
Abstract
Gene expression analysis provides an effective methodology to identify clinically relevant genes implicated in intervertebral disc (IVD) pathology. The analysis of gene profile in mesenchymal stem cells (MSCs) from human herniated IVD (H-IVD) and degenerated IVD (D-IVD) has not yet been investigated. We present in this study a characterization of MSCs isolated from clinically categorized H-IVD and D-IVD disc samples. H-IVD-MSCs and D-IVD-MSCs showed multipotent mesenchymal differentiation ability, expressing positivity for adipogenic, osteogenic, and chondrogenic markers with an immunophenotypical profile representative of MSCs. FACS analyses revealed a higher expression of CD44 in D-IVD-MSCs compared to H-IVD-MSCs. Gene expression profile revealed that most genes under investigation displayed large variations and were not significantly different in the two types of analyzed IVD-MSCs. Conversely, the gene expression of osteopontin (OPN), a protein involved in bone matrix mineralization and extracellular matrix destruction, was found markedly increased (more than 400-fold) in D-IVD-MSCs compared to H-IVD-MSCs. Moreover, the OPN protein expression was detectable only in D-IVD-MSCs, and its levels were directly related with D-IVD severity. These findings suggest that an abnormal expression of OPN in D-IVD-MSCs occurs and plays a pivotal role in the pathophysiological process of human disc degeneration. We speculate that the regulation of the OPN pathway might be a therapeutic target to counteract disc degeneration.
Introduction
I
A recent article on the expression profile of different inflammatory cytokines showed that the expression of interleukin (IL) 4, IL6, IL12, as well as that of interferon-γ was modest in D-IVDs, but relevant in H-IVDs, suggesting that these cytokines might be involved in the pathogenesis of disc herniation [8]. In support of a putative role of inflammatory processes in the onset of IVD disease, we recently reported that a subpopulation of cells in D-IVDs, named mesenchymal stem cells (MSCs), expresses neuroinflammatory markers during differentiation [9]. Prompted by this evidence, and aiming to contribute to clarify the pathophysiological mechanisms underlying the alterations of D-IVD, we isolated MSCs from both human H-IVDs and D-IVDs and analyzed their gene expression profile of inflammatory cytokines and receptors.
Materials and Methods
Participant and specimens
Approval was obtained from the Institutional Ethics Committee and patient informed consent was obtained from all patients before sample collection. IVD specimens were obtained at surgery from 21 patients.
Specimens were obtained from the lumbar (L4/L5 or L5/S1) region and categorized as H-IVD (n=10) or D-IVD (n=11) disease and Thompson grade was recorded from MRI scan [10] (Table 1). At the time of surgery, all H-IVD specimens were found fully extruded through the posterior annulus. D-IVD specimens were removed from discs of patients undergoing spinal stabilization in spondylolisthesis and spinal instability and were neither herniated nor bulging, being thus in structural continuity with the rest of the disc.
IVD, intervertebral disc.
Establishment of H-IVD-MSC and D-IVD-MSC cultures
After surgery, the whole disc samples (3.2±0.5g) were collected and placed in tubes with stem cells medium (SCM) containing DMEM-F-12 with basic fibroblast growth factor (human recombinant, 10 ng/mL; Peprotech, Rocky Hill, NJ, or Upstate Biotechnology, Lake Placid, NY), epidermal growth factor (human recombinant, 20 ng/mL; Sigma-Aldrich, Milan, Italy) supplemented with 10% fetal bovine serum (Gibco, Grand Island, NY), and 1% penicillin/streptomycin solution (Sigma-Aldrich) as previously described [9]. The disc tissue was then enzymatically digested with 0.25% Liberase Blendzyme2 (Roche Diagnostics, Indianapolis, IA) in Dulbecco's phosphate buffered saline (Euroclone, Milan, Italy) for 6 h. After filtration through a 100 μm cell strainer (Falcon, Becton Dickinson, Allschwil, Switzerland) to remove tissue debris, the disc lysate was centrifuged (300 g for 10 min). Disc cells were then resuspended in SCM seeded in a 75-cm2 flask (2×104 cells/cm2), and incubated at 37°C in a humidified atmosphere containing 5% CO2. Cells were weekly detached by TrypLE Select (Gibco) and replated as above. Cells were routinely observed with an inverted phase-contrast microscope (Nikon Eclipse TE300; Nikon, Shinjuku, Tokyo, Japan), and images were acquired with a digital camera (Carl Zeiss Microscopy GmbH, München, Germany).
