
Abstract
In vivo, bone marrow-derived multipotent mesenchymal stromal cells (MSC) have been identified at sites of tumors, suggesting that specific signals mobilize and activate MSC to migrate to areas surrounding tumors. The signals and migratory mechanisms that guide MSC are not well understood. Here, we investigated the migration of human MSC induced by conditioned medium of Huh-7 hepatoma cells (Huh-7 CM). Using a transwell migration system, we showed that human MSC migration was increased in the presence of Huh-7 CM. Using a human cytokine antibody array, we detected increased levels of MIP-1δ and MIP-3α in Huh-7 CM. Recombinant chemokines MIP-1δ and MIP-3α induced MSC migration. Anti-MIP-1δ and anti-MIP-3α antibodies added to Huh-7 CM decreased MSC migration, further suggesting that MIP-1δ and MIP-3α were implicated in the Huh-7 CM-induced MSC migration. By real-time polymerase chain reaction, we observed an absence of chemokine receptors CCR2 and CXCR2 and low expression of CCR1, CCR5, and CCR6 in MSC. Expression of these chemokine receptors was not regulated by Huh-7 CM. Furthermore, matrix metalloproteinase 1 (MMP-1) expression was strongly increased in MSC after incubation with Huh-7 CM, suggesting that MSC migration depends on MMP-1 activity. The signaling pathway MAPK/ERK was activated by Huh-7 CM but its inhibition by PD98059 did not impair Huh-7 CM-induced MSC migration. Further, long-term incubation of MSC with MIP-1δ increased α-smooth muscle actin expression, suggesting its implication in the Huh-7 CM-induced evolvement of MSC into myofibroblasts. In conclusion, we report that two inflammatory cytokines, MIP-1δ and MIP-3α, are able to increase MSC migration in vitro. These cytokines might be responsible for migration and evolvement of MSC into myofibroblasts around tumors.
Introduction
M
We recently demonstrated that MSC secrete anti-inflammatory molecules such as IL-1Ra, allowing an anti-fibrotic effect that attenuates liver fibrosis in mice [13]. Human MSC do not develop into tumors [14], but some reports indicate that MSC participate in the pathogenesis of cancer by transforming into cancer-associated fibroblasts (CAFs) [15 –19]. Some studies indicated that, when injected systemically, MSC migrate to sites of inflammation and diseased tissues [20 –23]. The in vivo tropism toward gliomas has been extensively studied. It appears that targeting gliomas by MSC is possible [24], but the ability of MSC to migrate over long distances to gliomas is still under debate [25,26]. In vitro, MSC migrate to conditioned medium from gliomas [27], breast cancer [28], and colorectal cancer [29] cells. Related to their tumor-homing properties, MSC have gained attention as potential therapeutic vehicles to deliver anti-tumor agents for cancer therapies [30 –32]. However, one major problem encountered with MSC is their poor migratory property once transplanted. Further insights of the mechanisms regulating MSC migration should help manipulate MSC for their use in specific applications.
So far, soluble factors, such as chemokines, seem to play an important role in inducing MSC migration and the attraction of MSC to tumors. Several chemokines and factors have been identified by systemic screening of conditioned medium of tumor cells, such as SDF-1 [29,33,34], IL-8 [35], MCP-1 [36], and platelet-derived growth factor BB (PDGF-BB) [26,37]. Further, extracellular proteolysis through matrix metalloproteinase (MMP) seems to be involved in the migratory activity of MSC [27,38,39]. Results from studies analyzing chemokines receptor expression in MSC populations are contradictory [35,40 –43]. However, several of these receptors were shown to be implicated in MSC migration in vitro such as CXCR1 and CXCR2 [35], as well as CCR2 and CXCR4 [36].
With the aim of further identifying chemokines involved in MSC migration, we analyzed the human hepatoma cell line conditioned medium (Huh-7 CM), which increased MSC migration similarly to PDGF-BB. In this study, we showed that several chemokines were present in the Huh-7 CM. Two of them, MIP-1δ and MIP-3α, increased MSC migration. This Huh-7 CM-induced migration was not regulated through differential expression of the chemokine receptors. We further observed that αSMA expression was induced in MSC after long-term treatment by Huh-7 CM and MIP-1δ. We concluded that in vitro migration of MSC can be stimulated by chemokines MIP-1δ and MIP-3α and that prolonged treatment of MSC with recombinant MIP-1δ favors the differentiation toward myofibroblasts. Hence, in vivo MIP-1δ and MIP-3α might be important chemoattractants that attract MSC to sites of injury and further favor differentiation into myofibroblasts.
Materials and Methods
Human bone marrow-derived multipotent MSC isolation and culture
Human adult bone marrow cells were collected from femoral heads of patients undergoing total hip arthroplasty after informed consent. This research project was approved by local ethics committees of the University Hospitals of Geneva, Switzerland. Cells were isolated from bone fragments, cultured, and characterized as previously described [44]. Medium used for culture was Iscove's modified Dulbecco's medium (Cambrex), supplemented with 10% fetal calf serum (FCS) (Gibco-Invitrogen), 100 IU/mL penicillin, 100 μg/mL streptomycin (P-S) (Gibco-Invitrogen), dithiothreitol (DTT; Sigma), and 10 ng/mL PDGF-BB (PeproTech EC Ltd.). Cells were expanded as previously described [45,46]. MSC were used between passages three and five.
Preparation of Huh-7, HepG2, and primary human hepatocyte conditioned media
Human hepatoma cells, Huh-7 and HepG2, as well as primary human hepatocytes, were cultured at subconfluence in a 150 cm2 bottle in Dulbecco's modified Eagle's medium (DMEM) supplemented with 5% FCS and P-S for 48 h. Conditioned media were harvested, filtered, and stored at −20°C until use.
