
Abstract
A growing body of evidence has revealed that stem-like cells in the posterior limbus of the eye between the corneal endothelium (CE) and trabecular meshwork (TM) may be able to rejuvenate these tissues in disease. However, these cells have not been clearly defined and we have named them PET cells (progenitor cells of the endothelium and trabeculum). A good and inexpensive animal model for PET cells is lacking, so we investigated bovine eyes as an effective large tissue source. We showed the presence of stem/progenitor cells in the bovine CE, transition zone, and TM in situ. Floating spheres cultured from the CE and TM showed similar stem cell marker expression patterns. Both the CE and TM spheres were bipotent and highly proliferative, but with limited secondary sphere-forming capability. They were highly prone to differentiate back into the cell type of their tissue of origin. It is speculated that the PET cells become more tissue-specific as they migrate away from their niche. Here, we showed that PET cells are present in the posterior limbus of bovine eyes and that they can be successfully cultured and expanded. PET cells represent an attractive target for developing new treatments to regenerate both the CE and TM, thereby reducing the requirement for donor tissue for corneal transplant and invasive treatments for glaucomatous patients.
Introduction
B
Accumulating evidence suggests that some stem-like cells reside in the transition zone between the CE and TM at the human posterior limbus. This population may be able to provide new cells for regeneration of the CE, TM, or possibly both. Studies of immunolocalization of stem cell markers in human tissues provide direct evidence for the presence of such stem-like cells in situ. Whikehart et al. [9] detected telomerase activity at the peripheral CE and bromodeoxyuridine (BrdU) labeling in the transition zone and TM. The BrdU staining extended into the CE following experimental mechanical injuries. These findings suggest that stem-like cells in the transition zone may help renew cells in the CE, especially after trauma. McGowan et al. [10] found the expression of stem cell markers nestin, alkaline phosphatase, and telomerase in some cells at the posterior limbus. More stem cell markers including octamer-binding transcription factor (Oct)3/4, paired box gene 6 (Pax6), Wnt1, and sex-determining region Y-related box gene (Sox2) were detected with wounded corneas. He et al. [11] reported that the expression of stem cell markers was largely restricted in the extreme periphery of the CE.
Raviola [12] was the first to describe a population of cells located just beyond a peripheral transition zone called Schwalbe's line in the rhesus monkey eye, which showed different ultrastructural characteristics from typical CE and TM cells. Challa et al. [13] later identified a novel cell type in human primary TM cell culture that highly expressed Ankyrin G (AnkG) and Breast Epithelial Antigen 46 (BA46). Kelley et al. [14] reported exclusive immunostaining of AnkG and BA46 in the human TM insert cells post-laser trabeculoplasty in an organ culture model. Cultured human TM insert cells were found to express BA46 [15]. It was speculated that the Schwalbe's line cells, novel cells, and TM insert cells may be one and the same and represent the putative stem cells in the transition zone at the posterior limbus. In fact, the putative stem cells in the peripheral CE, transition zone, and TM have not been clearly defined. Thus, we have collectively named them PET cells (progenitor cells of the endothelium and trabeculum) [6].
Despite the promising findings in human, there have been no reports on whether these stem-like cells are common to other non-primate species with marked anatomical diversity in the posterior limbus. Published works on the cultivation of CE or TM stem cells have been largely confined to the human species [16 –22]. Given the scarcity of human donor eyes, a supplementary animal model for studying the PET cells would be hugely advantageous. In this study, we examined the comparative anatomy of the human and bovine posterior limbus. We aimed to determine the presence and localization of the stem-like cells in the bovine posterior limbus by immunohistochemistry. We also sought to investigate whether the CE and TM stem/progenitor cells from bovine eyes can be isolated and expanded by the sphere culture technique. We then further characterized the phenotypical, proliferative, self-renewing, differentiation, and functional properties of these cells. Successful identification and isolation of these cells from bovine eyes provides an additional and valuable large tissue model for the study of CE or TM rejuvenation.
Materials and Methods
Human and bovine tissues
Human cadaver eyes (78±11 years old) not suitable for transplantation were acquired from the eye bank of the Royal Liverpool University Hospital with prior ethics approval from the National Research Ethics Service Committee (reference: 01/066) and handled in accordance to the tenets of the Declaration of Helsinki. Bovine eyes (1–2 years old) were obtained from a local abattoir.
Histology and immunohistochemistry (whole mount and section)
Whole eyes or dissected anterior segments (Fig. 1B, G) were fixed with 4% formaldehyde and embedded in paraffin. Sections (5 μm) were stained with hematoxylin and eosin for the anatomical studies [23]. Antigen was retrieved by trypsin (1 mg/mL; Sigma-Aldrich, St. Louis, MO) incubation for 30 min at 37°C, proteinase K (20 μg/mL; Sigma-Aldrich) treatment for 4 min at room temperature (RT), or incubation in Tris-EDTA buffer (10 mM Tris, 1 mM EDTA, 0.05% Tween 20, pH 9.0) for 20 min at 95°C. Primary antibody incubation was carried out overnight at 4°C (Table 1). Stem cell markers included ABCG2, AnkG, nestin, Oct4, Pax6, Sox2, STRO-1, and telomerase. Controls slides were incubated with corresponding isotype controls or normal rabbit IgG at the matched concentrations to the primary antibodies. The filamentous F-actin on whole mount tissues was detected by Alexa Fluor 488-conjugated phalloidin (1:20; Life Technologies, Carlsbad, CA). Vimentin (Table 1) staining of the TM was visualized with the EnVision+System-HRP (AEC) kit (Dako, Carpinteria, CA). The secondary antibodies were Alexa Fluor 488 Goat anti-rabbit IgG, Goat anti-mouse IgG or IgM (Invitrogen, Eugene, OR). Nuclei were counter-stained with 4′,6-diamidino-2-phenylindole (DAPI) (0.1 μg/mL; Sigma-Aldrich) or propidium iodide (PI) (1 μg/mL; Life Technologies). Stained sections were digitally scanned by an upright fluorescence microscope (Eclipse 80i; Nikon, Tokyo, Japan) or a confocal laser scanning microscope imaging system (MRC-600; Bio-Rad, Hemel Hempstead, United Kingdom).