Proliferation assays
H-IVDs (n=10) and D-IVD (n=11) cells were seeded in 25-cm2 flasks (2×104 cells/cm2) in 3.5 mL of SCM and incubated as above for 7 days. Cells were then harvested, counted, and seeded at the same initial density for the next passage count. The total number of dead and viable cells was counted at each passage by Trypan Blue dye exclusion assay in a Fuchs-Rosenthal chamber and the percentage of viability was assessed by the formula: n viable cells/(n viable cells+n dead cells)×100. Growth curves were obtained by counting the number of viable cells and calculating the cumulative population at each passage. Moreover, to evaluate the population doubling time (PDT), MSCs were plated (104 cells/cm2) in 25-cm2 culture flasks and grown to 100% confluence. Cells were then detached as above and counted. The PDT was calculated according to the following equation: ln(2)/relative growth rate (RGR). RGR was determined by the following formula: (logN−logN 0)/(t−t 0), where N 0 and N are the numbers of plated and harvested cells, respectively, and (t−t 0) is the time interval of the growth period in days.
Multipotent differentiation ability of H-IVD-MSCs and D-IVD-MSCs
H-IVD (n=5) and D-IVD (n=5) cells were tested for their capacity to differentiate into the three mesodermal lineages, according to the minimal criteria suggested by Dominici et al. [11]. Adipogenic, osteogenic, and chondrogenic cell differentiation was induced by the Human Mesenchymal Stem Cell Functional Identification Kit (R&D Systems, Minneapolis, MN), as previously described [9]. Briefly, cells were cultured in SCM in the presence of either adipogenic or osteogenic supplements. To test in vitro chondrogenic differentiation potential, MSCs were submitted to a micromass pellet culture in the presence of chondrogenic supplement. After 21 days, cells were fixed and stained with FABP-4, osteocalcin, or aggrecan antibodies (R&D Systems), to determine adipogenic, osteogenic, and chondrogenic differentiation, respectively. Moreover, H-IVD and D-IVD cells differentiated in chondrogenic supplement were assayed for the expression of glycosaminoglycans (GAG), by staining with Safranin O and Fast Green Solutions (Sigma-Aldrich). Nuclei were then counterstained with 4′-6-diamidino-2-phenylindole (DAPI) (1 mg/mL) (Chemicon, Millipore, Billerica, MA). Cells were viewed with a Zeiss Axiophot-2 microscope, and images were acquired by the Axion Vision Software (Zeiss).
Immunophenotypic analyses
For each analysis, 5×104 cells (passage 6–8) were incubated with the appropriate phycoerythrin or fluorescein isothiocyanate conjugated antibody to evaluate the expression of the following pattern of mesenchymal, hematopoietic, endothelial, and immunological cell surface markers: CD14, CD34, CD44, CD45, CD73 (BD Pharmingen, San Jose, CA), CD105 (AbDSerotec, Raleigh, NC), CD90 (Millipore, Temecula, CA), and CD19 (Beckman Coulter, Cassina de’ Pecchi, Milano, Italy) [9].
RT2 profiler analysis
For molecular analyses H-IVD (n=3) and D-IVD (n=3) cells were used at passage 6–8. Briefly, total RNA was extracted by the RNeasy kit (Qiagen, Valencia, CA) and reverse-transcribed using the iScript cDNA Synthesis Kit (Bio-Rad Laboratories, Hercules, CA), following the manufacturer's instructions. cDNA was then added to the RT2 SYBR Green qPCR Master Mix (SABiosciences, Frederick, MD) and aliquots from each sample were used for the Human Inflammatory Cytokines & Receptors RT2 Profiler PCR Array (SABioscience). Real-time polymerase chain reaction (PCR) was carried out on an ABI 7300 thermal cycler (Applied Biosystems, Foster City, CA). PCR array data were analyzed using the Free PCR Array Data Analysis Software provided by the manufacturer (
Western blotting
Cell lysates (40 μg proteins) from 5 H-IVD and 5 D-IVD from discs representative of different Thompson grades were analyzed by immunoblotting with anti-osteopontin (OPN) and anti-β-actin antibodies. In detail, H-IVD and D-IVD MSCs, passage 6–8, were lysed with RIPA lysis buffer (150 mM NaCl, 1% NP-40, 0.5% DOC, 0.1% SDS, 50 mM Tris-Cl, pH 8.0) in the presence of a protease inhibitor cocktail (Sigma-Aldrich). Cell proteins were resolved by SDS-PAGE on 12% polyacrylamide gels and transferred onto nitrocellulose membranes. Membranes were blocked in TBS-Tween 20/bovine serum albumin (0.1%:5%) for 1 h. After washing, membranes were first incubated (1 h, room temperature) with the anti-OPN antibody (1 μg/mL) (cod. ab63856; Abcam, Cambridge, United Kingdom), and then with a goat anti-rabbit horseradish peroxidase-conjugated secondary antibody (30 min, room temperature). β-actin was used as loading control. Bound antibodies were visualized by ECL (SuperSignal West Pico Sensitivity Chemiluminescent Substrate), and membranes were exposed to Kodak Biomax films.
Statistical analyses
The results are expressed as mean±SD. For all statistical analyses, one-way ANOVA two-tailed test was used, and performed with the GraphPad Prism software, version 4.0. P values of less than 0.05 were considered to be significant.
