
Abstract
The transcriptional regulation of neural stem/progenitor cells (NS/PCs) is of great interest in neural development and stem cell biology. The RNA-binding protein Musashi1 (Msi1), which is often employed as a marker for NS/PCs, regulates Notch signaling to maintain NS/PCs in undifferentiated states by the translational repression of Numb expression. Considering these critical roles of Msi1 in the maintenance of NS/PCs, it is extremely important to elucidate the regulatory mechanisms by which Msi1 is selectively expressed in these cells. However, the mechanism regulating Msi1 transcription is unclear. We previously reported that the transcriptional regulatory region of Msi1 is located in the sixth intron of the Msi1 locus in NS/PCs, based on in vitro experiments. In the present study, we generated reporter transgenic mice for the sixth intronic Msi1 enhancer (Msi1-6IE), which show the reporter expression corresponding with endogenous Msi1-positive cells in developing and adult NS/PCs. We found that the core element responsible for this reporter gene activity includes palindromic Regulatory factor X (Rfx) binding sites and that Msi1-6IE was activated by Rfx. Rfx4, which was highly expressed in NS/PCs positive for the Msi1-6IE reporter, bound to this region, and both of the palindromic Rfx binding sites were required for the transactivation of Msi1-6IE. Furthermore, ectopic Rfx4 expression in the developing mouse cerebral cortex transactivates Msi1 expression in the intermediate zone. This study suggests that ciliogenic Rfx transcription factors regulate Msi1 expression through Msi1-6IE in NS/PCs.
Introduction
N
The Musashi family is an evolutionarily conserved group of RNA-binding proteins that were isolated as mammalian homologues of the Drosophila Musashi [2 –5]. In mammals, two Musashi proteins, Msi1 and Msi2, have been identified by characterization of two RNA recognition motifs (RRMs) [3,6]. From our previous studies using knock-out mice, Msi1 and Msi2 are known to contribute to the maintenance of stemness of neural stem/progenitor cells (NS/PCs) by repressing the translation of target mRNAs [4,7]. We have identified a consensus RNA sequence that binds to the RRMs in both mammals [8] and Drosophila [9]. Msi1 binds to the 3′-untranslated region of the Notch signaling inhibitors m-numb [8] and p21WAF [10] through RRMs and is thought to control stemness and the cell cycle through that interaction.
The transcriptional regulation of stem-cell markers has been intensively investigated, and these studies have made crucial contributions to the elucidation of the regulatory mechanisms of stem-cell fate and developmental biology. Key roles for transcription factors in rewiring transcriptional networks have been revealed by the investigation of genomic regions. The NSC-specific regulatory region of the Nestin gene was found in its second intron and contains binding sites for SOX (Sry-like HMG box protein) and POU (Pit-Oct-Unc) factors [11,12]. Oct4, a member of the POU family, directly binds with Sox2, and it forms a core transcriptional network with Nanog to activate or repress downstream genes for maintaining embryonic stem cells (ESCs) [13]. During the transition from ESCs to NSCs, Sox2 changes its partner from the POU family member to Brn2, and they cooperatively regulate the second intron enhancer of Nestin [14,15]. Indeed, Brn2 is important for the induction of NSCs [15]. Many transcription factors, including Tcf3, Smad1, Stat3, Esrrb, Zfx and Klf4, have been shown to play important roles in the control of ESC state; remarkably, the transcription factors associated with the LIF, Wnt, and BMP4 signaling pathways (Stat3, Tcf3, and Smad1) tend to co-occupy enhancers bound by Oct4, Sox2, and Nanog [13]. Compared with the transcriptional network in ESCs, the mechanisms in NSCs are not well understood. Thus, further investigation into the transcriptional network in NSCs is required.
We have previously demonstrated that the regulatory region of Msi1 in NS/PCs is located in the sixth intron of the Msi1 genomic locus (Msi1-6IE): The activity of a 184 kb Msi1-BAC reporter is dependent on the Msi1 locus (exons and introns), including 595 bp of Msi1-6IE [16]. Msi1-6IE is highly conserved in mammals and is an active enhancer that is H3K4me1-positive and includes the p300-binding region in NS/PCs. The reporter gene containing Msi1-6IE shows activity specifically in Msi1-positive NS/PCs during the process of neural differentiation of ESCs to NS/PCs in vitro, and it does not show activity in differentiated neurons and astrocytes.
In the present study, we demonstrate that the Msi1-6IE reporter gene is specifically expressed in Msi1-positive NS/PCs in the embryonic and adult mouse brain and show that Msi1 is regulated by a master regulatory factor for ciliogenesis, the regulatory factor X (Rfx) transcription factor, through Msi1-6IE.
Materials and Methods
Generation of the transgenic mice
The Msi1-P1-ffLuc (Msi1-P1) and Msi1-6IE-ffLuc plasmids (Msi1-6IE) have been previously generated and described [16]. DNA was linearized at a site within the vector with BsiWI and ApaLI, and the plasmids were injected into the pronuclei of fertilized BDF1 background mouse eggs in RIKEN Brain Science Institute (BRC05764). Five lines of P1 and four lines of 6IE transgenic (tg) mice were obtained, and one line was chosen for the studies. All animal care and treatment procedures were performed in accordance with institutional guidelines approved by the Experimental Animal Care Committee of the Keio University School of Medicine (Approval number: 09091-10). Genotypes were determined by polymerase chain reaction (PCR) of genomic DNA prepared from tail biopsies. The primer sequences for the genotyping were as follows: 5′-CTACCTGAGCTACCAGTCCGCCCTG-3′ and 5′-CAGCTAACGACATCTACAACGAGCGCGAG-3′.
