
Abstract
Blastocyst formation represents the first lineage specification by segregation of the trophectoderm from the inner cell mass in early embryonic development. Transcriptional regulation of Cdx2, which is selectively expressed in and essential for the specification of trophectoderm, has been extensively studied. However, post-transcriptional regulation of Cdx2 remains largely unknown. In this study, we report that RNA-binding protein motif 46 (Rbm46), an RNA-binding motif protein with unknown function, directly binds to and stabilizes Cdx2 mRNA in early mouse embryos. In addition, knockdown of Rbm46 using RNA interference downregulated the majority of trophectoderm markers in mouse embryonic stem cells and blocked the allocation of blastomere cells to the trophectoderm in mouse embryos. Our study revealed a novel mechanism by which Rbm46 regulates trophectoderm specification through stabilizing Cdx2 mRNA in early mouse embryos.
Introduction
T
It has been proposed that the reciprocal expression of Cdx2 in the outer and Oct4 in the inner blastomeres of the blastocyst underlies the molecular basis for the segregation of trophectoderm from the ICM [6,10]. Mechanistically, in later morula stages, cell–cell interaction leads to Hippo signal-mediated phosphorylation and subsequent degradation of Yes-associated protein (YAP) in the inner blastomeres, whereas in the outer blastomere cells, nonphosphorylated YAP enters into the nucleus and facilitates TEAD4, a TEA domain/transcription enhancer factor, to activate Cdx2 expression [11 –13]. Subsequently, Cdx2 then interacts with other proteins, such as Brg1, a chromatin remodeling protein, to induce CpG methylation at the Oct4 promoter, thereby blocking Oct4 expression in the outer blastomeres [14]. As such, Cdx2 preferentially localizes at the apical side of the outer blastomeres and separates trophectoderm lineage from the ICM [15]. While the dynamic expression and functional role of Cdx2 are known to be critical for the segregation of trophectoderm from ICM, it remains unclear whether and how Cdx2 mRNA is regulated before its translation. For example, it has been noted that expression of Cdx2 mRNA varies from 2-cell to 8-cell stages as reported from different laboratories [15 –18], but Cdx2 protein cannot be detected until 8-cell to morula stages [15 –18]; we had a similar observation in our own studies (Fig. 1A, column 5 from the left). Our results were consistent with the previously reported fact that Cdx2 mRNA can be detected days earlier than the protein can [15 –18]. These observations raised a possibility of a post-transcriptional control of Cdx2 in the outer blastomeres.
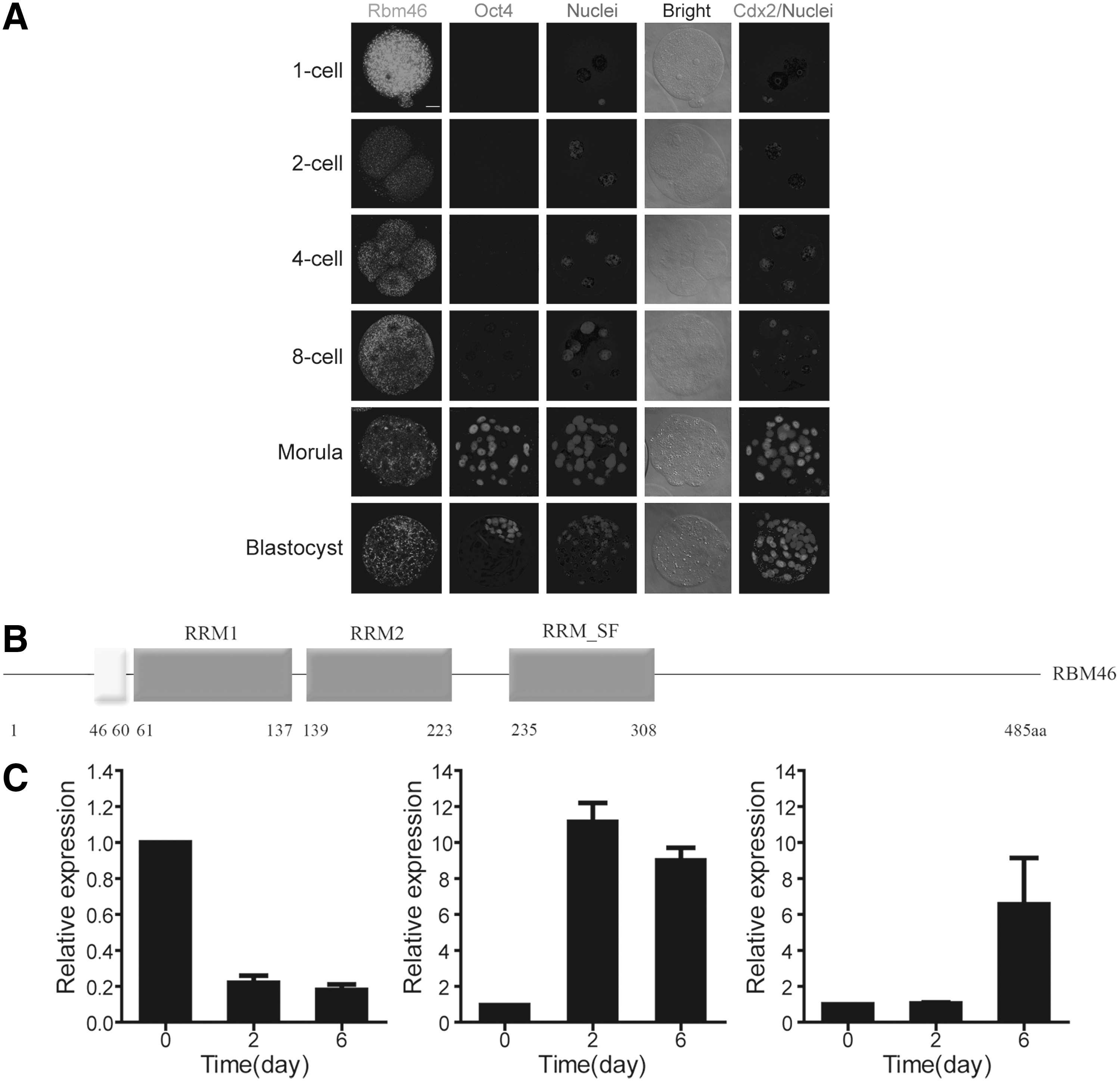
RNA-binding protein motif 46 (Rbm46) expression before Cdx2 expression at the early stage of mouse embryo development.
Multiple lines of evidence indicate that post-transcriptional regulation by RNA-binding proteins (RBPs) is involved in embryonic stem cells (ESCs) fate specification [19 –21]. For instance, it was reported that an RBP Elavl1/HuR regulates extraembryonic trophoblast late-stage differentiation probably by stabilizing Tbx4 mRNA and controls the placental labyrinth formation in the developmental embryo [22]. However, there has thus far been no report regarding the regulation of trophectoderm specification by RBPs.
