cell Signaling Technology, Danvers, MA), anti-C/EBPα (1:300; Santa Cruz Biotechnology, Santa Cruz, CA), and anti-β-actin (1:500; Sigma). The protein bands were visualized with the enhanced chemiluminescence system (Atto, Tokyo, Japan) and X-Omat film (Kodak, Rochester, NY). All blots were incubated overnight with a primary antibody at 4°C and with horseradish peroxidase-conjugated anti-mouse or anti-rabbit secondary antibodies (1:10,000; Pierce Biotechnology) for 2 h at room temperature.
Reverse transcription and quantitative real-time polymerase chain reaction
Total RNA was prepared from cultured cells using Trizol reagent (Qiagen, Inc., Hilden, Germany) and cDNAs were synthesized from 400 ng of each RNA sample by using an oligo(dT) primer and MMLV-RT enzyme (Promega, Madison, WI). Quantitative real-time polymerase chain reaction (PCR) was performed using the Takara Thermal Cycler Dice Real Time System TP800 (Takara) according to the Smart Cycler System's protocol (Takara) with the following primers: PPARγ-F (5′-ctgtcggtttcagaagtgcct-3), PPARγ-R (5′-cccaaacctgatggcattgtgagaca-3′), C/EBPα-F (5′-ccaagaagtcggtggacaaga-3), C/EBPα-R (5′-cggtcattgtcactggtcaact-3), aP2-F (5′-gaattcgatgaaatcaccgca-3), aP2-R (5′-ctctttattgtggtcgactttcca-3), Adipsin-F (5′-tccgcccctgaaccctacaa-3), Adipsin-R (5′-taatggtgactaccccgtca-3), Glut1-F (5′-cctgctcatcaatcgtaacgagg-3), Glut1-R (5′-cgaccctcttctttcatctcc-3), Glut4-F (5′-atagtctaaagcgcct-3), Glut4-R (5′-ccgagaccaacgtgaa-3), β-actin-F (5′-caaaagccacccccactcctaaga-3), and β-actin-R (5′-gccctggctgcctcaacacctc-3). All quantitative PCR reactions were performed by predenaturing at 95°C for 30 s, 43 cycles of two-step amplification at 95°C for 5 s, 60°C for 1 min, and a last cycle at 95°C for 15 s, 60°C for 30 s, 95°C for 15 s, irrespective of target genes.
Immunofluorescence microscopy
Differentiated cells were washed twice with PBS and fixed with methanol for 15 min. After washing thrice with PBS, cells were blocked for 30 min with PBS containing 1% FBS. Samples were then incubated overnight at 4°C with anti-C/EBPα (1:200; Santa Cruz Biotechnology) or anti-PPARγ (1:500; Cell Signaling Technology) primary antibody, washed thrice in PBS, and incubated for 1 h at room temperature with Alexa-555-conjugated secondary antibody (1:500; Invitrogen) diluted in blocking solution. Cells were further counterstained with 4′,6-diamidino-2-phenylindole (DAPI; Sigma). Fluorescent images were obtained with an inverted fluorescent microscope (Eclipse Ti; Nikon, Tokyo, Japan). The fluorescence signal was measured at an excitation wavelength of 553 nm (Alexa-555) and 345 nm (DAPI) and an emission wavelength of 568 nm (Alexa-555) and 455 nm (DAPI).
Animals
All animal studies were conducted in accordance with the regulations of the Committee of Animal Care and Use at the Laboratory Animal Research Center of Sungkyunkwan University. A total of 40 male mice of C57BL/6J, 6 weeks old, were purchased from Taconic (Germantown, NY) and randomly assigned to four groups (10 mice/group). Mice in the control group received a standard laboratory chow diet (SD; 10% kCal% fat) (#D12450B; Research Diets, Inc., New Brunswick, NJ) and water ad libitum. Three experimental groups included mice on SD treated with 300 mg/kg creatine, mice on a high-fat diet (HFD, 60% kCal% fat) (#D12492; Research Diets, Inc.), and mice on HFD treated with 300 mg/kg creatine. Creatine administration by intraperitoneal injection and measurement of animal weight were performed every 2 days from the beginning of the study.
