
Abstract
Bone marrow (BM)-derived mesenchymal stromal cells (MSCs), endowed with immunosuppressive and anti-inflammatory properties, represent a promising tool in immunoregulatory and regenerative cell therapy. Clarifying the interactions between MSCs and B-lymphocytes may be crucial for designing innovative MSC-based strategies in conditions in which B cells play a role, including systemic lupus erythematosus (SLE) and rejection of kidney transplantation. In this study, we show that, both in healthy subjects and in patients, in vitro B-cell proliferation, plasma-cell differentiation, and antibody production are inhibited by BM-derived MSCs when peripheral blood lymphocytes are stimulated with CpG, but not when sorted B cells are cultured with MSCs+CpG. Inhibition is restored in CpG+MSC cocultures when sorted T cells are added to sorted B cells, suggesting that this effect is mediated by T cells, with both CD4+ and CD8+ cells playing a role. Moreover, cell–cell contact between MSCs and T cells, but not between MSCs and B cells, is necessary to inhibit B-cell proliferation. Thus, the presence of functional T cells, as well as cell–cell contact between MSCs and T cells, are crucial for B-cell inhibition. This information can be relevant for implementing MSC-based therapeutic immune modulation in patients in whom T-cell function is impaired.
Introduction
M
Most of the reports suggested that B-cell proliferation, differentiation, and cytokine production are inhibited by MSCs [8]. Corcione et al. demonstrated that MSCs were able to suppress, in vitro, the proliferation of B cells activated with anti-immunoglobulin (Ig) antibodies, recombinant CD40L, and cytokines, as well as to interfere with their differentiation, antibody production, and migration [9]. Krampera et al. confirmed these results and showed that the inhibitory effect on B-cell proliferation depended on IFN-γ-induced indoleamine 2,3-dioxygenase (IDO) production by MSCs [10]. By contrast, Traggiai et al. reported that bone marrow (BM)-derived MSCs are able to promote in vitro proliferation and differentiation of transitional and naive B cells isolated from both healthy donors (HDs) and pediatric patients with systemic lupus erythematosus (SLE) upon stimulation with CpG, soluble CD40L, anti-Ig antibodies, and IL-2 [11]. These conflicting results on the interaction between MSCs and B lymphocytes may partly reflect differences in the experimental conditions. In particular, it is important to distinguish the direct action of MSCs on B cells from indirect effects mediated by other cell types contained in the different culture conditions.
In view of their immunosuppressive/anti-inflammatory properties, as well as of their role in sustaining tissue repair and tropism [12,13], MSCs represent a promising immunoregulatory and regenerative therapy for many conditions, including autoimmune disorders [14 –16]. Therefore, clarifying the interactions between MSCs and B-lymphocytes is also important for designing innovative strategies for B-cell mediated disorders.
In this study, we investigated the interactions between MSCs and B cells in vitro, documenting that the inhibitory effects of MSCs on B-cell proliferation, differentiation, and antibody production are largely mediated by T cells.
Materials and Methods
Patients and HDs
MSCs were obtained from residual cells from 15 HDs (age range: 5–32 years) who donated BM cells for transplantation at the Ospedale Pediatrico Bambino Gesù (OPBG), Roma. Peripheral blood from 20 HDs (age range: 23–50 years) was collected and used to perform control experiments. Peripheral blood from seven SLE patients and eight patients who had received kidney transplantation was also collected at the OPBG. The OPBG Institutional Review Board approved the study. All patients and donors or their legal guardian gave written informed consent to utilize cells. Patient clinical data, at the time of analysis, are described in Supplementary Tables S1 and S2 (Supplementary Data are available online at
Cell sorting
Peripheral blood mononuclear cells were isolated from heparinized peripheral blood by Ficoll-Paque™ Plus (Amersham Biosciences) by density-gradient centrifugation and stained with the following antibodies: clone ML5 (anti-CD24), clone UCHT1 (anti-CD3), clone B1.49.9 (anti-CD25), clone HIT8a (anti-CD8), clone RPA-T4 (anti-CD4), and clone M5E2 (anti-CD14) (BD Biosciences). Cells were sorted as following: B cells (CD24+), T cells (CD3+), regulatory T (Treg) cells (CD4+CD25+), monocytes (CD14+), peripheral blood lymphocytes (PBLs) without CD3+ cells, PBLs without CD14+ cells, PBLs without CD4+ cells, PBLs without CD8+ cells, and PBLs without Treg cells, using a FACSvantage SE (BD Biosciences). Cell purity was >98%.
Cell cultures
Total PBLs, CD24+ B cells, CD3+ T cells, CD14+ cells, Treg cells, PBLs without CD3+ cells, PBLs without Treg cells, PBLs without CD4+ cells, PBLs without CD8+ cells, and PBLs without CD14+ cells were labeled with 0.1 mg/mL of 5-chloromethylfluorescein diacetate (CellTracker CMFDA; Molecular Probes). For each culture condition, 2×105 cells were seeded on 96-well plates precoated with or without 2×104 MSCs/well and stimulated with RPMI 1640 (Gibco BRL, Life Technologies), 10% heat inactivated fetal bovine serum (Hyclone Laboratories), 2% L-glutamine (Gibco BRL), 5×10−5 M 2-βmercaptoethanol (Sigma) and 20 mg/mL gentamicin (Gibco BRL), supplemented or not with 2.5 μg/mL CpG-ODN (Hycult Biotechnology). Cell proliferation and phenotypic analysis was performed by flow cytometry using a FACSCalibur Flow Cytometer (BD Biosciences) on day 7 as previously described [17] and supporting information.