Measurement of cytokines in Huh-7 CM
Human cytokine antibody array (C series 1000; Raybiotech, Inc.) was used for the qualitative assessment of 60 cytokines in Huh-7 CM and control medium, as indicated by the manufacturer.
Cell migration assays
The migration capacity of MSC was analyzed using Costar transwell migration chambers (6.5 mm diameter inserts) with 8.0 μm pore size (Corning Incorporated). These transwell chambers (referred to as upper compartment) were inserted into a 24-well tissue culture plate (referred to as lower compartment). Cells were cultured for at least 4 h in DMEM 5% FCS before trypsinization. After trypsinization, cells were washed and 6×103 cells per 40 μL DMEM 5% FCS were plated in the upper compartment. One milliliter control medium (DMEM 5% FCS) containing recombinant human growth factors or cytokines MIP-1δ, MIP-3α, IGFBP-2, or PDGF-BB (20 ng/mL, all from Peprotech), angiogenin (Novoprotein) (100 ng/mL), inhibitor PD98059 (50 μM), or conditioned medium were added in the lower compartment. Neutralizing antibodies against human MIP-1δ (R&D Systems) (10 μg/mL) [47] and MIP-3α (Peprotech) (10 μg/mL) [48] as well as control IgG were used. After 18 h of migration, cells were fixed with methanol at −20°C for 2 min. Nonmigrating cells in the upper compartment were wiped off from the membrane with a cotton swab. Migrating cells were found on the lower side of the membrane and were colored with Hoechst. The total number of migrating cells was quantified by counting blue nuclei using Metamorph software. A migrating index was calculated by expressing the number of migrating cells as a fold increase of migrated cells in control condition (DMEM 5% FCS), which was set at 1.
Cell proliferation assay
MSC were seeded at 5,000 cells per well in a 24-well plate. After overnight incubation, cells were rinsed with phosphate-buffered saline (PBS) and incubated for 4 h in DMEM 5% FCS. Then, cells were exposed to control medium or medium containing MIP-1δ and MIP-3α (50 ng/mL) for 48 h. Cells were then rinsed, fixed with paraformaldehyde (PFA) 4%, and stained with Ki67 antibody. Ki67-positive cells were counted and expressed as a ratio of the total number of cells.
Cell differentiation assay
Differentiation assay was performed as previously described [44]. Briefly, MSC were trypsinized and seeded at high density (25,000 cells per cm2). MSC were exposed to either control medium or MIP-1δ and MIP-3α. Adipogenic differentiation was induced with adipogenic differentiation medium for 3 weeks. Cells were fixed with cold 10% formalin for 1 h, washed twice with water, and stained with Oil-red-O solution (Sigma) for 2 h at room temperature, to reveal triglyceride droplets in the cytoplasm. Cells were washed twice, coverslipped, and observed under an optical microscope (Zeiss Axiophot1; Carl Zeiss AG).
Mitogen-activated protein kinase/extracellular signal-related kinase activation assay
MSC were seeded at 3×104 cells per well on a 24-well plate. After overnight incubation, cells were rinsed twice with PBS and incubated for 4 h in medium without FCS. Then, cells were exposed to conditioned medium or medium containing growth factors for 30 min. Cells were rinsed with ice-cold PBS containing 1 mM orthovanadate (Na3Vo4). MSC were scrapped in DTT loading buffer 2×(0.4 M Tris pH 6.8, 20% glycerol, 4% SDS, 10% DTT), snap frozen, boiled at 95°C for 5 min, and submitted to electrophoresis.
SDS-polyacrylamide gel electrophoresis and western blot analysis
Whole cell extract proteins were separated on 12% SDS-polyacrylamide gel and electro-blotted on polyvinylidene fluoride (PVDF) membranes (Milipore). Membranes were blocked for 1 h with either 5% nonfat dry milk or 5% bovine serum albumin (BSA) in TBST (tris-buffered saline 150 mM NaCl, 20 mM Tris–HCl, pH 7.5) containing 0.1% Tween 20. Membranes were incubated overnight with the primary antibodies diluted in TBST with nonfat dry milk or BSA. Thereafter, membranes were washed in TBST and further incubated with secondary peroxidase-conjugated (HRP-goat anti-mouse or goat anti-rabbit; Bio-Rad) IgG antibody. Protein bands were visualized by the enhanced chemiluminescence system as recommended by the manufacturer (UptiLight HRP Blot Substrate-Reagent A). Antibodies against phosphorylated and total ERK were purchased from Cell Signaling, and anti-GAPDH was purchased from Santa Cruz Biotechnology. Antibody against MMP-1 was purchased from Abcam Antibody against αSMA and was a kind gift of Prof. C. Chaponnier (Geneva). The intensity of signals was evaluated by analyzing the optical density of the spots using Quantity One software.
Real-time polymerase chain reaction
Total RNA was extracted from expanded MSC and from MSC treated by control condition, PDGF-BB, or Huh-7 CM for 48 h using the RNeasy™ Mini Kit. cDNA was synthesized from 100 ng total RNA with SuperScript™. For real-time polymerase chain reaction (PCR), the following SYBR Green human QuantiTect Primers (from Qiagen) were used: MMP-1 (QT00014581), MMP-2 (QT00088396), TIMP1 (QT00084168), CCR2 (QT00000224), CXCR2 (QT00000518), and CCR6 (QT01666140). Other specific primers (Invitrogen) were used: CCR1 (forward TTTGGTGTCATCACCAGCAT and reverse GCCTGAAACAGCTTCCACTC) and CCR5 (forward CTGAGACATCCGTTCCCCTA and reverse CTGCGATTTGCTTCACATTG). Normalization was performed using three genes: glyceraldehyde 3-phosphate dehydrogenase (GAPDH), tata binding protein (TBP), and transferrin receptor protein 1 (TfR1).