Results
Creatine inhibits adipocyte differentiation of 3T3-L1, C3H10T1/2, and primary hMSCs
To test the effects of creatine on adipogenesis, three experimental cell models were used. 3T3-L1 cells are regarded to be committed to the adipocyte lineage [23,34], and C3H10T1/2 cells are multipotent stem cell lines that produce diverse lineages, including fat, muscle, and bone cells [35,36]. Primary hMSCs were also included in this study. When 3T3-L1 or C3H10T1/2 cells were cultured under adipogenic conditions in the presence of creatine during all differentiation stages (Fig. 1A), Oil Red O staining that visualizes accumulated lipids after adipocyte maturation showed that creatine inhibits differentiation into adipocytes in a dose-dependent manner with maximal inhibition at 50 mM (Fig. 1B). Differentiation of hMSCs was also efficiently blocked by creatine treatment (Fig. 1C). The inhibition was not due to a cytotoxic effect of creatine on these cells (Fig. 1D). These results indicate that creatine plays a repressive role during adipogenesis irrespective of progenitor cell type.
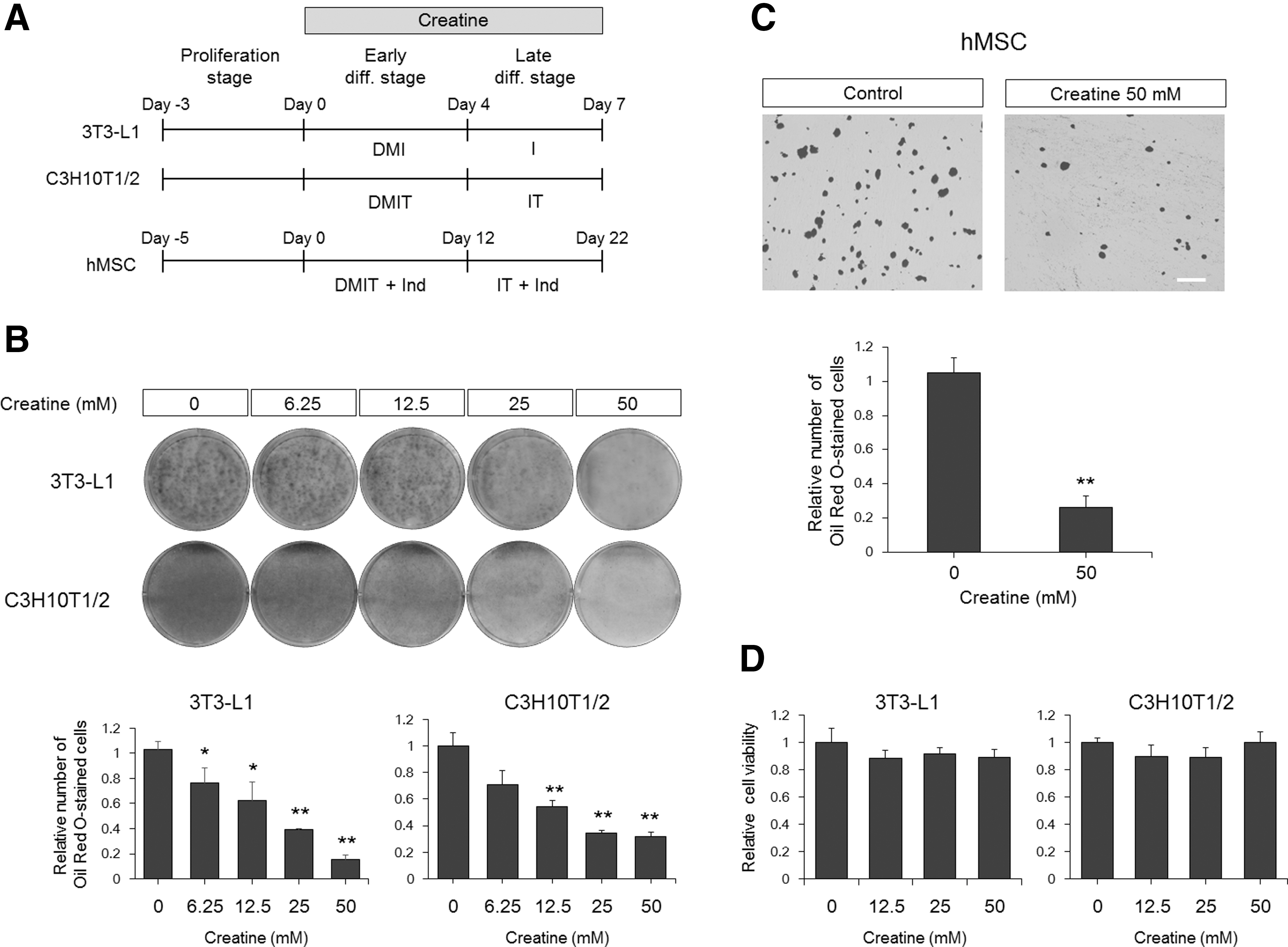
Creatine downregulates adipogenesis.