ESCs, derived from in vitro culture of the ICM of blastocysts, have often been used for studying trophectoderm differentiation [7], as it can be converted into trophectoderm cells by altering the expression of transcription factors [7,23]. Using this system, we sought to search for RBPs expressed in ESCs that may be involved in the post-transcriptional regulation of trophectoderm specification.
RNA-binding protein motif 46 (Rbm46) is one of the seven RBPs in the list of ESC-enriched factors [24]. Among these seven RBPs (Supplementary Table S1; Supplementary Data are available online at
In this report, we investigated the expression pattern of Rbm46 and tested our hypothesis that Rbm46 may be involved in the post-transcriptional regulation of Cdx2 in the early embryonic development.
Materials and Methods
Cell culture
E14Tg2a mouse ESCs were cultured without feeders on plastic coated with 0.1% gelatin and replated every 3 days at a split ratio of 1 in 10 following dissociation with 0.25% trypsin (Gibco; Cat. No. 25200-056). Cells were cultured in the DMEM supplemented with 15% (v/v) fetal calf serum (FCS; Hyclone, Logan, UT,
Embryo collection and culture
Embryos were collected into the M2 medium with 4 mg/mL bovine serum albumin (BSA) (M2+BSA) from F1 (C57Bl6×CBA) females superovulated with 10 IU of PMSG (Intervet) and 10 IU of hCG (Intervet) 48 h later and mated with F1, C57 mice. Embryos were cultured in KSOM with 4 mg/mL BSA (KSOM+BSA) under paraffin oil in 5% CO2 at 37.5°C. Animal handling and procedures were approved by the Peking University Health Science Center Institutional Animal Care and Use Committee.
Plasmids
Mouse Rbm46 individual full-length coding sequence was amplified from mouse testis mRNA and cloned into the pPyCAGIP vector with the following primers: 5′-ATCGCGCTCGAGATGGACTACAAGGACGACGATGACAAGATGAATGAAGAAAACACTGATGGTAC-3′ and 5′-ATATGCGGCCGCTCAGAAGAATGAGGCCTGATTG-3′. Modified pll3.7 plasmid (GFP fragment was replaced with puromycin-resistant gene) was employed to produce short hairpin nucleotides. Nucleotides targeting the firefly luciferase gene, which has no homology with mouse transcriptome, served as control short hairpin RNA (shRNA). The RNAi oligonucleotides sequences used are as follows: Rbm46 (siRbm46): 5′-TCCAACAATATCACTTGCTATTCATAGAGATAGCAAGTGATATTGTTGGTTTTTTC-3′. All constructs were verified by sequencing. To generate overexpression or knockdown cells, E14Tg2a ESCs were transfected separately with pPyCAGIP or shRNA constructs using Lipofectamine 2000 and 1 μg/mL of puromycin was added at 48 h post-transfection. About 10 days later, stably transfected cells were expanded and then stored for further analysis.
RNA isolation, reverse transcription, and quantitative real-time polymerase chain reaction
RNA was isolated using Trizol (Invitrogen). Reverse transcriptase (Promega) was employed for oligo (dT) primed first-strand cDNA synthesis. Real-time polymerase chain reaction (PCR) was carried out on the SYBR Premix Ex Taq II detection system (Takara) using SYBR dye. The ΔΔCt method was used to comparatively quantify the amount of mRNA level. Primers are listed in Supplementary Table S2.
Protein extraction, western blot
Cells were lysed in cold lysis buffer (20 mM Tris–HCl, pH 7.5, 150 mM NaCl, 0.5% Triton X-100, 12.5 mM β-glycerophosphate, 1.5 mM MgCl2, 2 mM ethylene glycol tetraacetic acid, 10 mM NaF, and 1 mM Na3VO4) containing protease inhibitors (Roche). Western blot was carried out by standard procedures; primary antibodies used for detection were anti-Oct4 (1:1,000; Santa Cruz, sc-365509), Nanog (1:1,000; Cosmo BioCo, REC-RCAB0001P), Sox2 (1:500; Santa Cruz, sc-20088). Proteins were visualized with an Odyssey Two-Color Infrared Imaging System (LI-COR Biosciences) according to the manufacturer's instructions.
Immunofluorescence microscopy
Cells on coverslips were fixed in cold methanol for 15 min. After three washes in phosphate-buffered saline (PBS), cells were blocked in PBS containing 5% BSA for 30 min and then incubated with mouse or rabbit primary antibodies in PBS containing 2.5% BSA for 2–12 h at 37°C, anti-stage-specific embryonic antigen-1 (SSEA-1) (1:50; Millipore), Oct4 (1:500; Santa Cruz, sc-365509), Nanog (1:1,000; Cosmo BioCo, REC-RCAB0001P), Cdx2 (1:50; Santa Cruz, sc-166830), and Gata6 (1:1,000; Abcam, ab22600). After three washes, cells were incubated with fluorescein isothiocyanate-conjugated α-mouse and PE-conjugated α-rabbit secondary antibodies for 1 h at 37°C and then with 4′, 6-diamidino-2-phenylindole (Roche) for 15 min. The coverslips were washed extensively and mounted onto slides. Imaging of the cells was carried out using the Zeiss LSM 510 Meta Confocal Microscope.
Flow cytometry
One million cultured cells per sample were dissociated with 0.25% trypsin (Gibco; Cat. No. 25200-056). Cells were then resuspended in 1:3 volumes of ice-cold PBS and 100% ethanol and then incubated on ice for 30 min. After centrifugation, the cell pellet was washed 3× with PBS and centrifuged for 5 min. Cells were then incubated in the dark in 1 mL PBS containing RNase (12.5 g/mL; Tiangen) for 30 min and propidium iodide (3 g/mL; Sigma) and analyzed by flow cytometry. Analyses were performed by flow cytometry (FACSCalibur; BD Biosciences) using CellQuest software (BD Biosciences) and the ModFit software (Verity Software House).
RNA immunoprecipitation
RNA immunoprecipitation (RIP) experiments were performed using the Magna RIP RNA-Binding Protein Immunoprecipitation Kit (Millipore; Cat No. 17-700) according to the manufacturer's instructions. The Anti-Flag M2 Magnetic Beads (Sigma; Cat No. M8823) were used for RIP. The coprecipitated RNAs were detected by reverse-transcription PCR (RT-PCR). Total RNAs (input controls) and isotype controls were assayed simultaneously to demonstrate that the detected signals were from RNAs specifically binding to Rbm46 (n=3 for each experiment).
mRNA half-life measurements
E14Tg2a mouse ESCs were transfected with vectors, and 48 h later, the cells were treated with actinomycin D (2 μg/mL) and were harvested in Trizol reagent (Invitrogen) at different time points (0, 0.5, 1, 2, 4 h). mRNA half-lives were calculated by real-time PCR and normalized against 18S rRNA and then different mRNA half-lives were calculated relative to control samples.