Histological analysis
Frozen sections (12 μm) of embryonic brains fixed with 4% (w/v) paraformaldehyde were prepared using a cryostat (CM3050S-IV; Leica). Adult brains were perfused with 4% paraformaldehyde, and the brains were dissected, postfixed with 4% paraformaldehyde at 4°C overnight, and sliced into 40-μm sections using a vibratome (VT1200S; Leica). To detect Mash1-positive cells, tissue sections were pretreated with ice-cold acetone for 30 s. The sections were preblocked with TNB blocking buffer (PerkinElmer) for 1 h at room temperature and incubated with primary antibodies at 4°C overnight, followed by incubation with Alexa Fluor-biotin-conjugated secondary antibodies for 1 h at room temperature. The Vectastain Elite ABC kit (Vector Laboratories) and Tyramide Signal Amplification-Red (PerkinElmer) were used for visualization. To detect BrdU incorporation, tissues were pretreated with 1 N HCl at 37°C for 30 min. The following antibodies were used: anti-Msi1 [rat immunoglobulin G (IgG); [43]; 1:400], anti-brain lipid binding protein (Blbp) (rabbit IgG; Chemicon AB9558; 1:200), anti-GFAP (rat IgG; Invitrogen 13-0300; 1:200), anti-Nestin (mouse IgG; BD 556309; 1:250), anti-Sox2 (rabbit IgG; Millipore mab377; 1:100), anti-Mash1 (mouse IgG; BD 556604; 1:200), anti-DCX (goat IgG; Santa Cruz Biotechnology sc-8066; 1:500), anti-βIII tubulin (mouse IgG; Sigma-Aldrich T8660; 1:500) anti-BrdU (rat IgG; Abcam ab6326; 1:100), and anti-GFP (chick IgY; Aves GFP-1010; 1:500). The images were captured by confocal laser-scanning microscopy (LSM700; Carl Zeiss).
Cell culture
Neurosphere culture
Cortical cells were dissected from E14.5 Msi1-6IE mice, pipetted 20 times and cultured in previously described media hormone mix (MHM) medium supplemented with 20 ng/mL fibroblast growth factor (FGF)-2 and 20 ng/mL epidermal growth factor (EGF) for 6 days [16,17].
Neuronal and glial differentiation of NS/PCs
The neurospheres were dissociated with TrypLE Select (Life Technologies), plated on poly-
Neural induction of ESCs
ESCs (EB3 tg14 line) were cultured in suspension conditions for 2 days and induced to neural lineage by the addition of all-trans retinoic acid (Sigma R2625) to a final concentration of 10−8 M for 4 days, as previously described [16,17].
Flow cytometer
Neurospheres were dissociated with TrypLE Select (Life Technologies), resuspended in MHM containing 1 mg/mL propidium iodide, and filtered (30 μm). Cell sorting and analyses were performed using a MoFlo (Beckman Coulter) flow cytometer/cell sorter equipped with CELLQuest software (Becton-Dickinson).
Plasmids
The genomic regions used to find the core enhancer in 6IE were generated with the following primers: 6IE 316 (5′-gtcgacGATCTGGGTCCAAGACGCAG-3′ and 5′-atgcatCTCGCTCCCTGGCAGCCTATTC-3′), 6IE 161 (5′-gtcgacGATCTGGGTCCAAGACGCAG-3′ and 5′-gtcgacCCTCCAATCACAGCTGCTGGATCAG-3′), 6IE 178 (5′-gtcgacGATCCAGCAGCTGTGATTGGAGG-3′ and 5′-gtcgacCCTCCAATCACAGCTGCTGGATCAG-3′), 6IE 279 (5′-gtcgacGGGGACAGCCGGGAATGCCAAG-3′ and 5′-gtcgacCTCCTGAGGCTGGCTGAG-3′), 6IE 181 (5′-gtcgacCCGCCATGGCTGCCACTCAG-3′ and 5′-gtcgacCTCCTGAGGCTGGCTGAG-3′).
Mutagenesis
The mutations of transcription-factor binding sites in 6IE were generated with the following primers: C/EBP (5′-AGCAGCTGTG
All Rfx expression vectors and NfI expression vectors for the luciferase assay were constructed in an entry vector (pENTR-3xHA) and recombined into CIIEF-RfA-IRES2-Venus.
Rfx expression vectors for chromatin immunoprecipitation (ChIP)-PCR and in utero electroporation were constructed on pENTR-3xHA and recombined into pCAGGS2-RfA.
Luciferase assays
Neurospheres derived from mouse E14.5 telencephalons were dissociated, and 1×105 cells were cultured on 24-well plates in MHM (+20 ng/mL FGF-2, +20 ng/mL EGF) for 2 days, after which they were transfected with 400 ng reporter plasmids and pRL-SV40 as an internal control, using GeneJuice (Novagen). After 2 days, the cells were washed with phosphate-buffered saline and then lysed with 100 μL lysis buffer (Toyo Ink) for 20 min at room temperature. For each sample, 25 μL luciferase substrate solution (Toyo Ink) was added to 5 μL lysate, and the luminescence was measured with a Berthold Centro LB960 luminometer. The assays were internally calibrated with a standardized Renilla luciferase solution (Toyo Ink) to ensure uniformity between experiments. For the reporter activity with transcription factors, 10 ng of the Rfx or NfI expression vectors was cotransfected into NIH3T3 cells, which were cultured for 2 days and measured for luminescence as described above.
Quantitative reverse transcription-polymerase chain reaction
Total RNA was isolated from cells with the RNeasy Mini Kit (Qiagen). Synthesis of cDNA was performed with Superscript II RNase H reverse transcriptase (Invitrogen) at 42°C for 50 min according to the manufacturer's instructions. Quantitative PCR was performed with SYBR Premix Ex Taq II (Tli RNaseH Plus) (RR820A; TaKaRa) in MX3000P Real-Time PCR Machine (Stratagene). The quantitative reverse transcription-polymerase chain reaction (qRT-PCR) assay was conducted with amplification conditions at 95°C 10 min for enzyme activation, 50 cycles of denaturation at 95°C for 30 s, 60°C for 1 min for annealing, 72°C for 30 s for extension. Serial dilutions ranging from 108, 106, 104, to 102 copies per 1 μL of DNA solution were prepared as standard samples by subcloning each PCR product into the Zero Blunt TOPO PCR Cloning Kit (K2800; Invitrogen). To analyze the relative expression in different samples, the amount of cDNA was normalized to the level of ubiquitously expressed β-actin mRNA. The primer sets used were as follows: GFP (5′-TGAACCGCATCGAGCTGAAGGG-3′ and 5′-TCCAGCAGGACCATGTGATCGC-3′), Msi1 (5′-GGGATGGACGCCTTCATGCTG-3′ and 5′-TGGCTTGGAACCCTGGGTAAC-3′), Rfx1 (5′-TAGTCCAGGCCACTCCACAGA-3′ and 5′-TACTGCACCTGGCTGGAATACAC-3′), Rfx2 (5′-CCAGTTCCTGCTCAAGTGGTC-3′ and 5′-ACACGGTGCTCCACCAGGTA-3′), Rfx3 (5′-CAAGCGCTGAAGCCCTATGAA-3′ and 5′-CTAGCAGCACTGCGCAAGGTTA-3′), Rfx4 (5′-TTGCCTCATGACACAGCCATC-3′ and 5′-TGCTCATCAAACAAGGTTCCATC-3′), Rfx5 (5′-TACAGAAATTCTCGGACAACGACAA-3′ and 5′-TGCGGATCCACCTATAGGCATAC-3′), Rfx6 (5′-CCAAGGGTCAACATTGCCTTC-3′ and 5′-GTTCACCATCGATCCAAGCCTA-3′), Rfx7 (5′-CCATTAAGTGCTGCTGACTTTGG-3′ and 5′-TGGGCAGTGTTGGCATATGAA-3′), β-actin (5′-GGTGGGCCGCCCTAGGCACCA-3′ and 5′-TTGGCCTTAGGGTTCAGGGGG-3′).
