
Abstract
Mesenchymal stem cells (MSC) are present in the bone marrow, from where they are thought to migrate through the blood stream to the sites of injury. However, virtually all tissues contain resident MSC that may contribute to local regenerative and immunomodulatory processes, thereby hypothetically preempting the need for recruiting MSC through the bloodstream. Although there is some indication for circulating MSC in animal models, there is little solid evidence for the mobilization and migration of MSC in the human circulation. In the present study, we were unable to detect MSC in the blood of healthy individuals. We then searched for MSC in the blood of ten patients with end-stage renal disease, ten patients with end-stage liver disease, and in eight heart transplant patients with biopsy-proven rejection by culturing of mononuclear cells under MSC-supporting culture conditions. In none of these patient categories, MSC were identified in the blood. MSC were, however, found in the blood of a severe trauma patient with multiple fractures, suggesting that disruption of bone marrow leads to the release of MSC into the blood stream. The conclusion of this study is that MSC are not recruited into the circulation in patients with injured solid organs and during aggressive immune responses after transplantation.
Introduction
M
There is, however, controversy about whether MSC can be detected in the circulation in human. While some studies failed to detect MSC in peripheral blood [6], it has been demonstrated that cells with a stromal appearance and osteogenic differentiation potential can be isolated from buffy coats of healthy individuals [7]. Yet, it has been reported that cells with a fibroblast morphology that derive from peripheral blood mononuclear cells (PBMC) in culture actually express a hematopoietic immunophenotype [8] and do not, therefore, fulfill the criteria for MSC set by the International Society for Cellular Therapy (ISCT) [9]. Others have indicated that MSC can be detected in the peripheral blood in rats after housing them for 3 weeks under hypoxic conditions [10], or in mice after the induction of liver injury by two injections with CCL4 [11]. These studies demonstrate that MSC can circulate at least under extreme conditions, but do not evidence that this occurs under more physiological conditions. Furthermore, the results may possibly be different in man and murine. In man, there is a report that shows that plastic-adherent cells with an MSC immunophenotype are present in the blood of hip fracture patients [12]. It is, however, questionable whether these cells actively enter the circulation through migratory processes or whether they end up in the blood stream through mechanical disruption of the bone tissue. So far, there is no convincing evidence that cells that adhere to the ISCT criteria for MSC enter the circulation through biological processes in human, whereas this has been demonstrated for endothelial progenitor cells [13], hematopoietic progenitor cells [14], and for MSC that are mobilized from graft tissue during organ transplantation [15].
If MSC are capable of migrating through the blood stream to areas of injury, then the highest chance of detecting them would be in diseased individuals. Therefore, in the present study, we set out to investigate whether MSC can be isolated from the blood of patients with severe inflammatory and degenerative diseases. For this purpose, blood samples were collected from end-stage liver failure patients and end-stage kidney failure patients just before organ transplantation and from heart transplant patients that went through a rejection episode early after transplantation and examined for the presence of MSC. As a positive control, blood was collected from severe trauma patients with multiple bone fractures.
Materials and Methods
Blood sample collection
Blood was collected from healthy individuals and from 10 end-stage liver disease and 10 end-stage kidney disease patients on the day before organ transplantation. Blood from eight heart transplant patients was collected at the time of a biopsy-proven rejection within the first weeks after transplantation. Blood from two trauma patients that fell from 10 meter heights, thereby acquiring multiple bone fractures and other injuries, was collected respectively 8 and 9 days after the accidents. Blood was collected in heparin tubes and PBMC were separated by the Ficoll-Paque PLUS (density 1.077 g/mL; GE Healthcare, Uppsala, Sweden). PBMC were stored at −150°C until usage. Blood sample collections were approved by the institutional Medical Ethics Committee (MEC-2010-022).