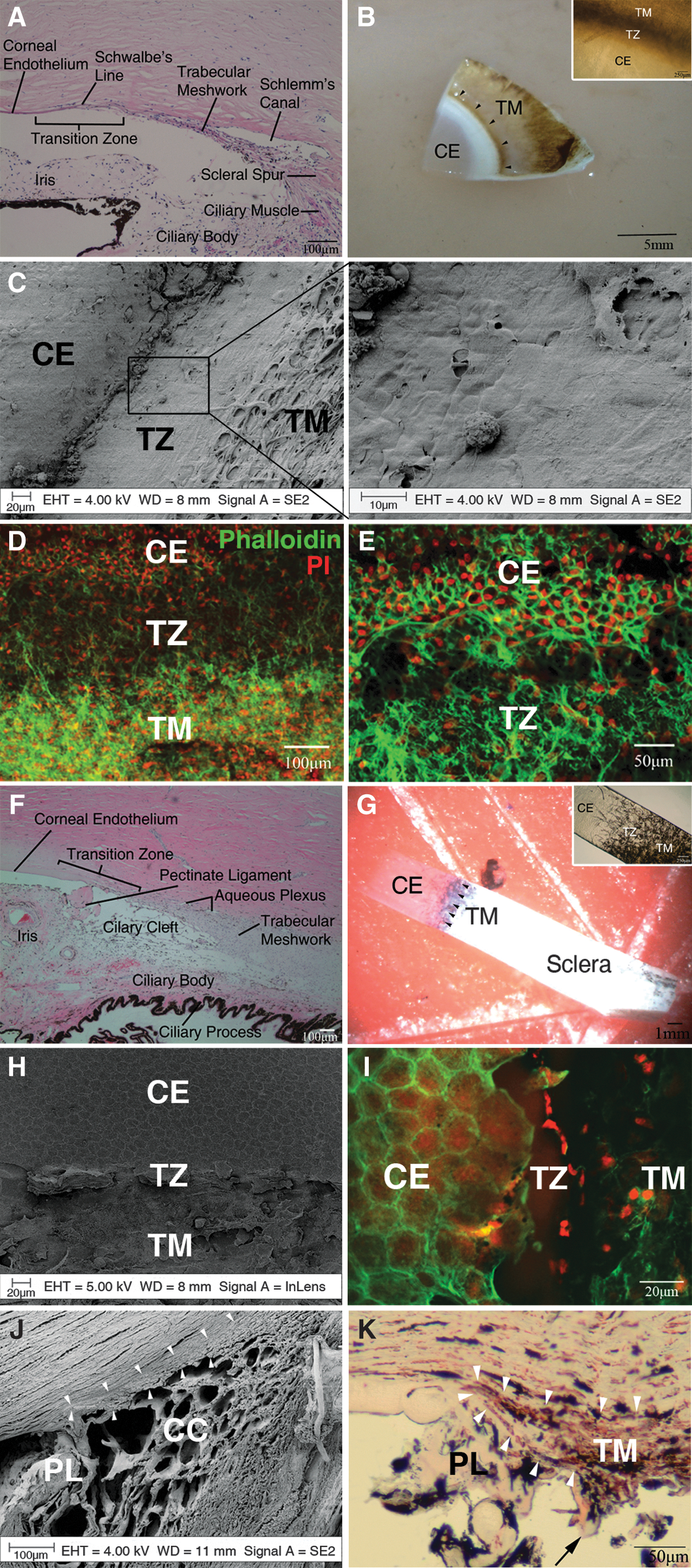
Comparative anatomy of the human and bovine posterior limbus.
ABCG2, ATP-binding cassette sub-family G member 2; AnkG, Ankyrin G; AQP1, aquaporin-1; BrdU, bromodeoxyuridine; CHI3L1, chitinase-3-like protein-1; GFAP, glial fibrillary acidic protein; Oct4, octamer-binding transcription factor 4; Pax6, paired box gene 6; Sox2, sex-determining region Y-related box gene; ZO-1, zonula occludens-1.
Quantitation of immunopositive cells in the transition zone
Montages of the chamber angle were created and analyzed by Photoshop CS5 (Adobe, San Jose, CA). The number of cells in the transition zone (those beyond the end of Descemet's membrane) showing positive immunoreactivity was counted manually. Results were expressed in percentage out of the total number of nuclei in the TM.
Tissue processing for scanning electron microscopy
Anterior segments were fixed in 2.5% glutaraldehyde for 24 h and post-fixed in 1% osmium tetroxide for 1.5 h. The tissues were then dehydrated through ascending concentrations of ethanol, critical point dried (Polaron, East Sussex, United Kingdom) and sputter coated with gold. A LEO 1550 field emission scanning electron microscopy (SEM) was used to observe the surfaces of the specimens with the secondary electron detector (accelerating voltage 3–5 keV, working distance 8–11 mm).
Isolation of primary CE and TM cells
Primary CE and TM cells were harvested respectively as previously described using Accutase (Life Technologies) for 30 min at 37°C [24 –28]. CE cells from the central cornea were harvested using a cloning ring with internal diameter of 12 mm. To harvest peripheral CE cells, the cells within the central 12 mm diameter were scraped off before Accutase incubation. The cells were cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS, Hyclone; Thermo Scientific, Waltham, MA), 1% penicillin–streptomycin (P/S), and 1% amphotericin B (Life Technologies) (DMEM/10% FBS) for 7 days at 37°C in a humidified atmosphere of 5% CO2. Medium was changed every 2 days.
Sphere culture
Primary bovine CE or TM cells were detached from the culture dish by incubation with Accutase for 10 min at 37°C. After centrifugation, the cells were resuspended in serum-free DMEM/F12 medium (1:1; Life Technologies) and dissociated into single cells. Finally, the cells were seeded on suspension culture dishes (Corning, Shanghai, China) at 2,500 viable cells per cm2 as determined by trypan blue exclusion test (Sigma-Aldrich) and cultured in stem cell medium [SCM; DMEM/F12 1:1, 2% B27, 20 ng/mL epidermal growth factor (Peprotech, London, UK), 20 ng/mL basic fibroblast growth factor (Peprotech), 5 μg/mL heparin (Sigma Aldrich), 0.1 mg/mL bovine serum albumin (Affymetrix, Santa Clara, CA), 2.5 mM
Immunocytochemical analysis of primary cells and spheres
Spheres or cell monolayers were fixed in 4% formaldehyde for 15 min, permeabilized with 0.1% Triton X-100 for 5 min, and processed for staining as described for immunohistochemistry. The cells were incubated with Alexa Fluor 488 or 568 secondary antibodies (Life Technologies) and counterstained with DAPI. Images were acquired with a confocal laser scanning microscope (LSM700; Carl Zeiss, Thornwood, NY) or an inverted fluorescent microscope (Eclipse TE2000-S; Nikon).