Results
H-IVD and D-IVD cultures exhibit multipotent differentiation ability
MSCs were isolated from tissue samples obtained from H-IVDs and D-IVDs. Once seeded, both cell types adhered to plastic, and exhibited a fibroblastic-like morphology (Fig. 1a, b). At early (2–3) passages, the number of viable cells from H-IVD (82.13%±4.06%) was found significantly higher than that from D-IVD (71.78%±4.67%) and a similar trend was maintained later on. After 6–8-passages, H-IVD cells viability was slightly higher (95.3%±5.16%) than that of D-IVD cells (91.65%±3.17%) (Fig. 1c). In both cell models, growth curves revealed that the cell number gradually increases with time. In particular, in both cases cells started proliferation after 3 weeks (passage 3), a time required for cell selection in SCM medium, and continued to propagate along with passages (Fig. 1d). Of note, H-IVD showed a higher ability to proliferate than D-IVD. As shown in Table 2, at initial passages, cell doubling time of the D-IVD cell population was significantly lower than that of H-IVD cells, and along with culture time, D-IVD-MSCs continued to grow more slowly than H-IVD cells. The most striking difference was found at the first passages, possibly indicating that D-IVD-MSCs required more time to adapt to culture conditions and efficiently proliferate than H-IVD cells.
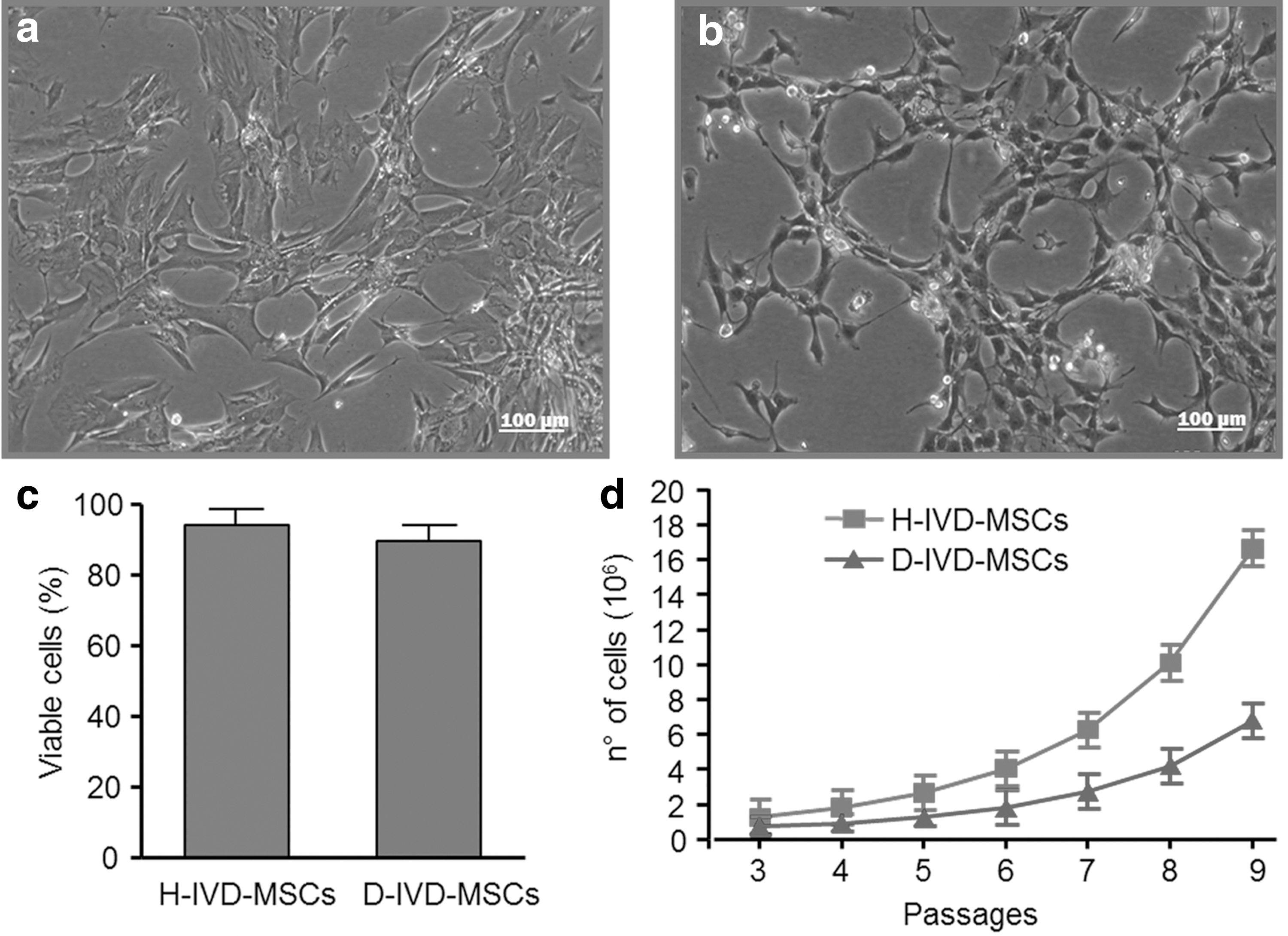
Proliferative characterization of H-IVD-MSCs and D-IVD-MSCs. Phase contrast representative images of H-IVD-MSCs
P<0.01.
