
Abstract
Human and mouse endometrium undergo dramatic cellular reorganization during pregnancy and postpartum. Somatic stem cells maintain homeostasis of the tissue by providing a cell reservoir for regeneration. We hypothesized that endometrial cells with quiescent properties (stem/progenitor cells) were involved in the regeneration of the endometrial tissue. Given that stem cells divide infrequently, they can retain the DNA synthesis label [bromodeoxyuridine (BrdU)] after a prolonged chase period. In this study, prepubertal mice were pulsed with BrdU and after a 6-week chase a small population of label-retaining stromal cells (LRSC) was located primarily beneath the luminal epithelium, adjacent to blood vessels, and near the endometrial–myometrial junction. Marker analyses suggested that they were of mesenchymal origin expressing CD44+, CD90+, CD140b+, CD146+, and Sca-1+. During pregnancy, nonproliferating LRSC predominately resided at the interimplantation/placental loci of the gestational endometrium. Immediately after parturition, a significant portion of the LRSC underwent proliferation (BrdU+/Ki-67+) and expressed total and active β-catenin. The β-catenin expression in the LRSC was transiently elevated at postpartum day (PPD) 1. The proliferation of LRSC resulted in a significant decline in the proportion of LRSC in the postpartum uterus. The LRSC returned to dormancy at PPD7, and the percentage of LRSC remained stable thereafter until 11 weeks. This study demonstrated that LRSC can respond efficiently to physiological stimuli upon initiation of uterine involution and return to its quiescent state after postpartum repair.
Introduction
T
Somatic stem/progenitor cells can be quiescent or slow cycling when situated in their specific stem cell niche [17]. Somatic stem cells maintain tissue homeostasis by acting as a cell reservoir for tissue repair and regeneration [18,19]. A well-established technique for understanding the stem/progenitor cells and their microenvironment is the label-retaining cell (LRC) approach. LRCs are cells that retain a DNA synthesis label after a prolonged chase period. Rapidly dividing cells, such as transit-amplifying cells, dilute the label through cell divisions. An alternative explanation for LRCs is that a stem cell selectively transmits one DNA strand of each chromosome to a daughter stem cell, while the newly synthesized DNA strands are inherited by the other daughter cells committed to differentiation, which will eventually be moved out of the tissue compartment [20,21]. The LRC approach has identified somatic stem/progenitor cells in various tissues, including the endometrium [15,22 –24].
Proliferation of endometrial epithelial LRCs and some stromal LRCs from cycling mice can be induced by estrogen, consistent with their proposed role in endometrial regeneration [15]. A functional response of endometrial epithelial LRCs upon endometrial repair was also observed in an induced decidualization, breakdown, and repair mouse model [25,26]. Nonetheless, the role of endometrial LRCs in the remodeling processes during pregnancy and postpartum remains largely unknown. In this study, we sought to identify and study the temporal change in the proportion of LRCs in the gestational and postpartum mouse endometrium, an essential step toward understanding the involvement of putative stem cells in these remodeling events. Our findings revealed that a small population of endometrial stromal cells retained the bromodeoxyuridine (BrdU) label after a 6-week chase. Therefore, they were termed as label-retaining stromal cells (LRSC). These LRSC are in a quiescent state before and after extensive remodeling, but respond efficiently to stimuli upon initiation of uterine involution. The mobilization of LRSC is associated with activation of β-catenin.
Materials and Methods
Animal and housing condition
Mice were obtained from the Laboratory Animal Unit at The University of Hong Kong. All procedures conducted in this study were approved by the Committee on Use of Live Animals in Teaching and Research, The University of Hong Kong, Hong Kong. Mice were housed under standard laboratory conditions with a 12-h light/12-h dark cycle and free access to food and water.
Study design
The experimental setup is outlined in Figure 1A. Day 19 prepubertal C57BL/6J female mice were administered with BrdU intraperitoneally (50 μg/g of body weight; Sigma-Aldrich) twice daily for four consecutive days and allowed to grow without further labeling. After sexual maturity age, the mice were mated with >6-week-old fertile C57BL/6J male mice and pregnancy was confirmed by the presence of a vaginal plug. Three mice were sacrificed 4 h after the last BrdU injection (0-week chased) to determine the initial labeling index. Other mice were sacrificed at the following time points: 5-week chased (prepregnant, n=3), 5+-week chased [gestational day; (GD)4, n=3], 6-week chased (GD7, n=4), 7-week chased (GD14, n=3), 8-week chased [postpartum day (PPD)1, n=13], 8+-week chased (PPD3, n=4), 9-week chased (PPD7, n=3), 10-week chased (PPD14, n=3), and 11-week chased (PPD21, n=4,). Virgin age-matched mice were scarified at 5-week chased (n=3), 5+-week (n=3), 6-week chased (n=3), 7-week chased (n=3), 8-week chased (n=3), 8+-weeks (n=3), 9-week chased (n=3), 10-week chased (n=3), and 11-week chased (n=3) as controls. Mice were euthanized and the uterine horns were collected, fixed overnight with 4% paraformaldehyde, processed into paraffin blocks by standard technique, and analyzed for BrdU-labeled nuclei by immunohistochemistry and immunofluorescence (IF).

Quantitative analysis of BrdU-labeled cells in gestational and postpartum endometrium. ), prepregnancy (as denoted by ψ), GD: 4, 7, 14 (as denoted by 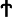 ), and PPD: 1, 3, 7, 14, 21 (as denoted by #).
), and PPD: 1, 3, 7, 14, 21 (as denoted by #).