Proliferative capacity of spheres
The spheres were stained with antibodies against Ki67 and BrdU. For BrdU labeling, spheres were incubated with 10 μM BrdU (Sigma-Aldrich) for 24 h before fixation in acid/alcohol (5 parts acetic acid and 95 parts of 95% ethanol) for 15 min at RT. The number of BrdU-positive cells was quantified with z-stack images (20×; zoom factor 1; step size of 3.5 μm) of the entire spheres using ImageJ (
Quantitative reverse transcription–polymerase chain reaction
Total RNA was extracted by RNAiso Plus reagent (Takara, Tokyo, Japan). At least three biological replicates were included in each group. First-strand cDNA was generated using High Capacity RNA-to-cDNA Master Mix (Life Technologies) in a thermal cycler (TaKaRa PCR Thermal Cycler Dice). Primers were designed using PrimerQuest (
Reverse transcription from RNA to cDNA were carried out for 5 min at 25°C, 30 min at 42°C, 5 min at 85°C, and followed by cooling at 4°C in a thermal cycler (TaKaRa PCR Thermal Cycler Dice). qRT-PCR was performed with StepOnePlus Real-Time PCR System (Applied Biosystems), with the cycling parameters shown as follows: 20 s at 95°C, 3 s at 95°C for 40 cycles, and 30 s at 60°C. Melt curve was generated with the following protocol: 15 s at 95°C, 1 s at 60°C and then 15 s at 95°C. The curves were analyzed to verify the amplification specificity.
qRT-PCR, quantitative reverse transcription–polymerase chain reaction.
Differentiation studies of the spheres
Non-directed differentiation
Cells dissociated from the CE or TM spheres were differentiated in DMEM/10% FBS for 7 days. Medium was changed every 2 days. The differentiated sphere progenies were stained with CE markers aquaporin-1 (AQP1), connexin 43, N-cadherin, Na+/K+ ATPase, and zonula occludens-1 (ZO-1) or TM markers AQP1 and chitinase-3-like protein-1 (CHI3L1) to investigate their differentiation status. For acetylated low density lipoprotein (Ac-LDL) labeling, the cells were incubated with DiI-conjugated Ac-LDL (20 μg/mL; Life Technologies) for 4.5 h at 37°C with 5% CO2 before fixation [29].
Neural differentiation
Cells were seeded at 2.5×104 cells per cm2 on glass coverslips coated with 100 μg/mL poly-
Mesenchymal differentiation
Mesenchymal differentiation of the spheres was carried out using the adipogenic, chondrogenic, and osteogenic differentiation kits (Cyagen, Guangzhou, China) according to the manufacturer's manuals. Mouse mesenchymal stem cells (mMSCs) (Cyagen) were cultured in parallel as a positive control.
Functional tests
Transendothelial electrical resistance measurement
To evaluate the quality of tight junction formation, primary CE cells and spheres were seeded at 2.5×104 cells per cm2 on Snapwell™ inserts (Corning) and cultured in DMEM/10% FBS for 7 days until confluence. The inserts were mounted in the Ussing Chamber (Physiological Instruments, San Diego, CA) for the resistance measurement. Fresh bovine corneas with the epithelium scrapped off were used as positive control. Each side of the chamber was filled with Ringer's buffer solution (pH 7.4) [30].
Melanin phagocytosis assay
To evaluate the phagocytic function of primary TM cells and TM sphere progenies, the cells were seeded at 1×104 cells per cm2 and cultured in DMEM/10% FBS for 7 days. On day 7, they were incubated with 10 μg/mL melanin (Sigma-Aldrich) for 24 h. The cells were then fixed with 4% formaldehyde, washed with 0.1% tween in phosphate-buffered saline, and counterstained with hematoxylin. Five random fields of view (2.3 mm2/field) from each well were photographed. The percentage area of phagocytosed melanin was calculated using ImageJ (
Statistical analysis
Statistical significance was determined by Student's t-test to compare mean values between two groups, or by one-way analysis of variance and Bonferroni's multiple comparison post hoc test to compare mean values between three groups using GraphPad Prism (San Diego, CA). Significant values were defined as P<0.05.
Results
Comparative anatomy of the human and bovine posterior limbus (progenitor cell niche)
The transition zone at the human posterior limbus contains the peripheral Descemet's membrane, Schwalbe's line, and the most anterior portion of the wedge-shaped TM (Fig. 1A). Approaching the transition zone, Descemet's membrane gradually becomes thinner and is continuous with the anterior trabecular beams, which taper and insert into the deeper corneal lamellae just beneath Descemet's membrane.
SEM examination illustrates a smooth zone between the CE and TM in human (Fig. 1C). The anterior border of the transition is covered by flattened polygonal cells, which becomes continuous with the trabeculae at the posterior border. This smooth zone varied from 80 to 130 μm in width and was present in all specimens we studied. Although transitional forms between the two cell types are present, there is generally an abrupt and well-demarcated change from CE to TM cells.
The anterior margin of the human transition zone is demarcated by the irregular outline of the peripheral CE cells (Fig. 1D, E). The zone appears to be an area particularly rich in cells. The posterior border of the transition is bound by the insertions of the anterior TM beams, some of which extend across the transition zone.
Corresponding to the anatomy of the human posterior limbus, the bovine transition zone begins at the peripheral CE and contains the most anterior portion of the TM (Fig. 1F). Near the transition, Descemet's membrane also gradually becomes thinner and is connected to the anterior beams of the TM.
The transition between the bovine CE and TM is even more abrupt than that of the human because there is no obvious smooth zone (Fig. 1H). At the transition, there is an irregular anterior border formed by the peripheral CE cells, followed by remnants of the base of pectinate ligaments and a rapid change into TM architecture (Fig. 1I). However, meridional examination of the transition zone clearly showed the anatomical extensions of the anterior trabecular beams beneath the Descemet's membrane (Fig. 1J, K), which is a distinctive human feature. The trabecular insertion varied between 125 and 350 μm measured from the termination of Descemet's membrane.