Injection of mRNA and dsRNA for Rbm46 and Cdx2
A full-length ORF Rbm46 or Cdx2 DNA construct was cloned into modified pcDNA3.1 plasmid (enhanced green fluorescent protein [EGFP] fragment was inserted between BamH1 and EcoR1). To overexpress Rbm46 or Cdx2, one blastomere was injected with mRNA for Rbm46 or Cdx2 (0.5 μg/μL) and EGFP (0.3 μg/μL) or, in controls, EGFP alone using mMESSAGE mMACHINE T7 Ultra Kit (Ambion). To downregulate Rbm46 or Cdx2, dsRNA was synthesized in vitro using T7 RNA polymerase (Ambion). A PCR using the construct described above, using primers specific to T7 RNA polymerase core sequence, synthesized the Cdx2 template primer as reported [17]. Primer dsRbm46 T7-F: GAATTAATACGACTCACTATAGGGAGAGAGGAGACAATTAAAGCAGAGTTCA; primer dsRbm46 T7-R: GAATTAATACGACTCACTATAGGGAGATCCCATCCTGACTGGTTGTT. Embryos were cultured at different stages and then observed under an inverted confocal microscope.
Statistics
Student's t-test was used for a comparison of two independent treatments. The two-way ANOVA statistical test was used for the analysis of CCK8 and mRNA half-life measurements results. The Mann–Whitney test was used to analyze the embryo injection data. For all tests, a P value of <0.001 was considered very significant (***).
Results
Rbm46 expression before Cdx2 expression at the early stage of mouse embryo development
Rbm46 contains three RNA recognition motifs (Fig. 1B) and is well conserved among different species. Mouse Rbm46 is expressed in zygotes and is continually present in embryos up to the blastocyst stage (Fig. 1A). As a comparison, neither Oct4 protein nor Cdx2 protein could be detected until the 8-cell stage or later (Fig. 1A). Maternal Rbm46 mRNA was detectable in zygotes, and expression levels were decreased in 2-cell stage embryos (data not shown). In addition, a burst of expression in Rbm46 activation was observed between the 4-cell and morula stages, which was earlier than the detection of Cdx2 during the early embryo development.
When we differentiated mouse ESCs in embryoid bodies (EBs) in vitro, 2 days after withdrawing LIF from the culture medium, Oct4 expression significantly declined (Fig. 1C), we found that Rbm46 expression was upregulated at day 2 and remained at a high level up to day 6. Interestingly, the Cdx2 expression was lagging and could not be detected until day 6 (Fig. 1C).
Based on these interesting expression patterns of Rbm46 versus Cdx2 in both embryonic development in vivo and mESCs differentiation in vitro, and taking into account the fact that Cdx2 mRNA is detected earlier than the protein [15], we next investigated the possibility that Rbm46 may be involved in the post-transcriptional regulation of Cdx2. As an RBP, Rbm46 may relate to a lot of genes, including Cdx2, and we therefore have an interest in the possible relation between these two genes.
Ectopic expression of Rbm46 promotes Cdx2 expression in mouse ESCs
To investigate the effect of Rbm46 on ESCs, a flag-tagged Rbm46 expression construct containing the puromycin-resistance gene was transfected into ESCs. After selection for puromycin-resistant cells for 10 days, the Rbm46-transfected cells (PuroMycin-resistant Rbm46-transfected cells: PM46 cells) were analyzed in comparison with ESCs transfected with an empty plasmid as a control (plasmid empty-transfected cells: PE cells). As expected, PM46 cells had a significant increase in Rbm46 expression compared with PE cells in both mRNA (Fig. 2A) and the protein level (Fig. 2B). We performed alkaline phosphatase (ALP) staining, an indicator for undifferentiated ESCs and found that control PE cells were almost 100% ALP-positive and closely compacted (Fig. 2C), a morphology of typical undifferentiated mouse ESCs. In contrast, PM46 cells were barely stained with ALP and grew in a loosely flat shape (Fig. 2C). This observation suggests that PM46 cells had undergone differentiation. We also conducted an apoptotic assay using the CCK-8 counting assay and showed that PM46 cells had significantly lower counts than PE cells (Fig. 2D, circled curve), ruling out the induction of cell death by forced Rbm46 expression. This was further confirmed by Annexin V staining and we did not find a difference between PM46 and PE cells (Supplementary Fig. S1A). We then analyzed cell proliferation and found that there was 8% less S-phase cells in PM46 than in PE cells (Supplementary Fig. S1B, Student's t-test, P<0.05).
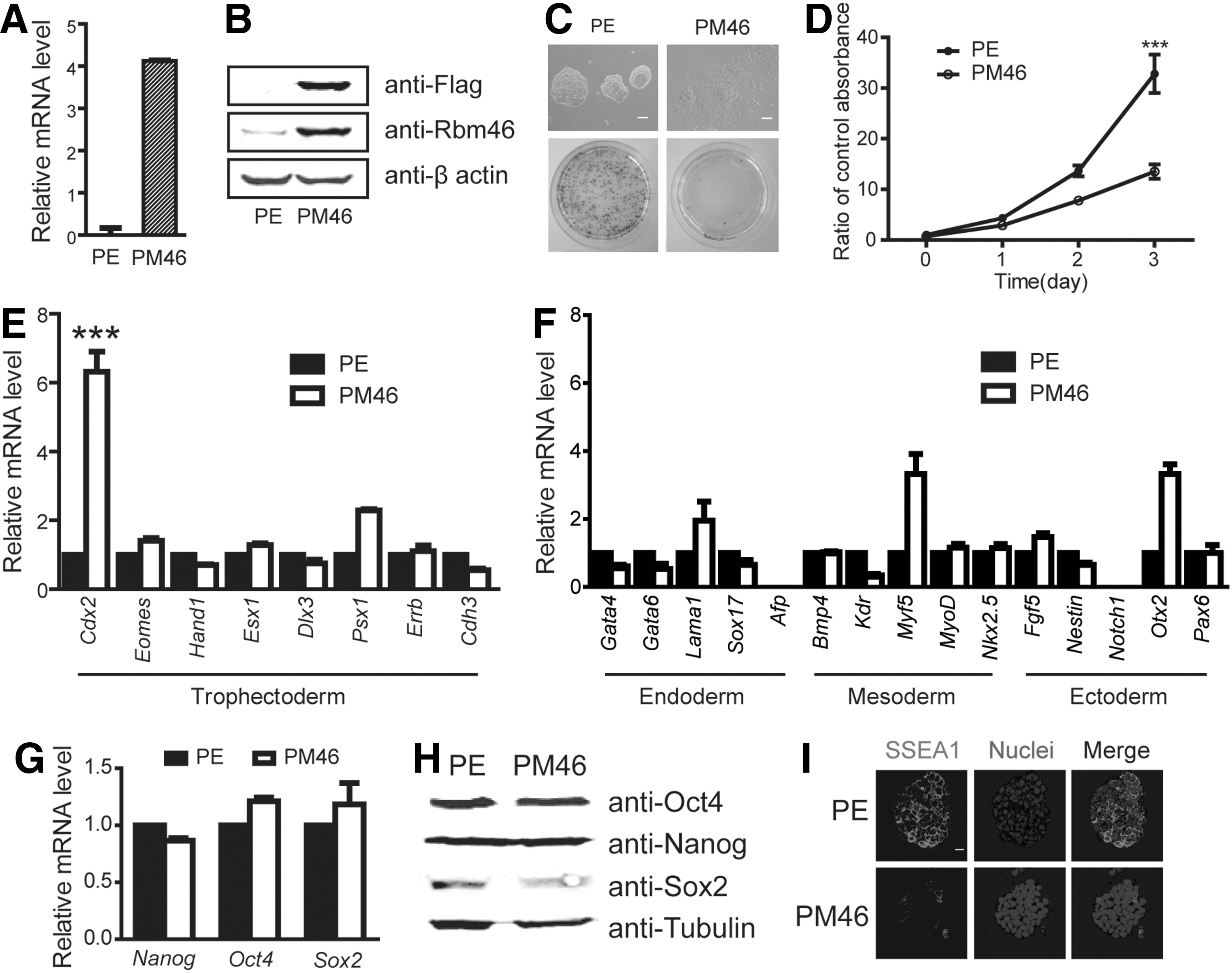
Ectopic expression of Rbm46 promotes Cdx2 expression.