BrdU immunohistochemistry
Paraffin sections (5 μm) were dewaxed in xylene, rehydrated in descending alcohol series and finally in water, underwent antigen retrieval using an antigen retrieval buffer (Dako) in a microwave oven, denatured with 0.1 N HCl for 45 min at room temperature to allow the access of antibodies to single-stranded DNA, and quenched in 3% hydrogen peroxide/methanol for 10 min. Subsequently, the slides were blocked with 10% donkey serum (Sigma-Aldrich) in phosphate-buffered saline (PBS) for 1 h to reduce nonspecific binding, followed by successive incubation with sheep anti-BrdU antibody (1:800 dilution; Abcam) at 4°C overnight, polyclonal donkey anti-sheep biotinylated secondary antibodies (Abcam) for 1 h, and Vectastain ABC reagent (Vector Laboratories) for 30 min. The sections were examined under a Zeiss Axioskop II microscope (Carl Zeiss) for color development with the substrate 3, 3′-diaminobenzidine (Dako). Nuclei were counterstained with Mayer's hematoxylin for 30 s and washed with distilled water. The slides were mounted with the aqueous mounting medium (Dako) and images were captured using a Photometrics CoolSNAP digital camera (Roper Scientific). All incubations were performed at room temperature unless otherwise specified and washes with PBS were conducted between each step.
Dual IF staining
Paraffin sections were deparaffinized, rehydrated, antigen retrieved, denatured as described above, and blocked with 2% bovine serum albumin (BSA) in PBS for 30 min. For dual IF staining, the two primary antibodies were coincubated in 2% BSA at 4°C overnight in a humidified chamber. Secondary antibodies according to the host of the primary antibodies were diluted in 2% BSA and incubated for 1 h. For triple staining, anti-BrdU staining was performed first, followed by incubation of the appropriate primary antibodies at 4°C overnight and subsequently the corresponding secondary antibodies the next day. Nuclei were counterstained with 4,6-diamidino-2-phenylindole dihydrochloride (DAPI 1:1,000; Invitrogen Life Technologies) for 5 min and mounted with a fluorescence mounting medium (Dako). Multispectrum fluorescence images were acquired using a Carl Zeiss LSM 700 inverted confocal microscope and Zeiss LSM ZEN 2010 software (Carl Zeiss) at the University of Hong Kong Core Facility.
Antibodies
The dilution and details of the primary and secondary antibodies used for IF protocols are presented in Supplementary Tables S1 and S2 (Supplementary Data are available online at
Enumeration of BrdU-labeled cells
For determination of BrdU+ cells, the cells were counted in a blinded manner on one section of each transverse and longitudinal section from a single uterine horn from 3 to 5 mice per group for each time point. Images of the entire mouse uterus were captured by a digital camera (Roper Scientific). BrdU+ cells and unstained (BrdU−) cells in epithelial and stromal compartments were counted using ImageJ software (NIH Image; National Institutes of Health)—At least 10,000 nuclei per uterine horn per mouse at each time point. Only whole nuclei-stained BrdU cells were counted as labeled cells. Those that had the speckled BrdU appearance were not considered as LRSC. The percentage of BrdU+ cells was determined by dividing the number of BrdU+ cells by the total number of nuclei counted in each section.
Enumeration of cell surface marker and BrdU coexpressing cells
Double immunofluorescent-stained slides were viewed under the Carl Zeiss LSM 700 inverted confocal microscope, and the number of BrdU+Marker+ cells was counted using ImageJ software. The percentage of colocalizing cells was determined from two longitudinal and transverse sections from four BrdU-labeled mice at 12 weeks of chase. The percentage of BrdU+Marker+ cells was calculated by dividing the number of BrdU+Marker+ cells by the total number of nuclei counted in each section. The percentages are presented as mean±standard error of the mean (SEM).
Statistical analysis
Data were analyzed using SPSS for Windows (version19.0; SPSS, Inc.). After testing for normal distribution using the D'Agostino-Pearson Kolmogorov-Smirnov test, statistical analysis was performed using Student's t test for comparisons of two independent groups. One-way analysis of variance followed by Fisher's least significant difference and Student–Newman–Keuls multiple comparisons post hoc tests were used for analysis of more than two independent groups. A difference with P-value<0.05 is considered as significant. Data are represented as mean±SEM.
Results
Gestational and postpartum endometrium contains label-retaining cells
To determine whether stem/progenitor cells were present during endometrial remodeling, we assessed the distribution of label-retaining cells in gestational and postpartum endometrium. Figure 1B shows the labeling index of BrdU in different chase periods. On day 0, after the labeling of prepubertal mice was completed, the BrdU labeling index was 66.8±18.6% for epithelial and 44.5±7.5% for stromal cells (Fig. 2A). Following a chase period of 5 weeks, only a few BrdU+ epithelial cells were detected in the luminal epithelium (0.21±0.28%, Fig. 2B) and were undetectable during gestation (Fig. 2C, D).
Localization of label-retaining cells in
The percentage of BrdU+ stromal cells also declined rapidly from 0- (∼45%) to 6-week chase (GD7: ∼1.4%, Fig. 2C, P<0.001, Fig. 1B), but remained steady until parturition (Fig. 2D, E). Between PPD3 and PPD7, another significant decline of BrdU+ cells occurred in the stromal compartment (PPD3: 0.88±0.06%, Fig. 2F and PPD7: 0.28±0.06%, Fig. 2G, P<0.05, Fig. 1B). The percentage of BrdU+ stromal cells was stable from the 9-week chase onward (PPD7, Fig. 2G and PPD14, Fig. 2H) and only a small proportion of labeled cells remained by the 11-week chase (PPD21: 0.41±0.14%, Fig. 2I).