P<0.05.
MSCs, mesenchymal stem cells.
To investigate the multilineage differentiation potential of H-IVD and D-IVD cells, we next performed in vitro adipogenic, osteogenic, and chondrogenic differentiation assays. After 3 weeks of adipogenic stimulation, most H- and D-IVD-MSCs were found positive to the intracellular lipid transport protein FABP4 (Fig. 2a, b), a recognized surface marker of adipocyte cells. Regarding osteogenic differentiation, after 21 days in the appropriate culture medium, both H- and D-IVD cells were positive to osteocalcin, a specific marker of osteoblasts (Fig. 2c, d). Moreover, differencing conditions favoring cell chondrogenic phenotype induced GAG deposition in both H-IVD-MSCs and D-IVD-MSCs, as revealed by positive immunoreactivity to aggrecan (Fig. 2e, f), as well as to Safranin O and Fast Green (Fig. 2g, h) in both MSC types. No differences were found between MSC differentiation potential from degenerated or herniated tissues (Fig. 2a–h).
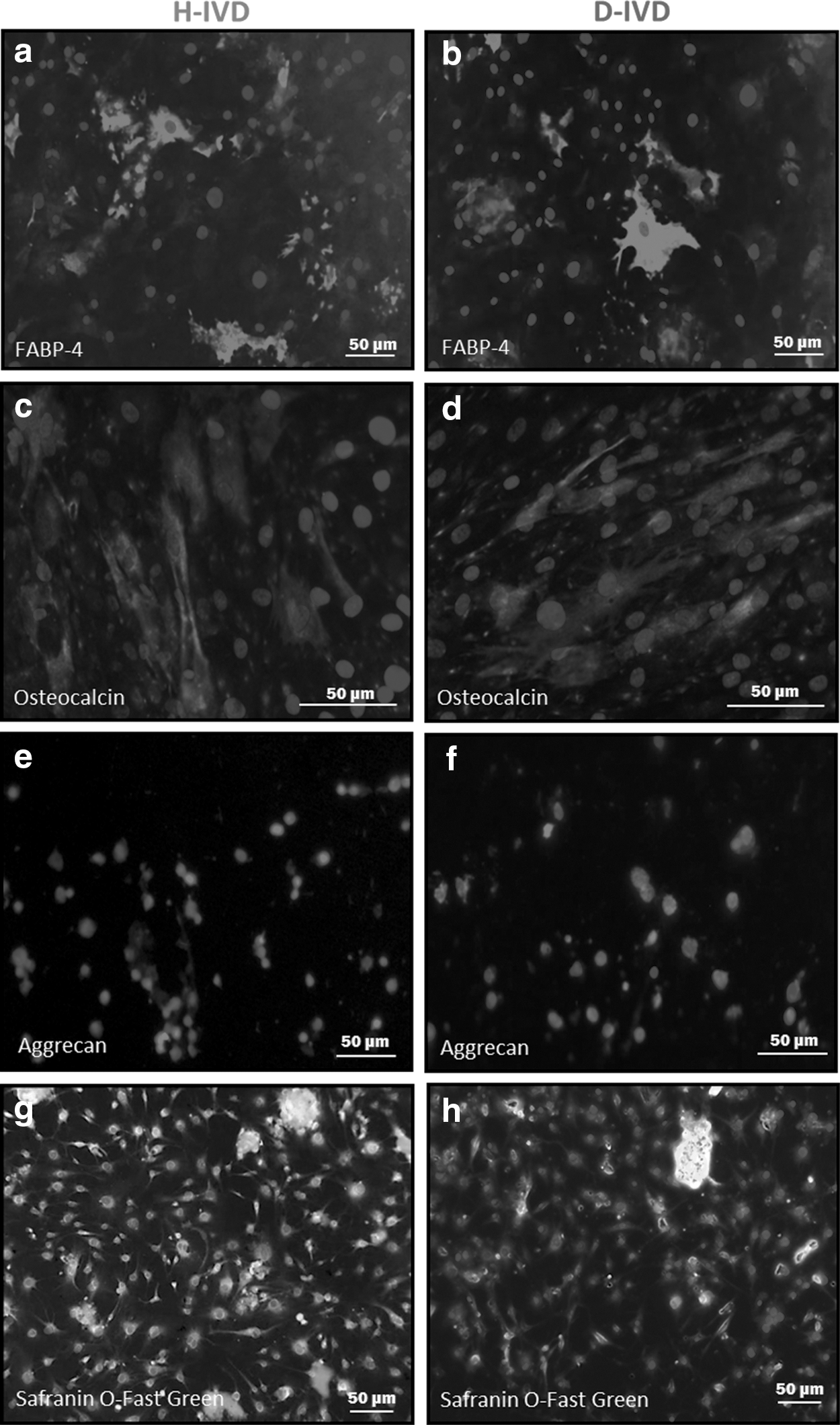
Multipotentiality of H-IVD-MSCs and D-IVD-MSCs. MSCs were maintained under adipogenic, osteogenic, and chondrogenic inducing media for 21 days. At the end, cells were fixed and stained for FABP-4
H-IVD and D-IVD MSCs express stem cell markers
Flow cytometric analysis revealed that cells from H- and D-IVDs were positive to different recognized markers of MSCs, including CD44, CD73, CD90, and CD105. Noteworthy, the percentage of CD44 positive cells was significantly higher in the D- than in the H-IVD cell population (Fig. 3). In addition, both cell types were negative for either the pan-monocytic antigen CD14, or the hematopoietic markers CD34 and CD45, or the pan-B-cell marker CD19.