This reduction of cell proliferation suggests a differentiation potential in PM46 cells compared with that in PE cells. We then compared the gene expression profiles between PM46 and PE cells and found that, among trophectoderm markers, Cdx2 was upregulated significantly in PM46 cells (Fig. 2E). We also observed that an endoderm marker Lam1, a mesoderm marker Myf5, and an ectoderm marker Otx2 were increased in PM46 cells compared with those in PE cells, but these markers were not significantly compared with Cdx2 levels (Fig. 2F). We also measured the expression of pluripotent factors, including Nanog, Oct4, and Sox2. To our surprise, none of the three factors was significantly changed in PM46 versus PE cells at the mRNA (Fig. 2G) or the protein level (Fig. 2H). However, the expression of SSEA-1 was substantially reduced in PM46 cells compared with that observed in PE cells (Fig. 2I). These data suggested that Rbm46 may prime ESCs for differentiation, but the pluripotent network, including Oct4, Nanog, and Sox2 still preserves the cells in pluripotency in the culture medium containing the LIF factor. In supporting this notion, we further found that PM46 cells were still able to form teratoma-containing ectoderm, mesoderm, and endoderm cells after transplantation into SCID-nude mice (Supplementary Fig. S2C), although the size of teratoma formed from PM46 was significantly smaller than that formed in PE cells (Supplementary Fig. S2A, B).
Knockdown of Rbm46 expression downregulates the majority of trophectoderm markers
To test whether Rbm46 function is involved in lineage differentiation, we knocked down Rbm46 expression in mouse ESCs (siRbm46 cells) using shRNA. Compared with the nonsilencing control ESCs, Rbm46 expression was reduced significantly in siRbm46 cells as measured by real-time PCR (Fig. 3A) and immunoblotting (Fig. 3B). Unlike the morphology of nonsilenced ESCs, siRbm46 cells were surrounded by skirt-shaped cells with a clear cell–cell boundary (Fig. 3C). The results showed that the majority of trophectoderm markers were downregulated in siRbm46 compared with the control cells (Fig. 3D). Interestingly, we observed that the endoderm markers Gata6, Lama1, and Sox17 were upregulated in siRbm46 cells, and similarly, the mesoderm markers Myf5, MyoD, and Nkx2.5 were also increased in siRbm46 cells (Fig. 3E). However, none of these pluripotent markers (Nanog, Oct4, and Sox2; Fig. 3F, G), apart from SSEA-1 (Fig. 3H), was changed in siRbm46 cells.
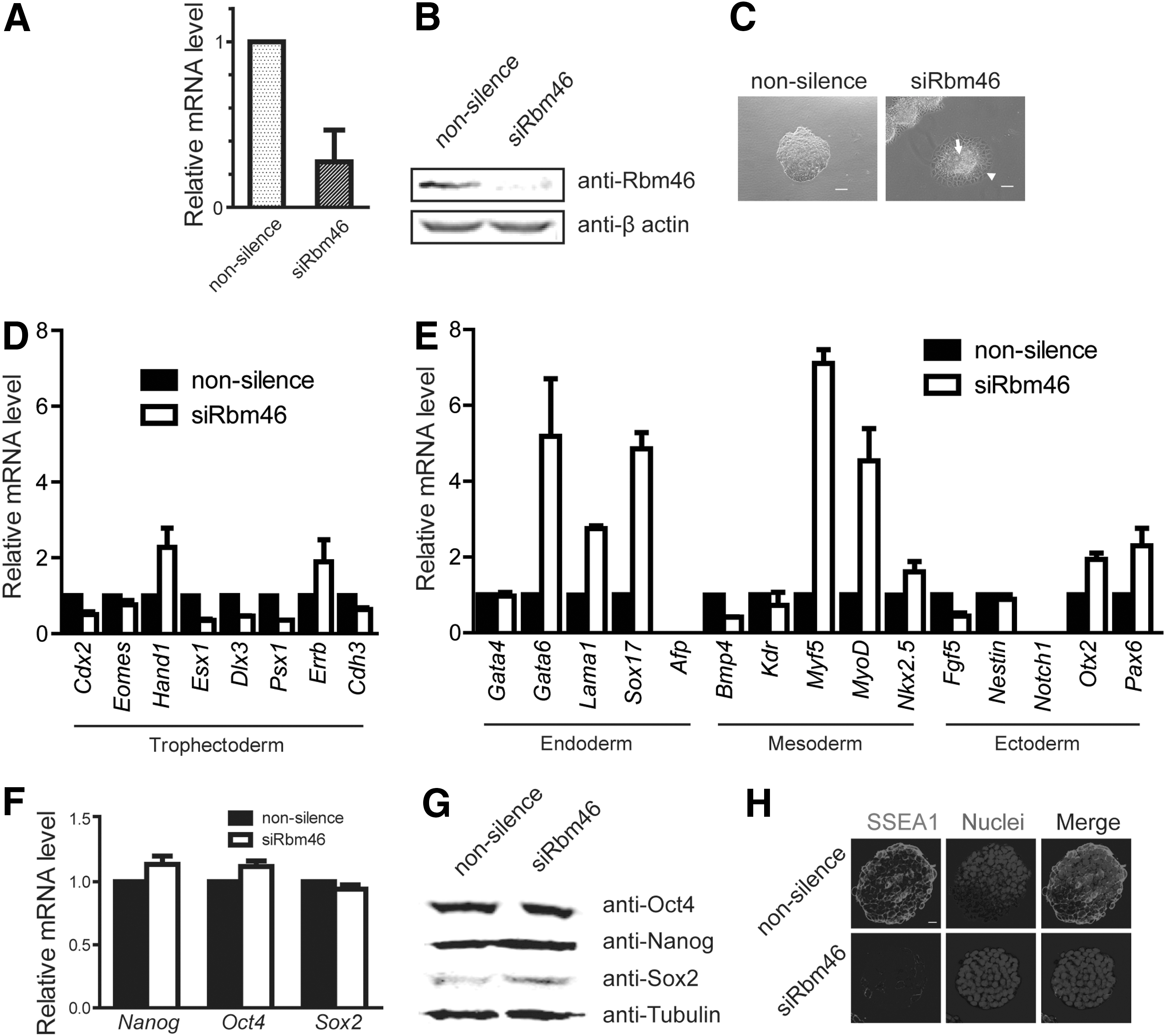
Knockdown of Rbm46 expression downregulates the majority of trophectoderm markers.
