
Abstract
Dysregulation of forkhead box protein A2 (Foxa2) expression in fetal ventral mesencephalon (VM)–derived neural precursor cells (NPCs) appears to be associated with the loss of their potential to differentiate into dopaminergic (DA) neurons after mitogenic expansion in vitro, hindering their efficient use as a transplantable cell source. Here, we report that epigenetic activation of Foxa2 in VM-derived NPCs by inducing histone hyperacetylation rescues the mitogenic-expansion-dependent decrease of differentiation potential to DA neurons. The silencing of Foxa2 gene expression after expansion is accompanied by repressive histone modifications, including hypoacetylation of histone H3 and H4 and trimethylation of H3K27 on the Foxa2 promoter, as well as on the global level. In addition, histone deacetylase 7 (HDAC7) is highly expressed during differentiation and recruited to the Foxa2 promoter. Induction of histone acetylation in VM-derived NPCs by either knockdown of HDAC7 or treatment with the HDAC inhibitor apicidin upregulates Foxa2 expression via hyperacetylation of H3 and a decrease in H3K27 trimethylation on the promoter regions, leading to the expression of DA neuron developmental genes and enhanced differentiation of DA neurons. These effects are antagonized by the expression of shRNAs specific for Foxa2 but enhanced by shRNA for HDAC7. Collectively, these findings indicate that loss of differentiation potential of expanded VM-derived NPCs is attributed to a decrease in Foxa2 expression and suggest that activation of the endogenous Foxa2 gene by epigenetic regulation might be an approach to enhance the generation of DA neurons.
Introduction
N
To resolve this issue, current efforts are underway to identify novel DA neuron development factors, particularly those involved in DA neuron survival and development. Multiple factors are involved in DA neuron differentiation from VM-derived NPCs. Among the factors, forkhead box protein A2 (Foxa2) is the first transcription factor expressed in the ventral midline of both the embryo neural tube and DA progenitors. Foxa2 is expressed in floor plate progenitors as early as embryonic day 8.0 (E8.0), making it a likely candidate factor that contributes to the specification of DA progenitor identity. Mice lacking Foxa2 fail to express DA-neuron-developmental genes and exhibit a reduction in DA neurons [9 –11]. In contrast, overexpression of Foxa2 results in rapid cell cycle arrest and maturation of NPCs toward DA neuronal phenotype [12]. Intriguingly, the transcription profile of NPCs has demonstrated that Foxa2 is downregulated in expansion conditions in vitro [1]. These studies indicate that the silencing of Foxa2 gene expression could impact DA neuron differentiation. However, the precise mechanism of the regulation of Foxa2 expression during DA neuron differentiation is unknown.
Maintenance of the NPC state and its neural differentiation are the result of transcriptional control in concert with chromatin remodeling and epigenetic modifications. NPC fate is strictly regulated by regional and temporal mechanisms involving precise epigenetic control [12]. Mouse models that are deficient in several histone acetylase genes, including CBP, p300, and GCN5, exhibit severe neural tube closure defects [12 –15]. Further, histone deacetylase (HDAC)-1, -3, -5, and -7 are highly expressed in NPCs, and HDAC-mediated transcriptional repression is required for the proliferation and differentiation of NPCs [16,17]. In contrast, HDAC2 and HDAC11 are preponderantly expressed in mature neurons [16]. Although these studies suggest that regulation of histone acetylation is important for lineage specification in neuronal cells, little is known about the role of epigenetic modifications in transcriptional control of DA-neuron-inducing genes. To address this, we passaged VM-derived NPCs and analyzed changes in epigenetic modifications on the Foxa2 promoter as well as at the global level during differentiation. Here, we report that active histone modifications on the Foxa2 promoter in VM-derived NPCs after expansion switched to repressive histone modifications during differentiation, leading to a decrease in differentiation potential. Induction of hyperacetylation by either the histone deacetylase inhibitor (HDACI) apicidin or knockdown of HDAC7 resulted in maintenance of differentiation potential through restoration of Foxa2 gene expression via active histone modifications.
Materials and Methods
Preparation of VM-derived NPCs
Ventral mesencephalons (VMs) were dissected from E12 mice (ICR strain) (19). ICR mice were purchased from Daehan Biolink. VMs were dissected in Ca2+- and Mg2+-free Hank's balanced salt solution (Corning). After dissection, VMs were placed in N2-serum-free medium and mechanically dissociated in 1 mL of N2 medium. The tissues were settled in a 37°C incubator for 5 min. Cells in the supernatant were collected and seeded at 2×104 cells on 10-cm culture dishes precoated with poly-
Virus infection
The lentiviral shRNA sets for Foxa2 were purchased from Open Biosystems. The lentivirus was harvested from 293FT cells according to the manufacturer's protocol. Passaged VM-derived NPCs (5×105 cells) were infected with lentivirus containing the Foxa2 shRNA for 24–48 h. The infected cells were used for experiments. The shRNA sets are as follows: Foxa2, 5′-CGCCACCGCATATTCGC-3′ and 5′-CCGCCATACCGATT-3′; HDAC7, 5′-CCGGTGCAGATCATTCTACAGCCATCTCGAGATGGCTGTAGAATGATCTGCATTTTTG-3′ and 5′-TTCGGCAACTTCTCAATAAACTCGAGTTTATTGAGAAGTTGCCGAAGTTTTTG-3′.
Immunocytochemistry
VM-derived NPCs were fixed in 4% paraformaldehyde and rinsed three times with cold phosphate buffered saline (PBS). The cells were incubated in 0.1% Triton-X100 in PBS for 15 min and rinsed three times with cold PBS. These cells were then blocked in 0.1% bovine serum albumin/10% goat serum in PBS for 1 h at room temperature and incubated with primary antibodies overnight at 4°C. For detecting endogenous-specific-protein-expressing cells, cells were treated with sodium dodecyl sulfate (1% in PBS) at room temperature for 5 min before the blocking procedure. The primary antibodies were detected using secondary antibodies conjugated to Alexa Flour 594 or 488 (Invitrogen). Cell nuclei were stained with 4′,6-diamidino-2-phenylindole (Sigma) and mounted using mounting solution. Confocal images were obtained using an Olympus FV-1000 confocal laser scanning microscope with an Apochromat 60× objective.
Cell counting and statistical analysis
Cell density of DA neurons was determined by counting the numbers of tyrosine hydroxylase (TH)–positive cells and the numbers of β-tubulin-positive cells per field at 40× magnification using a Zeiss Axioplan I fluorescent microscope (Carl Zeiss). Ten fields were randomly selected and counted for each sample, and cell densities were calculated by dividing the numbers of TH-positive cells by that of β-tubulin-positive cells. Numbers presented in figures represent the average percentage and standard error of the mean (SEM) of TH-positive cells over β-tubulin-positive cells from three samples per VM-derived NPCs. For statistical analysis, we performed analysis of variance (ANOVA) with an α level of 0.01 to determine possible statistical differences between group means.