The percentage of LRSC decreases after parturition
To determine whether the endometrial stem/progenitor cells contributed to the remodeling events after pregnancy, we assessed the distribution of labeled-retaining cells in age-matched virgin mice. Following a chase period of 6 weeks, the normal cycling endometrium in virgin mice contained a small percentage of BrdU-labeled stromal cells (1.19±0.58%) and remained unchanged at the 11-week chase (0.96±0.43%, Figs. 1B and 2J–L). A similar percentage of BrdU+ stromal cells was found in the gestational endometrium of the same chased period (GD7: 1.36±0.39%; P=0.763). We termed these stromal cells after a 6-week chase as LRSC. For virgin females, or those who had undergone gestation/postpartum remodeling, the locations of LRSC were found in the following three regions: beneath the luminal epithelium (Fig. 2F, H, J), adjacent to blood vessels (Fig. 2E, I), and near the endometrial–myometrial junction (Fig. 2H, I, L). Interestingly, we noticed that the LRSC at the 11-week chase were significantly higher in age-matched virgins compared with mice that had undergone gestation and postpartum remodeling (0.96±0.43% vs. 0.41±0.14%, respectively, P<0.05, Fig. 1B).
Surface phenotype of LRSC
Given that mouse endometrial stem/progenitor cells had no known cell surface markers, we evaluated their expression with a series of different surface markers (Table 1). Double IF of LRSC at PPD21 demonstrated that these cells express CD140b (Fig. 3A), CD146 (Fig. 3B), CD44 (Fig. 3C), CD90 (Fig. 3D), Sall4 (Fig. 3E), and Sca-1 (Fig. 3F), and are negative for ABCG2 (Supplementary Fig. S2A) and Musashi-1 (Supplementary Fig. S2B). There were wide variations in the expression pattern of these markers, ranging from 1.8% to 72% of the LRSC population (Table 1; Fig. 3G and Supplementary Fig. S3).

Phenotypic characterization of LRSC with various surface markers. Immunofluorescence images showing the LRSC and their colocalization (yellow arrow) with
LRSC, label-retaining stromal cells.
LRSC reside in the interimplantation loci of the endometrium during gestation
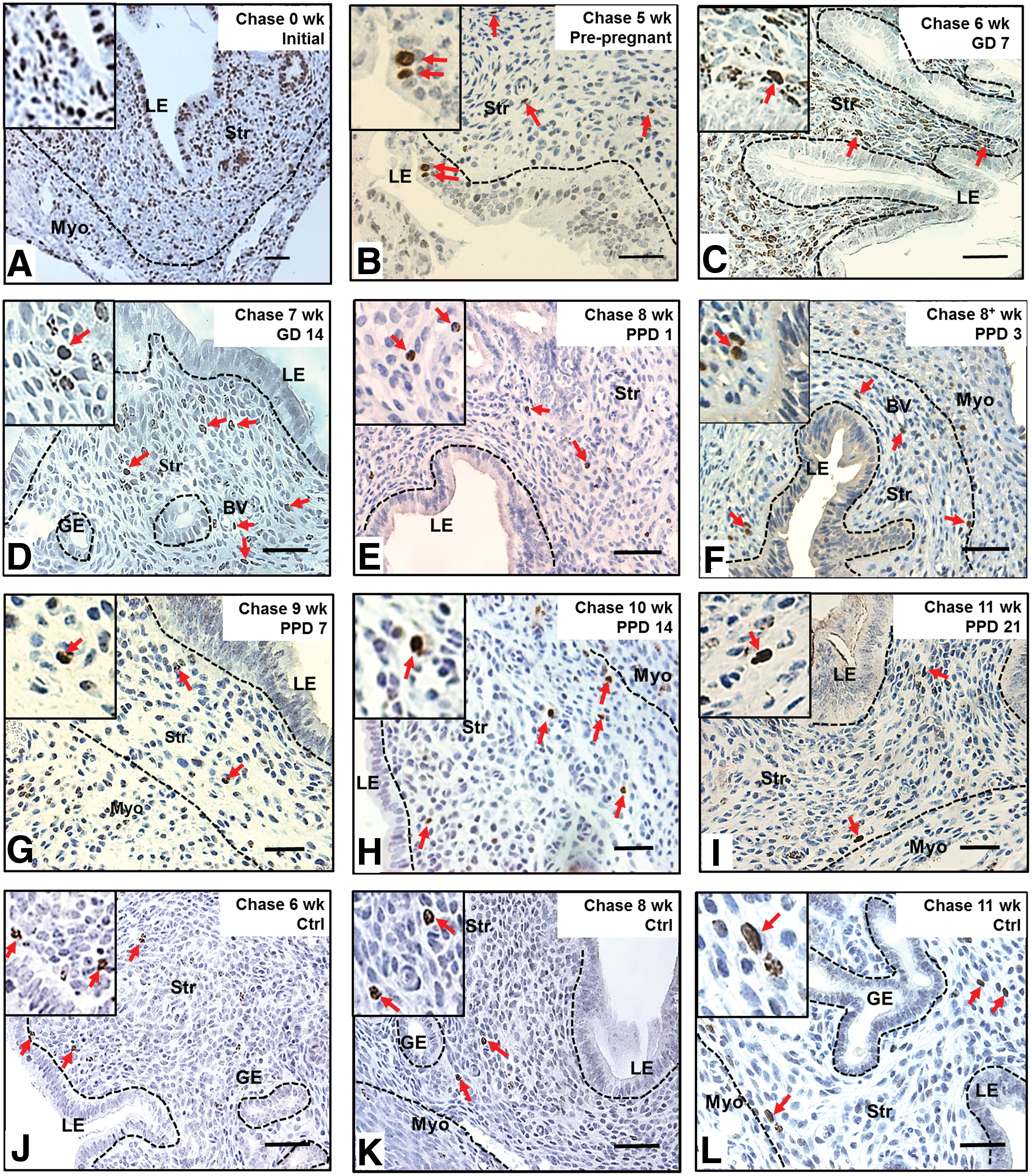
The gestational endometrium is characterized by distinct implantation sites (Fig. 4A). Given that connexin 43 is a widely used implantation marker in murine models and localized distinctively in implantation/placental loci [27 –30], this marker was used to assess the spatial distribution of LRSC during gestation (GD7, 14) and the early postpartum (PPD1) period. The connexin 43 expression was restricted to the implantation/placental loci of the gestational endometrium (Fig. 4C[ii]) and commonly associated with distinct large decidualized cells depicted by their round nucleus (Fig. 4D[i], [ii]).