Ig production
Quantitative analysis of IgA, IgG, and IgM concentration in culture supernatants was performed by ELISA, as previously described [17].
Cell–cell contact assay using transwells
Total PBLs, CD24+ B cells, and CD3+ T cells were labeled with 0.1 mg/mL CMFDA. For the cultures of total PBLs, 1.5×106 cells were seeded on the upper chamber of 24-transwell plates (6.5 mm Transwell® with 5.0 μm pore polycarbonate membrane insert; Corning). In the lower chamber, 1.5×105 MSCs/well were seeded. For the experiments with sorted cells, the upper chamber contained either CD24+ B cells mixed to sorted T cells at 1:10 ratio or B cells only. MSCs were always in the lower chamber either alone or with sorted T cells. The B/T ratio was always 1/10 and the total number of B+T was 1.5×106/well. CpG was used as previously described. Cell proliferation and phenotypic analysis was performed by flow cytometry using a FACSCalibur Flow Cytometer (BD Biosciences) on day 7.
Cytokine quantitation
Culture supernatants were diluted 1:10 and incubated with anti-cytokine beads (FlowCytomix; Bender Medsystems, eBiosciences) against IFN-γ, IL-2, IL-4, IL-6, IL-10, and TNF-α following the procedures indicated on the manufacturer's instructions. Sample analyses were performed on a FACSanto (Becton and Dickinson) and cytokine concentration was calculated using FlowCytomix-Pro Software (eBiosciences). IL-6 concentration, in culture supernatants, was also measured by ELISA following the manufacturer's instructions (Mabtech AB).
Statistical analysis
Data were analyzed using the StatView statistical program for MacIntosh (StatView Software) and P value was determined with the paired Student's t-test. P values <0.05 were considered to be statistically significant.
Results
Inhibition of B-cell function by MSCs in HDs
Peripheral blood B cells express toll-like receptor 9 (TLR9) and TLR9 ligation through CpG induces cell division and differentiation of memory B cells [17,18]. As CpG stimulates TLR9, an innate receptor that functions independently from the antigen-specific B-cell receptor, it works as a polyclonal activator and, thus, represents a good tool to evaluate B-cell function.
Memory B cells are implicated in autoimmune diseases and in the rejection of transplanted organ, because they produce autoantibodies [19] and anti-HLA class I and II antibodies [20], respectively.
We used CpG stimulation of total PBLs as a readout method to address the issue whether MSCs have immunomodulatory effects on B-cell function. We isolated total PBLs from 13 HDs and, in 13 independent experiments, we cultured the cells with or without CpG in the presence or absence of BM-derived MSCs. After 7-day culture, we measured B-cell proliferation (loss/reduction of CMFDA labeling) and plasmacell generation (identified as CD19+/lowCD27brightCD38bright cells) by flow cytometry and antibody production by ELISA. We found that, in the absence of CpG, MSCs did not affect B-cell survival (Fig. 1A, B, medium and medium+CpG and Supplementary Table S3). In contrast, in the presence of CpG, MSCs significantly inhibit B-cell proliferation as compared to CpG alone (Fig. 1A, upper panels, Fig. 1B and Supplementary Table S3). Frequency of CD19+ proliferating cells dropped from 13.3%±6.7% in CpG stimulated to 6.6%±5.3% (P<0.001) in CpG+MSC stimulated cultures. Moreover, in CpG+MSC cultures, B cells were unable to differentiate into plasmacells (Fig. 1A, lower panels, Sw plasmacells and IgM plasmacells). Impairment in plasmacell differentiation and antibody production was confirmed by the significant reduction of IgM, IgG, and IgA in culture supernatants (see also Fig. 1C for details).

B-cell proliferation and antibody production in peripheral blood lymphocytes (PBLs) stimulated with CpG for 7 days with or without mesenchymal stromal cells (MSCs). Total PBLs were cocultured with MSC in a medium or in a medium with CpG for 7 days. Cells were collected, stained for CD19, CD27, and IgM and analyzed by flow cytometry. Ig production was measured in culture supernatants by ELISA.
Previous studies have shown that MSCs increase proliferation and differentiation of purified peripheral blood B cells stimulated with soluble CD40L, anti-Ig, and IL-2 [11]. Also in our coculture conditions, using CpG, CD19+-sorted B cells showed increased proliferation and antibody secretion in the presence of MSCs (Fig. 2A, right panels). Mean proliferation frequency was 35.1%±15.1% in CpG stimulated cultures and 60.7%±15.8% when MSCs were added to the cultures (P=0.013) (Fig. 2D).