Statistical analysis
All values are presented as mean±SD. For statistical analyses, the Student's t-test was used. A P value <0.05 was considered statistically significant.
Results
Huh-7 cell conditioned medium and PDGF-BB induce MSC migration
To investigate MSC migration, we exposed MSC to conditioned medium of primary hepatocytes and of two different human hepatoma cell lines, Huh-7 and HepG2. We used PDGF-BB, a known inducer of migration, as a positive control. The migration, assessed using a transwell system, was increased by PDGF-BB (3.9±0.9-fold) and Huh-7 CM (3.5±1.3) compared with control (Fig. 1a). CM from hepatoma cell line HepG2 and from primary human hepatocytes did not activate MSC migration. Addition of PDGF-BB to Huh-7 CM did not potentiate MSC migration (2.7±0.2) compared with Huh-7 CM alone (Fig. 1b). Addition of an anti-PDGF-BB antibody in Huh-7 CM did not affect cell migration, suggesting that PDGF-BB was not the responsible factor in the Huh-7 CM (Fig. 1c). Other recombinant factors, reported to induce MSC migration, such as SDF-1 [49], IL-8 [35], and TGF-β1 [50], did not affect migration in our assay (data not shown).
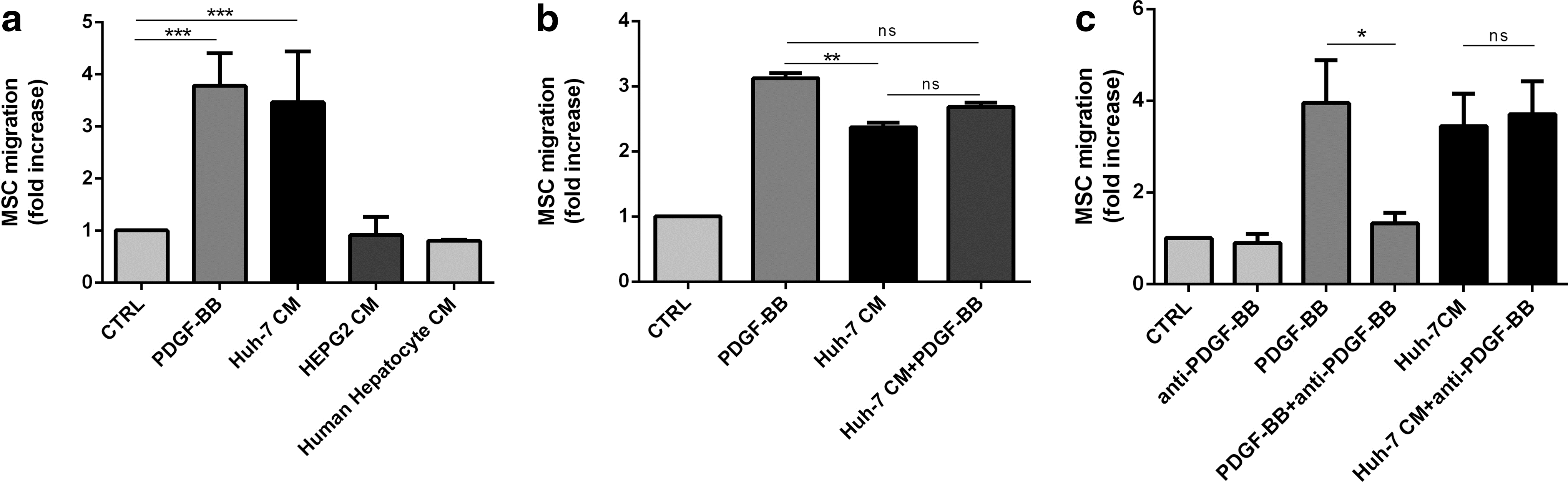
Huh-7 CM and PDGF-BB induces MSC migration.
Huh-7 cells produce angiogenin, IGFBP-2, MIP-1δ, and MIP-3α
To identify cytokines potentially involved in MSC migration induced by Huh-7 cells, we analyzed the cytokine profile of Huh-7 CM. We found that angiogenin, IGFBP-2, MIP-1δ (CCL15), and MIP-3α (CCL20) were secreted by Huh-7 cells (Fig. 2a, b). Intensity of the signals was evaluated by analyzing the optical density of the spots (Fig. 2c). As shown in the graph, chemokines known to be important for MSC migration, such as PDGF-BB, TGF-β1, SDF-1, IGF-1, IL-6, and IL-1β, were not present in the Huh-7 CM, suggesting that they are not responsible for Huh-7-induced MSC migration.

Huh-7 cells produce increased levels of angiogenin, IGFBP-2, MIP-1δ, and MIP-3α.
MIP-1δ and MIP-3α are involved in the increased migration of MSC
To investigate whether MIP-1δ and MIP-3α were involved in the Huh-7 CM-induced migration of MSC, we performed a migration assay in the presence of blocking antibodies directed against MIP-1δ and MIP-3α. As shown in Figure 3a, in experiments using MSC from four different donors, antibodies against MIP-1δ and MIP-3α slightly decreased MSC migration. However, inhibition of MSC migration was statistically significant only for MIP-1δ (Fig. 3a). No additive effect was observed when both antibodies are added simultaneously. Further, using recombinant proteins MIP-1δ and MIP-3α, MSC migration was induced in three out of five experiments when using a physiological concentration of recombinant proteins (50 ng/mL), whereas the use of angiogenin (100 ng/mL) and IGFBP-2 (20 ng/mL) did not modulate MSC migration (Fig. 3b). These results suggest that both chemokines MIP-1δ and MIP-3α are able to induce migration of MSC and also that Huh-7 CM most likely contains other factors increasing migration. We further investigated whether MIP-1δ and MIP-3α modulated adipogenic differentiation and proliferation of MSC, but these activities were not affected (data not shown).