Chromatin immunoprecipitation-polymerase chain reaction
For ChIP-PCR, 3×106 NS/PCs derived from E14.5 cortex were electroporated with 30 μg of CAGGS2-3xHA or pCAGGS2-Rfx4-3xHA using NEPA21 Type2 electroporator (NEPA GENE), and these cells were cultured on poly-
The primer sequences for the PCR reaction were as follows: 6IE core 1 (5′-GTGTCTTTGTGTCTGAGCACC-3′ and 5′-CTGGCAGCCTATTCTCCAGCTC-3′), 6IE core 2 (5′-CACTAATCTGATCCAGCAGCTG-3′ and 5′-GGTTTCACATGCCAACG-3′), Upstream (5′-CTTCTGTAGCCAGTTCAATCC-3′ and 5′-CTGAGAGCTAGTGACCTCTCAG-3′). Downstream (5′-CTATTAACAGCTGAAGCAGCTG-3′ and 5′-GATTCCACAGTCTGTAAAATGG-3′).
In utero electroporation
A plasmid expressing GFP driven by the CAG promoter was used with either an empty pCAGGS2-3xHA vector or pCAGGS2-Rfx4-3xHA. The introduction of DNA by in utero electroporation was performed essentially as described previously [18]. Briefly, ICR pregnant mice were anesthetized with isoflurane and nitrous oxide, and 1 μL of DNA solution was injected into the lateral ventricle at E13.5. The solution contained 0.3 μg GFP plasmid and 3 μg of either the Rfx4 plasmid or the empty vector. After injection, electroporation was performed using CUY-21 (NEPA GENE) to deliver eight cycles of square pulse (50-ms pulses of 37 V with 950-ms intervals) per embryo. Embryos were reimplanted in utero, and embryonic brains were harvested 3 days later.
Results
Msi1-6IE reporter gene is expressed in Msi1-positive NS/PCs in vivo
To evaluate Msi1-6IE reporter expression in vivo, we generated Msi1-6IE reporter tg mice. We used Msi1-6IE constructs containing 595 bp of 6IE aligned downstream of ffLuc, as previously reported ([16], 6IE is described as D5E2 in this paper) (Fig. 1A). The ffLuc reporter gene is a fused gene of Venus and firefly luciferase sequences, and it allows both visualization of the expressing cells by fluorescence and quantification of the transcriptional activity by bioluminescence [19]. A 1-kb Msi1 promoter upstream of the Msi1 transcription start site was placed in the upstream portion of ffLuc. Similar to the Venus expression in 180 kb Msi1-BAC tg mice [16], GFP fluorescence was observed in the CNS from forebrain to spinal cord of the E12.5 Msi1-6IE tg mice (four of four lines) (Fig. 1B), and this fluorescence was not detected with the 1-kb Msi1 promoter reporter tg without 6IE (Msi1-P1, 0 of 5). Next, we examined the immunohistochemical characterization of Venus (which can be detected with anti-GFP antibody) and endogenous Msi1 expression using embryos from P1 and 6IE tg mice (Fig. 1C, D). Venus expression was observed in the VZ and subventricular zone (SVZ) in Msi1-6IE tg mice, where NS/PCs are present, and colocalized with Msi1 in E14.5 cortex (Fig. 1D), midbrain (Fig. 1F) and hindbrain (Fig. 1G, H). Venus-expressing cells were not stained with the neuronal marker βIII tubulin (Fig. 1E). We could not detect the Venus expression in Msi1-P1 tg mice brain by immunostaining with GFP antibody (Fig. 1C). These results support our previous in vitro data showing that 6IE regulates Msi1 expression in the neural lineage and that the 1-kb promoter region is not sufficient for transactivation of the reporter gene.

Expression of the Msi1-6IE reporter corresponds to endogenous Msi1 expression in NS/PCs in vivo.
Msi1-6IE-positive cells are self-renewing and multipotent in vitro
To reveal the properties of Venus-expressing NS/PCs, we dissected and dissociated the cortex from E14.5 Msi1-6IE tg mice, and Venus-positive cells were sorted by flow cytometry. The Venus-positive cells proliferated in culture medium containing FGF-2 and EGF, which maintains Venus expression, and formed neurospheres in floating culture (Fig. 1I, J). The neurospheres were dissociated and cultured in differentiation medium without growth factors to test the multipotency of Msi1-6IE cells by immunocytochemistry. Under the growth conditions, Msi1-6IE cells expressed the NS/PC marker Nestin (Fig. 1K). Under the differentiation conditions, the purified NS/PCs produced β-III-tubulin-positive neurons, glial fibrillary acidic protein (GFAP)-positive astrocytes and 2′,3′-cyclic nucleotide 3′ phosphodiesterase-positive oligodendrocytes (Fig. 1K).