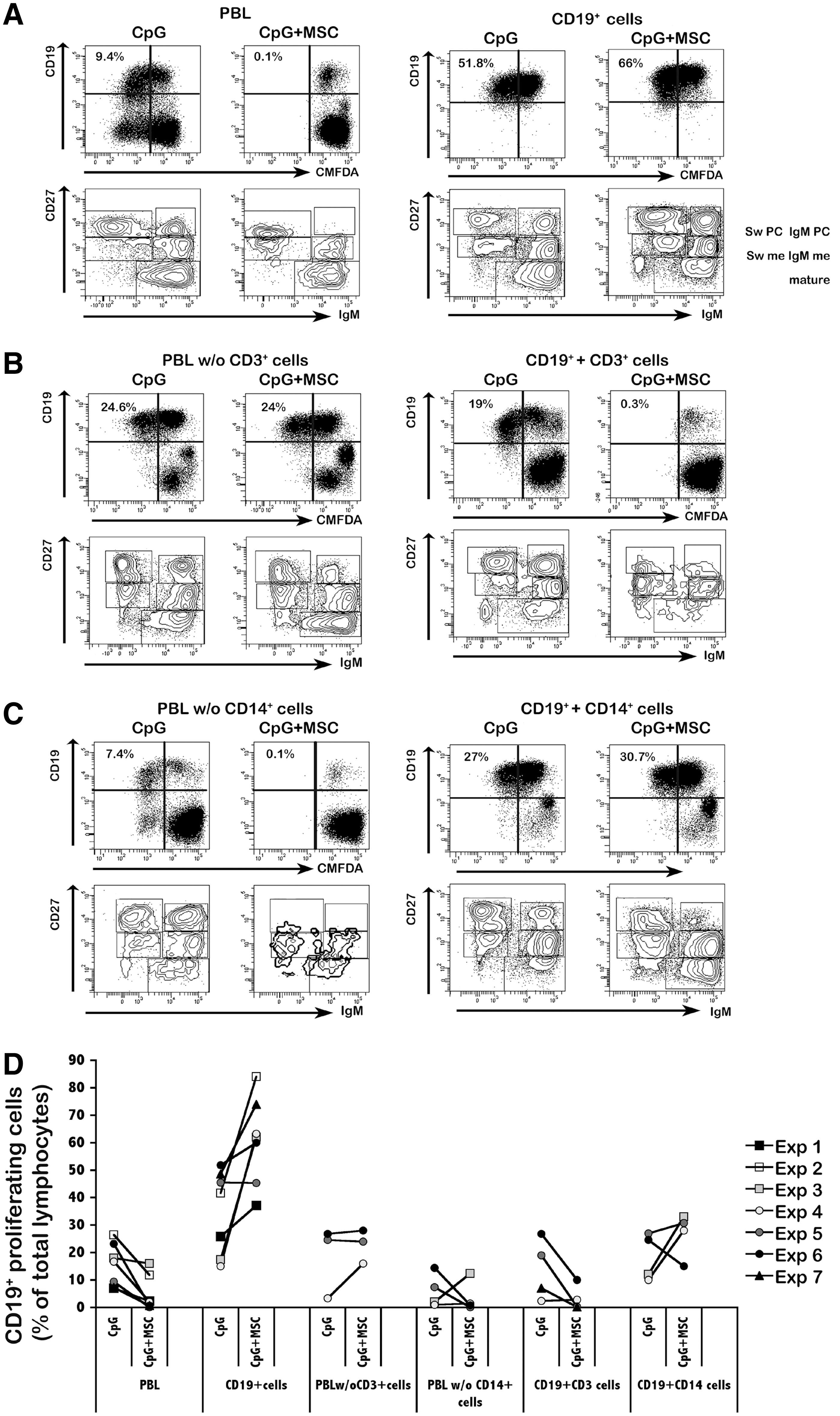
T-cell dependent MSC inhibition of CpG-induced B-cell proliferation. PBLs, sorted B cells (CD19+cells), PBLs depleted of CD3+ cells (PBLs w/o CD3+ cells), PBLs depleted of CD14+ cells (PBLs w/o CD14+ cells), sorted B cells mixed to sorted T cells (1:6) (CD19++CD3+ cells), and sorted B cells mixed to sorted CD14+ cells (1:6) (CD19++CD14+ cells) were stimulated with CpG in the presence or absence of MSCs for 7 days, stained for CD19, CD27, and IgM and analyzed by flow cytometry.
As B cells respond differently to CpG+MSC stimulation when cultured either alone or in the presence of unselected PBLs, we hypothesized that inhibition of B-cell proliferation and Ig secretion is not exerted directly by MSCs, but it is mediated by a third-party subset. To address this point, we performed a series of experiments in which we removed either CD3+ or CD14+ cells from total PBLs before stimulation with CpG with or without MSCs. In parallel, sorted CD3+ or CD14+ cells were added to sorted CD19+ B cells. We mixed the populations respecting the proportions normally observed in PBLs (ie, six CD3+ cells or CD14+ cells: one CD19+ cell). In the absence of CD3+ cells, MSCs did not inhibit B-cell proliferation (Fig. 2B, left panels). The frequency of proliferating B cells did not change or even increase upon CpG stimulation in cocultures with MSCs (Fig. 2D and Supplementary Fig. S1). Inhibition was restored only when purified T cells were added to sorted B cells (Fig. 2B, right panels and Fig. 2D). Both B-cell proliferation and plasmacell differentiation were impaired in CpG+MSC cultures using PBLs depleted of CD14+ cells (Fig. 2C, left panels), but containing T cells. Consistently, addition of purified CD14+ cells to purified CD19+ cells in the CpG+MSC coculture did not have any influence on B-cell proliferation (Fig. 2C, right panels and Fig. 2D).
Overall, these results strongly suggest that, in vitro, T cells play a crucial role in the modulation of B-cell function mediated by MSCs (Supplementary Table S4).