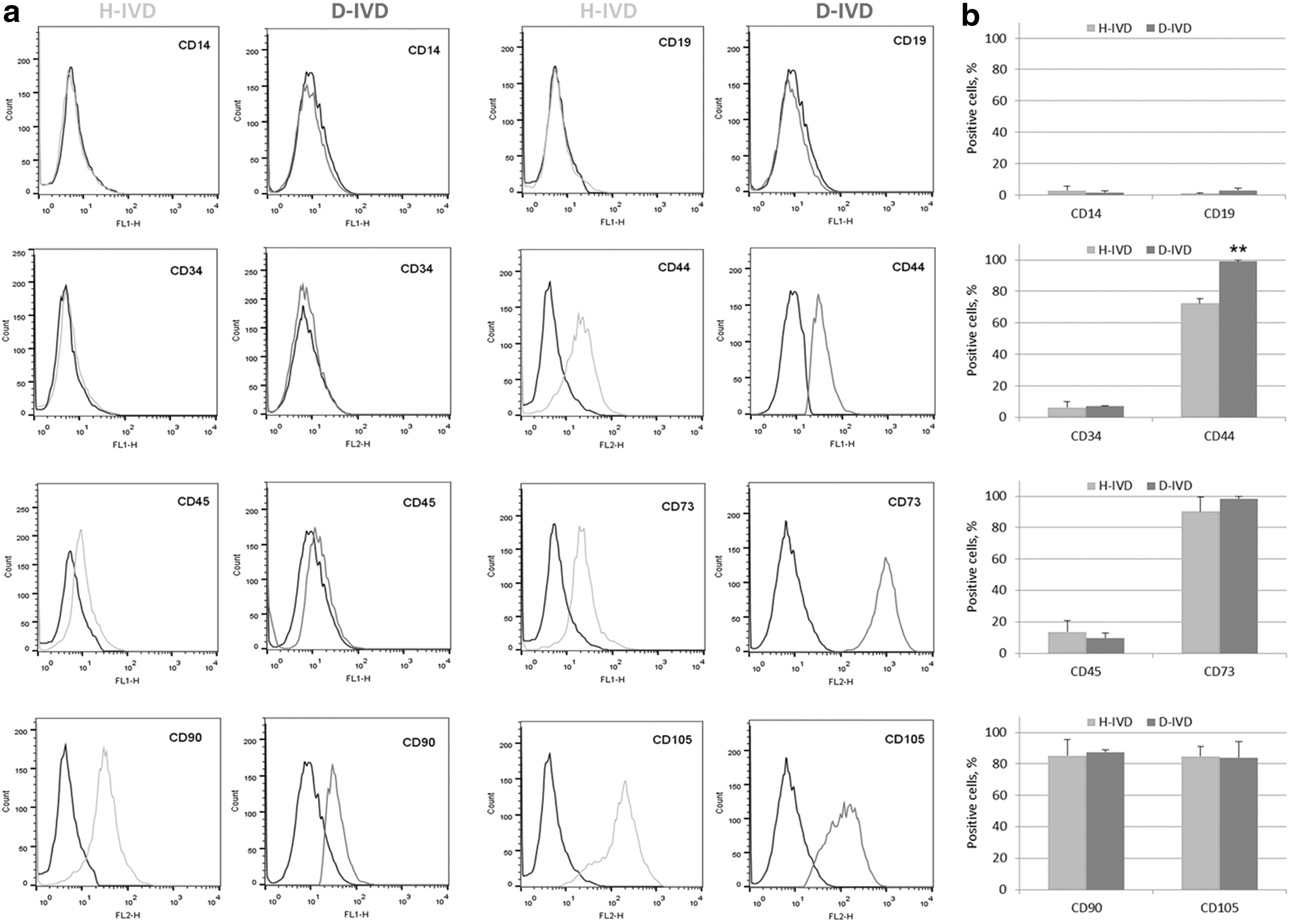
Immunophenotypic profile of MSCs from H-IVD and D-IVD. Results from flow cytofluorimetric analysis of different cell surface markers in MSCs isolated from H-IVD and D-IVD of independent experiments.