Spatial distribution of LRSC within the pregnant and early postpartum endometrium.
Figure 4B revealed that the LRSC were prominently enriched in interimplantation/placental loci of the GD7 (Fig. 4D[i]), GD14 (Fig. 4D[iii]), and PPD1 endometrium (Fig. 4D[v]). Significantly more LRSC resided within the interimplantation/placental loci (Fig. 4D[i], [iii], [v]) compared with the implantation/placental (Fig. 4D[ii], [iv], [vi]) loci (GD7: 18.31±2.04% vs. 0.84+0.16%, P<0.01; GD14: 12.43±0.65% vs. 0%, P<0.05; PPD1: 3.23±0.45% vs. 0.54±0.23%, P>0.05, Fig. 4B). Although LRSC were exclusively located in the interimplantation/placental loci of the endometrium at GD14, these cells soon distributed throughout the endometrium by PPD1 (Fig. 4[v], [vi]) and PPD3 after parturition (data not shown).
LRSC proliferate upon parturition
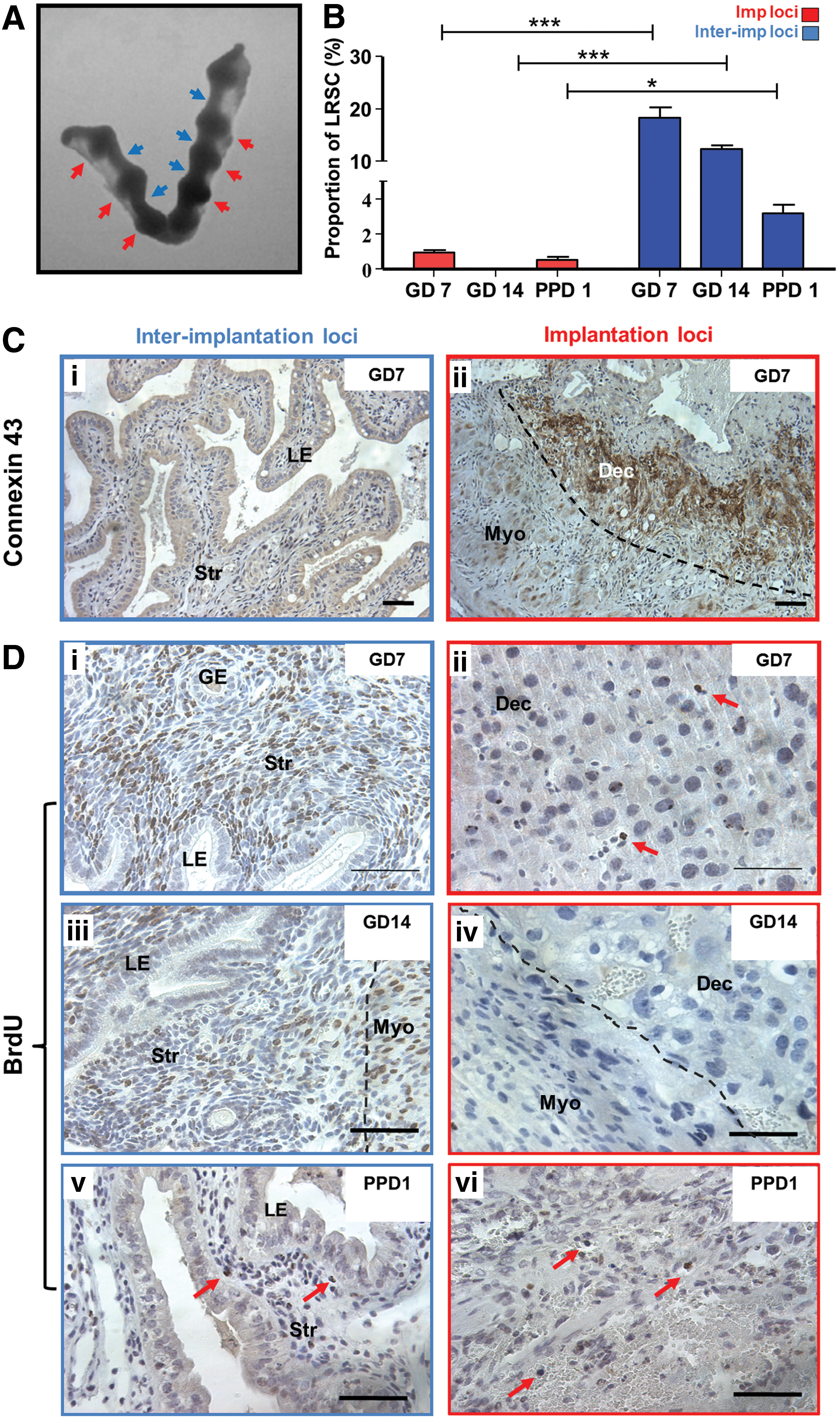
We assessed the ability of LRSC to proliferate in response to stimuli during gestation and postpartum. Figure 5A shows the distribution of proliferating LRSC (Ki67+/BrdU+) in the gestational (GD14) and postpartum endometrium (PPD1, 3). At the midgestational period (GD7 and 14) no proliferating LRSC were detected, suggesting that they were quiescent during this period (Fig. 5A[i], B). Dual IF revealed that the colocalization of Ki67 with BrdU was restricted to the endometrium at PPD1 (Fig. 5A[ii]) and PPD3 (Fig. 5A[iii]). The highest percentage of proliferating LRSC occurred at PPD1 (43.89±5.36%) and declined significantly at PPD3 (18.20±0.83%, P<0.05, Fig. 5A). Thus, LRSC were immediately active after parturition and their proliferative ability declined thereafter. Aside from proliferating LRSC, some nonproliferating LRSC persisted (Ki67−/BrdU+, Fig. 5A[ii], [iii]) during postpartum. Since proliferating LRSC were detected at PPD1 and PPD3, this resulted in the lower proportion of LRSC detected at PPD7 (PPD3: 0.88±0.06% vs. PPD7: 0.28±0.06%, P<0.05, Fig. 1B) and a higher amount of LRSC present in virgin mice at the 11-week chase (PPD21: 0.96±0.43% vs. virgin: 0.41±0.14%, P<0.05, Fig. 1B).