We then addressed the question whether CD4+ or CD8+ T cells contribute to MSC-mediated B-cell inhibition. We depleted PBLs of either CD4+ or CD8+ T cells and stimulated the cultures with CpG in the presence or absence of MSCs. In both coculture conditions, MSCs inhibited CpG-induced B-cell proliferation and antibody secretion (Fig. 3A–C). The frequency of proliferating B cells in MSC cocultures with CD8+ was 0.9%, whereas with CD4+ cells, was 1.6% (Fig. 3A, compare left and right panels).
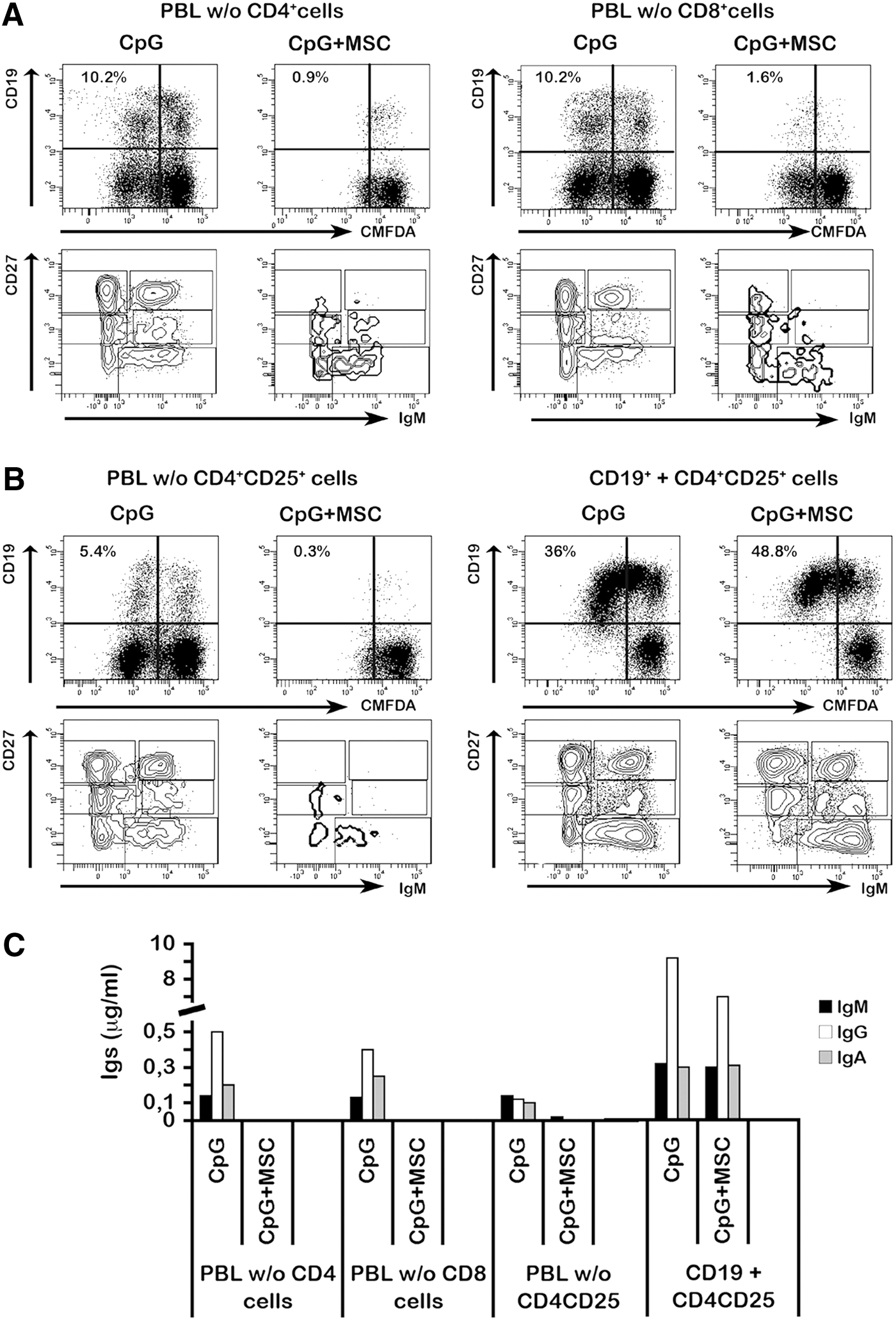
Effect of CD4+, CD8+ or CD4+CD25+ T-cells on MSC-mediated B cell inhibition.
Several studies have shown that in mixed lymphocyte reaction, using alloreactive T cells, MSCs induce differentiation of CD4+CD25+ Treg cells and suppress the proliferation of effector cells [21 –23]. A regulatory role for CD4+CD25+ T cells on B cells is still a matter of debate. In a mouse model of autoimmunity, depletion of Treg cells leads to loss of anergy in B cells and the production of autoantibodies [24]. We never observed an increase of Treg cells in our culture conditions. However, to address the question whether CD4+CD25+ Treg cells modulate B cell under the effect of MSCs, we measured B-cell proliferation after depletion of CD4+CD25+ T cells (PBL without Treg cells). In the absence of Treg cells, the frequency of proliferating B cells was reduced to the same extent as in the cultures containing Treg cells. Thus, removal of Treg cells does not alter MSC-dependent inhibition of B-cell proliferation (Fig. 3B, left panel) and antibody secretion (Fig. 3C). This result is further supported by the observation that the addition of sorted CD4+CD25+ Treg cells to purified B cells (1:4 ratio) does not influence the response of B cells to CpG (Fig. 3B, right panel). Also antibody production did not change when Treg cells were added to the experimental conditions (Fig. 3C). Overall, our results indicate that Treg cells do not play a role in the T-cell-dependent MSC-mediated inhibitory effect on B-cell proliferation and antibody production (Supplementary Table S4).