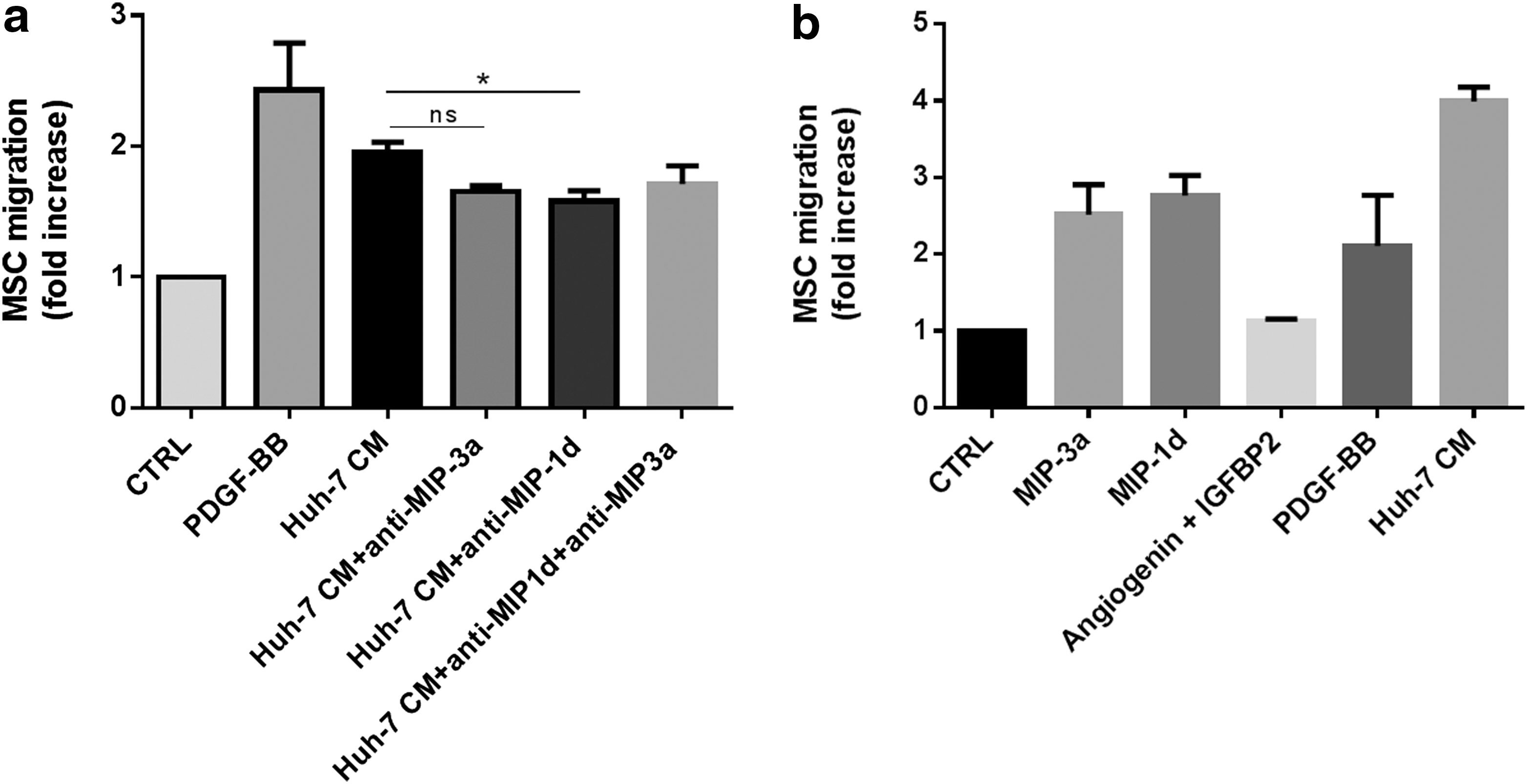
Recombinant chemokines increased transwell migration of MSC in three out of five experiments.
Expression of chemokine receptors and proteins involved in ECM degradation in MSC
The chemokine response can be regulated not only at the level of production of a ligand but also at the level of chemokine receptor expression in the target cell [51]. We thus investigated the expression level by real-time PCR not only of several cognate receptors for the identified chemokines but also of receptors CXCR2, CCR2 and the MMP-1, MMP-2 and their inhibitor TIMP-1, which was shown to be relevant in MSC migration and invasion [27,52]. Chemokine receptors CCR1, CCR6, and CCR5 were expressed at low levels in MSC at various passages from eight different donors. CCR6 was expressed at relatively high levels in four out of seven donors (Fig. 4a). Receptors, CCR2 and CXCR2 expressed at the lowest levels were considered absent. MMP-1, MMP-2, and TIMP-1 were strongly expressed in all MSC populations tested (Fig. 4b). To evaluate whether Huh-7 CM regulates the expression of chemokine receptors for MIP-3α, MIP-1δ, and IL-8 in MSC, we performed reverse transcription quantitative real time polymerase chain reaction (RT-qPCR) of MSC exposed to Huh-7 CM, PDGF-BB, and control medium for 48 h. We observed that expression of CCR1, the receptor for MIP-1δ, did not significantly change in both PDGF-BB and Huh-7 CM-treated cells. Further, the expression level of CCR6, CCR1, and CXCR2 was not significantly affected as well (Fig. 5a). Further, MMP-1, but not MMP-2 or TIMP-1, was significantly increased after exposure to Huh-7 CM and PDGF-BB (Fig. 5b). We further analyzed protein levels for MMP-1. MSC from three different donors were incubated with control, Huh-7 CM, MIP-1δ, and MIP-3α for 72 h and protein expression of MMP-1 was analyzed by western blotting. We showed an increase in MMP-1 protein level (Fig. 5c), which was quantified (Fig. 5d).