Rbm46 regulates ESCs differentiation into TE lineage
The gene profiling results suggested that Rbm46 might modulate Cdx2 expression in ESCs. To test this, we performed immunofluorescence analysis for Cdx2 protein level in PM46 cells (compared with control PE cells, Fig. 4A) and siRbm46 (compared with nonsilenced cells, Fig. 4B). PE cells did not show any Cdx2-staining, whereas PM46 cells showed positive staining for Cdx2 around the perimeter of undifferentiated colonies (Fig. 4A). Again, pluripotent factors remained positive in both PM46 and siRbm46 cells (Supplementary Fig. S3A, B). All these data suggested that Rbm46 is involved in the regulation of trophectoderm differentiation. But the above-described studies of Rbm46 using cell culture approach were carried in the LIF-dependent medium that favors ESC self-renewal by promoting pluripotent factor expression (Figs. 2G, H and 3F, G). To test the hypothesis that Rbm46 is involved in the regulation of trophectoderm differentiation, we used in vitro-induced differentiation of mouse ESCs to the TE lineage [16]. As expected, when the PM46 cells and siRbm46 cells were cultured in the presence of FGF4, heparin, and MEF-conditioned medium, PM46 cells gave rise to quite large and flat TE-like epithelial cells compared with PE cells (Fig. 4C). In contrast, siRbm46 maintained completely undifferentiated colonies with smooth edges, which was slightly different from nonsilence cells (Fig. 4D). Among these groups, only PM46 cells showed a large number of the enlarged or multiple nuclei and nucleus expression of Cdx2 (Fig. 4E, F), which was never detected in the other three groups. Real-time quantitative polymerase chain reaction analyses revealed TE lineage marker gene regulation by Rbm46, using a TS cell line 129R3 as a control [26] (Fig. 4G). These results showed that Rbm46 regulated ESCs differentiation into the TE lineage mainly by regulating Cdx2.
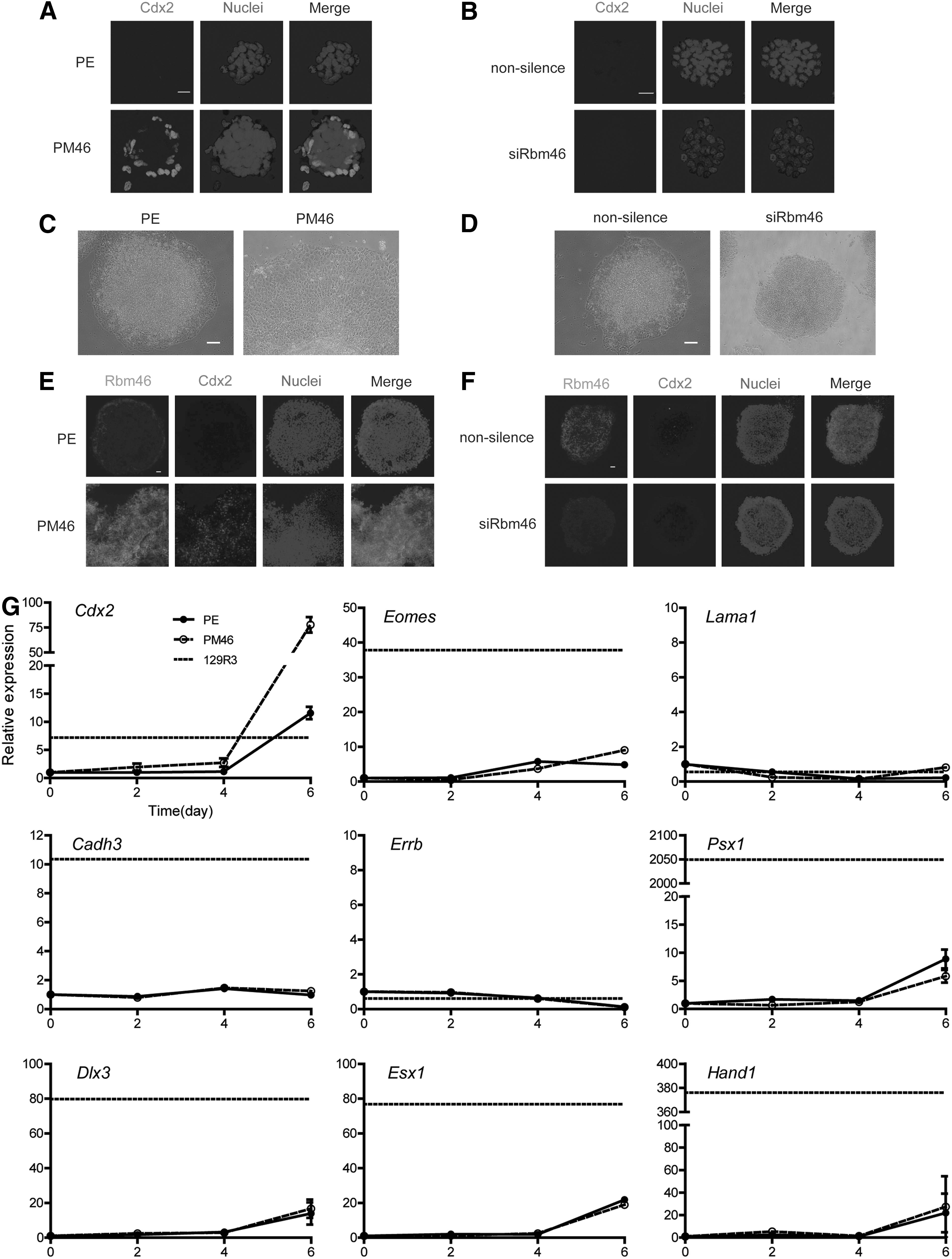
Rbm46 regulates ESCs differentiation into TE lineage.