Creatine represses the expression of adipogenesis- and adipocyte-related genes
Next, quantitative real-time PCR was performed to determine the expression of adipocyte marker genes and transcriptional regulators that are known to be important for adipogenic differentiation. mRNA levels of the mature adipocyte markers, adipocyte protein 2 [aP2, also known as fatty acid-binding protein 4 (Fabp4)] [37] and adipsin [38], were decreased by creatine in a similar manner as observed by Oil Red O staining (Fig. 2A).
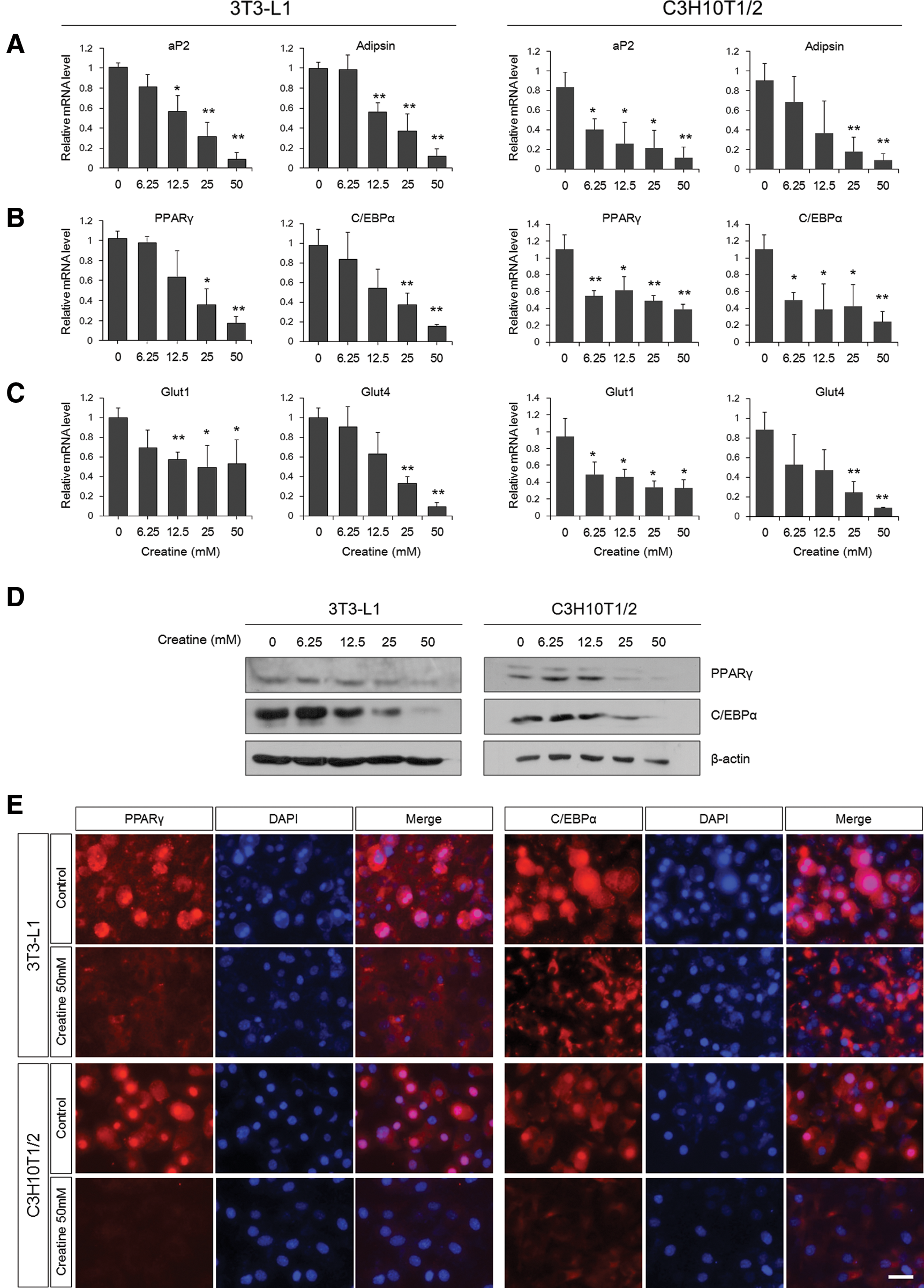
Decreased expression of adipogenesis-related genes upon creatine treatment. Quantitative real-time polymerase chain reaction (PCR) analysis to determine the mRNA levels of the adipogenic markers, aP2 and adipsin
Adipogenesis is a multistep process requiring induction of a cascade of transcription factors regulating downstream effector gene expression [3]. Both mRNA and protein expression of the key adipogenic transcription factors, PPARγ and C/EBPα, were reduced by creatine treatment during differentiation (Fig. 2B, D, E). The nuclear localization of transcription factors is necessary for target gene expression. In addition to decreased protein levels, PPARγ and C/EBPα were mainly detected in the cytoplasm of creatine-treated cells compared with strong nuclear accumulation in control cells (Fig. 2E).