Immunoblot analysis
For immunobloting, VM-derived NPCs were lysed with ice-cold lysis buffer [20 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (pH 7.5), 150 mM sodium chloride, 1% Nonidet P-40, 10% glycerol, 60 mM octyl β-glucoside, 10 mM sodium fluoride, 1 mM sodium orthovanadate, 1 mM phenyl methylsulfonyl fluoride, 2.5 mM nitrophenyl phosphate, 0.7 μg/mL pepstatin, and protease inhibitor cocktail tablet]. The lysates were kept on ice for 30 min and then centrifuged at 13,000 rpm for 15 min at 4°C. Samples were resolved by electrophoresis on 4%–12% sodium dodecyl sulfate polyacrylamide gel electrophoresis Bis–Tris gradient gels, and the proteins were transferred to polyvinylidene difluoride membranes using semidry transfer (Bio-Rad). The membranes were incubated overnight with the indicated primary antibodies. The membranes were incubated for 1 h at room temperature with a horseradish-peroxidase-coupled secondary antibody (Abcam or Millipore). Signals from immunoreactive proteins were detected using a Davinch-chemi imager v 0.0.3.5 CAS-400SM system. Additional details about antibodies are provided in Supplementary Materials and Methods (Supplementary Data are available online at
Real-time polymerase chain reaction analyses
RNA samples were obtained using Trizol reagent (Invitrogen). Reverse transcription (RT) was accompanied by optimal magnesium chloride (MgCl2), DNA polymerase concentration, and annealing temperature. The reaction was performed for 1 h at 42°C and terminated by incubating for 15 min at 70°C. Real-time polymerase chain reaction (RT-PCR) was performed on a C 1000™ thermal cycler (Bio-Rad) using SYBR fast qPCR mixture kit (Kapa Biosystems). The reaction mixture (20 μL) contained 1 ng/μL DNA template, 200 nM forward and reverse primers, 2× SYBR green 1 dye, 200 nM ROX reference dye, DNA polymerase, and 2.5 mM MgCl2. GAPDH mRNA was used as a normalization control. The primer sequences are described in Supplementary Materials and Methods.
Chromatin immunoprecipitation assay
VM-derived NPCs (8×105) were treated with 500 nM apicidin or dimethyl sulfoxide (DMSO) for 24 h and differentiated for 7 days. The chromatin immunoprecipitation (ChIP) assay was performed according to the manufacturer's protocol (Upstate Biotechnology). A small portion of the crosslinked, sheared chromatin solution was reserved as the input DNA, and the remainder was subjected to immunoprecipitation using histone modification antibodies. The immunoprecipitated DNA was purified using a QIA quick PCR purification kit (Qiagen) and analyzed by quantitative real-time PCR to identify histone modification patterns on the promoter of Foxa2 genes.
Dopamine release assay
Dopamine release was quantified by reverse-phase high-performance liquid chromatography (HPLC) using the culture supernatants of VM-derived NPCs. Passaged VM-derived NPCs were treated with apicidin (500 nM) or DMSO for 24 h and differentiated for 7 days in 24-well plates with 500 μL of N2 medium+ascorbic acid for 24 h or 15 min of in the same medium supplements with 56 mM potassium chloride (KCl) for 15 min. The medium was collected from the plates and immediately treated with 100 μL trichloroacetic acid and 100 pmol isoproterenol (internal standard). The buffer was centrifuged at 13,000 rpm for 5 min at 4°C. The supernatants were gathered and stabilized in 0.1 mM ethylenediaminetetraacetic acid (EDTA). The final reaction mixture (50 μL) was injected into an HPLC system. DA levels were calculated using a standard (1–50 nM dopamine; Sigma) injected immediately before and after each experiment. The HPLC column used for dopamine detection was the ESA HR-80 column (3 μm, octadecyl silica gel, 80×4.6 mm). The speed of mobile phase (7.5 mM sodium phosphate monobasic, 1.7 mM 1-octanesulfonic acid, 0.05% triethylamine, 0.1 mM EDTA, and 10% acetonitrile, pH 3.0 with phosphoric acid) was 0.6 mL/min. For this experiment, the Shimadzu HPLC system was used.
Statistical analysis
The data presented are the means±SEMs from three or more independent experiments. Statistical analysis was performed by using one-way ANOVA and the Dunnett test. Statistical significance (*P<0.05 and **P<0.01) was determined with Sigma Stat software (Systat Software, Inc.).
Results
Expansion of VM-derived NPCs causes loss of differentiation potential to DA neurons, concomitant with suppression of Foxa2 expression
First, to investigate whether loss of differentiation potential by expansion is associated with alteration of Foxa2 expression pattern, we differentiated VM-derived NPCs after mitogenic expansion for up to 20 days to DA neurons and analyzed the expression level of neuronal markers. Both mRNA and protein expression levels of Foxa2 significantly decreased compared with unexpanded differentiation and showed an expansion-time-dependent decline (Fig. 1A–C and Supplementary Fig. S1A, C). Other neuronal factors (LIM homeobox transcription factor 1a/b/engrailed 1) essential for DA neurogenesis also showed an expression pattern similar to that of Foxa2 following expansion (Supplementary Fig. S3A–C). Consistent with the reduced expression of these neuronal factors, the portion of TH-positive neurons/total cells after expansion for 5 days decreased by almost half compared with unexpanded differentiated NPCs (Fig. 1D–F and Supplementary Fig. S1B, C). Also, the expression pattern of paired-like homeodomain transcription factor 3 (Pitx-3), another DA neuron marker, was similar to that of TH (Supplementary Fig. S2A–C). In contrast, glial fibrillary acidic protein (GFAP), an astrocyte marker, dramatically increased in expanded culture (Supplementary Fig. S2D–F). No drastic slowing of proliferation or obvious phenotypic change was observed after expansion for 10 or 20 days (data not shown). Taken together, these observations indicate that inefficient differentiation of expanded VM-derived NPCs into DA neurons may be attributed to the suppression of Foxa2 expression.