Immunolocalization of stem cell markers in the bovine posterior limbus
Expression of ABCG2, nestin, Oct4, Pax6, Sox2, STRO-1, and telomerase were observed in the entire CE (Fig. 2A), isolated cells within the transition zone (Fig. 2B), and the entire TM (Fig. 2C). Positive staining of AnkG, which is a marker for TM insert cells, was exclusively found in the transition zone (Fig. 2D). The immunofluorescent cells in this zone were found in deeper layers just beneath the end of the peripheral CE and seemed to be continuous with the anterior TM. Overall, they comprised ∼1%–5% of the total TM cell population.
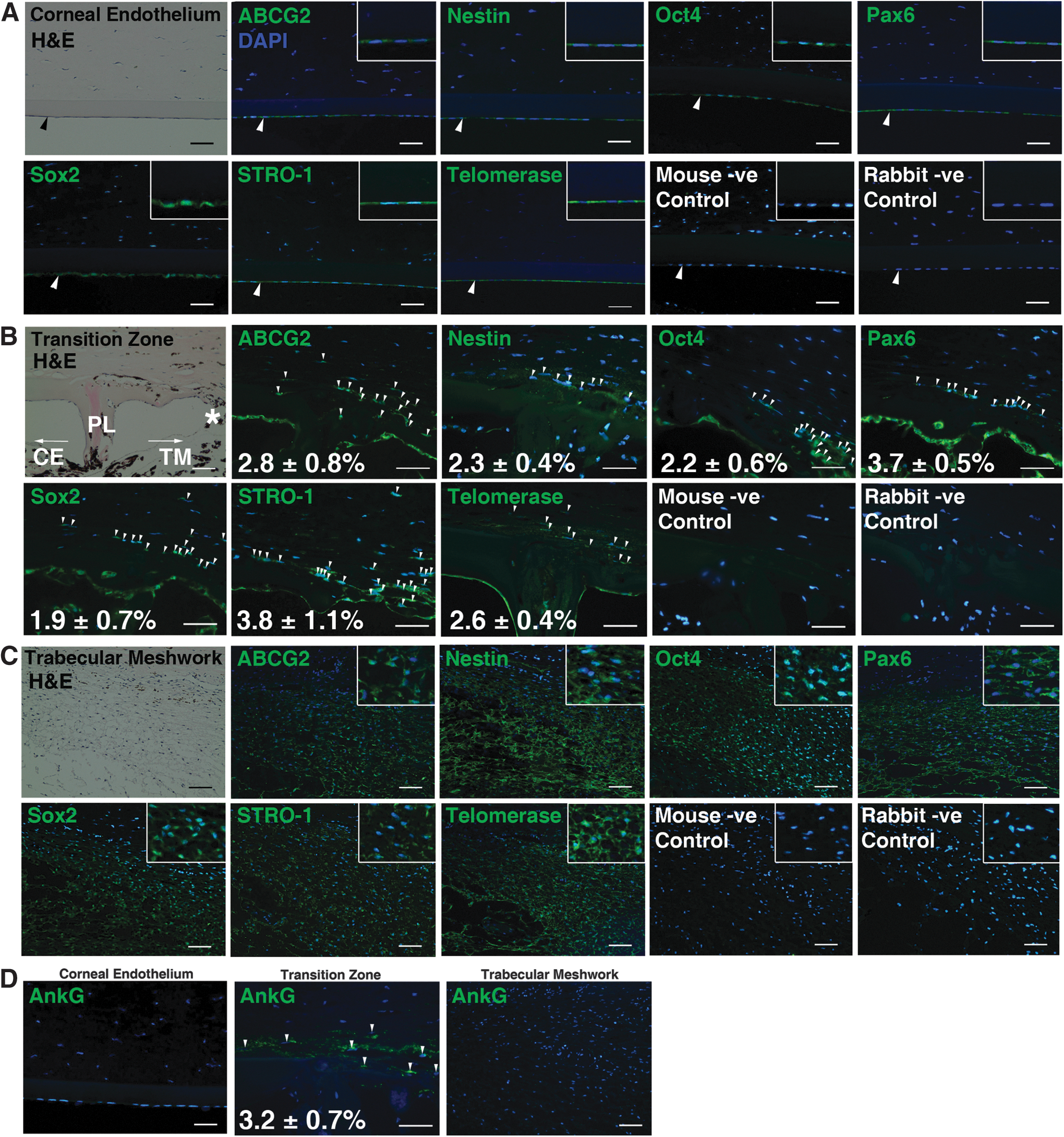
Immunolocalization of stem cell markers in the bovine posterior limbus.
Properties of the CE spheres
CE cells from the peripheral cornea generated more spheres
Single CE cells seeded at clonal density level grew and formed spheres after 7 days of culture. The peripheral CE cells generated significantly higher number of spheres (29.8±2.8 spheres/10,000 cells) than the central CE cells (20.8±1.7 spheres/10,000 cells) (Fig. 3A).