Proliferation of LRSC.
Activated LRSC expressed the Wnt/β-catenin signal pathway components
The Wnt/β-catenin signaling pathway plays an important role in stem cell self-renewal in vivo and in vitro [31,32], asymmetric cell division of stem cells [33], and is a key mediator of human endometrial homeostasis and proliferation [34]. Therefore, we investigated the expression of total β-catenin and its active form in LRSC. The expression of total β-catenin (Fig. 6A) and active β-catenin (ABC) (Fig. 6B) on LRSC at PPD1 was 34.50±13.17% and 7.95±4.64%, respectively (Fig. 6C). In virgin mice, 14.17±11.76% of LRSC expressed total β-catenin and no LRSC expressed ABC (Fig. 6C). Moreover, higher proportions of LRSC at PPD1 coexpressed with total β-catenin and ABC than other gestational and PPDs (Fig. 6D, E). Notably, 9.95±1.89% of LRSC coexpressed with Ki67 and total β-catenin (Fig. 6F, G[i]), and 5.27±0.91% of LRSC coexpressed with Ki67 and ABC at PPD1 (Fig. 6F, G[ii]). The coexpression of Ki67 with total β-catenin or ABC was not detected in virgin mice (Fig. 6F).
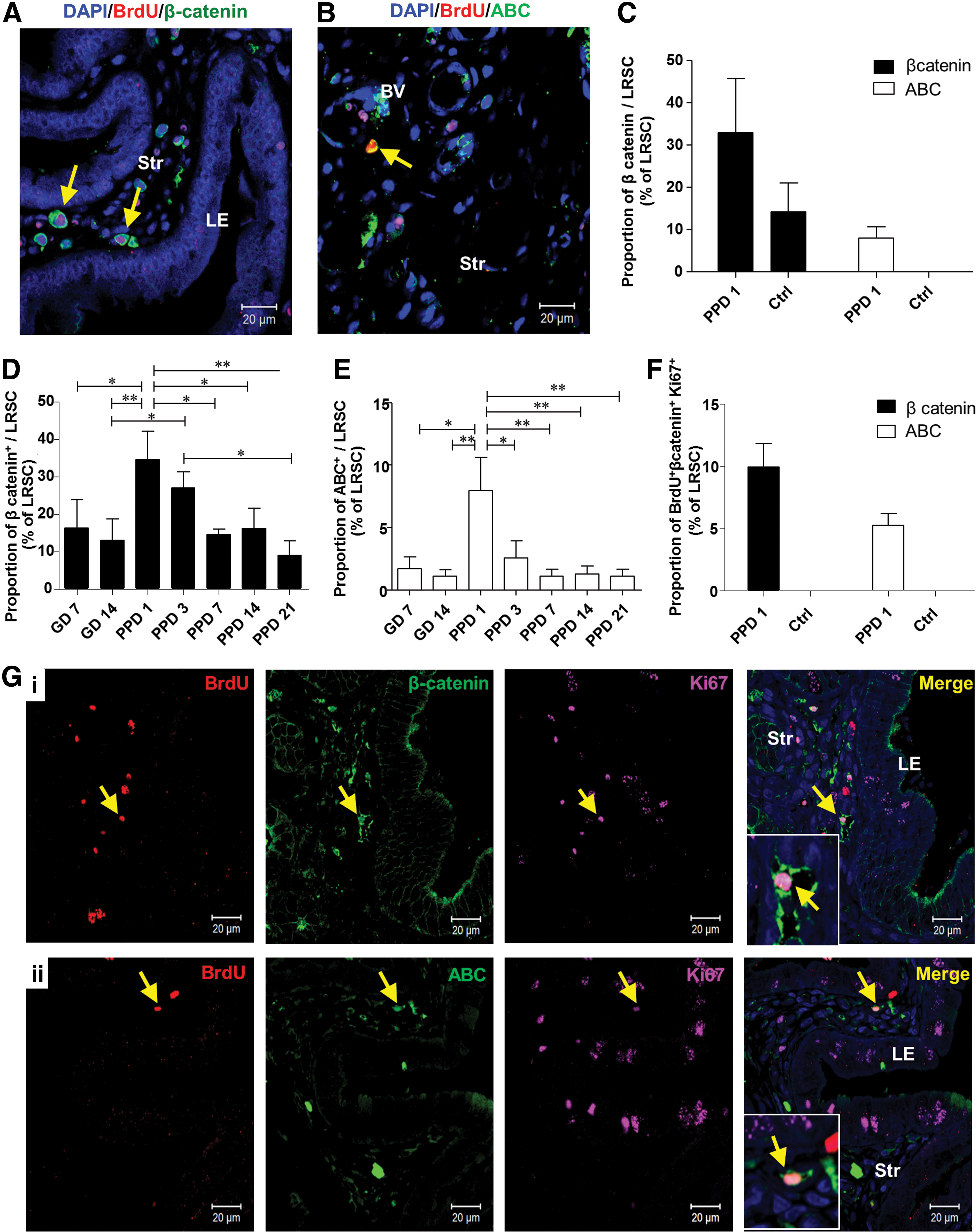
Proliferating LRSC express total β-catenin and ABC. Representative immunofluorescence images show LRSC at PPD1 colocalizing with
Identification of endometrium epithelial BrdU+ cells after parturition
The high cellular turnover of epithelial cells at gestation resulted in depletion of epithelial BrdU+ cells from the 5+-week chase (GD4, Fig. 1B). Interestingly, at PPD1, a few epithelial cells with strong nuclei BrdU staining were observed in the luminal epithelium (Fig. 7A, B) and none in the glandular epithelium. The time of tissue collection after parturition was a crucial factor for the identification of these epithelial BrdU+ cells. Of the 10 mice analyzed at PPD1, only two mice harvested within 6 h after parturition contained occasional epithelial BrdU+ cells.
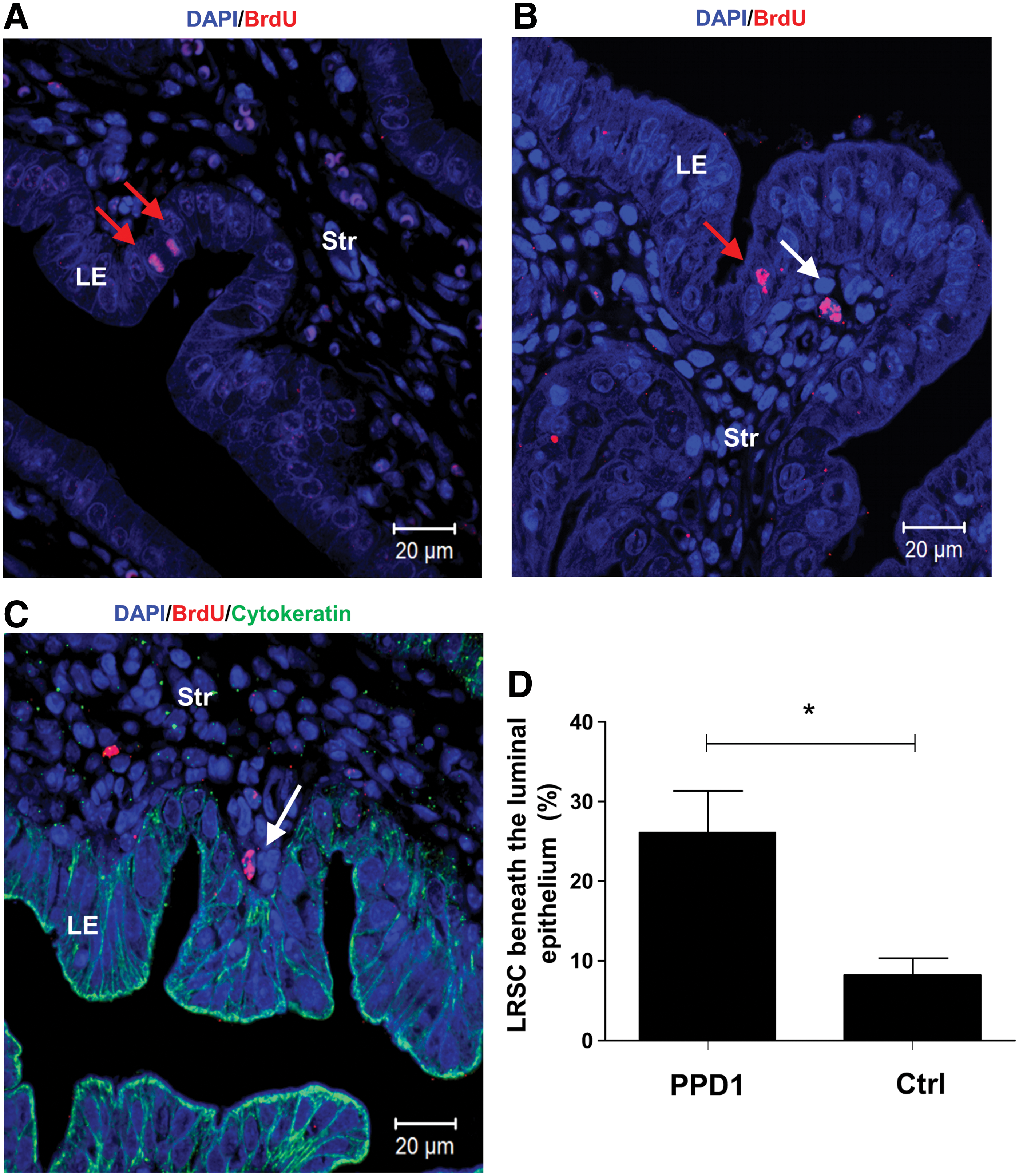
Identification of epithelial BrdU+ cells and location of LRSC on PPD1.