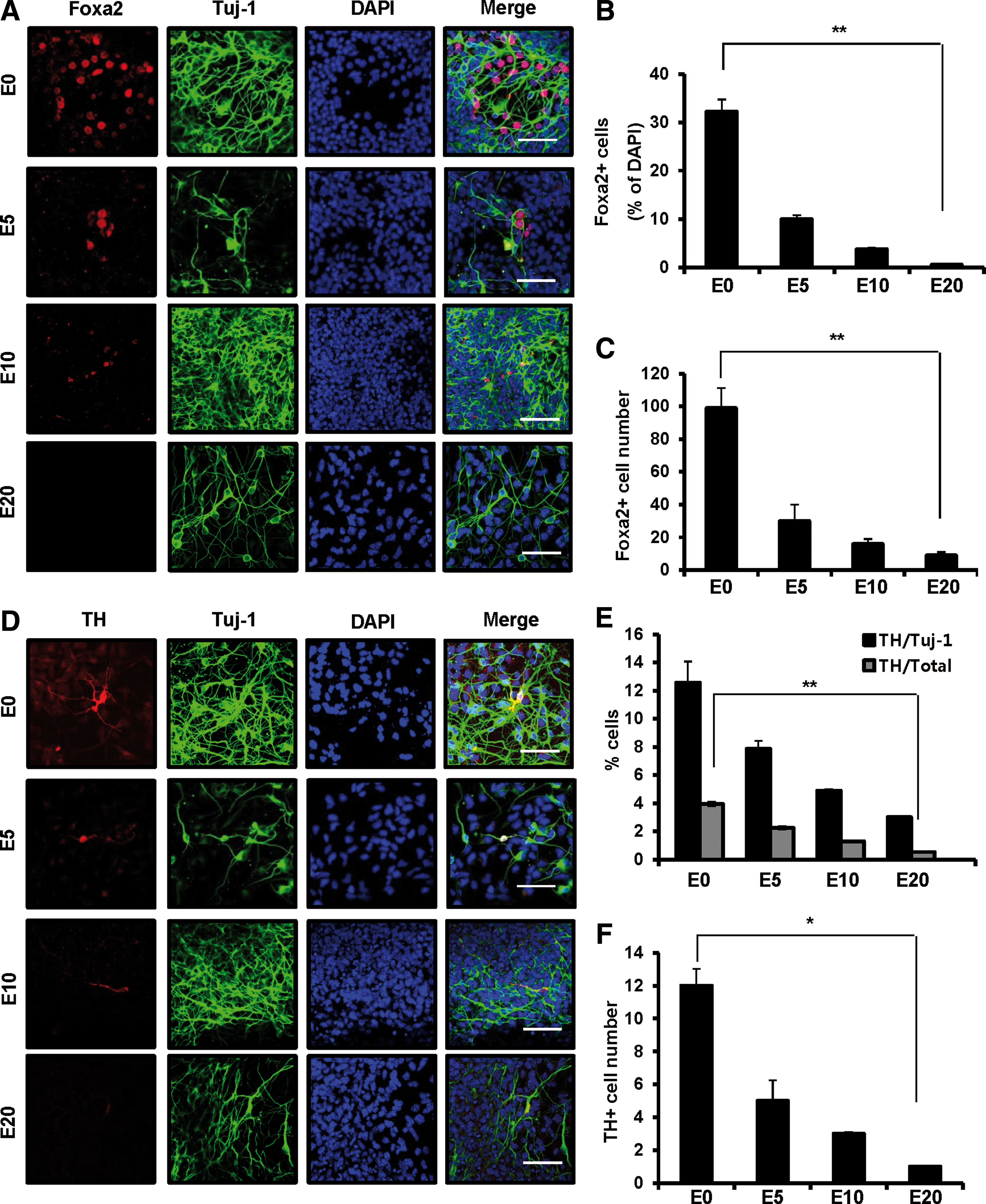
Developmental potency of VM-derived NPCs to DA neurons after mitogenic expansion. VM-derived NPCs were unexpanded (E0) or expanded in the presence of bFGF and bEGF in N2/B27 medium for up to 20 days before differentiation. Expanded cells were passaged and differentiated in N2/B27 medium without bFGF and EGF for 7 days.
Global and local histone modification patterns are altered during DA neuron differentiation after expansion
To examine the involvement of epigenetic alteration in loss of differentiation potential, we first measured the global histone modification patterns during differentiation of NPCs expanded for 5 days when significant loss of differentiation potential (about 50%) was observed (Fig. 1E). Interestingly, the level of H3K27me3, an indicator of gene repression, was globally increased, whereas histone acetylation, an indicator of active gene expression, was decreased. Other histone modifications were marginally changed (Fig. 2A–C). We next examined the nature of the histone modifications using a ChIP assay. In agreement with changes in global histone modifications, acetylation of histone H3, an activating histone modification, on the Foxa2 promoter dramatically decreased during DA neuron differentiation, while trimethylation of H3K27, a repressive histone modification, significantly increased (Fig. 2D). However, trimethylation of H3K9 on the promoter was not changed, although it was globally decreased (Fig. 2A, D). These data suggest that the decrease of Foxa2 expression during DA neuron differentiation is attributed to epigenetic silencing of the Foxa2 gene through repressive histone modifications, leading to the low efficiency of DA neuron differentiation in mitogenic expanded culture.
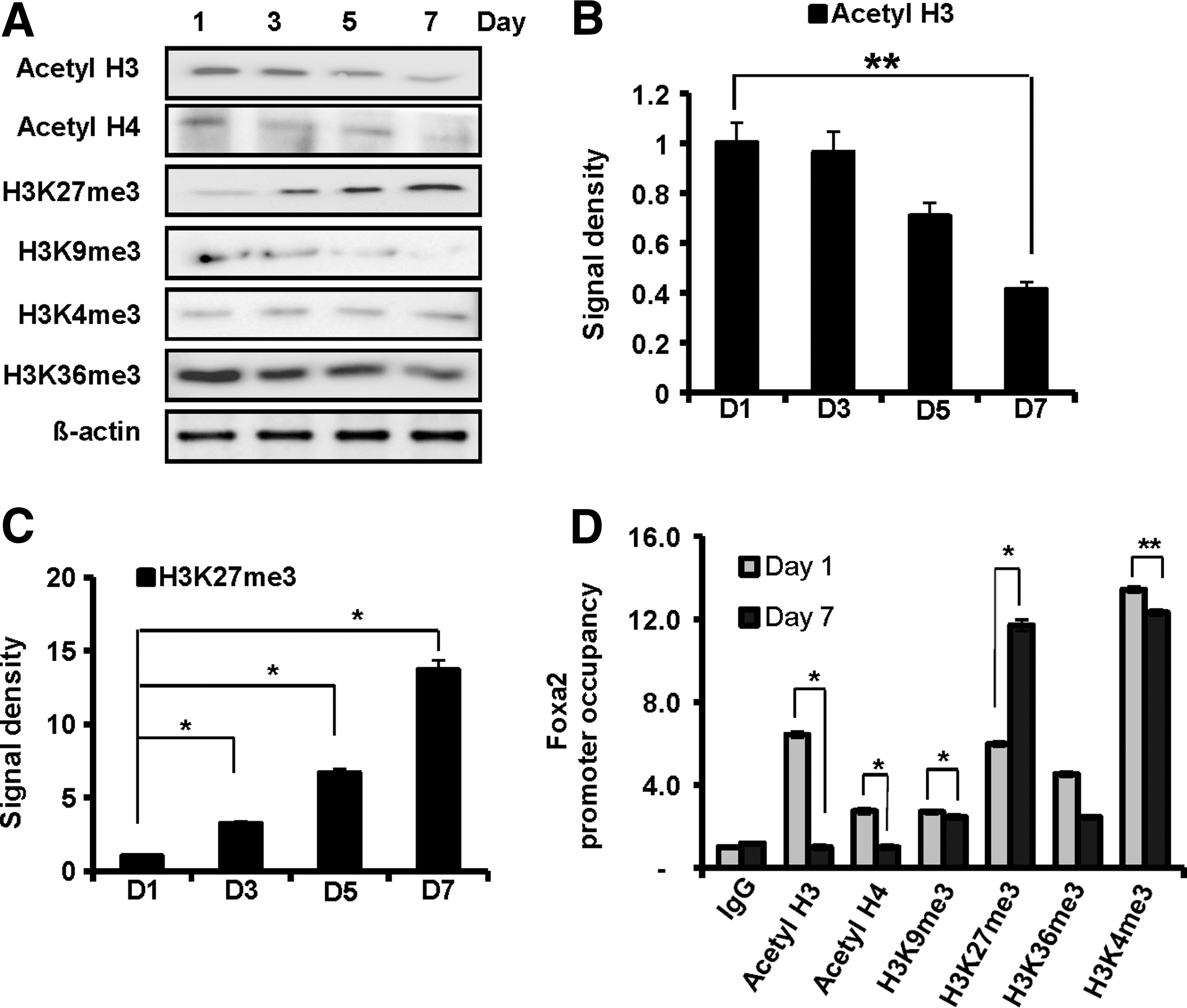
Alternation of histone modification patterns during DA neuron differentiation. VM-derived NPCs were expanded in vitro for 5 days and then differentiated for 7 days.