Msi1-6IE reporter recapitulates the spatio-temporal regulation of endogenous Msi1
To gain insight into the temporal regulation of 6IE, we immunostained adult Msi1-6IE tg mouse brain and analyzed the distribution of Venus-positive cells in the hippocampus and cortex, where NS/PCs are present. In the subgranular zone (SGZ) of hippocampus, radial-glia-like stem cell (Type 1 cell) gives rise to three consecutive stages of transiently amplifying PCs (Type 2a, Type 2b and Type 3 cells). These stages lead to a differentiation of transient postmitotic granule cells (GCs) following to terminally differentiated mature GCs [20]. Endogenous Msi1 is mostly expressed in the SGZ, colocalized with cells expressing Blbp, a marker of Type 1 and Type 2a cells, and it is not expressed in NeuN-positive mature neuronal cells (Supplementary Fig. S1A, C; Supplementary Data are available online at
In the cortex, Venus-positive cells were observed in subependymal zone along the lateral ventricle. These cells showed good co-localization with Msi1 (Supplementary Fig. S1T). The majority of endogenous Msi1-positive cells were observed in Sox2-positive and Nestin-positive Type B radial glia-like cells or Type C precursors (Supplementary Fig. S1P, Q), and some PSA-NCAM-positive and Dcx-positive Type A neuroblasts were Msi1 positive cells during neuronal differentiation process (Supplementary Fig. S1R, S). The Msi1-6IE reporter was able to mimic the expression in these cells (Mash1 was used for labeling Type C cells. See Supplementary Fig. S1U–W). Based on these observations, we concluded that the transactivation of the sixth intronic enhancer can reflect the authentic spatio-temporal expression of Msi1.
The 178-bp core enhancer site is responsible for the transactivation of Msi1-6IE
Msi1-6IE includes many transcription-factor binding sites, and several candidate binding sites have been observed from the correlation of the localization with Msi1 and/or the functional aspects with Msi1, as we previously reported [16]. For example, Sox and Ap-2 are expressed in NS/PCs in a subtype-specific fashion and control the balance between proliferation and differentiation [21 –24]. To elucidate which transcription factors regulate Msi1, we performed a deletion study of the reporter gene to find the regulatory region responsible for the transactivation of 6IE. We first generated two Msi1-6IE reporter constructs that were gradually shortened from the 3′ end of 6IE (6IE-316 and 6IE-161) (Fig. 2A). The luciferase activity of ffLuc in 6IE-316-transfected cells was significantly higher than with the 1-kb promoter alone (P1) at a level similar to the full length of the 6IE reporter gene in NS/PCs derived from E14.5 cortex. However, the activity of 6IE-161 was markedly decreased, to almost the same level as P1 (Fig. 2B). These results indicated that the critical regulatory region was included in the region from 161 to 316 bp of 6IE. To confirm this result, we generated reporter genes including the common sequences between 6IE-161 and 6IE-316 (6IE-178), the region between full-length 6IE and 6IE-316 (6IE-279), and the 3′ 181 bp of 6IE (6IE-181). 6IE-178 had transcriptional activity at the same level as 6IE-316, whereas 6IE-279 and 6IE-181 showed low activity, consistent with their relationships with 6IE-316. These data show that 6IE-178 is required for the transactivation in NS/PCs as a cis-element, suggesting the presence of important transcription factor(s)-binding sites regulating Msi1 transcription in this 178-bp genomic region.
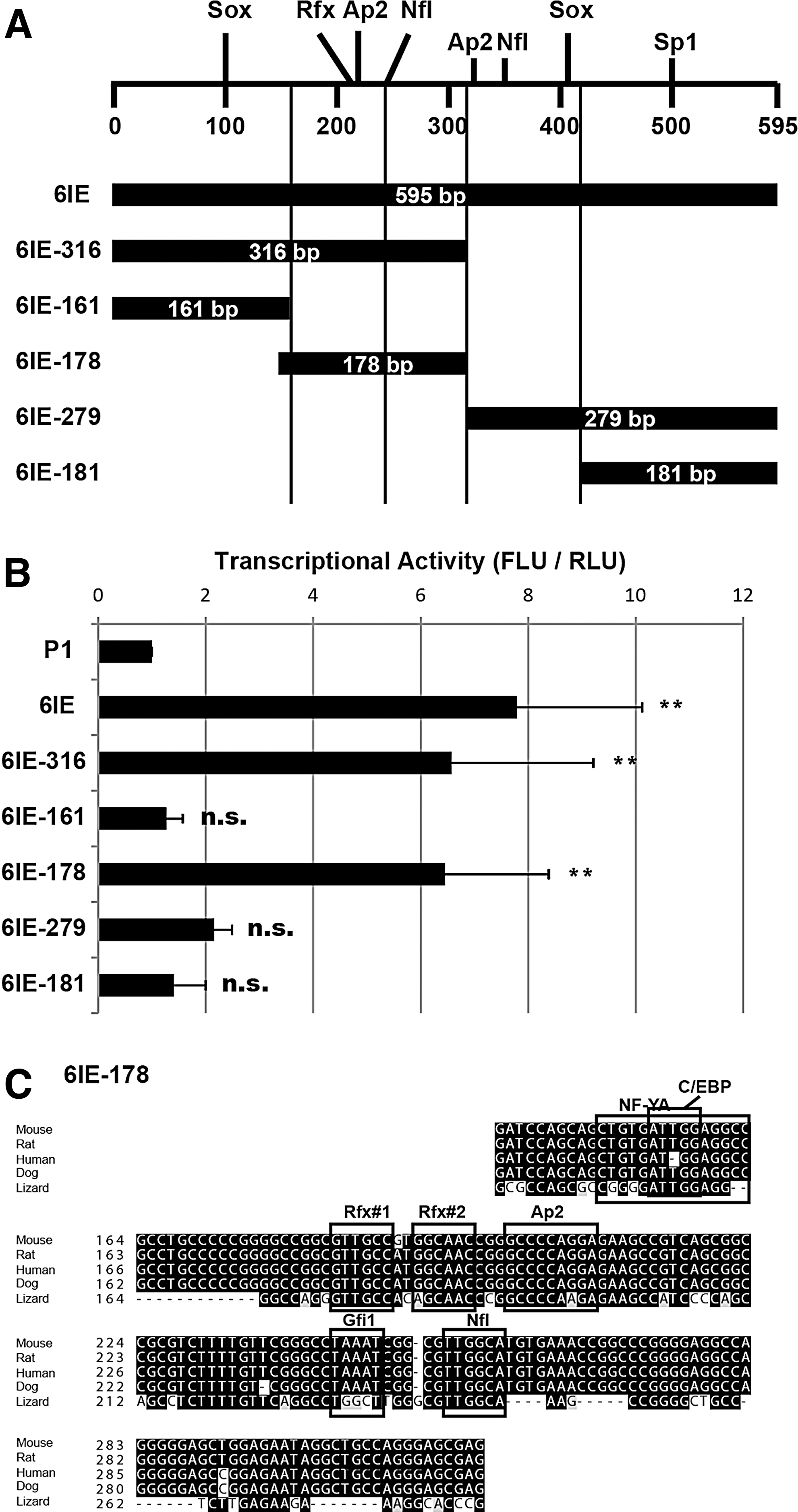
Identification of core enhancer site in 6IE.