Glucose transporter 1 and 4 (Glut1 and Glut4) are important for insulin-induced glucose uptake across the plasma membranes of adipocytes, and simultaneous knockdown of Glut1 and Glut4 abolishes insulin-stimulated glucose transport [39,40]. Therefore, the principal mechanism of glucose disposal, insulin-stimulated glucose uptake, and efficient energy storage by adipocytes requires these glucose transporters. As expected, creatine-treated cells have much less Glut1 and Glut4 expression than control cells (Fig. 2C). However, the reduction of Glut1 was less significant than that of Glut4. This may be due to the possibility that unlike Glut4, which is expressed only in mature adipocytes, Glut1 is produced in the progenitor cells at a moderate level and is also responsible for glucose uptake in nonfat cells. Taken together, these results indicate that creatine blocks the expression of adipogenesis- and adipocyte-related genes, such as adipogenic inducers, adipocyte markers, and genes that are important for mature adipocyte function.
Creatine blocks adipogenesis at the early stages of differentiation
We then asked whether creatine affects adipogenesis during the early or late stages of differentiation. C3H10T1/2 cells were treated with early differentiation media containing creatine (Fig. 3A). After 4 days, the media were replaced with fresh late media without creatine, and lipid accumulation was assessed after 3 additional days. Addition of creatine at the early stages of differentiation resulted in efficient inhibition of adipogenesis, whereas it showed no overt effect when it was presented at the proliferation or late stages of adipogenesis (Fig. 3B). These results indicate that creatine plays a regulatory role in the early events of adipogenesis and it cannot block fat cell maturation once adipogenesis is triggered.

Creatine inhibits adipogenesis when it is present during the early stages of differentiation.
Creatine inhibits adipogenesis independent of disturbances in energy metabolism
The most well-known function of creatine in the body is the regulation of energy levels by receiving or transferring high-energy phosphates from ATP or to ADP, respectively [14]. Therefore, inhibition of adipogenesis by creatine may result from disturbances in energy metabolism during differentiation. To test this possibility, we applied phosphocreatine to C3H10T1/2 cells during all differentiation stages and assessed the effects on adipogenic differentiation (Fig. 4A). Phosphocreatine also efficiently downregulated adipogenic differentiation of C3H10T1/2 cells in a dose-dependent manner similar to what was observed with creatine (Fig. 4B). The results of ATP supplementation provided more direct evidence of independence of the cellular energy level from creatine action on adipogenesis. Addition of ATP to creatine-treated cultures from the first day of differentiation did not rescue reduced lipid accumulation (Fig. 4C, D). These results suggest that the creatine moiety itself exerts a negative effect on adipogenesis, regardless of its phosphorylation state, and imply that creatine-mediated repression of adipogenesis does not result from altered energy supplies within cells.