H-IVD and D-IVD MSCs differ in gene profile and OPN expression
Growing evidence suggests that the inflammatory process plays an important role in the pathophysiology of both H-IVD and D-IVD [8]. At present, the molecular mechanisms involved in this process remain largely unknown. Therefore, we evaluated the expression profile of 84 key genes involved in immune cascade reactions during inflammation (Supplementary Table S1; Supplementary Data are available online at
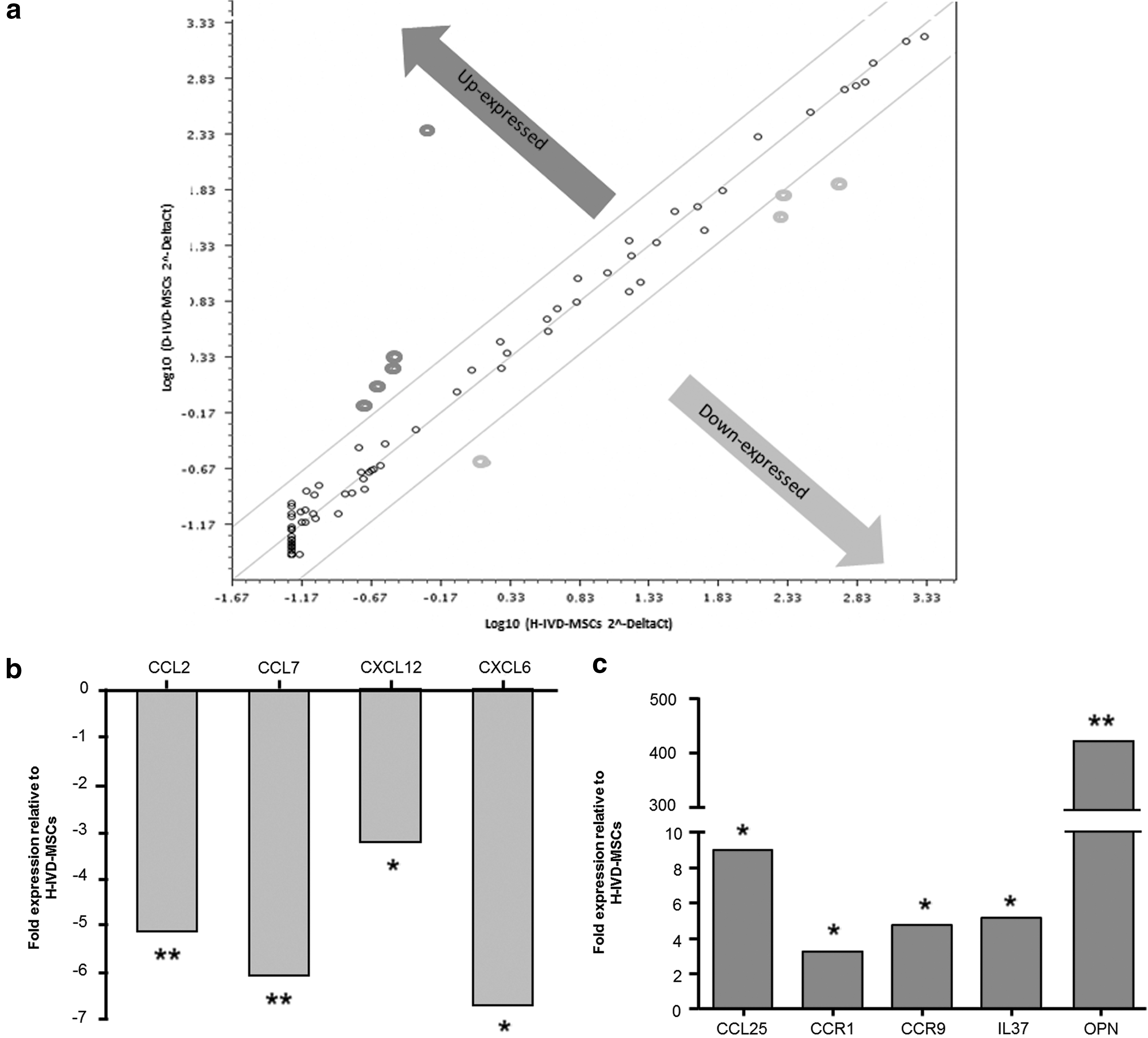
Molecular characterization of H-IVD-MSCs and D-IVD-MSCs. RT2 Profiler PCR array of human inflammatory cytokines & receptors was performed on cDNA from H-IVD-MSCs and D-IVD-MSCs at passage 6–8.