Rbm46 stabilizes Cdx2 mRNAs
To address how Rbm46 regulates Cdx2 production, we next tested our hypothesis that Rbm46, as an RBP, regulates Cdx2 production by stabilizing Cdx2 mRNA. We first immunoprecipitated flag-tagged Rbm46 protein from PM46 cells, extracted total RNA from the immunoprecipitated complex, and then subjected this to reverse transcription as well as subsequent RT-PCR amplification. As shown in Figure 5A, Cdx2 mRNA, but not Oct4 mRNA, can be reverse-transcribed from the immunoprecipitated complex in mouse ESCs. Furthermore, real-time PCR analysis indicated that Cdx2 mRNA increased significantly in PM46 cells than in PE cells (Fig. 5B), whereas Oct4 expression did not show any differences between PM46 and PE cells (Fig. 5B). We also measured the half-life to reflect the decay of Cdx2 mRNA in PM46 and PE cells, respectively, after adding actinomycin D in the cells to inhibit de novo transcription. We calculated that the Cdx2 half-life in PM46 was almost 3-fold longer than that in PE cells (Fig. 5C, two-way ANOVA statistical test, P<0.05). As a comparison, there was little change in the decay time of Oct4 mRNA between the two types of cells (Fig. 5C). These results indicated that Rbm46 binds directly to Cdx2 mRNA and stabilizes Cdx2 mRNA in PM cells, revealing the molecular mechanism regarding how Rbm46 regulates and stabilizes Cdx2 mRNA.
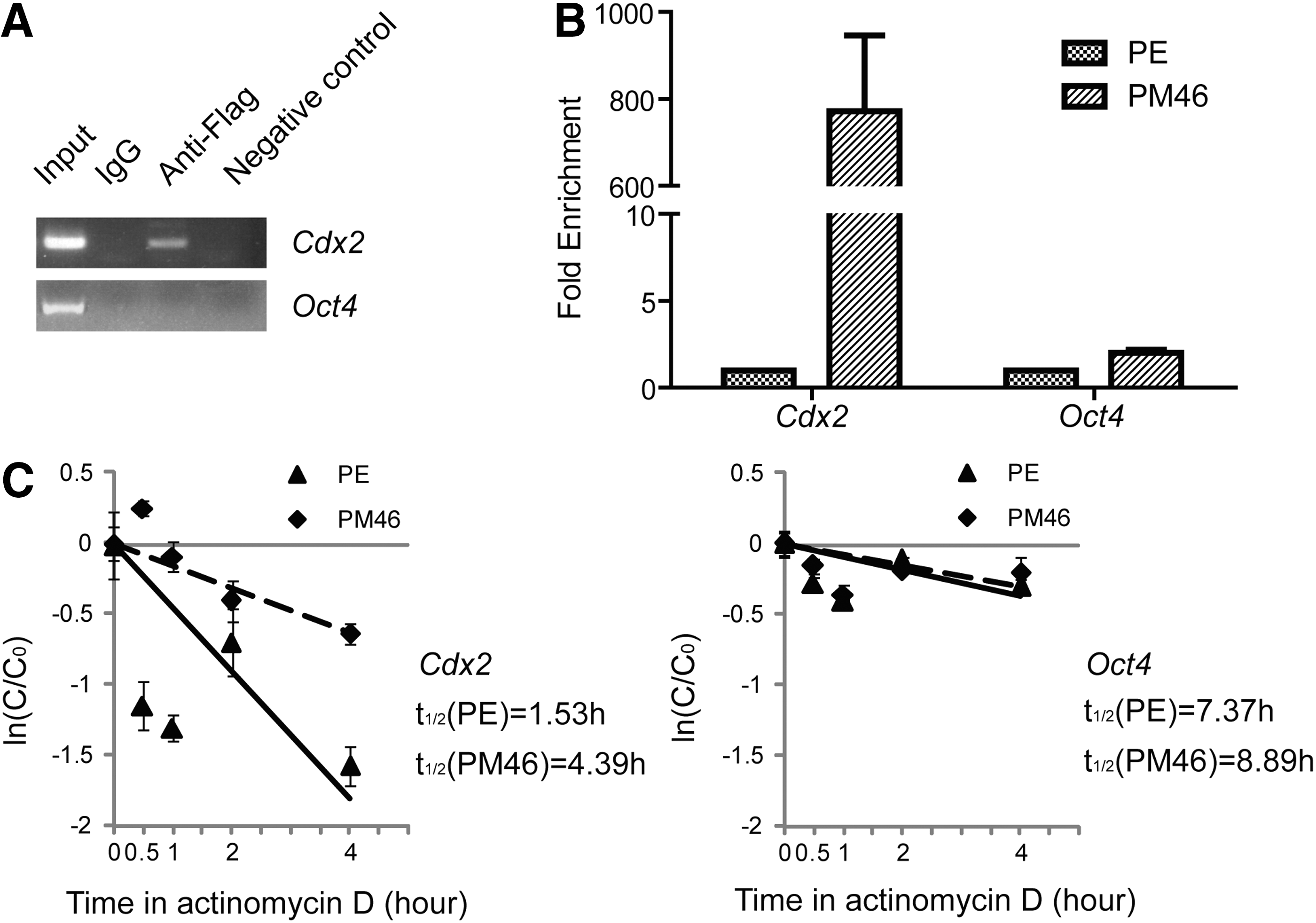
Rbm46 stabilizes Cdx2 mRNAs.