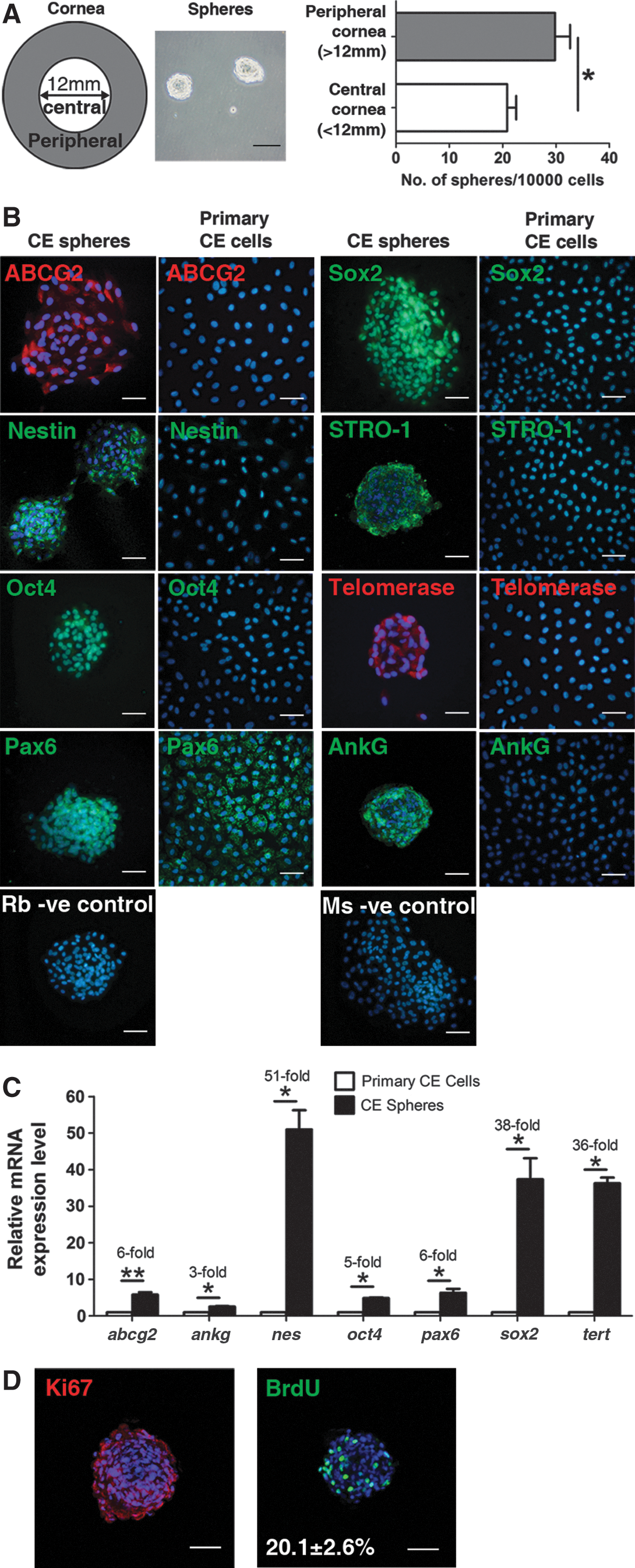
Characterization of the CE spheres.
CE spheres expressed a panel of stem cell markers
Immunocytochemical characterization showed that cells within the spheres expressed stem cell marker proteins ABCG2, nestin, Oct4, Pax6, Sox2, STRO-1, and telomerase (Fig. 3B). They were also AnkG-positive. Furthermore, the spheres expressed higher levels of stem cell gene transcripts such as nes, sox2, and telomerase reverse transcriptase (tert) than primary CE cells (Fig. 3C).
Self-renewal and proliferative capacity of CE spheres
Self-renewal and proliferative capacity of CE spheres were assessed by secondary spheres formation, Ki67 staining, and BrdU incorporation. CE spheres were able to form secondary spheres until the fifth cyclical passage. Over 90% of the cells within the CE spheres were positively stained with Ki67, while 20.1%±2.6% of the cells in the spheres incorporated BrdU (Fig. 3D).
Non-directed differentiation of CE spheres
Cell monolayers with a cobblestone appearance formed after 7 days. They reached confluence earlier than primary CE cells. These differentiated CE sphere progenies expressed CE differentiation marker proteins AQP1, N-cadherin, ZO-1, and Na+/K+ ATPase, in addition to connexin 43 (Fig. 4A). Sphere progenies expressed less stem cell genes transcript than primary CE cells, yet both of them showed comparable CE differentiation gene expression (Fig. 4B).
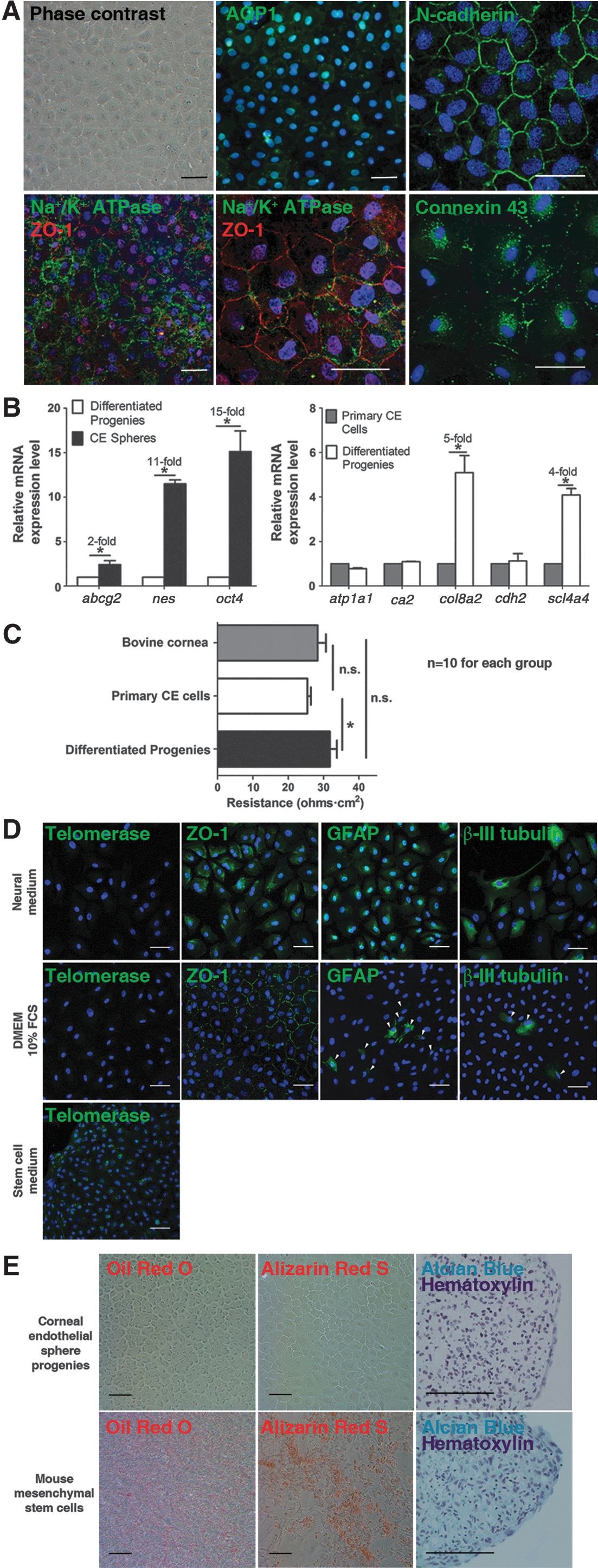
Differentiation properties of the CE spheres.