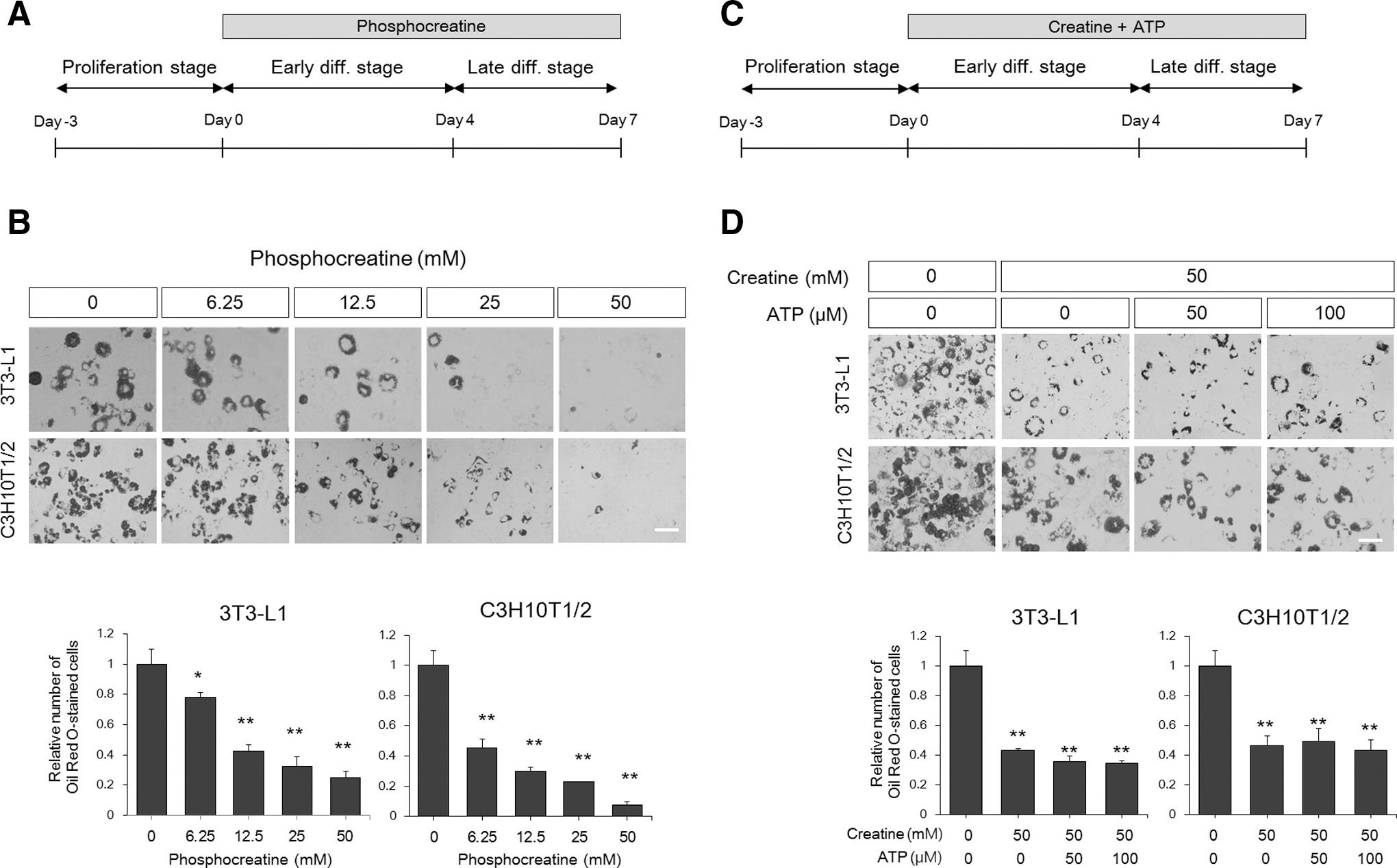
Creatine inhibits adipogenesis independently of the cellular energy level.
Creatine inhibits adipogenesis through suppression of PI3K-Akt-PPARγ signaling cascade activation
Insulin is a critical initiator for differentiation into fat cells. Among many intracellular signal transducers, the activation of PI3K, Akt, mitogen-activated protein kinase (MAPK), and glycogen synthase kinase 3β (GSK3β) was analyzed because of their involvement in adipogenesis as key downstream effectors of the insulin action cascade [41 –43]. Upon acute insulin stimulation, creatine addition resulted in pronounced inhibition of PI3K and Akt levels, but not MAPK and GSK3β, as shown by western blotting (Fig. 5A). The importance of PI3K suppression by creatine was confirmed by rescue experiments using CA Akt and the master adipogenic transcription factor PPARγ. Expression of CA Akt or PPARγ completely reversed the inhibitory effects of creatine on adipogenesis to a normal level (Fig. 5B, C). These results indicate that creatine is an upstream regulator of Akt and PPARγ that cannot reverse the effects of Akt activation or PPARγ-induced adipogenesis, and this hypothesis was further confirmed by rescue experiments using a CA PI3K, the CAAX motif-containing p110 subunit of PI3K. The inhibitory effect of creatine was relieved by introduction of CA PI3K (Fig. 5D). Taken together, these results suggest that creatine inhibits insulin-induced adipogenesis through the blockade of PI3K activation.