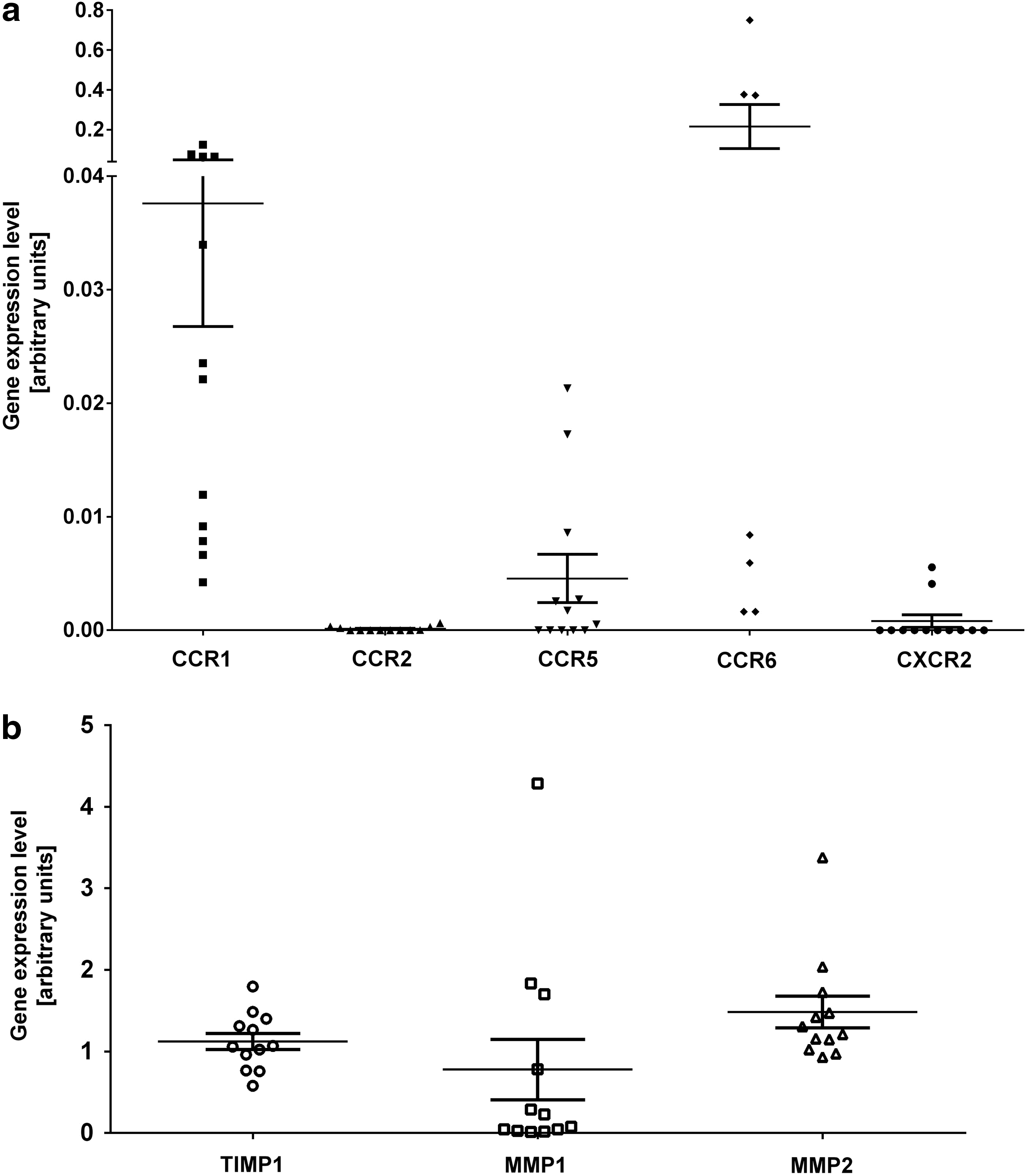
MSC express low levels of chemokine receptors and high levels of proteins involved in EM degradation. MSC isolated from at least six different donors and expanded for three to five passages have been used to perform real-time PCR on total RNA extractions.