Apicidin increases Foxa2 expression by upregulating H3 acetylation
To test the effect of histone H3 acetylation on Foxa2 expression during DA neuron differentiation, we increased the level of H3 acetylation using apicidin, a specific cyclic tetrapeptide HDACI [18,19]. Acetyl-H3 and acetyl-H4 appeared to peak after 12–24 h of 500 nM apicidin treatment (Supplementary Fig. S4A). Apicidin treatment for 24 h on the first day of DA neuron differentiation increased global acetyl-H3 in a dose-dependent manner, while the levels of H3K36me3 and H3K4me3 remained unchanged (Fig. 3B and Supplementary Fig. S4B). In parallel, acetylated histone H3 was enriched on the Foxa2 promoter upon apicidin treatment, whereas H3K27me3 was depleted (Fig. 3G). Consistent with the alteration of histone modifications, the number of cells expressing Foxa2 (Fig. 1B and Fig. 3C, D; 9.98% vs. 33.21%) and the level of Foxa2 mRNA (Fig. 3E) were increased by apicidin treatment. Apicidin did not affect cell viability and the total number of Tuj-1-positive cells during mitogenic expanded DA neuron differentiation (Fig. 3F and Supplementary Fig. S4C, D). These data indicate that the induction of H3 acetylation is sufficient to initiate the expression of the Foxa2 gene, which is suppressed during DA neuron differentiation, suggesting that the downregulation of acetyl-H3 during DA neuron differentiation is associated with the reduction of Foxa2 expression.
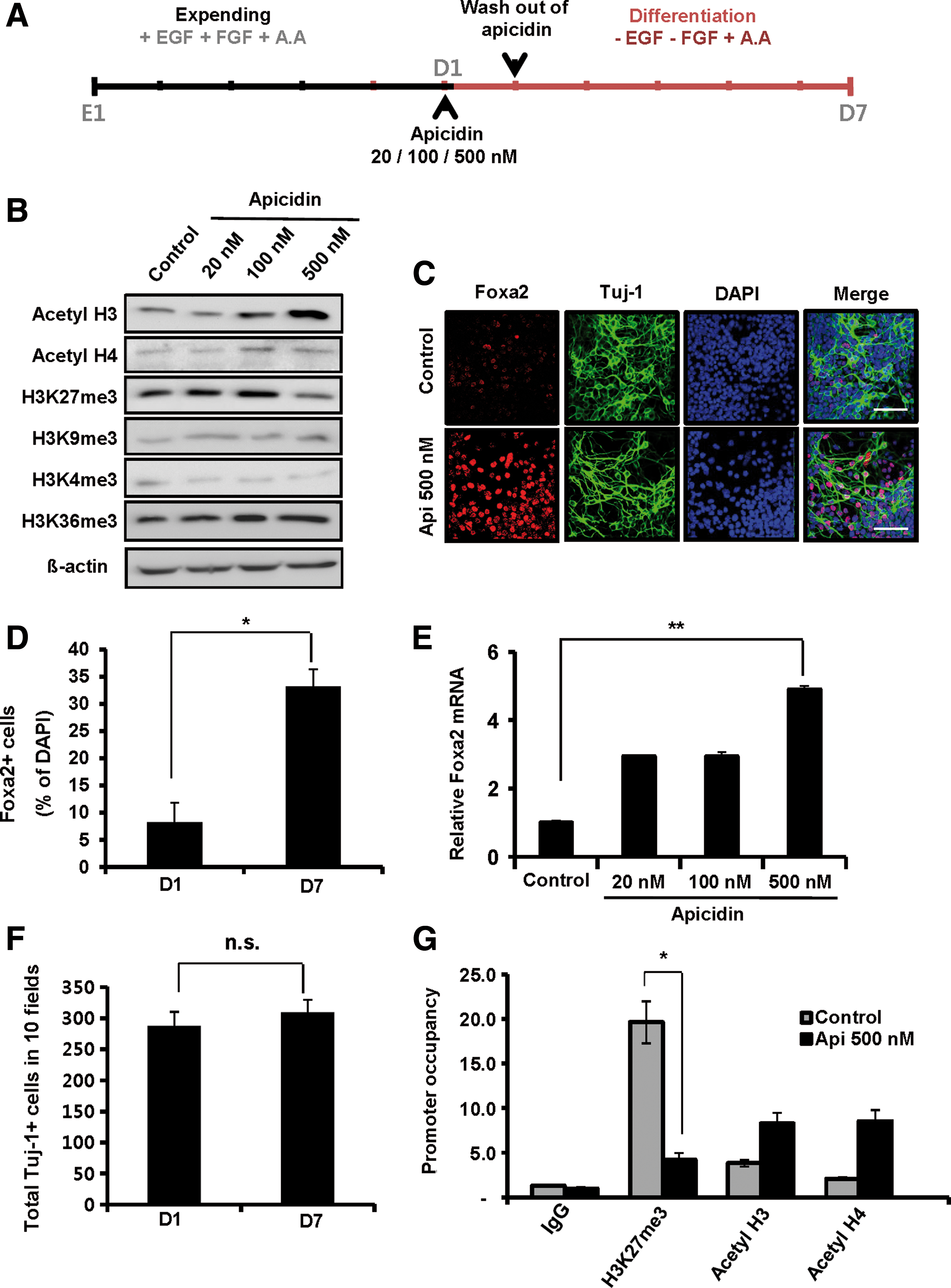
Apicidin increases Foxa2 expression by upregulating histone H3 acetylation. VM-derived NPCs from embryonic day 12 mice were expanded for 5 days before differentiation. After expansion, cells were plated on newly coated dishes and treated with different doses (20, 100, and 500 nM) of apicidin in N2/B27 medium for 24 h on the first day of DA neuron differentiation. Cell differentiation was induced for the subsequent 7 days.