Flow cytometric analysis of PBMC
PBMC were stained for CD45, CD73, CD90, and CD105 (all BD Biosciences, San Diego, CA) and analyzed on a FACSCanto II flow cytometer with the BD FACSDiva software (BD Biosciences). At least 500,000 CD45− events were gated and analyzed for the expression of the MSC cell surface markers CD73, CD105, and CD90. To detect the specificity of the staining, cells stained only for CD45 were used.
Culture of PBMC
PBMC were thawed, washed, and seeded at approximately 200,000 cells per cm2 in minimum Essential medium Eagle's alpha modification (MEM-α) (Sigma-Aldrich, St. Louis, MO) supplemented with 2 mM
Culture of MSC from adipose tissue
The stromal vascular fraction (SVF) of adipose tissue is a rich source of MSC. To determine whether Ficoll separation would affect the culture of MSC, the SVF was isolated as described before [16] and subjected to Ficoll-Paque PLUS. In brief, subcutaneous adipose tissue was collected from healthy individuals that underwent surgery for kidney donation after written informed consent as approved by the institutional Medical Ethics Committee. Adipose tissue was mechanically disrupted and enzymatically digested with collagenase type IV (Life Technologies, Paisley, United Kingdom). The obtained SVF was put on Ficoll-Paque PLUS and the mononuclear fraction cultured in the culture medium as described above. The culture medium was refreshed twice weekly.
Culture of MSC liver perfusates
We have previously demonstrated that the perfusion fluid obtained from liver grafts contains liver-derived MSC, which were found to be highly similar to bone marrow MSC with regard to differentiation potential, gene expression, immunosuppressive and regenerative capacities [15,17]. To test whether Ficoll separation and freezing of cells affected the efficiency of MSC culture expansion, liver perfusate cells were separated by Ficoll-Paque PLUS and mononuclear fractions were stored frozen at −150°C until usage. Upon thawing, the cells were treated as described above to serve as positive controls. The use of liver perfusate cells was approved by the institutional Medical Ethics Committee.
Microscopy
Cultures were inspected for the presence of cells with MSC morphology using an Axiovert 40 microscope (Zeiss, Goettingen, Germany). Images were made using the Canon EOS 1000D digital camera and Axiovision software (Zeiss).
Flow cytometric analysis of cultured cells
Cells were removed from culture flasks using 0.05% trypsin-EDTA (Life Technologies, Paisley, Great Britain) and stained for CD13, CD31, CD45, CD73, CD90, and CD105 (all BD Bioscience). Unstained cells were used as negative controls. Cells were analyzed on a FACSCanto II flow cytometer with the BD FACSDiva software.
Results
Circulating MSC in healthy individuals
To examine whether cells with an MSC immunophenotype (CD45−CD73+CD90+CD105+) were detectable in the peripheral blood of healthy individuals, PBMC of six healthy volunteers were Ficoll-separated and analyzed by flow cytometry. CD45+ cells were out gated and at least 500,000 CD45− cells, comprising hematopoietic and endothelial progenitor cells, thrombocytes, and potentially MSC, were analyzed for expression of the MSC markers CD73, CD90, and CD105. No CD45− cells that coexpressed CD73, CD90, and CD105 were detected (Fig. 1), suggesting cells with a MSC immunophenotype are not present in the blood of healthy individuals. To confirm the lack of MSC in blood by a more sensitive method and to circumvent the possibility that circulating MSC have a different immunophenotype making them undetectable by flow cytometry, PBMC were cultured in MSC supporting medium to detect the outgrowth of MSC colonies from potentially single cells. No MSC colonies were found in cultures of PBMC from healthy individuals (n=5, data not shown).