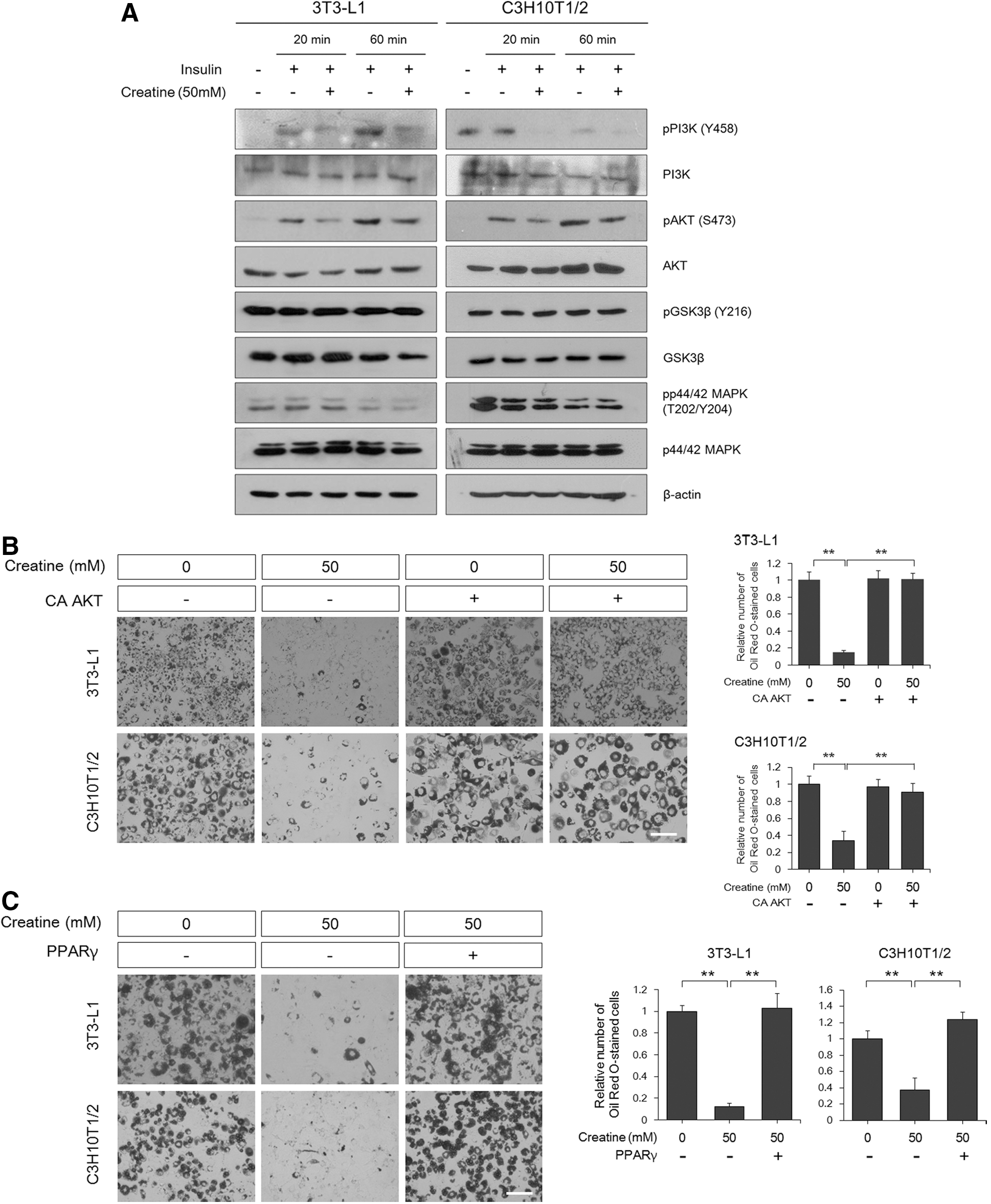
Phosphatidylinositol 3-kinase (PI3K) is a major inhibitory target of creatine during adipogenesis.

Creatine administration into mice in vivo recapitulated the effects observed in humans
Finally, the in vivo effects of creatine were tested using mouse models on a normal-fat diet or HFD. When 6-week-old male C57BL/6J mice were placed on a normal diet, the body weight curves diverged over a 5-week period, with increased weight gain in mice administered with 300 mg/kg creatine every 2 days (Fig. 6A). Conversely, a slight, but statistically insignificant, decrease of abdominal fat accumulation in creatine-treated animals was observed (Fig. 6B). Therefore, the ratio of abdominal fat weight to total body mass was significantly reduced by creatine administration on a normal diet (Fig. 6C). These results mirror the outcomes of creatine supplementation in humans. However, under HFD conditions, no significant changes were observed upon creatine administration. This may be due to caveats of the experimental regimen used in this study.