Apicidin enhances differentiation of mitogenic expanded VM-derived NPCs into DA neurons
To examine the effect of apicidin on the efficiency of DA neuron differentiation, we added 500 nM apicidin for 24 h on the first day of expanded DA neuron differentiation and measured the expression of several DA neuronal markers during DA neuron differentiation. Apicidin treatment caused a dose-dependent increase in both nuclear receptor related 1 (Nurr1) and Pitx3 mRNAs (approximately 3.88-fold and 2.74-fold, respectively) (Supplementary Fig. S5A), and Pitx3 protein also increased by ∼2.3-fold (Fig. 4A, B). In addition, the portion of TH + cells/Tuj-1 + cells and TH + cells/total cells also increased by approximately 2.2-fold and 3.8-fold, respectively, upon apicidin treatment (Fig. 4C, D). Real-time PCR further confirmed the increase of TH mRNA after apicidin treatment (Supplementary Fig. S5B). Similarly, the mRNA level of dopamine transporter (DAT), a late DA marker, significantly increased in a dose-dependent manner (Supplementary Fig. S5B). However, apicidin did not affect the total number of Tuj-1-positive cells (Fig. 4E). In contrast, the expression of astrocyte makers (GFAP and glutamine synthetase) was reduced by apicidin treatment (Supplementary Fig. S5C–E), indicating that the effect of apicidin is specific to DA neuron differentiation.
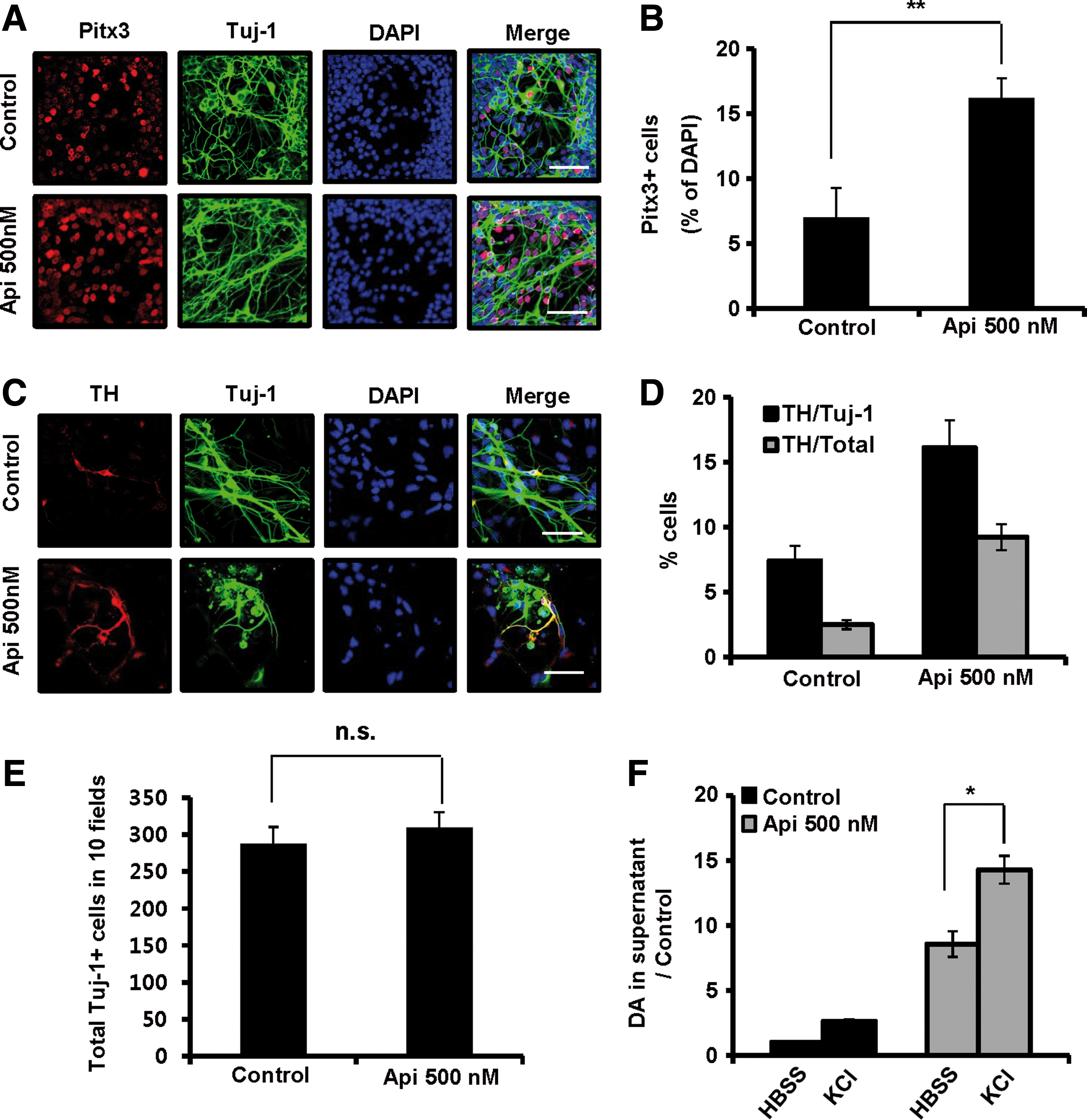
The yield of DA neuron differentiation is increased by apicidin treatment. Mitogenic expanded VM-derived NPCs from embryonic day 12 mice were treated with different doses (20, 100, and 500 nM) of apicidin in the absence of bFGF and EGF in N2/B27 medium for 24 h. Cells at the indicated times were then probed with various antibodies and stained with fluorescently labeled secondary antibodies.
To test whether apicidin-induced expression of specific DA markers leads to generation of more efficient functional DA neurons, we next examined the ability of the cells to release DA after membrane depolarization. In-vitro-differentiated cells from expanded NPCs in the presence or absence of apicidin were treated with 50 mM KCl, and the released DA in the media was analyzed by HPLC or enzyme-linked immunosorbent assay (ELISA). In response to membrane depolarization from expanded cells, apicidin treatment caused a fivefold increase of DA release compared with untreated control cells (Fig. 4F). These data suggest that apicidin-induced Foxa2 expression may maintain the differentiation potential of expanded and passaged VM-derived NPCs into DA neurons by specifically regulating the expression of DA neuronal factors.
HDAC7 is expressed in a time-dependent manner and recruited to the Foxa2 promoter during DA neuron differentiation
Our observations of the decreased levels of histone acetylation after expansion also suggest a possible role for HDACs during DA neuron differentiation. To assess this possibility, we first examined the expression patterns of HDACs during DA neuron differentiation after expansion by immunocytochemistry, western blot, and real-time polymerase chain reaction. The expression levels of HDAC1, HDAC2, HDAC3, and HDAC8, which are class I HDACs, showed no significant changes during expansion and differentiation (Fig. 5A and Supplementary Figs S6A–D and S7A–D). Like class I HDACs, the expression levels of HDAC4 and HDAC5 (class IIa HDACs) and HDAC6 (a class IIb HDAC) appeared to change only slightly (Fig. 5A, and Supplementary Figs S6E, F, and H and S7E–G). However, among the class IIa HDACs, mRNA and protein expression levels of HDAC7 were significantly increased in a time-dependent manner during expansion and differentiation (Fig. 5A, B and Supplementary Fig. S6G). To further test the involvement of HDAC7 in Foxa2 gene expression, we analyzed the pattern of HDAC7 recruitment to the promoter of the Foxa2 gene during DA neuron differentiation. In agreement with upregulation of HDAC7 expression, HDAC7 and its enzyme activity enhancer (HDAC2 and HDAC3) were significantly enriched at the Foxa2 promoter (Fig. 5C).