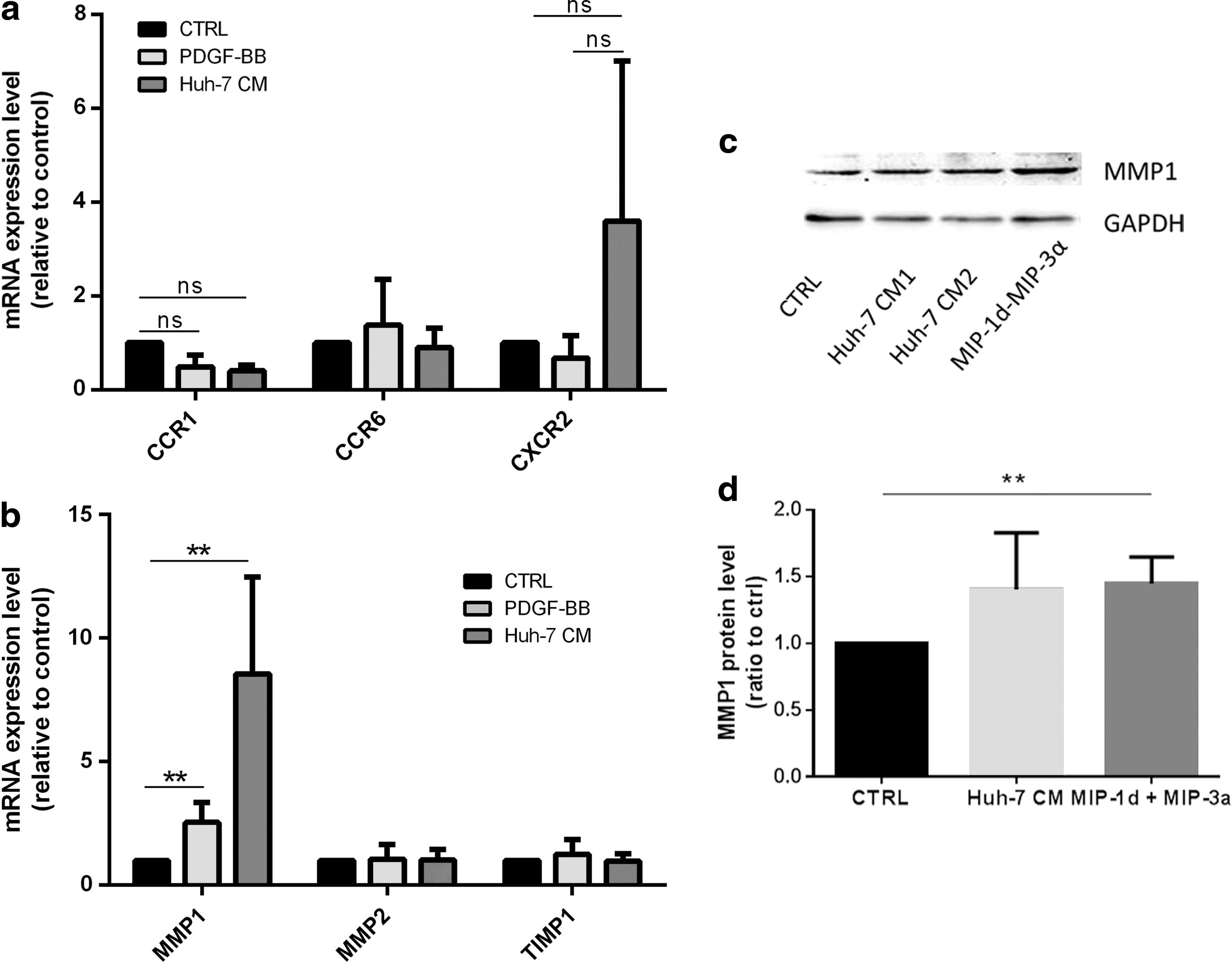
Expression of chemokine receptors and of proteins involved in EM degradation after incubation with Huh-7 CM. MSC isolated from four different donors have been treated with control condition (CTRL), PDGF-BB, and Huh-7 CM for 24 h.
MIP-1δ and MIP-3α mediates activation of ERK pathway in MSC
Considering that ligand binding to CCR1 and CCR6 receptor activates ERK signaling, and that ERK signaling is, in turn, implicated in MSC migration [53,54], we investigated the activation of the ERK pathway. We observed an increased phosphorylation of ERK after 30 min of stimulation with PDGF-BB, Huh-7 CM, as well as MIP-1δ and MIP-3α, when compared with control medium. Adding the inhibitor of MAPK/ERK kinase (MEK) PD98059 decreased phosphorylation of ERK to a basal level of phosphorylation in all stimulated conditions (Fig. 6a, b). Further, the role of ERK phosphorylation in migration was tested by exposing MSC to PD98059 during the migration assay. We observed that PD98059 was most efficiently interfering with migration in PDGF-BB-stimulated MSC (Fig. 6c); however, although not significantly, MSC migration was also decreased in Huh-7 CM, MIP-1δ and MIP-3α-treated cells (Fig. 6d), suggesting that treatment with Huh-7 CM, MIP-1δ, and MIP-3α activates other signaling pathways involved in MSC migration.

Huh-7 CM, MIP-1δ, and MIP-3α mediate activation of ERK pathway in MSC.
MIP-1δ induces αSMA expression in MSC
Bone marrow fibrocyte migration into CCl4-damaged liver is regulated by CCR1 and CCR2 [55]. As MIP-1δ is secreted by Huh-7 cells and induces MSC migration, we investigated its implication in the differentiation of MSC into myofibroblasts. Treatment of MSC with TGF-β1 and Huh-7 CM strongly induced αSMA expression in MSC after 5 days of culture (Fig. 7a, b). In addition, MIP-1δ treatment for 5 days (but not MIP-3α, data not shown) increased αSMA expression in MSC (Fig. 7a, b), suggesting that prolonged exposure to MIP-1δ is implicated in the evolvement of MSC into myofibroblasts.