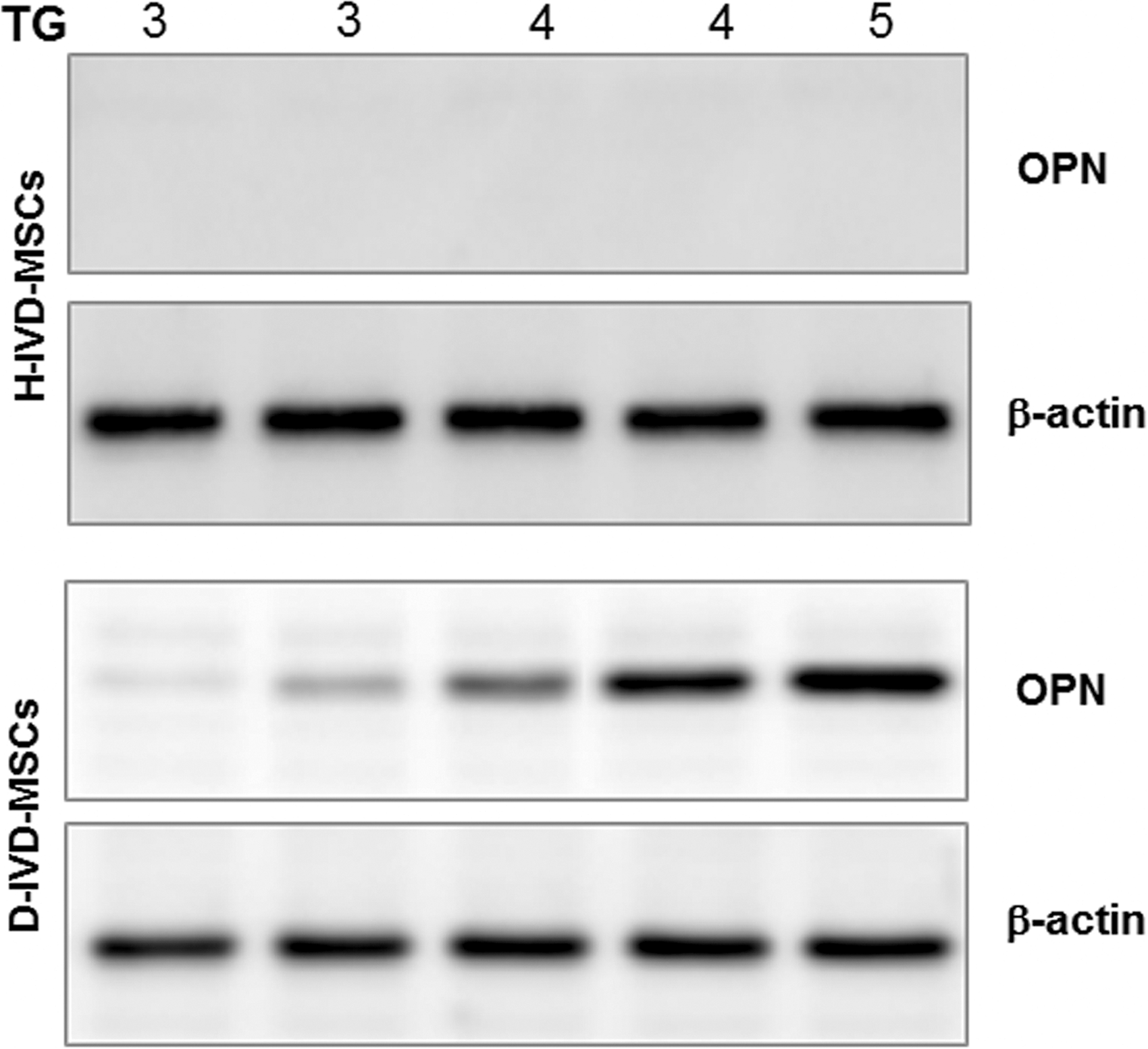
Expression of the OPN protein in H-IVD-MSCs and D-IVD-MSCs. MSCs lysates from H-IVD and D-IVD representative of different TG were analyzed by immunoblotting with anti-OPN and anti-β-actin antibodies. The images are representative of one out of three immunoblottings. OPN, osteopontin; TG, Thompson grades.
Discussion
Lumbar disc degeneration is a multifaceted condition, in which, besides biomechanical factors, aging, genetic, systemic, and toxic factors contribute to influence the degeneration process [12 –14]. Furthermore, IVD is often associated to LBP [15], a chronic debilitating condition, which may have a serious negative impact on the quality of life. The actual treatments for IVD degeneration are mainly conservative and often inconclusive, not restoring full spine function. Thus, to improve the living conditions of patients with disc pathology, there is a need to develop novel effective treatments. A better understanding of disc matrix biology may help determine preventative and therapeutic measures for LBP.
Previous studies on D-IVD and H-IVD specimens were mainly histological [8]. Growing evidence suggests that a subpopulation of MSCs is present in both H-IVDs and D-IVDs, but, up to date, its functional and potential role in degeneration remains unexplored.