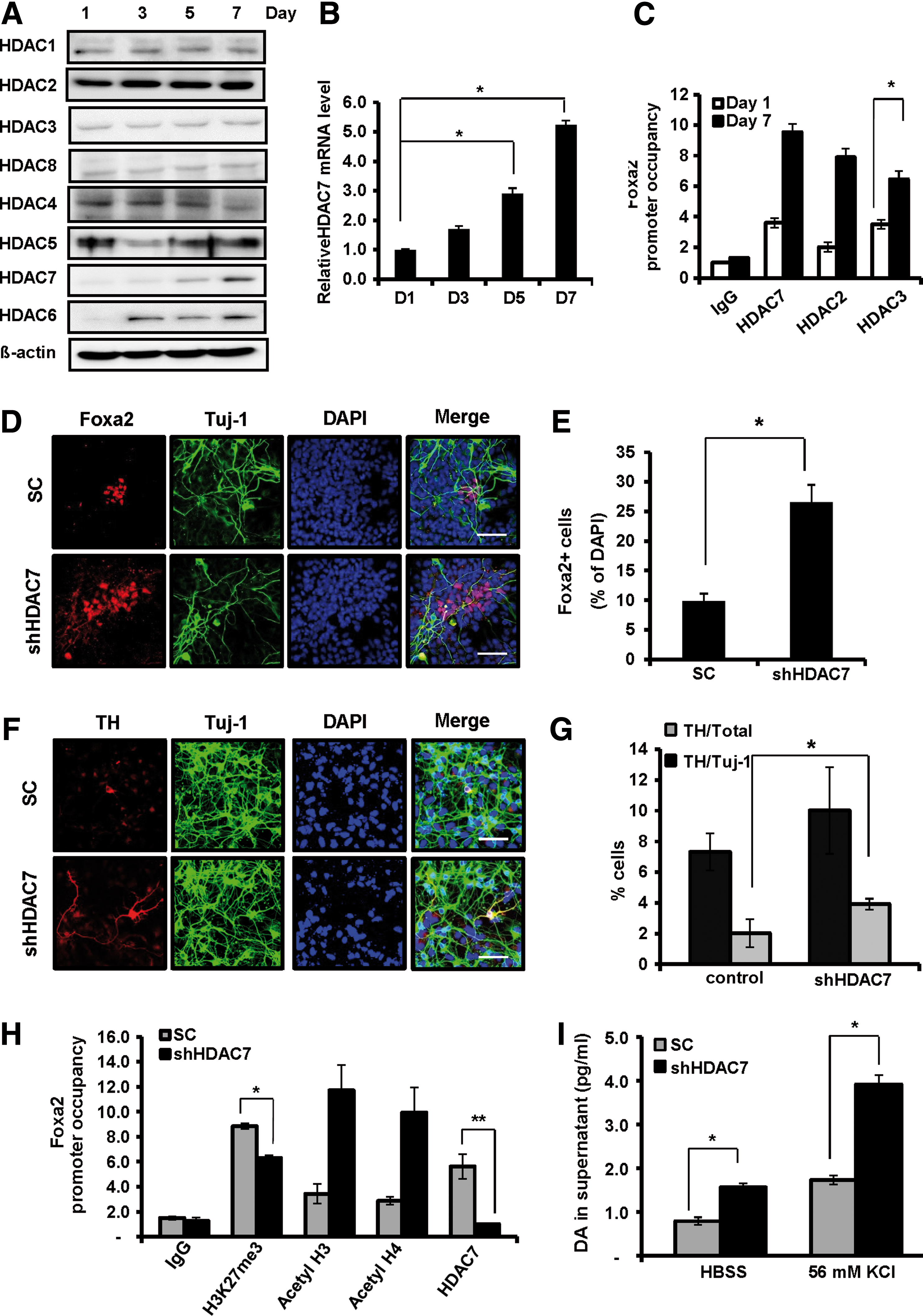
HDAC7 is highly associated with expanded VM-derived NPC differentiation due to its regulation of Foxa2 expression. VM-derived NPCs were expanded in vitro for 5 days and then differentiated for 7 days. HDAC protein was prepared from the indicated differentiation time period.
To test whether the reduced expression of Foxa2 was mediated by HDAC7 during DA neuron differentiation, we examined the expression of Foxa2 after knockdown of HDAC7 using HDAC7-shRNA-inserted lentivirus. First, three sets of shRNAs targeting different regions of HDAC7 mRNA were transiently expressed in NPCs, and their efficiency was examined (Supplementary Fig. S8A, B). Following these tests, we selected group 2 shRNA for HDAC7 deletion (Fig. 5D–I). HDAC7 depletion increased the expression of Foxa2 compared with the control group during DA neuron differentiation (2.5-fold) (Fig. 5D, E). Next, we investigated whether decreased expression of HDAC7 affects DA neural factors during DA differentiation. Knockdown of HDAC7 markedly increased the expression of TH and Pitx3 (Fig. 5F, G and Supplementary Fig. S8C, D) but did not affect the total number of Tuj-1-positive cells during DA neuron differentiation (Supplementary Fig. S8E). Real-time PCR further confirmed the increase of mRNA expression of DA neuronal markers after depletion of HDAC7 (Fig. 6D). These results indicate that depletion of HDAC7 is sufficient to rescue the differentiation potential of expanded VM-derived NPCs to DA neurons comparable to that of the unexpanded NPCs (Fig. 1C, D). In addition, depletion of HDAC7 caused increased occupancy of acetylated histone H3 and H4 at the Foxa2 promoter (Fig. 5H) as well as the global acetylation (Supplementary Fig. S9A–C). In good agreement with this result, cells depleted of HDAC7 showed more efficient functional DA neurons, as determined by DA detection analysis using ELISA (Fig. 5I). Taken together, these data indicate that among the HDACs we tested, HDAC7 showed not only the most dramatic increase in expression but also a high occupancy on the Foxa2 gene promoter during differentiation, suggesting that HDAC7 may play a critical role in DA neuron differentiation.

Apicidin influences HDAC7 expression and its enzymatic activity in the expanded VM-derived NPC differentiation.
To test whether HDAC7 is affected by apicidin, we first analyzed both the mRNA and protein expression levels of HDAC7. Apicidin treatment suppressed the expression of HDAC7 in a dose-dependent manner (Fig. 6A, B) as well as the enrichment of HDAC7, HDAC2, and HDAC3 on the Foxa2 promoter (Fig. 6C). In contrast, Foxa2 expression as well as DA neuronal markers was increased by apicidin (Fig. 6D), indicating that the effect of apicidin on Foxa2 expression might be mediated by the downregulation of HDAC7 expression.