Inhibition of B-cell function by MSCs in SLE and recipients of kidney transplantation
Traggiai et al. reported that MSCs are unable to inhibit B-cell proliferation and antibody secretion of SLE patients [11]. In view of the findings obtained using cells of HDs, we analyzed B-cell response to CpG in total PBLs, alone or with MSCs, isolated from seven SLE patients and eight subjects that had received kidney transplantation. This choice lies on the rationale that B cells play an important role in the pathogenesis of both SLE and humoral transplant rejection [25 –27].
Since all patients were taking immunosuppressive drugs, the majority of them showed a modest response to CpG stimulation (Supplementary Table S3). Moreover, in contrast to what we observed in HDs, in three out of eight recipients of kidney transplantation (Patient number 1, 5, and 7) and in 2/7 SLE (Patient number 3 and 5) MSCs increased B-cell proliferation (Fig. 4A, B). Inhibition of B-cell proliferation mediated by MSCs was observed only in 3/8 (37.5%) kidney transplant recipients and in 4/7 (57%) SLE patients (Supplementary Table S3).
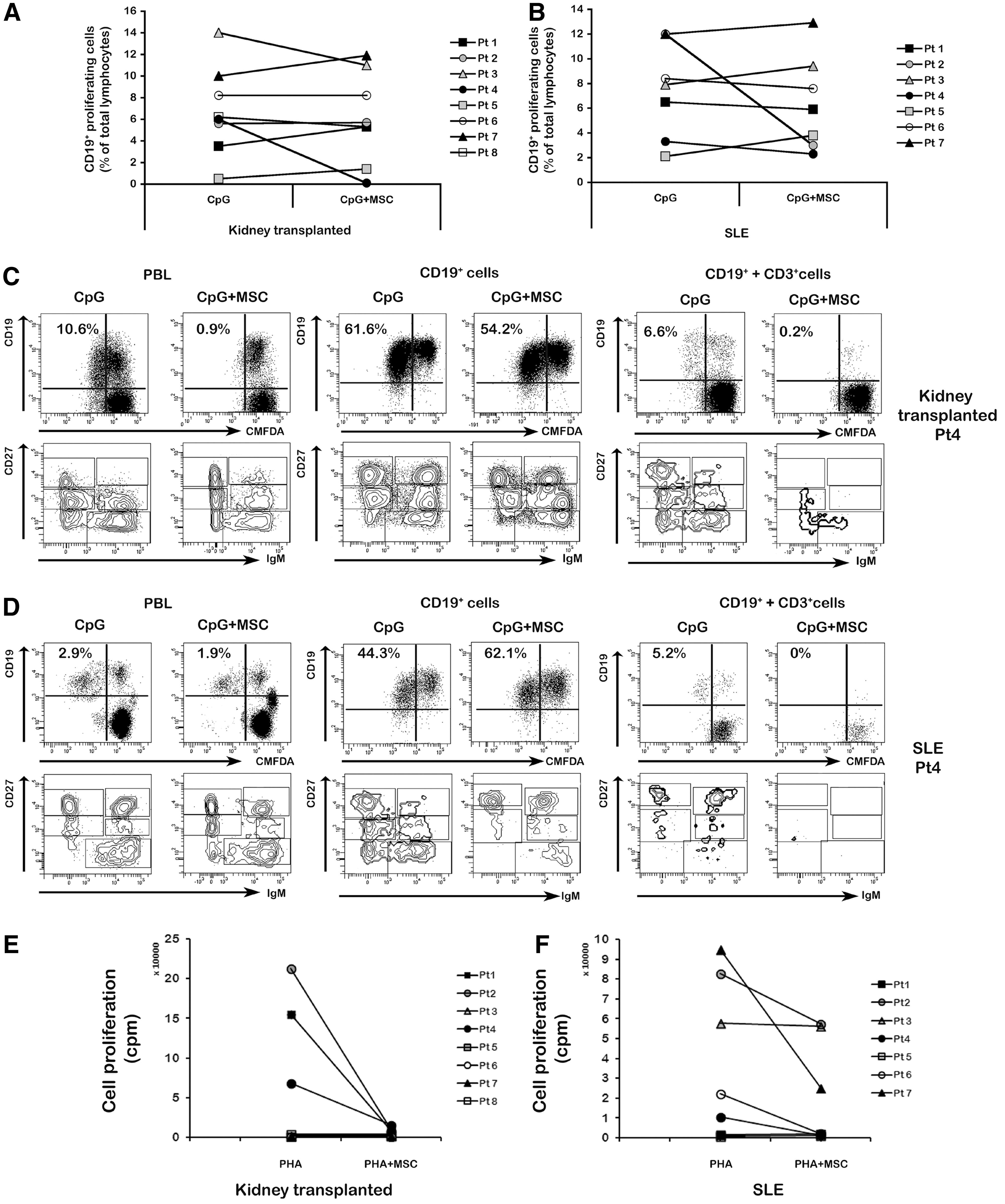
T-cell mediated MSC inhibition of B-cell proliferation and antibody secretion in recipients of kidney transplantation and systemic lupus erythematosus (SLE) patients.