Rbm46 regulates trophectoderm specification in mouse embryos
Based on the observation in mouse ESCs, we sought to investigate whether Rbm46 indeed plays a role in regulation of trophectoderm specification in early mouse embryos. Embryo injection experiments were performed in three independent repetitions for every group (n=20–30), and we injected the Rbm46 mRNA along with EGFP mRNA into single blastomere of a 2-cell stage embryo. When the embryos developed into blastocysts, we monitored the allocation of EGFP positive cells to the ICM or to the surrounding trophectoderm to reflect a role of Rbm46 in regulation of trophectoderm specification. In this experimental system, EGFP mRNA was coinjected as a tracer, and the embryo was immunostained with anti-Oct4 antibodies for ICM identity [17]. When analyzed at the 4-cell and 8-cell stages, embryos injected with Rbm46 mRNA (Supplementary Fig. S4A, B) had the same effect on the allocation of EGFP positive cells. We used embryos injected with Cdx2 mRNA as a positive control or with the negative control of EGFP mRNA alone (Supplementary Fig. S4A, B). Similarly, at both the 4-cell and 8-cell stages, knockdown of Rbm46 or Cdx2 did not affect the EGFP positive cell distribution (Supplementary Fig. S4C, D). At the blastocyst stage, however, embryos injected with Rbm46 showed a significant increase in the allocation of the EGFP positive cells to the trophectoderm, whereas Oct4-positive ICM cells did not see EGFP positive signal (Fig. 6A). The distribution of EGFP green cells observed in the embryos injected with Rbm46 mRNA was similar to that observed in embryos injected with Cdx2 mRNA (Fig. 6A). To further confirm this observation, we showed nine individual confocal sections of the blastocysts injected with either Rbm46 mRNA or Cdx2 mRNA (Fig. 6B) and found that all EGFP green cells were restricted to trophectoderm cells but not to the ICM. We have quantified the in vivo embryo data; embryos were then grouped into EGFP-positive cell distribution in the trophectoderm (named Trophectoderm) and EGFP-positive cell distribution in both the trophectoderm and ICM (named Trophectoderm+ICM), as shown in Supplementary Figure S4E, and the Mann–Whitney test was used to analyze the differences between each group and the EGFP mRNA-injected group (negative control group). Elevating Rbm46 expression significantly increased the proportional contribution of the clone to the trophectoderm compared with the noninjected blastomere of the same embryo (n=61 embryos, Mann–Whitney test, P<0.001) (Fig. 6E). Similarly, 90.3% of the embryos from the Cdx2-injected group have EGFP green cells contributing to trophectoderm, while 9.7% contributed to both trophectoderm and ICM (n=62 embryos, Mann–Whitney test, P<0.001). There was no significant difference between the above two groups (Mann–Whitney test, P=0.976) (Fig. 6E). This observed increase in contribution to the trophectoderm and decrease in contribution to the ICM was not due to the injection procedure itself because the injection of EGFP mRNA alone did not lead to any significant changes in the contribution to specific lineages. Thus, 36.3% of the EGFP-expressing embryos contributed to the trophectoderm and 63.6% of both the trophectoderm and ICM (n=66 embryos) (Fig. 6E). All these data indicated that Rbm46 is sufficient to promote trophectoderm specification in mouse embryos.
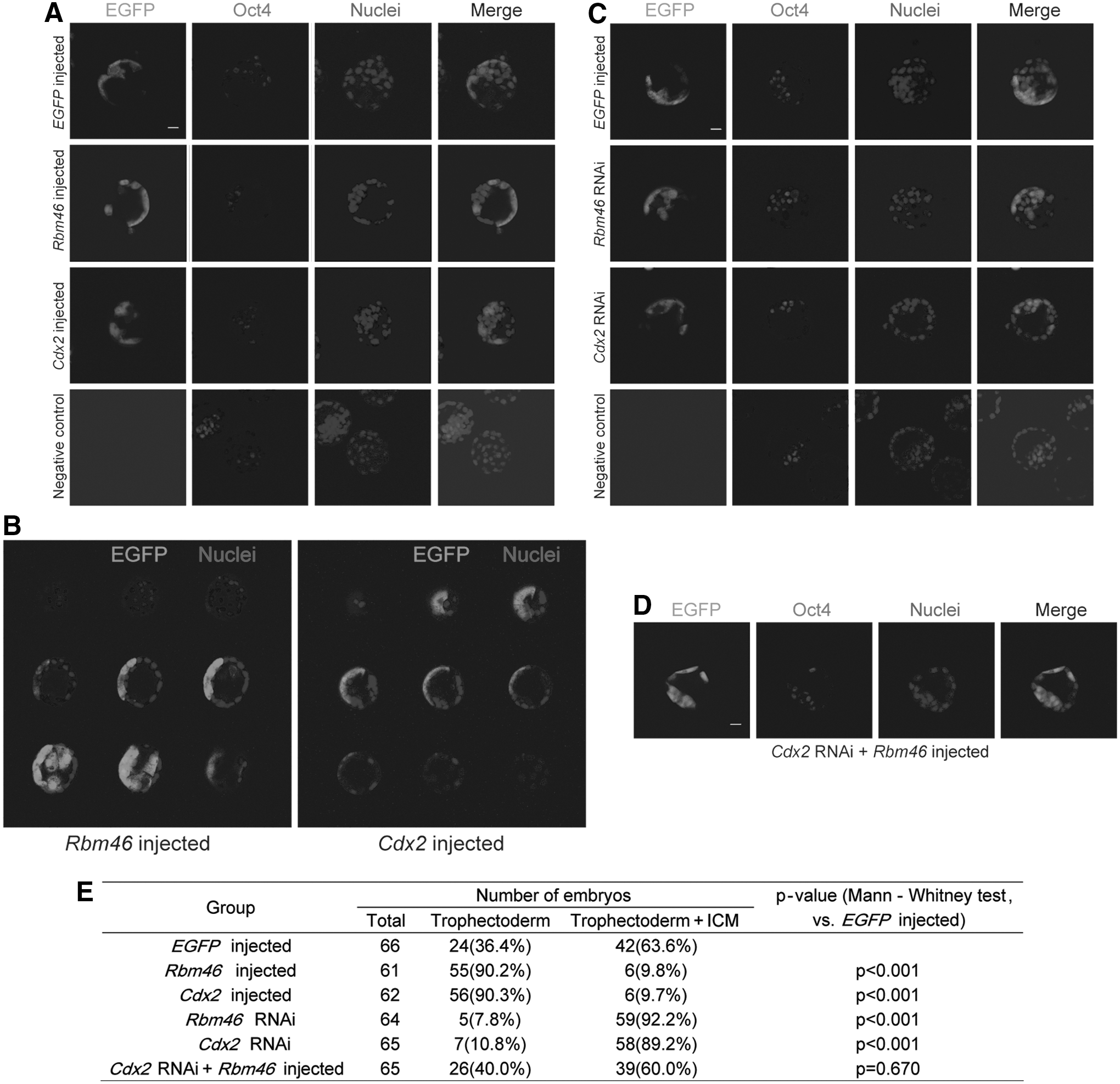
Rbm46 regulates trophectoderm specification in mouse early embryos.
To confirm the role of Rbm46 in regulation of trophectoderm specification, we injected Rbm46 dsRNA along with an EGFP tracer into single blastomere of a 2-cell embryo and inspected the EGFP green cell distribution in the blastocyst. Cdx2 dsRNA was also injected as a positive control. As predicted, inhibition of Rbm46 significantly decreased EGFP green cell allocation to the trophectoderm (Fig. 6C) and meanwhile increased EGFP-positive cells in ICM (Fig. 6C). 7.8% of embryos in the Rbm46 dsRNA-injected group have EGFP green cells contributing to trophectoderm, while 92.2% contributed to both trophectoderm and ICM (n=64 embryos, Mann–Whitney test, P<0.001) (Fig. 6E). A similar distribution was also made in the embryos injected with Cdx2 dsRNA (n=65 embryos, Mann–Whitney test, P<0.001) (Fig. 6E). There was no significant difference between Rbm46 RNAi and Cdx2 RNAi groups (Mann–Whitney test, P=0.565) (Fig. 6C, E). Most strikingly, embryos injected with Cdx2 dsRNA along with Rbm46 revealed an equal allocation of EGFP green cells in the blastocyst (n=65 embryos, Mann–Whitney test, P=0.670) (Fig. 6D, E), indicating that the defect in blastomere allocation caused by Cdx2 inhibition was reversed by Rbm46 mRNA injection (Fig. 6C). All these data indicates that Rbm46 plays a crucial role in regulation of trophectoderm specification by increasing Cdx2 production.