Palindromic Rfx transcription-factor binding sites determine the efficiency of transactivation in the 6IE region
To identify the transcription factor(s) regulating Msi1, we searched for potential transcription-factor binding sites in 6IE-178 by in silico analysis, and we observed that there were highly homologous motifs with consensus sequences for NF-Y (or C/EBP), Rfx, Ap-2, Gfi1 and NfI (Fig. 2C). Two palindromic Rfx-binding sites (5′-GTTGCC-3′) were present across two bases of 5′-GT-3′ (Rfx#1 and Rfx#2). We compared the genomic sequences of 6IE-178 among 60 species including 40 mammals and 20 other species including fishes, reptiles, birds and amphibians (UCSC genome browser conservation track). Each binding site was identical among the most of mammals including mouse, rat, human and dog, and Rfx, Ap-2 and NfI sites were completely conserved even in lizard (Anolis carolinensis). To test whether these binding sites were required for activation of 6IE, we disrupted them by introducing mutations (see the “Materials and Methods” section for their mutation sequences). Unexpectedly, when either of the mutations in the Rfx-binding sites was introduced into 595 bp of Msi1-6IE reporter gene, enhancer activity was obviously attenuated in NS/PCs (Fig. 3A, B). The mutation in the NfI-binding site also showed mildly but significantly decreased activity against wild type (no mut.), but the mutations in other binding sites were not significantly different. Noteworthy, simultaneous mutations of both Rfx#1 and Rfx#2 sites did not exhibit any additive effect compared to single mutants. These results suggest that the Rfx binding sites and the NfI binding site are involved in the regulation of Msi1-6IE.

Rfx transcription factor binding sites regulate 6IE.
Genes encoding seven Rfx proteins have been identified in vertebrates, based on a highly conserved DNA binding domain [25 –27]. The conserved dimerization domains, which mediate homo- and heterodimerization, are present in Rfx1 through 4 but are lacking in Rfx5 and Rfx7. Rfx proteins recognize a highly conserved regulatory element called the X-box, which is frequently observed as a palindromic sequence (5′-GTNRCC (0-3N) RGYAAC-3′) [28].
To establish the contribution of Rfx transcription factors to the regulation of the Msi1-6IE, we exogenously expressed Rfx1-4 in NIH3T3 cells (Fig. 3C). When each Rfx was expressed, the cotransfected Msi1-6IE reporter was activated (no mut. in the Rfx expression) compared to reporter alone (no gene). We also tested whether functionally disrupted Rfx4 expression (Rfx4L298P) could activate wild-type 6IE. Rfx4L298P has a point mutation in the dimerization domain and shows impaired transcriptional activation ability due to its failure of nuclear localization, as previously reported [29]. As expected, the Rfx4L298P could not activate the Msi1-6IE reporter compared to wild-type Rfx4 expression (Fig. 3C).
To confirm that the reporter activities induced by Rfx transcription factor were dependent on Rfx binding sites, Msi1-6IE reporters carrying either Rfx#1 or Rfx#2 mutations or Rfx#1 and Rfx#2 double mutation were used for the luciferase assay (Fig. 3C). The overactivation produced by the expression of each Rfx was significantly attenuated when mutations in #1, #2 or both (#1 and #2) were introduced into the 6IE sequence. Each single mutation repressed the activity to the same degree as the double mutations. The similar results were obtained by the reporter assay in NS/PCs (Fig. 3A). Thus, the palindromic Rfx binding sites are necessary for efficient 6IE activation, suggesting the dimer of Rfx transcription factors may regulate 6IE.
Rfx expression in embryonic forebrain and NS/PCs
Rfx members belong to the winged-helix subfamily of helix-turn-helix transcription factors, which have been identified in various eukaryotic organisms from yeast to human [30]. The expression levels of Rfx family transcription factors vary among mouse tissues [25]. Rfx1, Rfx5 and Rfx7 are ubiquitously expressed in essentially all types of tissues, whereas Rfx6 shows highly tissue-specific expression in liver, heart and pancreas. Despite these many differences, all Rfx transcription factors except for Rfx6 are highly expressed in brain. The loss of Rfx4 function results in pleiotropic brain defects, and ablation of Rfx3 produced hydrocephalus and abnormal differentiation of ependymal cells [28,29,31]. Knockdown of Rfx7 results in failure of neural tube closure in Xenopus embryos [32]. However, the precise localization of Rfx genes in brain is poorly understood. To gain a quantitative understanding of the expression of Rfx in NS/PCs, we quantified the mRNA levels of Rfx genes in neurospheres and embryonic forebrains by qRT-PCR. All of the Rfx genes except for Rfx6 were detected in the forebrain from E12.5 to E18.5 (Fig. 4A). Rfx1, Rfx3, Rfx4 and Rfx7 were also strongly expressed in neurospheres dissociated from the E12.5 to E18.5 cerebral cortices. Noteworthy, at any stage examined, the expression levels of Rfx4 were much higher in neurospheres than in the corresponding forebrains (Fig. 4B). These results suggest that Rfx4 is highly expressed in cortical NS/PCs. To address the regulation of Msi1-6IE by Rfx, NS/PCs dissociated from neurospheres derived from the cortices of E14.5 Msi1-6IE tg mice were divided into Venus-positive and Venus-negative cells by flow cytometry, and the expression of Rfx genes was quantified in each population (Fig. 4C). Consistent with the higher expression in neurosphere culture, the expression of Rfx4 was significantly highly enriched in Venus-positive NS/PCs compared with Venus-negative cells. In contrast, Rfx1, Rfx3, and Rfx7 did not exhibit such an enrichment in Venus-positive cells. We also analyzed the expression of Rfx genes in Nestin-second intronic enhancer reporter mice (Nestin-d4-Venus) in embryonic forebrain to confirm this finding in NS/PCs (Supplementary Fig. S2A) [33]. Consistent with the expression of Rfx4 in VZ of forebrain revealed by in situ hybridization [28], significantly higher amounts of Rfx4 mRNA were found in Nestin-Venus positive NS/PCs than in Venus-negative cells. Rfx3 had a similar expression pattern to Rfx4, probably because these samples included the ventral cortex area. Rfx3 was detected in the ganglionic eminence by immunohistochemistry, but it was very weakly expressed in dorsal cortex (data not shown). Further analysis in embryoid bodies derived from ESCs was conducted to compare Rfx expression to Msi1 expression during the process of differentiation to NS/PCs (Supplementary Fig. S2B). Msi1 expression was increased along with neural differentiation, and it was relatively high in neural induced cells (+RA) and low in −RA cells. Rfx4 expression was increased in a similar way as the Msi1, and it was specifically expressed by neural induction, whereas non-neural Rfx6 showed opposite results. All of these expression profiles suggested that Rfx4 was the most likely candidate gene involved in the transcriptional regulation of Msi1 transcription among the seven Rfx genes, although there may be redundant regulation by Rfx1, Rfx3 and Rfx7 in NS/PCs.