The reappearance of epithelial BrdU cells in the luminal epithelium, led to the investigation of surrounding LRSC at PPD1. We found postpartum females containing a higher proportion of LRSC beneath the luminal epithelium (within one-cell diameter) compared with age-matched virgin mice (26.13±9.08% vs. 8.20±4.36%, P<0.05, Fig. 7B–D).
Discussion
Adult stem cells are rare specialized cells that self-renew and differentiate to supply a progeny of lineage-specific cells for cellular replacement after injury-like events [35]. Despite the identification of stem cells in endometrial homeostasis, the roles of these cells in highly dynamic states such as pregnancy and postpartum remain unclear. Using the BrdU pulse-chase experiment, this study investigated the in vivo location of LRSC and their possible role in uterine regeneration. If all endometrial cells divide evenly during uterine remodeling, this would cause homogenous dilution and eventual loss of the label, like the pancreas [36]. Instead, the identification of cells retaining BrdU label after an 11-week chase in the virgin females indicates some endometrial cells divide much less frequently than the surrounding cells or may have undergone asymmetric division. In addition, we demonstrate the functional activity of LRSC in response to parturition stimuli.
LRSC in the gestational and postpartum endometrium
The mouse endometrium is a highly renewing adult tissue in mammals and serves as an excellent model for studying the role of tissue stem/progenitor cells in response to local niche stimuli. In an artificial mouse endometrial repair model, LRSC near vasculatures proliferate upon tissue breakdown [25]. On the other hand, LRCs enriched at the distal oviduct and the endocervical transition zone did not respond to endometrial regeneration events [37]. These findings support the existence of a functional stem cell compartment in the mouse endometrium. Since, decidualization and postpartum involution are hallmarks of endometrial remodeling events [4], we examined the location of the LRSC during pregnancy and at postpartum.
As expected, the extensive structural and functional differentiation during early pregnancy resulted in a significant decline of LRSC. In this study, the percentage of BrdU+ cells in the virgin mice and the pregnant mice dropped within the duration of the chase period and reached the same stable level after a 6-week chase. We termed these cells LRSC, and they could represent the endometrial stromal stem/progenitor cell population.
Although the LRC approach has been widely used to study the distribution and functions of putative stem cells in different adult tissues, it is not specific for stem cells. Arrest cells in vivo may also retain the BrdU label, hence producing false-positive errors. Since our LRSC were able to proliferate upon specific stimuli, it is unlikely that such a phenomenon occurred in this study.
The histological and morphological appearance of the uterine horns returned to its nonpregnant state in less than 1 week after parturition. Hence, after completion of endometrial remodeling only 0.4% of LRSC remained at PPD21 and resided in regions reported in cycling mice, that is, beneath the luminal epithelium, around the blood vessels, and near the myometrium–endometrium junction [15].
The phenotypic characterization of LRSC
Several evidences suggest that endometrial repair can trigger recruitment of endometrial stem/progenitor cells from the bone marrow through vasculature [25,38]. Hence, we examined three markers (CD44, CD90, and Sca-1), which have been reported to be expressed on mesenchymal stem-like cells in the female reproductive system, including the human endometrium [39], myometrium [14], menstrual blood [40], and mammary glands [41]. The percentage of LRSC expressing CD44+ and CD90+ were 23.72±2.25% and 45.26±12.88%, respectively, suggesting that a significant proportion of the endometrial stem/progenitor cells could be of mesenchymal origin. Interestingly, 72.04±15.01% of the LRSC were Sca-1+, a marker used to enrich hematopoietic stem cells, mammary stem/progenitor cell, and other mesenchymal stem cells [42]. Previous work demonstrated that LRSC in the cycling endometrium, myometrium, and distal oviduct lack Sca-1 expression [43,44]. However, endometrial postpartum cells with a side population phenotype showed abundant expression of Sca-1 [45]. The discrepancies in these observations may be related to difference in the studied cell populations (eg, LRSC vs. side population) and/or physiological state of the animals (eg, estrus cycling vs. postpartum). In general, decidualized endometrial stromal cells express abundant amount of Sca-1 during implantation [46].
In the human endometrium, stromal cells coexpressing perivascular markers, CD146 and CD140b, exhibit stem cell-like activity based on in vitro assays and differentiation ability [47]. About half of the LRSC expressed CD140b (44.51±2.33%), but the CD146 expression was very low (1.81±0.37%). Hence, we speculate that the putative stem/progenitor cell population in mice may not share the same phenotype as in humans [47]. This aside, we also evaluated several cellular markers related to stem cells, but were unable to identify a marker signature that uniquely identifies LRSC in the mouse endometrium. Thus, there is still no specific marker for mouse endometrial stem/progenitor cells.
Environmental niches and LRSC
During pregnancy
Depending on the environmental cues, niche cells can maintain stem cells in an undifferentiated state or mobilize resident stem cells to function [48,49]. During pregnancy, nonproliferating LRSC preferentially localize within the interimplantation sites during the late gestational stage, suggesting that these regions maintain the LRSC in a quiescent state. The lack of cell proliferation regulator genes (cyclin D1, D2, and D3) in the interimplantation sites in comparison with the implantation sites [50] supports that these sites are relatively dormant. Such a microenvironment would be critical for maintaining a reservoir of LRSC after each successive pregnancy.