In this context, the isolation and expansion of the cellular component of the disc matrix and the study of MSC biology in H-IVDs and D-IVDs may be of relevance to better elucidate tissue arrangements and pathophysiological processes underlying IVD diseases. Despite progenitor cells representing a minor cell population in the IVD matrix, they most likely provide a significant contribution in orchestrating the pathological process. In agreement, Risbud et al. [16] identified a proliferating population of skeletal progenitors within the NP and AF of moderately degenerated human IVDs, and demonstrated that these progenitors express stem cell markers, such as CD90 and CD105, and are able to differentiate in vitro into the three mesodermal lineages. Consistent with these and our previous findings on MSCs isolated from NP of human D-IVDs [9], the present study shows that all the investigated IVD cells express different MSC markers, and, upon appropriate stimulations, exhibit pluripotent differentiation capabilities being able to acquire adipogenic, chondrogenic, and osteogenic markers. All these capabilities were found similar in MSCs from H-IVDs and D-IVDs.
We performed RT2 Profiler PCR array analysis of IVD-MSCs, focusing on inflammatory cytokines and their receptors. It was shown that several cytokines are locally increased in IVD diseases [17] and MSCs were suggested to be implicated in their production [8,18]. In cells isolated from human D-IVDs and H-IVDs, we observed an extensive cytokine and chemokine expression profile, reinforcing the potential of these factors to act as intercellular signaling molecules under pathological conditions. Notably, our results show that the gene expression of CCL2, CCL7, CXCL12, and CXCL6 is downregulated in D-IVD versus H-IVD cells. Furthermore, our data demonstrate an overexpression of CCR1, CCR9, IL37, CCL25, and OPN genes in D-IVD. We cannot exclude that the observed differences between the two cell populations might be influenced by the H-IVD microenvironment, cell population could have been significantly influenced by its microenvironment, and/or contaminating cells from surrounding tissues. Notwithstanding, different alterations observed in H-IVD cells appear important and deserve some comments. First, it has been demonstrated that IL37 has an anti-inflammatory effect by regulating excessive inflammatory responses through a feedback loop that contributes to restore the cytokine balance in various chronic inflammatory conditions [19]. Then, it was reported that CCL25, but not CCL2, represents a good candidate for the recruitment of human subchondral mesenchymal progenitors in synovial fluid of patients affected by cartilage defects [20]. Thus, the increased expression of IL37 and CCL25 by MSCs may represent an endogenous effort during the pathophysiological process in which the damaged cartilage in the D-IVD appears to counteract tissue degeneration.
Intriguingly, when compared to H-IVD-MSCs, D-IVD-MSCs showed a significant overexpression of OPN, a protein involved in both cell attachment and calcification of mineralized tissue [21,22]. Since the IVD is primarily an avascular tissue, it is likely that the inflammatory cytokines and chemokines produced by disc cells are not cleared from the tissue and remain there with time. Thus it appears that D-IVD-MSCs reside in a proinflammatory cytokine-rich microenvironment, which might favor OPN gene expression and protein synthesis. The OPN increase found in D-IVD cells leads us to speculate that this alteration might favor degenerative changes in D-IVD-MSCs, possibly through mineralization-inducing conditions, as it occurs in AF cells [23]. In agreement, a very recent article suggested that AF cells have characteristics of progenitor cells and, under appropriate stimuli, are capable of differentiating into chondrocytes and osteoblasts both in vitro and in vivo [24].
Intriguingly, we found that OPN level in D-IVD cells is directly related to the IVD Thompson grade. Although more studies are necessary to confirm this possibility, this finding suggests that OPN represents a potential marker of IVD degeneration and that its level might be an indicator of degeneration severity in patients affected by D-IVD disease.
It is well known that OPN acts through the interaction with CD44, an important mediator in chondrocyte cell–matrix interactions, which involve proteoglycan and hyaluronan link protein aggregates [25]. Noteworthy, besides OPN, our results show that also CD44 expression is enhanced in D-IVD-MSCs. It was reported that OPN is able to induce tissue mineralization through CD44 interaction as well as that after integrin binding, it prompts disc matrix degradation through the activation of some MMPs [26]. Thus, it appears likely that high levels of OPN together with those of CD44 might exacerbate ECM degeneration and/or mineralization.
Of relevance, the results of this study implicate for the first time the expression of OPN in the IVD degeneration process. Since OPN is involved in several pathophysiological pathways, including inflammatory and immune response, as well as in pathological disc mineralization, the correction of this and other biochemical imbalances within the degenerated disc might facilitate the recovery and maintenance of normal disc morphology, thereby improving the biomechanical function of the disc and ultimately altering the pathological course of this disease. Further studies are necessary to clarify the OPN role in IVD degeneration. These will hopefully offer new chances to identify novel molecular targets for conservative therapies, and be of significance in autologous IVD cell transplantation approaches.
Footnotes
Acknowledgments
This work was supported by IRCCS Foundation Ca’ Granda Ospedale Policlinico Milan, and University of Milan, Italy.
Some of the data reported in this article were presented to the XX Cardiac Growth and Regeneration Congress, 22th–25th June, 2014, Viterbo, Italy.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.