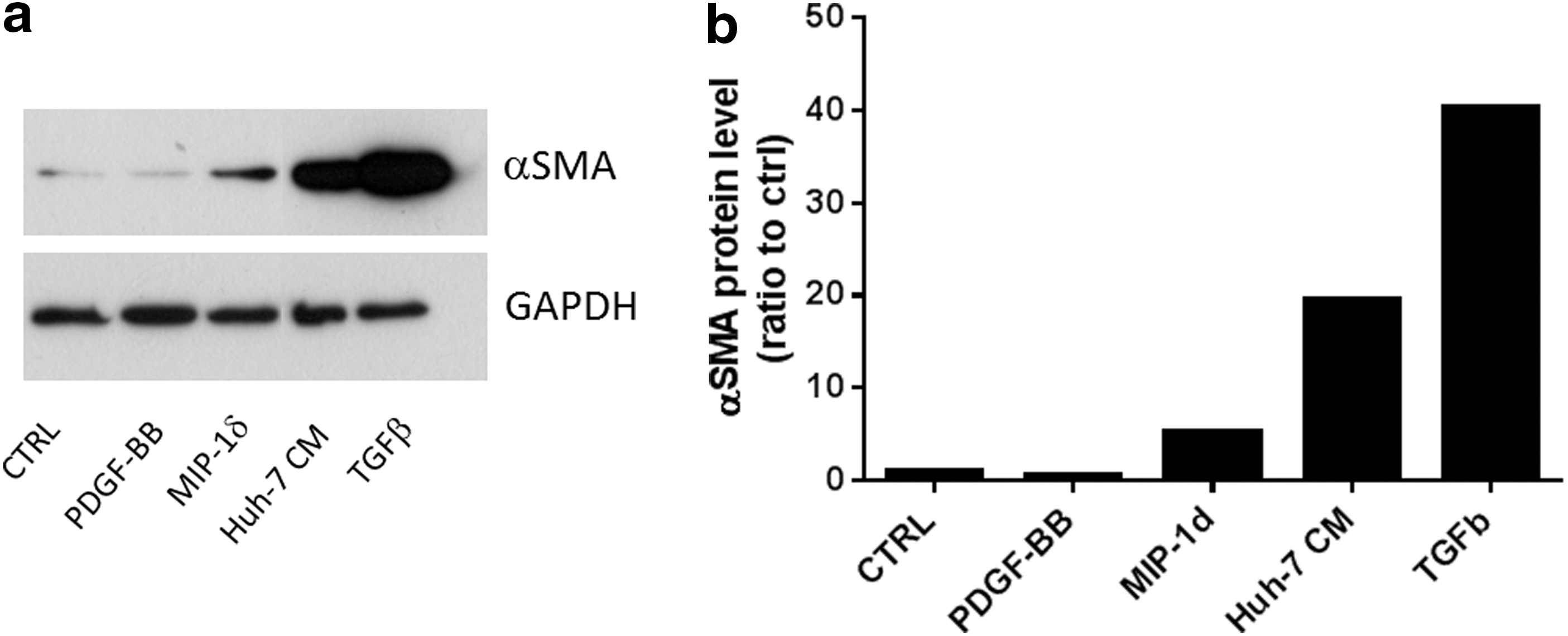
MIP-1δ induces α-SMA expression in MSC after 5 days of exposure. MSC were seeded at a density of 30,000 cells per well in a 24-well plate. Cells were incubated with control medium (CTRL), control medium containing PDGF-BB (25 ng/mL), MIP-1δ (25 ng/mL), or TGF-β (50 ng/mL) as well as with Huh-7 CM.
Discussion
MSC migration is controlled by common mechanisms identified in leukocytes, and it has been proposed that exploiting such cellular mechanisms may help improve cellular therapies [56]. However, the mechanisms involved in MSC and leukocytes migration might be different, since the expression of adhesion molecules such as P-selectins, necessary for tethering in leukocytes, was not unequivocally demonstrated in MSC [40,57]. Other studies showed the importance of CD44 in MSC migration [58]. Conversion of the native CD44 glycoform on MSC into hematopoietic cell E-selectin/L-selectin ligand increases the homing capacity of MSC into the bone marrow [59]. The involvement of chemokines, cytokines, and growth factors and their receptors in the attraction of MSC into wounds or tumors still need to be clarified both in vitro and in vivo.
Here, we investigated the implication of chemokines and factors in molecular trafficking mechanisms of MSC using a hepatoma cell conditioned medium that we found to induce MSC migration. In line with others [60], we observed that conditioned medium from the human cell line Huh-7 was able to induce efficient migration in MSC similar to PDGF-BB, whereas conditioned medium from freshly isolated human hepatocytes and another hepatoma cell line, HepG2, did not. Therefore, the effect observed with the Huh-7 CM is specific to this type of cell line and is probably related to their unique secretory profile that we have analyzed. We attempted to identify chemokines or growth factors involved in the activation of migration of MSC specific to Huh-7 CM. Using a chemokine antibody array, we detected increased levels of angiogenin, MIP-1δ (CCL15), MIP-3α (CCL20), and IGFBP-2 in the Huh-7 CM. Factors previously reported to induce MSC migration such as PDGF-BB [26,61], TGF-β [62], IL-1β [63], TNF-α [64], MCP-1 [28], and SDF-1 [33,65] were not increased in Huh-7 CM compared with control medium, excluding the possibility of an association between these factors and the induced migration in this setting. Neither angiogenin nor IGFBP-2 has an effect on MSC migration, which is not surprising since angiogenin is relevant for angiogenesis and more likely to be involved in inducing tumor vascularization [66] and IGFBP-2 was shown to act mainly on mitogenic and survival capacity of cells [67,68]. We focused on MIP-1δ and MIP-3α, both of which are cytokines regulating immune cell migration [69]. MIP-1δ stimulated chemotactic migration in primary endothelial cells. It was further proposed to play a role in angiogenesis [47] and recently considered a serum biomarker for hepatocellular carcinoma [70]. Using recombinant MIP-1δ and MIP-3α in the transwell migration assay, we showed that these soluble factors increased MSC migration and that specific blocking antibodies against MIP-1δ and MIP-3α slightly decreased MSC migration, with a significant result only with anti-MIP-1δ antibody.
Cell migration in response to chemokines has been shown to be modulated through the regulation of receptor expression. In neuronal stem cells, IL-10 or IL-4 up-regulated CXCR4 and CCR5 chemokine receptor expression, thereby increasing the migratory response to IL-4 and regulated on activation normal T cell expressed (RANTES) [71]. In human MSC, modulation of chemokine receptor expression was observed after exposure to several factors [56]. Treatments with cytokines TNF-α and IFN-β increased expression of chemokines receptors CXCR4 and CCR3, leading to increased migration in response to SDF-1 in vitro [72]. However, the chemokine receptor's expression profile in human MSC is broad and the expression of a chemokine receptor is not necessarily related to a function in migration [35]. Discrepancies exist between studies from different groups about the expression of CCR2 and its function [35,42]. CCR1 expression and its involvement in migration had also been reported [73]; however, others did not detect CCR1 [40].