As in HDs, the inhibition of B-cell proliferation and antibody secretion is not directly mediated by MSCs, but is a T-cell dependent effect, and we selected one patient from each group and studied B-cell proliferation under the influence of T cells. To this purpose, we selected patients in whom MSCs had an inhibitory effect on B-cell proliferation and differentiation. We purified CD19+ cells and CD3+ cells from PBLs of the kidney recipient number 4 and from the SLE patient number 4 and cultured CD19+ cells either alone or mixed to CD3+ cells (at 1:6 ratio), together with either CpG or CpG+MSCs. In the kidney recipient, we observed that sorted CD19+ cells stimulated with CpG proliferated equally well in cultures with or without MSCs (proliferating B cells were 54.2% and 61.6%, respectively, Fig. 4C, middle panels). Proliferation was inhibited in the cultures with MSCs when sorted B cells were mixed with sorted CD3+ cells (Fig. 4C, right panels). Differentiation into plasmacells and antibody production were inhibited, in both total PBLs and CD19++CD3+ cell cultures stimulated with CpG in the presence of MSCs (Supplementary Fig. S2A). Similarly, in the SLE patient, we observed that sorted B cells stimulated with CpG in the presence or absence of MSCs proliferated to a comparable extent (44.3% vs. 51.1%, Fig. 4D, middle panels). Coculture of sorted CD3+ cells with sorted B cells in the presence of MSCs abrogated B-cell proliferation (Fig. 4D, left panels) and antibody production (Supplementary Fig. S2B) induced by CpG.
As T-cell function is crucial for the inhibitory effect of MSCs on B-cell function, we evaluated phytohemagglutinin (PHA)-induced T-cell proliferation in our SLE and kidney transplanted patients receiving immunosuppressive treatment both in the presence and absence of MSCs and compared it with that of HDs. As expected, PBLs from HDs stimulated for 3 days with PHA strongly proliferated; in the presence of MSCs (MSC: PBL ratio 1:2), we observed a 68-fold reduction of T-cell proliferation (mean counts per minute, cpm, dropped from 65,354 to 952, see Supplementary Table S5). In contrast, PHA did not induce sizeable T-cell proliferation in 5/8 kidney transplanted patients, probably because of the heavy and long-standing pharmacological immunosuppression that patients had been receiving. In the three patients who responded to PHA, MSCs had strong inhibitory effect on T-cell (Fig. 4E) and a very modest inhibitory effect on B-cell (Fig. 4A) function. In SLE patients, the PHA response was evident in five out of seven cases, but only in three patients MSCs were able to strongly inhibit T-cell proliferation (Fig. 4F). In these three cases, CpG-induced activation of B cells was modestly inhibited by MSCs (Fig. 4B). Overall, our results indicate that, also in patients with autoimmune disorders or who had received organ transplantation, the immunosuppressive effects of MSCs on B cells are mediated by T cells; moreover, our results suggest that in individuals with compromised T-cell function MSCs can be less capable to suppress B-cell activation.
The role of cell contact and soluble factors
Previous studies have shown that the immunoregulatory function of MSCs is exerted both through cell–cell contact and release of soluble factors, including IFN-γ, TNF-α, IL-2, IL-6, IL-10, and TGF-β [5 –7].
To investigate whether the contact with MSCs is necessary to mediate B-cell inhibition, we first stimulated total PBLs with CpG in a transwell chamber in which PBLs were physically separated from MSCs. We found that, in these culture conditions, B-cell proliferation was not inhibited by MSCs indicating that cell contact between MSCs and PBLs is needed (Fig. 5A, lower left panel). We also sorted B and T cells and cultured them with CpG in two conditions. In the first condition, sorted B cells were in the upper chamber and T cells in the lower, together with MSCs. In the second condition, both B and T cells were in the upper chamber and MSCs in the lower. We found that MSC-T cell contact, but not MSC-B cell contact, is crucial for the inhibition of B-cell proliferation (Fig. 5A lower right panel and 5B). These results strongly suggest that the inhibition of B-cell proliferation depends on soluble factors produced either by MSCs or T cells only when MSCs are in contact with T cells.