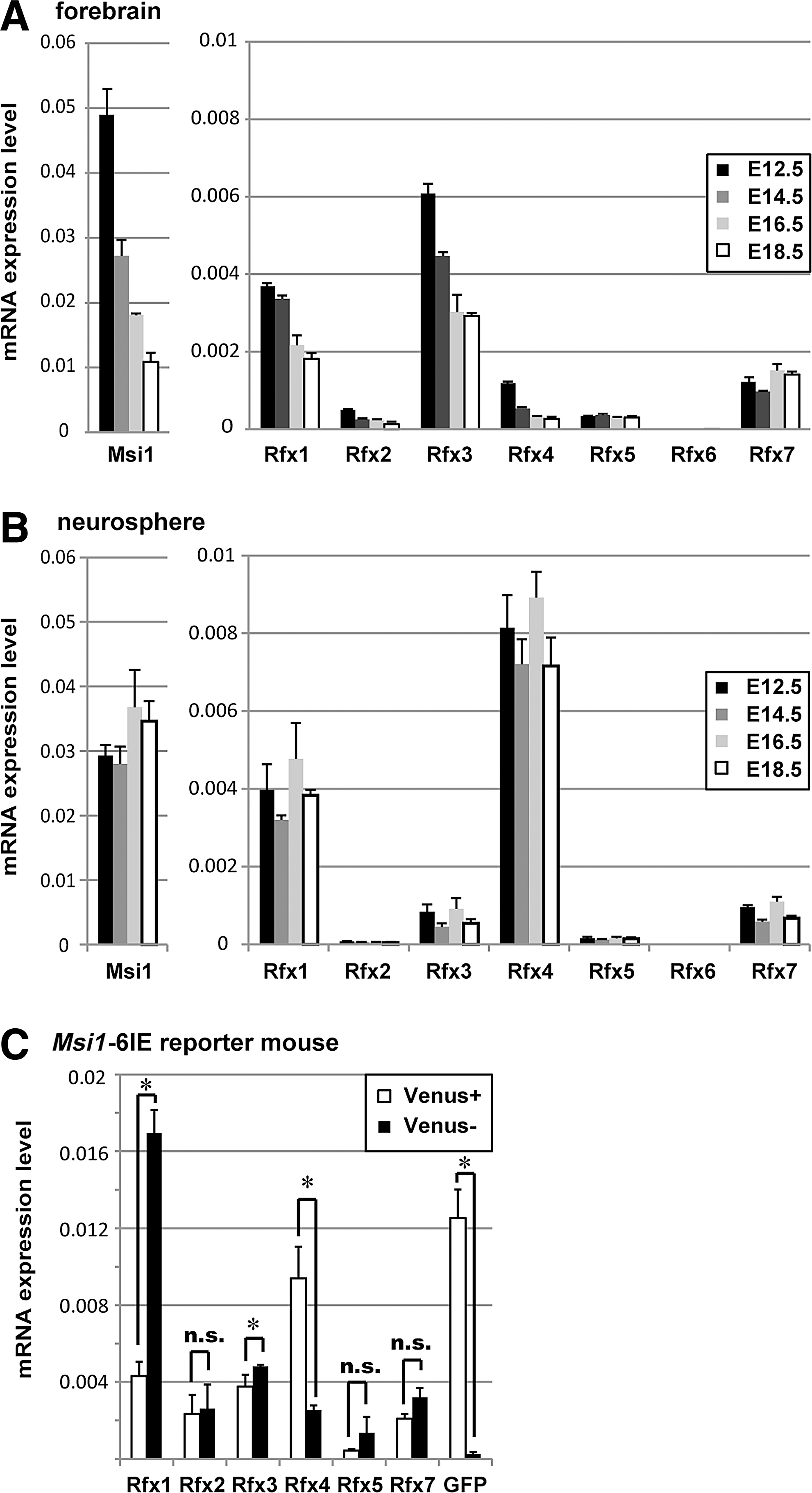
Quantification of Rfx expression in NS/PCs.
Binding of Rfx4 to the Msi1-6IE in NS/PCs
To test whether Rfx4 binds to the 6IE in NS/PCs, ChIP followed by PCR (ChIP-PCR) was performed (Fig. 5). The NS/PCs dissociated from the neurospheres derived from E12.5 cortex were electroporated with control plasmid or HA-tagged Rfx4 expression plasmid. The Rfx4-binding genomic regions were immunoprecipitated with anti-HA antibody. Only the 6IE region precipitated by Rfx4 was amplified, suggesting that Rfx4 binds to 6IE but not to each region upstream or downstream of 6IE.