Apicidin treatment induces DA neuron differentiation in a Foxa2-dependent manner
To test whether the effects of apicidin on mitogenic expanded DA neuron differentiation were mediated by Foxa2 expression, we examined the expression levels of DA neural factors after knockdown of Foxa2 using Foxa2-shRNA-inserted lentivirus (Supplementary Fig. S10A–C). Foxa2 depletion decreased the expression of Nurr1 compared with the control group during mitogenic expanded DA neuron differentiation (Fig. 7F). Moreover, Foxa2 depletion suppressed the apicidin-induced upregulation of Nurr1 (Fig. 7F), indicating that the effect of apicidin on Nurr1 relies on Foxa2 expression. The expression level of Foxa2 was not recovered by apicidin treatment after Foxa2 knockdown (Fig. 7A, C). Consistent with these results, apicidin treatment did not upregulate the expression of TH, DAT, or Pitx3 in cells lacking Foxa2 (Fig. 7B, D–F and Supplementary Fig. S10D). These results indicate that the effect of apicidin might be mediated by restoring the expression of Foxa2.
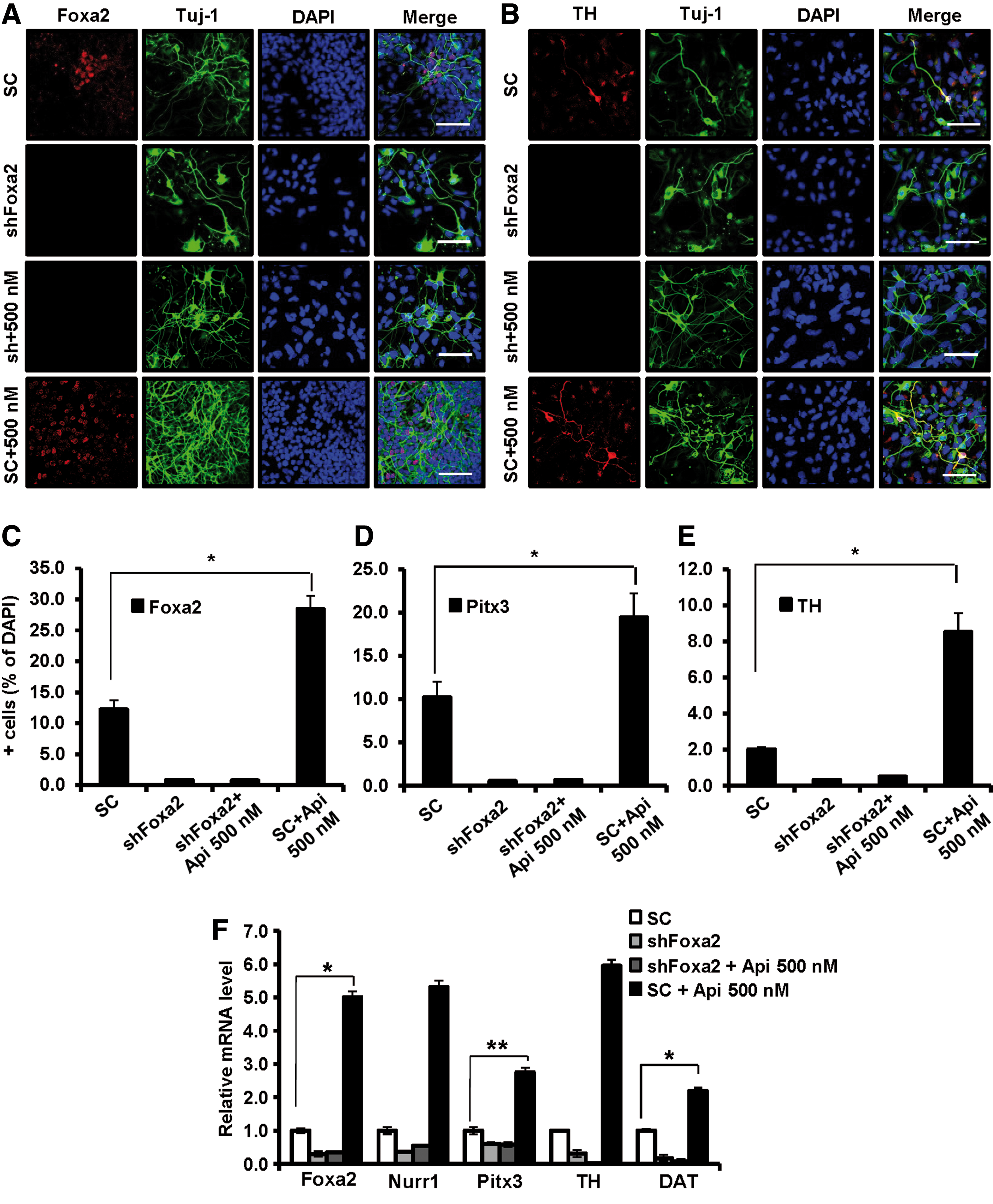
DA neuron differentiation by apicidin is dependent on Foxa2 expression. Mitogenic expanded VM-derived NPCs from embryonic day 12 mice were infected with lentivirus-expressing shRNA against Foxa2 for 48–72 h. One day after transduction, infected cells were treated with 500 nM apicidin. The cells were differentiated for 5 days.
Discussion
PD is characterized by the loss of DA neurons, making it the most likely of neurodegenerative diseases to be treated by cell replacement therapy [20]. Two representative cell sources are considered potential donor cells for PD. The first source is embryonic stem cells (ESCs) from the inner cell mass of the blastocyst. The second candidate source is neural stem cells (NSCs) derived from fetal or adult brain. NSCs are multipotent cells derived from neural tissues and can differentiate into various neural cell lineages with appropriate extrinsic signal [21,22]. NSCs derived from the ventral mesencephalon can differentiate to DA neurons. The advantages of NSCs over ESCs are a lower potential to form tumors after transplantation and greater genomic stability than ESCs, which show karyotypic changes [23 –25]. These advantages make VM-derived NPCs the strongest candidate for cell replacement in PD. To obtain a sufficient number of DA precursors and neurons for cell replacement treatment, cell expansion is an absolute necessity. However, VM-derived NPCs usually show signs of senescence after mitotic expansion and repeated passaging [26], hindering the production of DA neurons from passaged VM-derived NSCs. Although expanded and passaged VM-derived NSCs can be genetically modified to express transcription factors involved in DA neurogenesis, the total number of TH-positive cells is <2% [26]. Despite repeated attempts by many research groups, the production of DA neurons from mouse VM-derived NPCs has failed, even from short-term-expanded NPCs. In spite of the successful production of DA neurons from short-term-expanded mitotic neural precursors, researchers have failed to generate DA neurons from NPCs with self-renewal and multipotency.
To clarify this issue, the present study demonstrates that DA neuron developmental genes are continuously reduced during differentiation of VM-derived NPCs into DA neurons, whereas astrocyte marker genes are increased. Interestingly, we found that Foxa2 was downregulated in passaged VM-derived NPCs compared with unexpanded NPCs. Thus, recovery of silenced Foxa2 expression appears to be necessary to generate DA neurons from expanded mouse VM-derived NPCs. In line with the previous observations that show that Foxa2 is sufficient to induce and required for maintaining the expression of other neuronal factors, such as Lmx1a/b, in DA progenitors [27], the suppression of Foxa2 expression during expansion was accompanied by the reduction of Lmx1a/b and EN-1 (Supplementary Fig. S3A–C). These results indicate that Foxa2 acts in concert with other neuronal factors, including Lmx1a/b and EN-1, to drive a complex pattern of gene expression leading to DA neurogenesis.