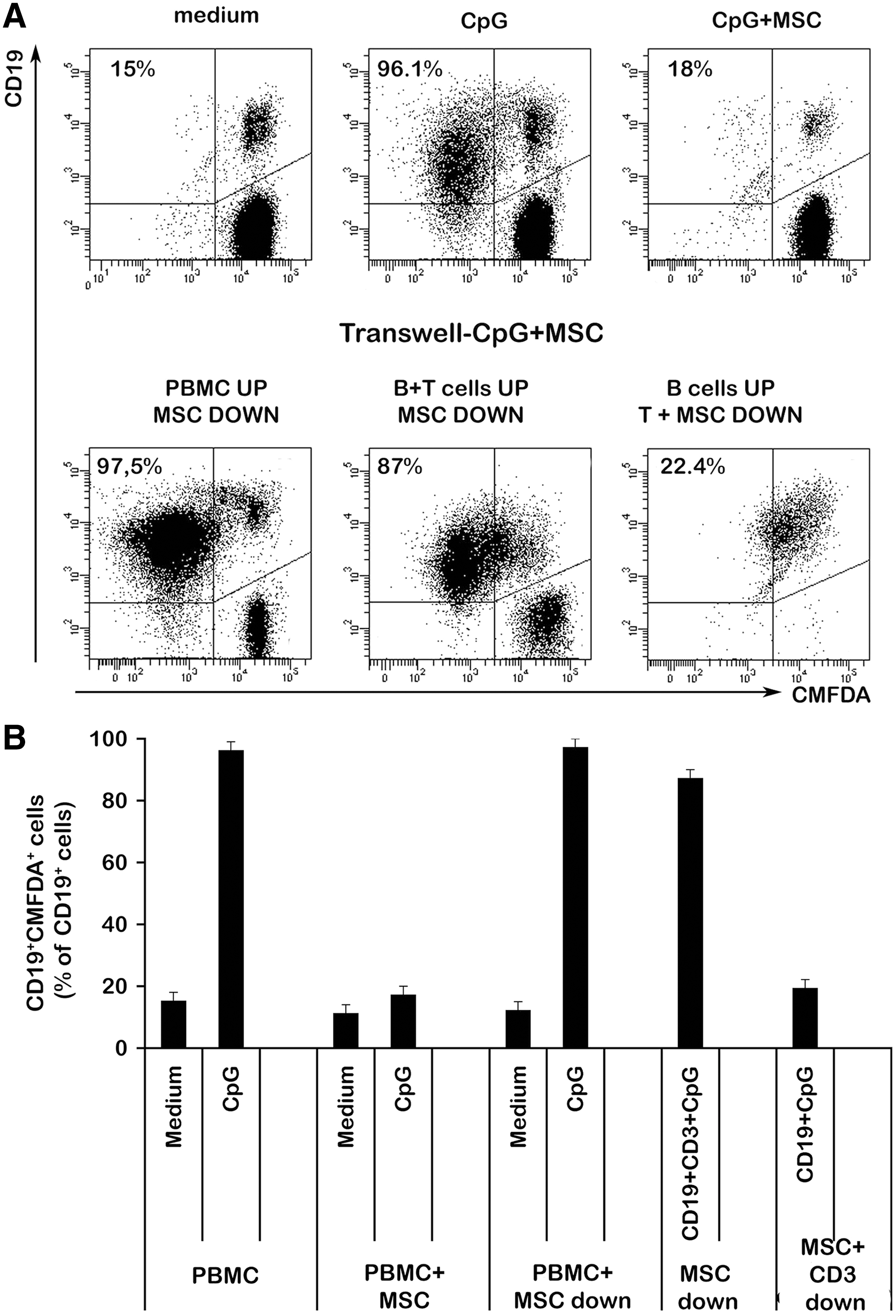
Cell–cell contact effect of MSCs on B-cell proliferation. Total PBMCs and sorted CD19+ B cells mixed to sorted CD3+ T cells (1:10 ratio) were seeded at the upper chamber of a transwell plate previously coated with MSCs and stimulated with CpG for 7 days. In another set of transwell plates, sorted CD3+ T cells were added to MSCs coating the lower chamber and sorted CD19+ B cells were seeded in the upper chamber and stimulated as before. B cell proliferation was analyzed by flow cytometry.
Since cytokines are potential mediators of MSC-dependent inhibition of B-cell function, we measured the concentration of IL-2, IL-4, IL-6, IL-10, IFN-γ, and TNF-α in culture supernatants. We quantified cytokine production in either CpG or CpG+MSC-stimulated PBLs, purified B cells, and sorted B cells mixed to sorted T cells. We found that in all culture conditions while the levels of IL-2, IL-4, and TNF-α were below the detection threshold (<20 pg/mL) (three to five independent experiments), there was an increase of IL-6 in the cocultures with CpG+MSCs (Supplementary Fig. S3A). It has been published that MSCs respond to CpG stimulation by producing IL-6 and IFN-γ [11]. We were unable to detect IFN-γ in culture supernatants of sorted B cells stimulated with CpG+MSCs (Supplementary Fig. S3B), whereas we observed a decrease of IFN-γ in total PBLs stimulated with CpG+MSCs (Supplementary Fig. S3B), as compared with PBLs stimulated with CpG alone. However, IFN-γ remained low in CpG+MSC cultures when sorted B cells were mixed with sorted T cells (Supplementary Fig. S3B), this demonstrating that neither T cells nor MSCs play their inhibitory role through this cytokine.
MSCs led to a reduction of IL-10 in cultures of PBLs and, to a less extent, in sorted B cells mixed with sorted T cells (Supplementary Fig. S3C) stimulated with CpG.
We also analyzed the concentration of IL-6 and IFN-γ in culture supernatants of CpG stimulated B cells in the transwell experiments. We found that, regardless of cell–cell contact, MSCs induced an increase of IL-6 in CpG stimulated cultures as compared to CpG stimulated cultures in the absence of MSCs. As to IFN-γ concentrations they were very similar in all culture conditions.
Discussion
BM-derived MSCs display, in vitro and in vivo, immunosuppressive activity on activated T cells [28], dendritic cells [29,30], and NK cells [31], whereas a direct immunomodulatory effect of MSCs on B cells is still a matter of controversy [9,11,26,32,33]. Similar experimental set-ups, from independent laboratories, using cocultures of MSCs and purified/sorted B cells stimulated with CpG, CD40L, anti-Ig antibodies, IL-2, IL-4, and IL-10 gave conflicting results for what concerns B-cell proliferation and antibody production [8,9,11].
In this study, we show that, in HDs, B-cell proliferation, plasmacell differentiation, and antibody production are inhibited by MSCs in PBLs stimulated with CpG, but not when purified B cells are used. Inhibition is restored in CpG stimulated cocultures with MSCs when sorted T cells are added to the culture of purified B cells. Our observations strongly suggest that inhibition of B-cell proliferation and antibody secretion, by BM-derived MSCs, is mediated by T cells. We also demonstrated that both CD4+ and CD8+ T cell subsets contribute to the MSC-mediated inhibition.