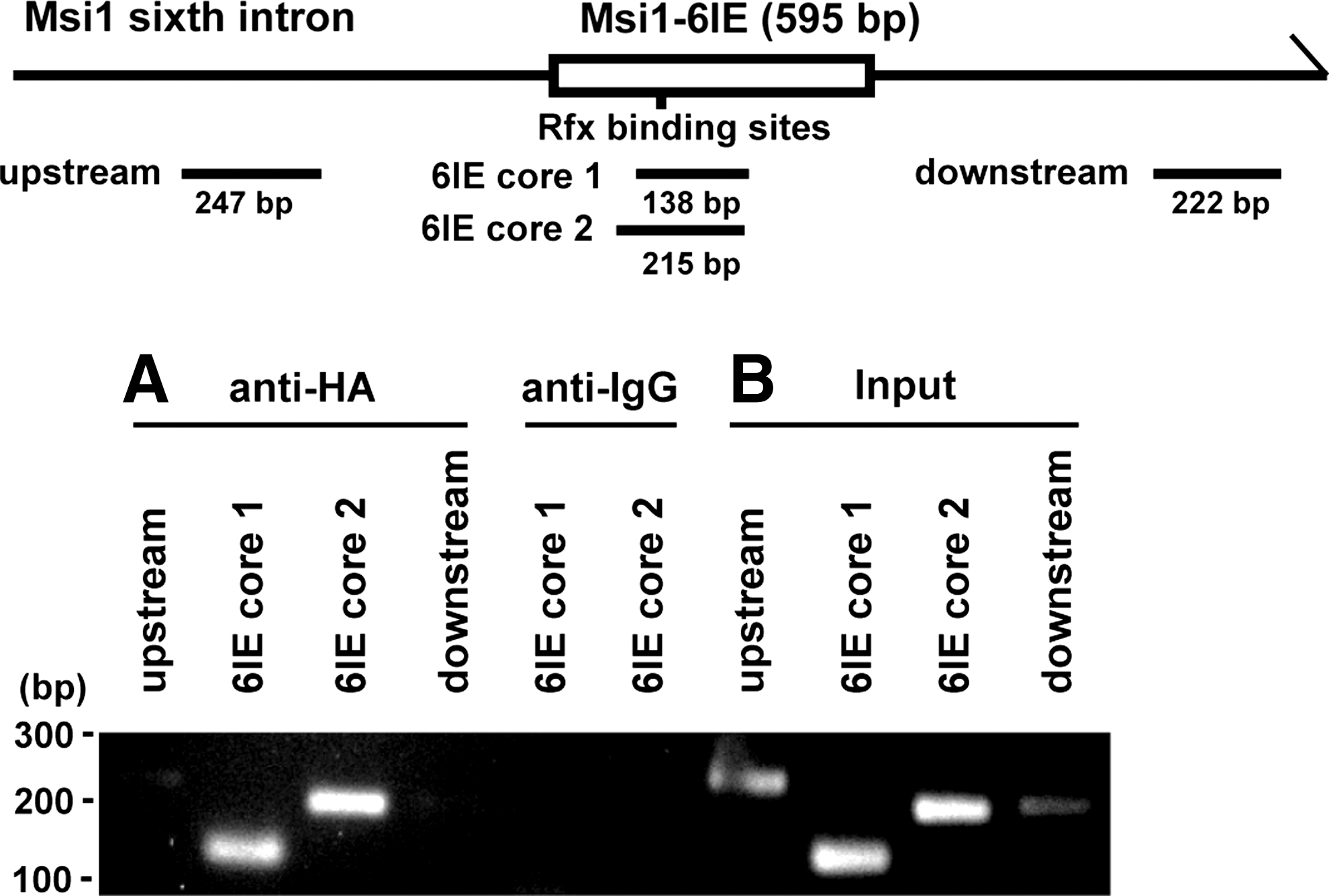
Rfx4 binds to the Msi1-6IE region. chromatin immunoprecipitation-PCR analysis of Rfx4 binding to Msi1-6IE in NS/PCs.
Overexpression of Rfx4 shows ectopic Msi1 expression in vivo
Because Rfx4 transactivated the regulatory region of Msi1 in NS/PCs, we examined whether overexpression of Rfx4 could ectopically promote the endogenous Msi1 expression in the cerebral cortex. HA-Rfx4, along with GFP, was overexpressed in the developing dorsal cortex by in utero electroporation at E13.5. At E15.5, the control GFP-expressing cells were mainly observed in the intermediate zone (IZ), and those cells did not express Msi1 (Fig. 6). In contrast, HA-Rfx4-expressing cells in the IZ ectopically and highly expressed Msi1. Taken together, these results demonstrate that Rfx4 induces endogenous Msi1 expression through Msi1-6IE in the developing nervous system.

Rfx4 expression shows ectopic Msi1 expression in IZ. Immunostaining of cortical periventricular layers at E15.5 after in utero electroporation of control plasmid (upper) and Rfx4 (lower) at E13.5. Msi1 expression in IZ was observed only in Rfx4-electroporated slices with colocalization with GFP. IZ, intermediate zone; SVZ, subventricular zone; VZ, ventricular zone. Scale bars: 50 μm.
Discussion
To better understand the regulation of NS/PCs, we focused on the transcriptional regulation of the Msi1 gene, which marks these cells. Our study took advantage of the properties of Msi1 based on the previous observation that Msi1 is expressed preferentially in NS/PCs in embryonic brain and functions to maintain the NS/PCs in an undifferentiated state. In this study, we show that Msi1 expression is regulated by Rfx transcription factors through the sixth intronic enhancer of the Msi1 gene in NS/PCs.
We previously identified the NS/PC-specific enhancer of the Msi1 gene in the sixth intron [16]. The stable lines of ESCs containing the Msi1-6IE reporter gene showed the increased activity by the induction to neural lineage in embryoid bodies, and they continued the expression of reporter gene in tertiary neurosphere culture in vitro. Therefore, 6IE was thought to be involved in Msi1 expression from developmental stages to adult brain in vivo. In the present study, we confirmed this temporal expression by using the Msi1-6IE tg mouse in both embryonic and adult brain. Further, the Msi1-6IE reporter expression was well colocalized with endogenous Msi1-positive NS/PCs, and these cells were negative for neuron markers. Our present data indicate that 6IE is the authentic regulatory region for Msi1 transcription, and it is useful for understanding the transcriptional regulation of Msi1. While endogenous Msi1 is highly expressed in NS/PCs, it is also weakly expressed in GFAP-positive astrocytes in adult brain [34]. The reporter expression in Msi1-BAC tg mouse was confirmed in GFAP-positive astrocytes [16]. In contrast to these expression patterns, Msi1-6IE reporter expression was not detected in GFAP-positive astrocytes in the cerebral cortical layer and in the area outside the SGZ by immunostaining with anti-GFP antibody (Supplementary Fig. S1E). This result suggests that Msi1-6IE is an active genomic region specifically in NS/PCs, and the potential regulatory regions for Msi1 transcription in astrocytes are present in other regulatory regions included in the 180-kb Msi1-BAC reporter.