Previous studies showed that lowered oxygen conditions enhance proliferation, survival, and DA differentiation of NPCs [6,30]. In addition, overexpression of Foxa2 in human embryonic stem cell-NPCs cultures increased cell survival following exposure to the toxins hydrogen peroxide and MPP+ [13], and Foxa2 is still expressed in DA neurons until adult stage, indicating that Foxa2 might have important roles in adult DA neurons, such as protecting against oxidative stress [28,29]. Thus, we could not rule out the possibility that the loss of differentiation potency of NPCs toward DA neurons in normal oxygen conditions is, in part, due to oxidative stress, which might be induced by the silencing of Foxa2 gene expression. Although the Foxa homolog in Caenorhabditis elegans is reported to induce antioxidant enzymes and increase life span in response to diet restriction [31], there is no evidence that Foxa2 directly regulates the induction of antioxidant enzymes in NPCs during expansion and differentiation. Therefore, the elucidation of antioxidant genes as Foxa2 target gene during expansion and differentiation of NPCs will be an important future study, which may provide us with important insight into the molecular mechanisms by which Foxa2 rescues the mitogenic-expansion-dependent decrease of differentiation potential of NPCs to DA neurons.
It is still unclear what factor controls the Foxa2 gene in DA neurons. In this study, we analyzed the Foxa2 promoter region to identify epigenetic regulators of Foxa2 expression through a ChIP assay and found that various histone modifications occurred at the Foxa2 promoter. Interestingly, HDAC7 was gradually upregulated and the inhibition of Foxa2 expression was transcriptionally mediated by direct association of HDAC7 with the Foxa2 promoter during DA neuron differentiation. In contrast with class I HDACs (HDAC1, -2, -3, and -8), class IIa (HDAC4, -5, -7, and -9) show weak enzymatic activity by themselves, but through recruitment of class I HDACs and interaction with the SMRT/NCoR corepressor complex carry out deacetylase function [32,34]. Consistent with this evidence, we showed that HDAC7 was recruited on the Foxa2 promoter together with HDAC2 and HDAC3. In line with this finding, histone H3 and H4 acetylation on the Foxa2 promoter was decreased significantly during expanded and passaged VM-derived NPC differentiation into DA neurons, suggesting that Foxa2 expression is directly upregulated by histone H3 and H4 acetylation. These results indicate that HDAC7 recruitment causes histone hypoacetylation on Foxa2 promoter, leading to downregulation of Foxa2 expression. These results also support our idea that increased histone acetylation acts as an enhancer of DA neuronal factors. However, the role of HDAC7 in NPCs and DA neurons has not been identified. Further study should reveal the role of HDAC7 in neurogenesis.
It is clear that histone modifications play a critical role in determining the timing of the cellular progression from NPC maintenance to mature neuronal differentiation. Generally, the genes involved in neuronal differentiation in NPCs have both repression-associated (H3K27me3) and activation-associated histone markers (H3K4me3) at their transcription start site or promoter [11,33,35]. This bivalent state allows genes to remain in either an inactive or active state, depending on the genetic cue. Many transcription factors or epigenetic regulators can act as a cue to alter the epigenetic state of target gene promoters. Treatment of NPCs with HDACI acts as an appropriate activator to change the correlation between the physical association of HDAC1/HDAC2 with RE1 sites present in specific neuronal promoters and promote the expression of neuronal genes [36]. This evidence suggests that HDACs are required, at least in part, for the repressor element silencing transcription factor-mediated silencing of many neuron-specific genes in NPCs. Further, treatment of rat subventricular zone-derived neurospheres with HDACIs promotes neuronal differentiation [37].
Taken together, these lines of evidence suggest that HDACs play an important role in the induction of neuronal differentiation and that HDACI treatment could increase the differentiation of mature neurons from NPCs. Indeed, HDACIs increase DA neuronal differentiation [37,38]. However, the molecular mechanisms related to this event remain unknown. In this study, we provide new insights into DA neuron differentiation by investigating the connection between histone acetylation and the expression of DA neuronal genes such as Foxa2. Intriguingly, apicidin treatment upregulates the expression of endogenous Foxa2 along with the increased expression of DA neuron markers. Apicidin is a novel cyclic tetrapeptide with potent, broad-spectrum antiprotozoal activity against apicomplexan parasites, and its structure is related to trapoxin, a potent HDACI. The antiparasitic activity of apicidin at low-nanomolar concentrations appears to be attributed to the inhibition of apicomplexan HDACs [19]. One-day treatment with apicidin significantly increased Foxa2 expression and DA neuron generation, suggesting that apicidin treatment, and HDACI treatment in general, could be a suitable alternative method of differentiating DA neurons from VM-derived NPCs in expanded and passaged culture.
Moreover, we have revealed the epigenetic mechanism for the reduction of Foxa2 expression during DA neuron differentiation. Histone H3 acetylation in the Foxa2 promoter region was suppressed, accompanied by the reduction in Foxa2 expression during DA neuron differentiation. Apicidin treatment inhibited the reduction of histone H3 acetylation in the Foxa2 promoter region and increased Foxa2 expression during DA neuron differentiation. These data suggest that histone acetylation is a key step to target to improve DA neuron differentiation. In addition to histone acetylation, histone methylation on specific lysine and arginine residues has an essential role in neuron generation and differentiation. The promoters of genes involved in DA neuron differentiation are methylated on H3K27 and H3K4, which are bivalent histone markers [38]. Indeed, one histone H3 trimethyl K27 demethylase, JMJD3, can activate specific components of the neurogenetic program, suggesting a critical role in epigenetic control during neurogenesis [12]. Recently, the polycomb-repressive complexes (PRC) proteins have emerged as key players in neurogenesis. During ESC differentiation into neural lineages, repression by PRC proteins and H3K27 trimethylation decrease in PRC-target neural-specific genes [39,40]. Collectively, previous reports strongly suggest that histone methyltransferases and histone demethylases are required for the proper development of the nervous system and regulate specific cohorts of genes depending on developmental timing and cellular context.
Interestingly, our data reveal that among the numerous types of histone methylation, only H3K27me3 was increased during DA neural differentiation (Fig. 3A, C). However, little is known about the precise biological function and regulatory mechanism of H3K27 methylation during DA neural differentiation. Future work should explore how and when H3K27 becomes methylated and which enzymes trigger methylation and demethylation, as well as the precise role of HDAC7 and its function. In addition, human DA stem cells immortalized with the myc oncogene showed rapid proliferation with karyotype stability and differentiated into electrophysiologically active neurons [41]. Thus, it would be of great interest to elucidate the determining factors for human fetal-derived NSCs to be maturated into functional neurons, and to determine the histone modification patterns on their promoter regions.
Footnotes
Acknowledgments
This work was supported by the Stem Cell Research Center of the 21st Century Frontier Research (2012-0006429) and Medical Research Center (NRF-2012R1A5A2A28671860) programs funded by the Ministry of Science and Technology, Republic of Korea.
Author Disclosure Statement
The authors declare no competing financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.