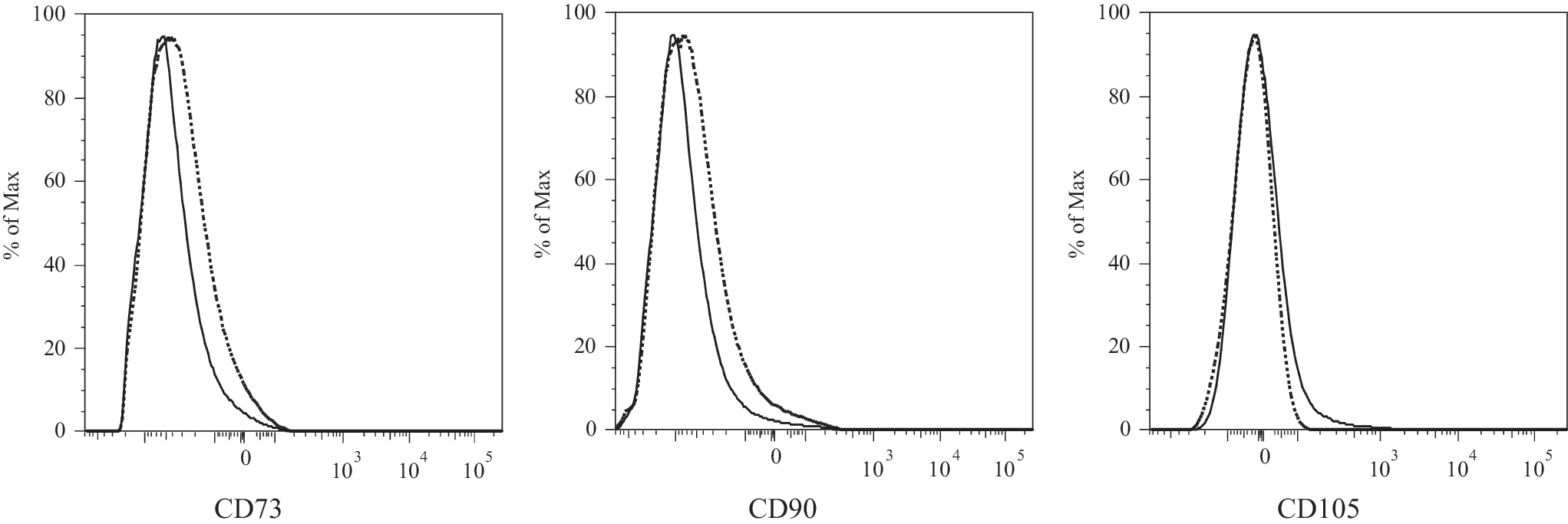
Flow cytometric analysis of peripheral blood mononuclear cells (PBMC) of healthy individuals for the presence of mesenchymal stem cells (MSC). At least 500,000 CD45− events were gated and analyzed for expression of the MSC markers CD73, CD90, and CD105. Solid line histograms represent stained cells, dotted histograms represent unstained cells. A representative experiment is shown.
Circulating MSC in organ failure patients
The lack of MSC in the blood of healthy individuals may be explained by the absence of injury or inflammation, which may act as a trigger for the mobilization and attraction of MSC. To examine whether MSC are present in the blood of patients with injury and/or inflammation, blood was collected from three groups of organ failure patients with abundant inflammation and tissue damage. The first group comprised of end-stage kidney disease patients on dialysis from whom blood was collected on the day before kidney transplantation (n=10). The second group were end-stage liver failure patients from whom blood was taken on the day before liver transplantation (n=10). The third group were heart transplant patients up to 2 months after transplantation. Blood was taken at the time of biopsy-proven rejection (n=8). Patient characteristics are shown in Table 1.
M, male; F, female; HTx, heart transplantation; MMF, mycophenolate mofetil; MSC, mesenchymal stem cells.
PBMC were put in culture in MSC-supporting culture medium. In all cultures, plastic-adherent cells were visible within days, but no cells with an MSC/fibroblastic morphology were observed. Instead, the cells showed a macrophage-like morphology (Fig. 2A). Flow cytometric analysis of the cultures after 2–3 weeks demonstrated that all cells were CD45+, and a subset was CD13+, and weakly CD31+ and CD105+ (Fig. 3A). The cells, thereby, did not meet the MSC immunophenotype (CD45−, CD31−, CD13+, CD73+, CD90+, CD105+).