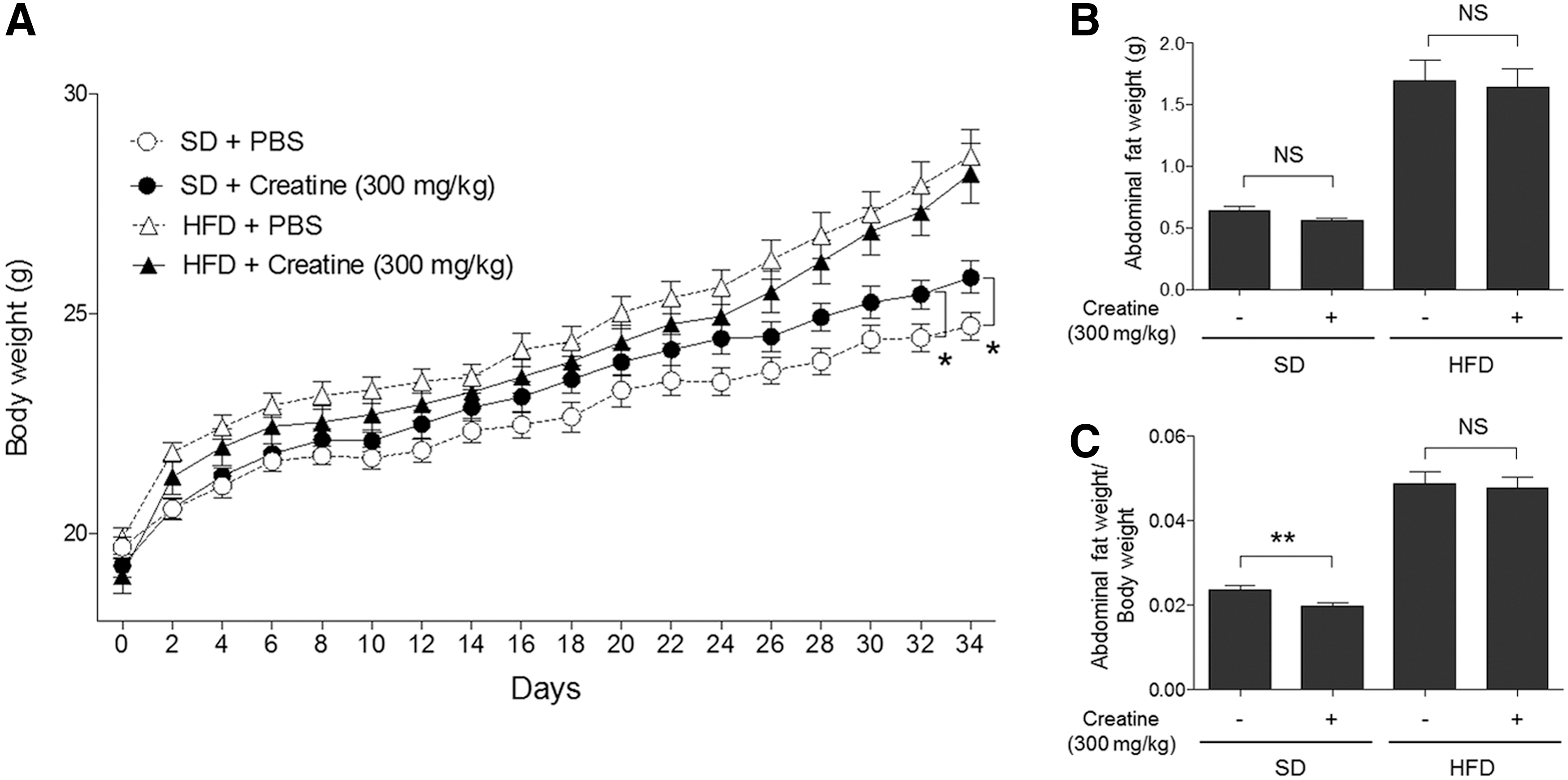
Effects of creatine on body weight gain and abdominal fat accumulation in vivo.
Discussion
Thus far, creatine was known to have two major distinct roles, balancing energy metabolism by regulating ATP generation at the cellular level and increasing fat-free body mass resulting in enhanced exercise performance at the whole-body level [44,45]. Several mechanisms, such as elevated protein synthesis [46,47], reduced protein catabolism [48], and increased satellite cell mitotic activity [49], have been proposed to explain creatine-induced fat-free mass gain. Some reports suggested that enhanced exercise performance upon creatine ingestion may also be directly associated with increased myogenesis; creatine induces muscle-specific gene expression and myogenic differentiation of precursor cells [18 –20]. These observations imply that creatine may act as a modulator of cell signaling pathways and affect the differentiation of unspecified progenitor cells. In addition to myogenesis, creatine was also known to stimulate the differentiation and mineralization of osteoblast-like cells [21]. Both muscle and bone cells share a common progenitor cell type, MSCs, which can also give rise to fat cells, or adipocytes [5].