Several studies have shown that MSCs exert immunomodulatory effects on T cells through different pathways [21,23,34,35]. To understand the mechanisms through which MSCs and T cells modulate B-cell function, we investigated the role of cell–cell contact, measured cytokine production, as well as the frequency and function of Treg cells in our culture conditions.
We found that inhibition of B-cell proliferation in CpG stimulated cultures, is dependent on cell–cell contact between MSCs and T cells. Thus, MSCs+T cells modulate the function of B cells through the release of soluble factors. We found a reduction of IFN-γ production in CpG-stimulated PBLs in the presence of MSCs as compared with CpG alone, suggesting that this cytokine may directly or indirectly be involved in the MSC-mediated inhibition observed using total PBLs. IFN-γ, however, was not detected in cultures of sorted B and T cells, this finding raising the hypothesis that IFN-γ is produced only in the presence of other cell types. As to the anti-inflammatory cytokine IL-10, we found that its production was reduced in the presence of MSCs, most probably because of the block of endogenous production by B cells [17].
In our culture conditions, MSCs did not induce the generation of Treg cells. Moreover, in CpG stimulated PBL cultures depleted of Treg cells, MSCs inhibited B-cell proliferation and antibody secretion to a similar extent as that observed in total PBLs. Conversely, sorted Treg cells added to sorted B cells had no inhibitory effect on B-cell proliferation induced by CpG. Thus, Treg cells do not seem to play a role in the inhibition of B-cell function.
The authors and others, have shown that stimulation of memory B cells with CpG induces differentiation in antibody secreting cells in a BCR independent way [17,36]. Thus, exposure to unmethylated DNA, for example bacterial DNA, works as a mechanism of natural recall of memory B cells that contributes to the maintenance of serologic memory. In the case of autoimmune diseases, such as SLE, this mechanism may be the cause of relapse associated with the occurrence of infections. We have previously demonstrated that inhibition of TLR9 signal on B cells from SLE patients blocks secretion of autoantibodies [19]. In this study, we show that MSCs are also able to impair B-cell activation and antibody secretion induced by CpG in SLE patients, provided that T-cell function is preserved.
Infusion of MSCs in animal models of experimental autoimmune encephalomyelitis [37], diabetes [3], and inflammatory bowel diseases [38] demonstrated their efficacy in ameliorating manifestations of the autoimmune disorder. Also in humans suffering from immune-mediated/inflammatory diseases, beneficial effects have been obtained by BM-derived MSC injection in Crohn's patients with refractory fistulas [39]. Several groups have treated SLE-prone mice with MSCs. Although the majority of the studies described successful results [15,16,40], in one report, MSCs not only did not protect mice from developing autoimmunity, but also enhanced autoantibody production and kidney disease [41]. Clinical studies on MSC infusion in SLE patients are still missing, but our results suggest that MSCs can be beneficial only in those patients in whom T-cell function is preserved. In this regard, in vitro inhibition of T- and B-cell function by MSCs might become a predictive test of successful MSC-based treatment in these patients.
Activated B cells promote allograft rejection by producing antibodies against HLA class-I and class-II molecules and by presenting antigens to T cells. Plasmapheresis after transplantation or B-cell depletion at the time of transplantation impairs T-cell mediated alloresponses with increased graft survival [42,43]. Conversely, B cells can promote tolerance either directly by producing IL-10 [44] or by favoring the emergence of Treg cells from naïve T cells [45]. Thus, B-cell depleting strategies may be a suitable strategy to avoid humoral rejection [42,43], but they can be associated with loss of regulatory mechanisms and increased risk of infections [46]. Our experimental results suggest that MSCs are able to impair B-cell function in the presence of T cells. Indeed, in experimental models of kidney transplantation, the success of MSC therapy strongly depends on trafficking and homing of injected MSCs and by the time elapsing between transplantation and MSC infusion [4,47,48]. Discrepancies observed in animal models probably reflect the complexity of the mechanism of action of MSCs, as well as of the different inflammatory conditions [49].
In conclusion, we show for the first time, in both HDs and patients, that the presence of functional T cells is crucial for MSC-mediated inhibition of B-cell proliferation and antibody secretion. Both CD4+ and CD8+ T cells are capable of inducing MSC-dependent B-cell inhibition, whereas Treg cells do not seem to play any role. Cell–cell contact between T cells and MSCs is crucial to modulate B-cell function. Use of allogeneic BM-derived MSCs may represent a good tool to control B-cell mediated diseases, such as graft rejection and autoimmunity. While more studies are needed to develop clinical protocols, our results suggest that stimulation with CpG of total PBLs in the presence of MSCs could be a good test to predict the success of MSC therapy in patients either with autoimmune diseases or at risk of solid organ rejection.
Footnotes
Acknowledgments
The authors thank Dr. C. Pioli for critical review of the manuscript. This work has been partly supported by grants from Istituto Superiore di Sanità (National Program on Stem Cells), MIUR (Ministero dell'Istruzione, dell'Università e della Ricerca, Progetti di Rilevante Interesse Nazionale, PRIN), and by the special grant 5×1000 from AIRC to F.L.; grants from AIRC IG9062 and Bando Giovani Ricercatori 2008 to M.E.B.
Author Disclosure Statement
The authors declare that no competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.