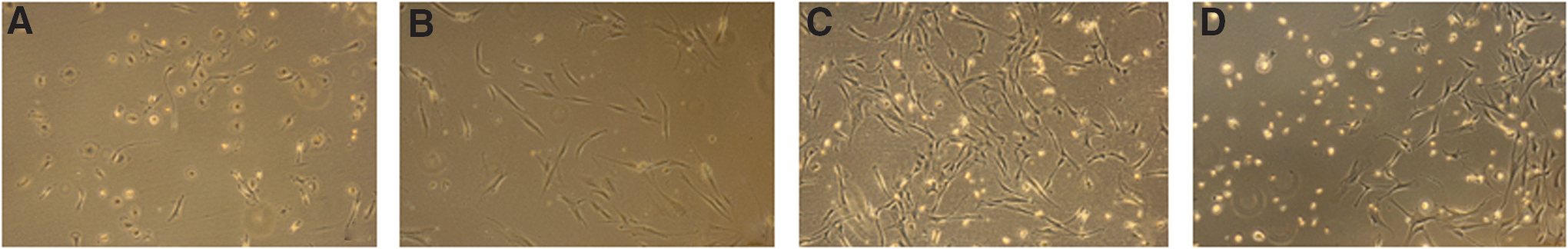
Cultures of PBMC from blood of organ failure patients do not contain cells with an MSC morphology, whereas adipose tissue, liver perfusates, and PBMC from blood of a trauma patient do.

Flow cytometric analysis of adherent cells in cultures of blood from a kidney disease patient
MSC from adipose tissue, liver perfusate, and blood from trauma patients
To rule out that the Ficoll separation procedure was toxic for MSC and to confirm that the culture conditions used were supportive for MSC, cells of the SVF of adipose tissue, known to be rich of MSC [18] and routinely isolated and cultured in our laboratory [16,19], were put on Ficoll and brought into culture (n=1). In addition, liver transplant perfusates, previously demonstrated to contain MSC [15], were put on Ficoll and the obtained cells were stored frozen. Subsequently, cells were thawed and brought into culture (n=2). Adherent cells with a fibroblastic morphology appeared within days after seeding of the SVF and liver perfusate cell suspensions (Fig. 2B, C). Finally, as MSC were previously reported in the blood of hip fracture patients [12], PBMC were collected from the blood of trauma patients with multiple bone fractures (see Table 1 for details) and brought into culture. In one of the patients, cells with an MSC morphology appeared in the cultures (Fig. 2D). This indicates that viable MSC can be present in the blood and can be brought into culture, although the cells most likely originated from a single colony.
Flow cytometric analysis of the cells cultured from adipose tissue and liver perfusates demonstrated that all cells expressed the MSC cell surface markers CD13, CD73, CD90, and CD105 and were negative for CD31 and CD45 (Fig. 3B, C). In a culture of PBMC from one of the trauma patients, a subset of cells was found that was CD45− and CD31− (Fig. 3D). Analysis of the CD45− cells demonstrated that the majority of these cells were CD13+, CD73+, CD90+, and CD105+, confirming their MSC phenotype (Fig. 3E).
These results indicate that the isolation and culture procedures used in the present study were supportive for the culture of MSC, but that MSC are not present in the blood of organ failure patients.
Discussion
The role of MSC in mediating tissue repair and modulating immune responses has been considered in many research and review papers. The mechanisms through which these processes may occur are, however, largely unclear and, therefore, widely speculated upon. One of the key questions on MSC biology is whether these cells can migrate toward sites of injury and inflammation. There is some evidence that infused MSC home to such sites [3 –5], but other studies indicated that although MSC have a beneficial effect, they act from a distance and are short lived after infusion [20,21].