Electrical resistance measurement
The transendothelial electrical resistance of the native bovine cornea, primary CE cell layer, and differentiated sphere progenies was 28.3±4.1, 25.4±2.8, and 31.8±4.5 Ω·cm2 respectively. A significantly (P<0.05) higher resistance was found in monolayers derived from differentiated sphere progenies than primary CE cell monolayers (Fig. 4C).
Neural differentiation of CE spheres
Upon differentiation for 7 days, telomerase expression was down-regulated. In neural medium, over 50% of the sphere progenies were positive for the tight junction marker ZO-1 (cytoplasmic), glial marker GFAP, and neuronal marker TUJ1. Whereas with DMEM-cultured controls, most of the cells formed a monolayer of polygonal cells expressing ZO-1 (cell membrane). Only 4.5%±0.9% and 2.9%±1.2% of the cells expressed GFAP and TUJ1 respectively (Fig. 4D).
Mesenchymal differentiation of CE spheres
Cells derived from CE spheres showed polygonal morphology in adipogenic and osteogenic differentiation medium (Fig. 4E). Neither oil red O nor alizarin red S was observed. In chondrogenic differentiation, cell pellets were formed after 21 days of culture. However, paraffin sections of the pellets were negative for alcian blue staining, indicating the absence of proteoglycan synthesis. On the contrary, mMSCs readily differentiated into adipocytes, osteocytes, and chondrocytes.
Properties of the TM spheres
Sphere culture of TM cells
Single TM cells seeded at clonal density level grew and formed spheres after 7 days (27.8±3.2 spheres/10,000 cells).
TM spheres expressed a panel of stem cell markers
Similar to the CE spheres, the TM spheres also expressed stem cell markers ABCG2, nestin, Oct4, Pax6, Sox2, STRO-1 and telomerase (Fig. 5A) and were immunoreactive to AnkG. Compared to primary TM cells, TM spheres expressed significantly higher levels of the examined stem cell gene transcripts, particularly nes, pax6, sox2, and tert (Fig. 5B).

Characterization of the TM spheres.
Self-renewal and proliferative capacity of TM spheres
There was a lack of TM sphere formation beyond the sixth passage, indicating their loss of stemness. The TM spheres contained proliferative cells. Over 90% of the cells within the spheres were immunoreactive to Ki67 while 25.4%±3.1% of the cells in the spheres incorporated BrdU (Fig. 5C).
Non-directed differentiation of TM spheres
A confluent monolayer of TM-like cells was formed after 7 days. The TM-like cells expressed the TM markers AQP1, CHI3L1, and Ac-LDL (Fig. 6A). The TM-like sphere progenies expressed significantly less stem cell gene transcripts than the TM spheres. The TM-like cells showed significantly higher levels of TM differentiation marker genes than the TM spheres, but comparable levels when compared to primary TM cells (Fig. 6B).
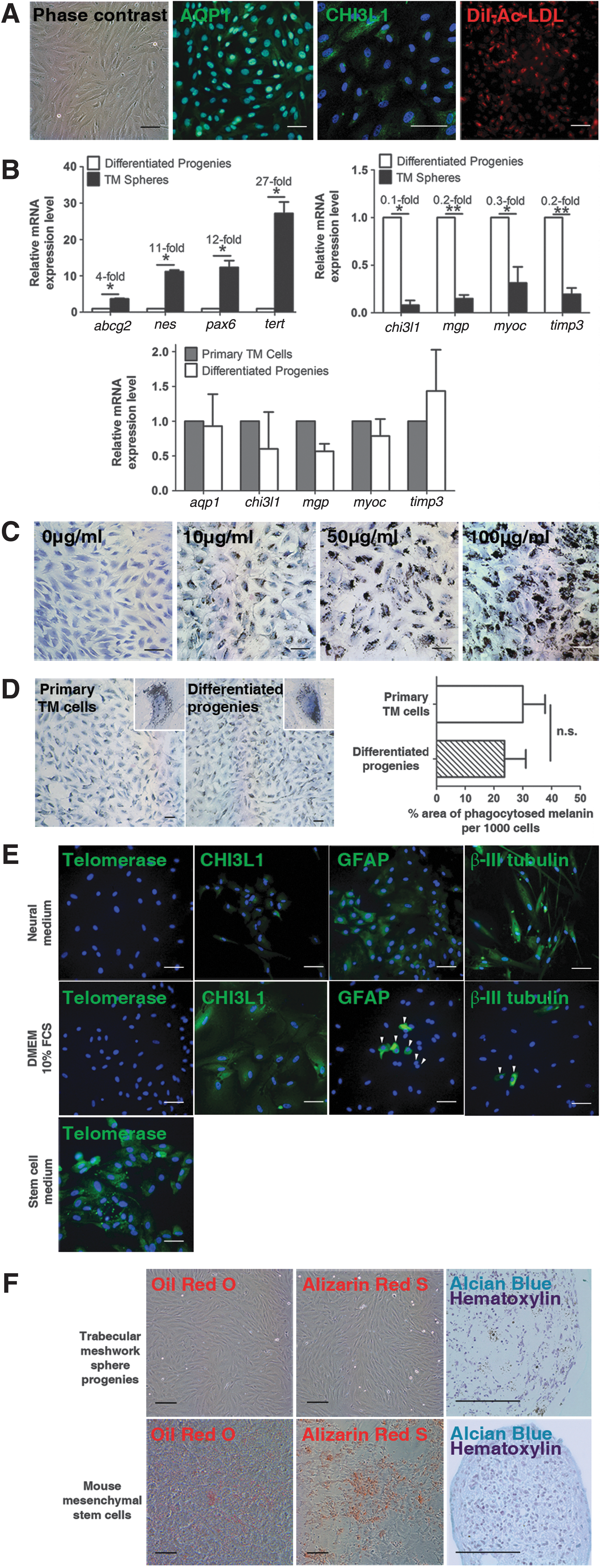
Differentiation properties of the TM spheres.
Melanin phagocytosis assay
The TM-like cells differentiated from the TM spheres demonstrated a strong capacity to phagocytose melanin granules in a concentration-dependent manner (Fig. 6C). However, the uptake was not uniform with a minority of cells being more phagocytic than the rest. When the cells were exposed to 100 μg/mL melanin, some cells took up such large amounts of melanin that they appeared round and engorged.