Postpartum remodeling
The regeneration of the endometrium occurs immediately after parturition [51,52]. Therefore, it was not surprising that a proportion of the LRSC proliferated soon after parturition (PPD1). We speculate that the LRSC are activated to reorganize the endometrium after the birth of newborns. The distinct proliferating LRSC subset identified after parity may correspond to the repopulating cells. A similar observation occurs in the mammary compartments [53]. To protect stem cell exhaustion, these cells exhibit increased cell proliferation and differentiation, but are able to return to dormancy through inhibitory signals from the niche environment. Such a functional response of the stem cells to physiological and pathological stimulations is a common phenomenon in a variety of tissues, including the skin [48,54,55], intestine [49,55,56], mammary gland [41], and pancreas [57]. These adult stem cells must be mobilized periodically to generate cells committed to replenish certain cell lineages [58]. Bulge LRCs expelled from their niche, migrate upward [55], and proliferate to repair the damaged skin epidermis [59]. Hematopoietic stem cells respond readily to injuries and reversibly switch to dormancy after reestablishment of homeostasis [60]. In the rat kidney, papillary LRCs migrate toward other regions of the kidney, and proliferate after acute kidney injury [22]. In the mouse ovary, H2B-GFP LRCs are quiescent before ovulation and proliferate upon estrous cycling changes [23]. In this study, we demonstrate that certain LRSC at the interimplantation/placental loci during pregnancy can facilitate the endometrial remodeling after parturition.
Wnt/β-catenin signaling pathway involvement in the activation of LRSC
The role of the Wnt/β-catenin signaling pathway on stem cell function is well documented. Habib et al. [33] demonstrated that Wnt directed the mitotic division plane and oriented the asymmetric distribution of the centrosome in embryonic stem cells. Lowry et al. showed β-catenin acted directly on the stem cells of the hair follicle to induce their mobilization or exit from a quiescent state to provide proliferating transit-amplifying cells [61]. In mouse mammary stem cells, the Wnt protein can promote their long-term expansion [62].
In this study, we found greater expression of total β-catenin and active β-catenin in LRSC at PPD1 compared with the virgin age-matched mice. Such a phenomenon was only detected after partition and not in other gestation and postpartum periods. This finding provides the first evidence on the possible role of the Wnt/β-catenin signaling pathway on the activation of putative stem/progenitor cells in the mouse endometrial regeneration. Further investigation is necessary to understand the mechanisms of the Wnt/β-catenin pathway on endometrial stem/progenitor cell activation.
LRSC may facilitate the regeneration of the epithelium during the postpartum period
After parturition and expulsion of the placenta, the epithelial basement membrane is completely lost in the first 24 h. An incomplete layer of epithelial cells and stroma containing numerous microvasculatures reappears by 48 h. These observations reflect that the regeneration takes place immediately and rapidly after parturition. In this study, no epithelial BrdU+ cells were observed during pregnancy. At PPD1, a few epithelial BrdU+ cells reappeared in the luminal epithelium. The number of LRSC beneath the luminal epithelium in these mice was significantly more compared with the virgin mice. Previously, using genetic fate-mapping techniques, Huang et al. proved that a proportion of stromal cells differentiates into both luminal and glandular epithelial cells after parturition and contributes to the reepithelialization process [63]. Our observations, together with other reports, provide evidences that the endometrial epithelial cells in mice could be derived from the stromal cells during endometrial regeneration [63,64].
We speculate that the transformation of LRSC into epithelial cells requires mesenchymal–epithelial transition (MET). Patterson et al. [64] demonstrated MET in epithelial repair after decidualization. Notably, cells in the transitional state between the mesenchyme (vimentin+) and epithelium (pan-cytokeratin+) were found migrating from the endometrium–myometrium junction to the lumen to facilitate the reepithelialization after artificial decidualization. During MET, the mesenchymal cells progressively lose their mesenchymal traits and obtain the epithelial characteristics [65]. In this study, LRSC located near the luminal epithelium did not express cytokeratin, suggesting that these cells are in the initial stages of the MET.
Since, there were abundantly more LRSC locating near the luminal epithelium in the PPD1 endometrium than in virgin mice, we propose that LRSC may have the ability to migrate to the disrupted luminal epithelium and transform into epithelial-like cells for reepithelialization after parturition, suggestive of a dual epithelial/stromal differentiation potential. Interestingly, similar cyclic rupture, repair, and regeneration occur in the ovary after each ovulation. Resident stem cells located in the coelomic epithelium at the sites of the ovulation wound can facilitate repeated reepithelialization processes and display characteristics of epithelia and mesenchyme [23]. These data strengthen our theory that in preparation for the remodeling of the endometrium after giving birth, certain environment cues may elicit surrounding LRSC to proliferate and undergo MET for the reconstitution of the epithelial compartment in postpartum endometrium. However, we cannot exclude the possibility that the epithelial BrdU+ cells may originate from a circulating source such as the bone marrow. Morelli et al. [66] showed bone marrow cells can serve as a cellular source for cyclic replenishment of multiple endometrial cell types. Bone marrow-derived stem cells can transdifferentiate into endometrial epithelial and stromal cells [67]. They also contribute toward the regeneration of the endometrium after ischemia/reperfusion injury [68].
In conclusion, we identified and characterized mouse endometrial LRSC after dramatic physiological remodeling. LRSC primarily located in the interimplantation loci proliferated after parturition and highlight the possible involvement of the Wnt/β-catenin pathway in endometrial stem cell activation. It is essential that future studies examine the role of the Wnt/β-catenin pathway on endometrial stem cell regulation.
Footnotes
Acknowledgments
The authors thank all the staff at the Laboratory Animal Unit and Faculty Core Facility, the University of Hong Kong, for their assistance in this study. This work was supported by a grant from the General Research Fund (GRF), Research Grant Council (HKU 7822/09M) to W.S.B.Y.
Author Disclosure Statement
The authors declare no financial or commercial conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.