Numerous factors and multiple signaling pathways that regulate the differentiation of MSCs (adipogenesis, myogenesis, and osteoblastogenesis) have been identified. In many cases, inducers of differentiation into one lineage often inhibit differentiation into other lineages [10]. For example, strong inducers of osteoblastogenesis, such as sonic hedgehog, inhibit adipogenesis [50]. These facts led us to examine the effects of creatine on adipogenesis. From our results, we conclude that creatine is a potent negative regulator in the early events of fat cell generation.
Based on our findings that a CA form of Akt and PPARγ could reverse the inhibitory effects of creatine on adipogenesis, we suggested that creatine acts at a very upstream level of insulin signaling. However, CA Akt and PPARγ are known to be strong adipogenic inducers that can stimulate adipogenesis even without hormone stimulation [51,52]. Therefore, we cannot formally exclude the possibility that creatine acts independently of the Akt/PPARγ pathways because the adipogenic driving force of CA Akt and PPARγ might have been stronger than the inhibitory effect of creatine. However, we attempted to further narrow down the target of creatine action and showed that creatine negatively acts on PI3K activation from rescue experiments using a CA PI3K (p110-CAAX). The role of the CAAX motif in the constitutive activation of target proteins is to facilitate the attachment of proteins to cell membranes. Therefore, since the catalytic- and substrate-binding domains remain identical between wild-type PI3K and CA PI3K, and CA PI3K relieved inhibition of adipogenesis by creatine, it is highly likely that creatine affects the membrane translocation of PI3K, and not the catalytic site of PI3K, upon insulin stimulation.
Since creatine acts as an energy acceptor that receives a phosphate group from ATP [14,17], it is formally possible that the higher than normal concentrations of creatine may lower ATP amounts in the progenitor cells, and this ATP deficiency might cause the block in differentiation, although there are no previous reports on a causal relationship between energy levels and adipogenic differentiation. We provide evidence that phosphocreatine, which is expected to act in an opposing manner to creatine in terms of ATP metabolism, also efficiently interferes with adipogenic differentiation with similar dose dependencies as creatine. Thus, we can conclude that the repressive effects of creatine on adipogenesis are not due to altered energy metabolism, but caused by the creatine moiety itself. This hypothesis was further supported by the observation that ATP addition did not reverse the inhibitory effect of creatine.
We obtained surprising results showing that creatine administration into mice resulted in body mass increase without fat accumulation under normal diet conditions, which mirror the outcomes observed in humans. However, in the HFD model, injection of creatine exhibited a slight, but statistically insignificant, delay in body mass increase during all experimental days and less obvious effects on fat accumulation. This might be due to several possible reasons: first, the low concentration of creatine administered into animals because of its low solubility in water; second, the very short-term period of assessment (∼1 month); and third, prolonged acute HFD conditions may not be the ideal setting to assess the potential benefits of creatine in vivo. In this context, we note that the effects of creatine on the normal diet model that we observed are remarkable, and there is a possibility that the long-term daily administration of creatine at a higher dosage would produce better experimental outcomes both under normal-fat diet and HFD conditions.
Contrary to drug development studies, the aim of this study was to explain the molecular and cellular basis for the human outcome of fat-free body mass increase by creatine through a reverse-translational approach. We provide a clearer understanding of this physiological manifestation in terms of stem cell differentiation. Repression of adipogenesis and concurrent induction of myogenesis of MSCs may result in body mass gain without fat accumulation. In addition, the combined results from this and other studies showing that creatine promotes differentiation of MSCs toward osteogenic cell fates at the expense of adipogenesis also open the possibility that creatine can be applied to the simultaneous treatment of obesity and osteoporosis. Since creatine has been widely used as a supplement, use of creatine as a serious therapeutic agent is expected to be highly cost-effective and less likely to cause potential side effects and safety concerns. Further in-depth and careful in vivo studies that evaluate the potential therapeutic benefits of creatine are still required.
Footnotes
Acknowledgments
This work was supported by the Basic Science Research Program through the National Research Foundation of Korea (#2012R1A1A2003185) funded by the Ministry of Education, Science, and Technology (MEST) and the Samsung Research Fund, Sungkyunkwan University (#2009-0474-000).
Author Disclosure Statement
No competing financial interests exist.