Both the primary TM cells and sphere progenies were capable of engulfing melanin at the 10 μg/mL concentration (Fig. 6D). The mean percentage area of phagocytosed melanin of the differentiated sphere progenies was about 79% of that of the primary TM cells.
Neural differentiation of TM spheres
After 7 days of neural induction, the sphere progenies showed a weak expression of the TM marker CHI3L1. However, majority of the sphere progenies stained positively for GFAP and TUJ1. Telomerase was only expressed when the cells were kept in SCM. In controls cultured with DMEM, most of the cells formed a monolayer of cells that expressed CHI3L1. 6.9%±2.1% and 4.8%±1.4% of the cells expressed GFAP and TUJ1 respectively (Fig. 6E).
Mesenchymal differentiation of TM spheres
Cells derived from the TM spheres showed the normal TM cell morphology in adipogenic and osteogenic differentiation media. No staining for oil red O or alizarin red S was observed. Cell pellets formed in chondrogenic differentiation were negative for alcian blue staining. On the contrary, mMSCs readily differentiated into adipocytes, osteocytes, and chondrocytes (Fig. 6F).
Discussion
Several other non-primate species have been studied for research purposes [31 –38], however, bovine CE and TM have been useful tissue culture sources since the 1980s [21,22]. In addition, the sheer size of the bovine outflow system and chamber angle structures plus the ready availability and inexpensiveness of bovine eyes would make it a compelling animal model if stem/progenitor cells were shown to be present in the target tissues.
In contrast to human, the bovine transition does not contain a smooth zone. However, meridional examination shows that the most anterior portion of the bovine TM also taper to insert beneath the peripheral cornea. This observation is novel as far as we know, leading to the speculation that stem-like cells may also be present in the bovine transition area, since the transition in primates is considered stem cell-rich [6].
The immunolocalization studies confirmed the presence of stem/progenitor cells in the bovine posterior limbus. Apart from the transition zone-confined AnkG staining, CE and TM showed no differential markers expression. However in human, stem cell markers were expressed in the peripheral or extreme peripheral CE, Schwalbe's line, and TM [9,11]. The discrepancy could be explained by the relative young age of the animals (1–2 years old) and difference of proliferative capacity between the human and bovine CE [1].
Similar to rabbit and rat, bovine CE cells can be readily grown in culture, but human CE cells are more difficult to culture, especially cells from older donors [25,39 –41]. In fact, bovine CE cells derived from older donors also showed lower proliferative capacity [42]. In addition, age-related decrease of the CE cell density was observed in rabbits and rats [43,44]. These suggest that albeit with relatively higher proliferative capacity, the replenish rate of the animal CE cells is lower than the rate of cell loss, which is also the case in human [1].
A plausible justification for the stem cell markers expression in the entire CE and TM is that they represent a population(s) of progenitor cells, instead of undifferentiated stem cells. The idea that stem-like cells may be present in the transition zone primarily came from the observation of an increase in TM cell division at the anterior portion of the TM post-argon laser trabeculoplasty [7]. It was believed that the laser treatment invigorated the putative stem cells in the anterior TM, which in turn stimulated TM repopulation. A small population of cells from primary human TM cell culture were found to escape replicative senescence and express AnkG [13]. Cells in the TM insert region exclusively expressed AnkG after laser trabeculoplasty [14]. Consistently, our data on bovine tissue (AnkG staining in the transition zone) suggests that cells at the TM insertion may be more stem-like. It is speculated that the cells gain more determined fates when they migrate away from the niche to the TM and/or CE (Fig. 7). Indeed, the cellular components of the microenvironment where the stem cells reside, and the signals emanating from the support cells have been found to influence directly the maintenance of “stemness” of a stem cell population [45,46].

Schematic diagram of the distribution of stem/progenitor cells in the bovine anterior chamber angle. It is speculated that the PET cells resides in a niche at the TM insertion within the transition zone. As they migrate away from the niche towards the CE or TM, they become more tissue-specific. PET cells, progenitor cells of the endothelium and trabeculum.
Cells at the extreme periphery of the CE (0.2 mm from the end of Descemet's membrane) were found to show greater expression of stem cell markers [11]. These cells were present as cell clusters between Hassall-Henle warts and located in deeper layers from the CE surface. Given that the TM insertion extends behind the end of Descemet's as far as 100 μm [47], it is tempting to speculate that the cell clusters may have originated from the cells at the TM insertion. They may be responsible for providing new cells that migrate continuously toward the center of the CE, while cell clusters are formed with the accumulation of Hassall-Henle bodies as we age. In response to TM injury, such as in argon laser treatment for glaucoma, this niche at the TM insertion may also provide new cells to repopulate the TM. It was found that laser placed close to Schwalbe's line advanced the uncontrolled endothelial overgrowth to cover the TM surface [48 –50]. We speculate that this response may be due to disruption of the niche, which activates the aberrant regeneration process. This phenomenon may be similar to the conjunctivalization response on the ocular surface, in which the conjunctiva overgrows the cornea when the epithelial limbal niche is damaged [51].
Having located the stem cell niche, we wanted to investigate whether the progenitor cells could be isolated and expanded in culture. However, due to the lack of specific markers, scarcity and complicated anatomical location of the PET cells, direct harvest of these cells from their niche in the transition zone can be very difficult, even with the much bigger bovine eyes. Hence, we utilized the sphere culture technique to enrich the progenitor cells from the CE and TM respectively.
The cells within the spheres expressed a panel of stem cell marker transcripts and proteins. They also expressed AnkG, a putative marker for PET cells [13 –15]. On the contrary, the primary cells only expressed Pax6. The young age of the animals may account for the presence of Pax6 protein, which is involved in cellular migration and differentiation in the CE and TM development [52]. The stem cell markers were present in the bovine CE and TM in situ; yet they were not expressed in the cultured primary cells. One possible explanation may be that the cells readily obtained a differentiated phenotype when they were maintained in DMEM/10% FBS medium. It is, however, intriguing to note that despite the apparent diverse proliferative potential of the CE in different mammals [1,53 –55], the CE spheres having been isolated from human, rabbits, and cows appear to share a highly conserved phenotype.