We analyzed several receptors, such as CXCR2, CCR2, and CCR5, which were not related to the factors that we found to be increased in the Huh-7 CM. CXCR2 and CCR2 are receptors for IL-8 and MCP-1, respectively. Both were shown to be involved in MSC migration [28,33]. In our experiment, both receptors were considered absent. Others have demonstrated the expression of CCR2 and CXCR2 by FACS analysis in bone marrow-derived MSC [64,74]. These discrepancies might be related to different time periods of expansion of MSC in various media, which may select subpopulations of MSC with different receptor expressions. Such subpopulations might also further reflect the broad range of functions that MSC are expected to fulfill. Further, we detected low but consistent expression of CCR5 and CCR1 and highly variable expression of CCR6. Interestingly, the observation that the expression of receptors remained unchanged on treatment with Huh-7 CM and PDGF-BB suggests that migration of MSC was independent of mechanisms regulating their expression. As mentioned, the expression of CCR6 was highly variable in MSC populations from different donors, suggesting a regulation of this receptor independently of migration. Further, in our experiments, expression of the receptor for IL-8, CXCR2, which was previously shown to be involved in MSC migration [33], was highly variable, although not significantly increased in three independent experiments. This result leads us to hypothesize that Huh-7-induced migration of MSC might not be mainly regulated by receptor expression. However, we only analyzed receptor expression at 24 h after treatment, which might be too short to observe long-term regulatory mechanisms.
Cell migration is a complex phenomenon, also implicating dynamic modifications of the extracellular matrix through the release of metalloproteinases [75]. In MSC, previous studies revealed that MMP-1 was implicated in MSC migration to gliomas and induced by glioma conditioned medium [27], and that the migration/invasion capacity of hMSCs via MMP-1 was regulated by the Wnt signaling pathway [39]. Exposure of MSC to factors such as IL-1β or TGF-β increased the production of MMPs [52]. In this study, we also observed an up-regulation of MMP-1 by PDGF-BB and, to a higher extent, by the Huh-7 CM. This up-regulation was specific for MMP-1 since the expression of MMP-2 remained unchanged. Whether the identified inflammatory chemokines are involved in the up-regulation of MMP-1 remains to be investigated.
Several studies have shown that MSC migration is regulated through the activation of the MAPK/ERK signaling pathway [49,76]. We found that Huh-7 CM activated the ERK pathway similarly to PDGF-BB. However, PDGF-BB-induced migration was significantly impaired after inhibition of ERK phosphorylation by MEK inhibitor PD98059, whereas Huh-7 CM, MIP-1δ, and MIP-3α-induced migration was not, suggesting that other pathways are implicated under these conditions. In MSC, activation of several other pathways had been demonstrated. In human umbilical cord blood-derived MSC, Akt, ERK, and p38 signaling pathways were shown to be activated on SDF-1-induced migration [76]. Sphingosine-1-phosphate-induced MSC mobilization and migration requires cooperation of MMPs with the RhoA/Rho kinase and MAP/ERK signaling pathways [38]. Interestingly, MSC migration induced by TNF-α increased expression of p-ERK and p38; however, only the inhibitor SB203580 and not PD98059 suppressed migration of MSC [77]. Here, the induction of MSC migration by MIP-1δ and MIP-3α may activate several signaling pathways, also including the cooperation of MMP-1.
We further observed that αSMA expression was increased after prolonged culture of MSC with Huh-7 CM and MIP-1δ. At first sight, this observation appears as counterintuitive, since increased αSMA expression is accompanied with increased adhesion strength and contractibility of the cell [78], which lead to reduced cell migration [79]. However, we observed αSMA expression after 5 days, suggesting that migration and αSMA expression are sequential events where MSC migrate first and then start expressing αSMA. In tumors, tumor-derived factors not only activate local fibroblasts but may also attract circulating precursor cells, contributing to a tumor permissive microenvironment [80,81]. It has been proposed that MSC are a source of CAFs, which contain αSMA-positive contractile stromal cells with increased secretion of extracellular matrix and increased expression of SDF-1 [82 –84]. Interestingly, MSC exposed to conditioned medium of human breast cancer cells adopted a CAF-like phenotype [83]. MSC showed increased migration when exposed to the MDAMB231-conditioned medium without previous exposure of MSC, whereas MSC that had been exposed for several days to tumor conditioned medium migrated less, suggesting that adopting a CAF-like phenotype decreases migration. It will be of interest to further analyze whether MSC under long-term incubation with Huh-7 CM or MIP-1δ adopt a CAF phenotype.
In conclusion, we identified two chemokines, MIP-1δ and MIP-3α in the Huh-7 CM that are implicated in the Huh-7 CM-induced MSC migration; one of them, MIP-1δ, is involved in the induction of αSMA expression. Both MIP-1δ and MIP-3α activated the ERK pathway, but inhibition of this pathway in MSC did not significantly impair migration, suggesting the activation of other pathways. Activation of MSC migration was independent of regulation of receptors expression. Further, prolonged exposure to MIP-1δ induced a myofibroblast phenotype in MSC, characterized by increased levels of αSMA. This might be relevant for clinical application of MSC, since MSC are recruited to sites of injury or tumors producing chemokines and factors. MSC might be susceptible to undergo phenotypic changes once exposed to micro-environmental conditions. Characterizing chemokine receptors in MSC as well as analyzing inflammatory chemokines expressed by tumors might be important for patient-tailored therapies.
Footnotes
Acknowledgments
This study was funded by a research grant no. 05-1-II from the Research and Development Budget of the University Hospitals of Geneva, Switzerland to C.G.G. The authors gratefully acknowledge Corinne Sinigaglia, Nadine Pernin, and David Matthey-Doret for their technical help and Sergei Startchik and Olivier Brun (Bioimaging Core Facility, Faculty of Medicine, Geneva, Switzerland) for their help in using microscope and Metamorph settings for the quantification of cell migration.
Author Disclosure Statement
The authors disclose that no competing financial interests exist.