In the present study, we investigated whether MSC could be detected in the circulation of patients with severe organ failure. While MSC could not be found in the blood of healthy individuals by us and others [8,22], it was hypothesized that they would be recruited into the bloodstream by signals coming from injured organs. Although several studies demonstrated the mobilization of hematopoietic stem cells in organ injury [23,24], we could not detect MSC in the blood of patients with end-stage kidney failure, end-stage liver failure, or in patients that underwent a rejection episode shortly after heart transplantation. We could only detect MSC in the blood of a trauma patient with multiple bone fractures, which is in line with a previous study that detected MSC in the blood of hip fracture patients [12]. In bone fracture patients, MSC may be released into the blood stream by mechanical disruption of the bone marrow rather than by active migration.
The results of this study imply that MSC are not recruited to sites of injury through the bloodstream. It is possible that the medication taken by the large majority of the patients is harmful to the circulating MSC. However, medication was very variable between all patients. Furthermore, MSC can be isolated from bone marrow and adipose tissue of patients with end-stage renal disease that are on similar medication [25,26]. Therefore, it is reasonable to assume that the blood stream is not the route of migration for MSC. It was recently demonstrated that adipose tissue-derived MSC can be found in lymph fluid, suggesting that the lymph system is used by MSC as a route for migration [27]. A possible limitation of the present study is that the organ injuries of the studied patients were not acute enough for recruitment of MSC from distant sites. In particular, the kidney disease patients suffered from chronic disease, whereas the liver disease patients contained both patients with acute and chronic disease. The heart transplant patients suffered from acute rejection, but the time of diagnosis of rejection may be some time after the onset of the rejection. We cannot totally exclude the possibility that MSC are recruited into the bloodstream shortly after severe organ injury.
The discrepancy between the results of the present study and other, in particular, animal studies may in the first place depend on the differences between the biology of experimental animals and humans. Rodents show better regenerative responses than human and this might rely on the recruitment of distant MSC in rodents and the lack of this regenerative mechanism in humans. Furthermore, circulating rodent MSC may show a different phenotype than human MSC, making them easier detectable by flow cytometry. Possibly rodent circulatory MSC are better adapted to cell culture conditions than human circulatory MSC, making them better available after seeding in a culture dish. Finally, experimental animals are usually exposed to more extreme conditions than humans and can be studied in a more controlled manner. Trauma can be timed and varied in severity and blood can be taken on chosen time points in animal studies. This is not possible and not desirable in human studies.
Our data from organ transplant patients are in support of the idea that MSC do not migrate through the blood stream toward injured organs. Transplantation is a unique model to pursuit migrating MSC as recipient cells migrating to the donor organ can be detected by HLA typing. Furthermore, transplanted organs are initially not connected to the lymph vasculature, leaving the circulation as the only route toward the organ. We have previously demonstrated that recipient MSC are not present in heart transplants up to 6 years after transplantation [28]. These heart transplants suffered from brain death and ischemia-reperfusion related injury and were exposed to alloreactivity for years, but even so were found to contain only donor MSC. A study in lung transplants found very similar data [29]. These data are supportive for the lack of evidence for circulating MSC.
Whereas MSC and hematopoietic cells are considered to belong to diverged lineages with specialized functions, there is some evidence that the distinction between the cells may not be as sharp as commonly thought. Circulating CD14+ monocytes can be transformed in multipotent cells in vitro that have endothelial and mesenchymal differentiation potential [30]. Adipose tissue-derived hematopoietic cells contain a population of macrophages that, like MSC, can differentiate into osteogenic, adipogenic, and chondrogenic lineages [31]. It is, therefore, not unlikely that under particular conditions hematopoietic cells can be mistaken for MSC. Hypothetically, hematopoietic cells with differentiation capacity migrate to injured organs to contribute to regeneration and immune modulation.
In conclusion, MSC, with the phenotype and characteristics as described by the ISCT, are not present in the circulation. All organs contain resident MSC that can contribute to organ repair and immunomodulation, making the recruitment of MSC from distant sites obsolete.
Footnotes
Author Disclosure Statement
No competing financial interests exist.