In agreement with previous findings in human and rabbits, our findings demonstrated that progenitor cells are present in the central and peripheral bovine CE, with a higher density at the periphery [20,22]. The peripheral CE cells may correlate to the subset of cells that showed higher proliferative potential in the bovine CE as reported by Huang et al. [56]. It will be interesting to study whether there is any difference between the sphere-forming efficiency of cells derived from the anterior and posterior portion of the TM. However, this comparison is hard to achieve due to the difficulty in separating the two portions of TM during dissection.
By way of comparison, some stem/progenitor cells that were isolated by sphere assay from other organ systems showed a far higher self-renewal property, while some were similar to our CE or TM spheres [57 –59]. The restricted renewal capacity of the spheres can be attributed to the occurrence of more asymmetric divisions in spheres of subsequent passages [59]. Increased number of differentiated and/or senescent cells with advancing passage number may be another possibility [60]. Whether it is the presence of apoptotic or necrotic cells in the CE and TM spheres that reduces their renewal capacity is another possibility that needs to be investigated.
The highly proliferative property of the CE and TM spheres as shown by Ki67 and BrdU staining justifies our earlier discussion that the spheres were more progenitor-like. It appears that long-term maintenance of the progenitor cells in serial passages becomes less critical in terms of clinical application, as we may have obtained sufficient functional differentiated cells from the highly proliferative primary spheres for transplantation.
When the spheres from CE and TM were differentiated without any induction factors, they predominantly adopted a CE-like or TM-like lineage, respectively. The sphere progenies became functional differentiated cells. Transcellular electrical resistance indicates the presence of functional tight junction [61]. Our data are comparable to those reported in the literature [62 –65] and suggest better tight junction formation in the differentiated CE sphere progenies than primary CE cells. In fact, it was found that tight junction proteins such as ZO-1 and claudin-1 were downregulated as neural crest cells differentiate [66,67]. Besides, the densities of Na+/K+ ATPase pump sites in the CE cells were found to increase as the CE become more mature [68]. Given the embryologic origin of the CE [69], our data seems reasonable in that the resistance was lower in the more differentiated primary CE cells.
Phagocytosis is one of the key functions of TM cells that help maintain normal aqueous outflow [70]. We showed that the TM-like cells differentiated from TM spheres had strong melanin phagocytosis capacity, which was comparable to that of the primary TM cells (∼80%). The propensity of CE or TM spheres to differentiate back into functional CE-like or TM-like cells is an important finding because cells destined to be transplanted need to be functional to rejuvenate the tissue.
Most of the cells differentiated from the spheres efficiently took on the neural lineages upon induction. This could be attributed to elevated expression of neural marker transcripts sox2 and nes. However, the spheres did not differentiate toward the mesenchymal lineages upon induction despite expressing MSC markers ABCG2 and STRO-1. In fact, human TM cells were shown to have certain similarities with adipose-derived MSC but with distinct differentiation properties [71]. Our data suggest that the spheres had a bipotent potential and were more prone to adopt their original lineage, indicating that they may be more appropriate for regeneration purposes than pluripotent or multipotent stem cells.
The evidence suggesting the presence of stem cells in the human TM has been accumulating for some time [6,7,9,12,15]. To date, there have been five studies that successfully cultivated the TM stem cells in vitro [16 –18,72,73]. These data proved that stem/progenitor cells from the human TM could be cultivated and home-in to their tissue of origin. Nonetheless, it remains obscure whether the cells isolated via different reported methods are actually the same population. In fact, there have been mixed results in the differentiation characteristics of these cells. It is likely that the age of the donor from which the stem cells are derived will affect the differentiation properties [74]. Together with our findings, it appears that the TM stem/progenitor cells are present in both primate and non-primate animals. In addition, sphere culture provides us a useful tool to enrich the CE and TM progenitor cells, which may be used for transplantation as tried or suggested by others [75 –77]. As a novel PET cell transplant approach might be applied in patients with CE diseases or glaucoma in the future, further research on these cells is required so that the finding of a replete non-primate source of PET cells is of enormous benefit.
Taken all together, our results showed that PET cells reside in a niche at the transition zone in the posterior limbus of bovine eyes and they can be isolated and cultured in vitro. The easy access, large number of cells and degree of certainty in harvesting cells that are well committed are prerequisites for clinical application. Given the shortage of human donor eyes, bovine eyes provide a readily available and large tissue source for the development of cell replacement therapies for CE diseases and/or glaucoma. With reliable harvesting of bovine PET cells, further detailed research and even xenotransplantation may be possible in the future, making our research highly applicable to regenerative medicine. As yet, it is important to note that more markers need to be identified to allow even more specific and effective isolation of PET cells.
Footnotes
Acknowledgments
We are grateful to Dr. Chi-wai Do and Mr. King-kit Li for their expert advice on the Ussing chamber experiments, Dr. Sharon Mason for the assistance on sphere culture, and Prof. Rachel Williams and Dr. Victoria Kearns for the help with SEM. We also acknowledge the assistance of the University of Hong Kong Li Ka Shing Faculty of Medicine Faculty Core Facility. This work was supported from grants from the International Glaucoma Association (IGA) and the Foundation for Prevention of Blindness.
Prior conference presentation: Yu WY, Grierson I, Sheridan C, Lo ACY, So KF, and Wong D. Bovine corneal endothelial precursors isolated by sphere-forming assay. UK National Stem Cell Network Scientific Conference 2011, York, UK, March 30–April 1, 2011.
Yu WY, Sheridan C, Grierson I, Lo ACY and Wong D. Enhanced survival and expansion of bovine corneal endothelial progenitors using accutase. The Association for Research in Vision and Ophthalmology Annual Meeting, Fort Lauderdale, US, May 6–10, 2012.
Yu WY, Sheridan C, Grierson I, Lo ACY and Wong D. Progenitors from the bovine trabecular meshwork express neural markers. Mercia Stem Cell Alliance Annual Scientific Meeting, Manchester, UK, December 19, 2012.
Yu WY, Sheridan C, Grierson I, Lo ACY and Wong D. Expression of stem cell markers in the bovine corneal endothelium, insert region and trabecular meshwork. The Association for Research in Vision and Ophthalmology Annual Meeting, Seattle, US, May 5–9, 2013.
Author Disclosure Statement
The authors indicate no potential conflicts of interest.