We identified a 178-bp core element of 6IE containing NF-YA (or C/EBP), palindromic Rfx, Ap-2, Gfi1 and NfI binding sites. It has been reported that the NF-Y transcription factor regulates Sox3 promoter activity: Sox3 is expressed in NSCs from the earliest stages of development [35,36]. It is also known that NF-Y consists of three subunits, NF-YA, NF-YB, and NF-YC, and binds to the CCAAT sequence of the Y box in the major histocompatibility complex class II (MHC-II) promoter [37]. Rfxank forms a complex with the Rfx5 dimer and Rfxap, and this complex associates with NF-Y to transactivate the MHC-II gene [38,39]. From these reports, we hypothesized that NF-Y and Rfx form a complex for Msi1 transcription in the CNS. Therefore, we disrupted each binding site in Msi1-6IE. The reporter activity was significantly repressed by the mutation in tandem Rfx binding sites, but it was not affected by mutation of the NF-Y binding site in NS/PCs. This result indicates that Rfx regulates 6IE, but NF-Y is not required for the transactivation. The mutation in the NfI binding site in the Msi1-6IE reporter gene also reduced transcriptional activity. NfIa, NfIb, and NfIx are expressed in the developing mouse brain by E15 and E16, and NfIa and NfIb are expressed in the VZ and SVZ [40]. NfI targets the GFAP promoter prior to the induction of astrocyte differentiation in NS/PCs [41,42]. Because endogenous Msi1 is also expressed in both NS/PCs and in the astrocyte [43], we tested whether the Msi1-6IE reporter was activated by NfIa or NfIb expression in NIH3T3 cells. However, these NfI proteins could not transactivate the reporter. This result is consistent with the Msi1-6IE reporter expression in Msi1-6IE tg mice, that is, the activity was specific to NS/PCs and was not detected in astrocytes in the cerebral cortex or hippocampus. Thus, it was conceivable that other transcription factors regulate this NfI-binding site. AP2 and Gfi1 were other candidates for 6IE regulatory factors. The AP2 binding sequence in 6IE-178 (GCCCCAGG) was homologous to the consensus AP2γ binding sequence (GCCC/T/G G/C G/A/CGGG/C) [44]. AP2γ is expressed in Pax6-positive NSCs, where Msi1 is expressed, and a subset of apical VZ progenitors, and it specifies basal progenitor fate in the occipital cortex [45]. The Gfi1 proto-oncogene encodes a zinc-finger protein that binds to the sequence AATC, and the mRNA of Gfi1 is expressed in the developing brain and in several specialized sensory cells of the peripheral nervous system [46]. The binding sites for AP2 and Gfi1 are conserved in mammals; thus, we disrupted the AP2 and Gfi1 binding sequences, but significant differences of reporter activities were not observed with mutant constructs compared to the wild-type sequence of the 6IE reporter.
In addition to the repression of the Msi1-6IE reporter activity by the introduction of mutations in the Rfx binding sites, the expression of all four Rfx transcription factors could significantly activate the Msi1-6IE reporter gene. ChIP-PCR experiment revealed that Rfx4 binds to the 6IE region of the genome in NS/PCs (Fig. 5). Further supporting ChIP-sequence data was reported in Creyghton et al. [53] that clearly showed that Rfx1 also binds to 6IE with an active enhancer signature (H3K27ac) in NS/PCs (Supplementary Fig. S3) [15]. Moreover, the exogenous Rfx4 expression in the developing cortex could transactivate the endogenous Msi1 expression ectopically in the IZ. Rfx transcription factors bind to tandem binding sites, called the “X-box,” in the regulatory regions of several genes, including IL-5Ra, HLA-DRA, and c-myc, and disruption of one of the two binding sites inhibits the DNA binding of Rfx [30]. Our luciferase assays for Msi1-6IE activity agreed well with these observations. The transcriptional activities of single mutations in each Rfx binding site were sufficient to reduce the activity to the same level as the mutations of both binding sites. Taken together, we concluded that Rfx transcription factors regulate Msi1 expression through Msi1-6IE.
Rfx is known as a master transcription factor for the development of both primary cilia and motile cilia. The loss of a Rfx transcription factor known as Daf-19 in Caenorhabditis elegans causes the absence of cilia [47], and Rfx2 and Rfx3 are broadly required for the proper development of cilia in mammals [48,49]. Rfx4 has been reported to be a key regulator of primary cilia development that modulates sonic hedgehog signaling in the neural tube [29,50]. The lack of Rfx4 transcripts or the functional disruption of Rfx4 (Rfx4 L298P) lead to dorsal midline defects in the forebrain, hydrocephalus, and patterning defects in the telencephalon [28,29]. Because the Msi1 knockout mouse also shows hydrocephalus, there might be a relationship with the phenotype of the Rfx4-disrupted mouse. Although the functional role of Rfx transcription factors in NS/PCs has not been reported, primary cilia are required for the expansion and establishment of postnatal hippocampal progenitors [50 –52]. Thus, it is speculated that Rfx transcription factors contribute to the regulation of NS/PCs regarding their maintenance or differentiation. Interestingly, Rfx binding site was enriched at H3K4 me1 positive enhancers and Sox2-Brn2 co-occupied regions in NS/PCs [15,53]. We quantified the mRNA expression of Rfx in NS/PCs; Rfx1, 3, 4 and 7 were strongly expressed in NS/PCs. Rfx1 is ubiquitously expressed in brain, however, it transcriptionally represses neuronal marker MAP1A gene in non-neuronal cells but not in neuronal cells, and Rfx3 also shows repressive regulation of MAP1A in non-neuronal cells [54]. This indicates Rfx transcription factors function redundantly and in a cell type specific manner. Furthermore, both Rfx1 and Rfx3 were recently suggested to regulate the transcription of GPR56 in NS/PCs, and the deletion of Rfx binding sites in GPR56 E1m promoter correlated with pathogenesis of polymicrogyria in entire cortex through putative decreased proliferation of NS/PCs and reduced neuronal migration [55]. Therefore, Rfx transcription factors are good candidates for the regulation of NS/PCs. Our present study revealed that Rfx transcription factors were involved in NS/PC marker expression, and this insight will be applied to the functional analysis of these genes in NS/PCs.
Footnotes
Acknowledgments
We thank Suyama Satoshi and Masato Yano for provision of the materials and technical advice. This work was supported by a Grant-in Aid for Young Scientist (B) (2012–2014, 24700334) from the Ministry of Education, Culture, Sports, Science, and Technology (MEXT) of Japan, and the Funding Program for World-leading Innovative R&D on Science and Technology to H.O.
Author Disclosure Statement
H.O. is a scientific consultant of San Bio, Inc. and Daiichi Sankyo Co., Ltd. The remaining authors have no conflicts of interest to declare.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.