Discussion
Reciprocal expression of Cdx2 in trophectoderm and Oct4 in the ICM is the molecular basis for the first lineage specification in the blastocyst of early embryos [5]. Cdx2, which blocks Oct4 expression through interacting with other factors [14], is essential for the trophectoderm specification. However, post-transcriptional regulation of Cdx2 has not yet been documented. Cell-fate determination is a crucial process of development, and it remains unclear whether post-transcriptional regulation by RBP is involved or not. In this study, we report a novel finding that Rbm46 with unknown functions regulates the segregation of trophectoderm from ICM by stabilizing Cdx2 mRNA in early mouse embryos. Since RNA interference of Rbm46 reduces the allocation of blastomere cells to the trophectoderm (Fig. 6), this further supports the notion that Rbm46 plays a role in controlling the segregation of trophectoderm from the ICM in early mouse embryos.
At the same time, there are questions that can be further investigated. In TE lineage differentiation analysis, for example, it has been noted that mouse ESCs can transdifferentiate into trophectodermal cells only to a very limited extent [27]. Such transdifferentiation requires forced expression of a trophoblast-promoting factor, such as Cdx2 [16]. Our experiment provides evidence that Rbm46 is able to regulate ESCs differentiation into TE lineage in vitro. In this differentiation system, a shift in Fgf4+/heparin+culture condition will not trigger trophoblast differentiation of ESCs but has been shown to maintain TS cells in an undifferentiated diploid state. In our study, we did not find any significant change in other TE markers except for Cdx2 at our tested time window between PM46 cells and PE cells, and the trophoblast markers were only modestly activated, and thus, even after 6 days, the expression of other marker genes was increased but did not reach the levels of that seen in TS cells.
Overexpression of Rbm46 in ESCs regulated trophectoderm differentiation; trophectoderm markers were upregulated so that even knockdown of Rbm46 expression barely changed trophectoderm markers, but interestingly, some endoderm markers, such as Gata6 and Sox17, and mesoderm markers, such as Myf5 and MyoD, were increased when Rbm46 was knocked down (Fig. 3E). Some of them were downregulated when Rbm46 was overexpressed (Fig. 2F). This inverse correlation of endoderm markers to Rbm46 expression suggests that Rbm46-mediated trophectoderm differentiation may be partially due to inhibition of endoderm differentiation. Cell-fate determination is a crucial process of development. Specifically, ESCs were easier to differentiate into other mesodermal tissue, such as endoderm and mesoderm; this might be due to the changed state of cell populations mediated by knockdown of Rbm46.
Also, in Figure 1C, the expression of Cdx2 in EB is not derived from trophectoderm, which may come from neuroectoderm and endoderm cells. Rbm46 may post-transcriptionally regulate Cdx2 mRNA in all the three-layered embryos. Given the observation that overexpression of Rbm46 can regulate trophectoderm differentiation in ESCs and in mouse embryos, we focus on the regulation of trophectoderm by Rbm46 in this study. We are also interested in Rbm46 function in other systems, which will be our future work.
Mouse ESCs have been generally used for studying the regulation and the molecular mechanism of trophectoderm differentiation [28,29]. However, a caveat in using ESCs for such studies is that cells generated in this way may exhibit nonpredictable characteristics. For example, mouse ESCs can be converted into trophectoderm cells by ectopic expression of trophectoderm-determining transcription factors [7,23]. PM46 cells were obtained by introducing Rbm46 into mouse ESCs, and these cells showed a bias toward differentiating into trophectoderm cells (Fig. 4). Yet, at the same time, we found that PM46 cells also contain pluripotent factors, including Nanog, Oct4, and Sox2 protein (Fig. 2H). In real mouse blastocysts, Oct4 and Cdx2 may only be coexpressed in blastomeres for a short amount of time, which never exceeds 10 days. One explanation for this unusual observation is that the LIF-added culturing medium regulates the expression of the pluripotent factors to support ESC self-renewal. Meanwhile, the ESCs overexpressing Rbm46 in the culturing medium were in the early stage of differentiation with active pluripotent factors, and such predifferentiated cell populations were still predominantly comprised of undifferentiated ESCs. This early stage, mentioned above, is similar to a period at a much earlier point after the initiation of differentiation in the study by Hailesellasse et al. [30]. That is why ESCs overexpressing Rbm46 showed continued proliferation when expressing pluripotency-associated genes, such as Oct4, Nanog, and Sox2. Alternatively, Cdx2 may require more factors, which may not be present in PM46 cells, to block Oct4 expression.
Technically, the PM46 cells used in our studies were generated by introducing an episomal vector-delivered Rbm46 gene [27,31]. The advantage of this approach was that the possible side effect of random genome insertion was avoided while maintaining high transfection efficiency. The disadvantage is that Rbm46 expression levels vary from passage to passage. Using a doxycycline-inducible episomal vector in future studies may help to avoid possible variations in Rbm46 expression levels in the study of PM46 cell differentiation [32]. Meanwhile, the ESCs overexpressing Rbm46 in the culturing medium were in the early stage of differentiation with active pluripotent factors, and such predifferentiated cell populations were still predominantly composed of undifferentiated ESCs. Quantitative real-time PCR data indicated that the mRNA level of Rbm46 is much higher than the physiological level. According to the gray-scale scanning of the western blot bands (Fig. 2B), the protein level of Rbm46 is 10×higher in the PM46 cells than in the PE cells (108.398 DPI: 10.302 DPI). Thus, we combined knockdown experiments to investigate the physiological function of Rbm46.
While Rbm46 was expressed in all cells in the PM46 colonies (Supplementary Fig. S3C), so was Oct4 (Supplementary Fig. S3A). We observed that only the cells surrounding the ESC colony showed Cdx2 staining in the PM46 colony (Fig. 4A). A possible interpretation is that differentiation often occurs in those cells that are around the perimeter of undifferentiated colonies [33]. Consequently, Cdx2 would be expressed in those perimeter cells and Rbm46 could then stabilize Cdx2 mRNA and increase Cdx2 production. The fact that we are unable to detect Cdx2 staining in PE cells cultured (Fig. 4A) supports that Rbm46 is required for promoting Cdx2 production.
Based on the mouse ESCs to TE lineage differentiation analyses, as well as the blastomere injection approach, we provide evidence that Rbm46 plays a role in the segregation of trophectoderm from the ICM in mouse embryos. Whether the function of Rbm46 we observed in mouse development is applicable to human cases remains unknown. Our future studies will include the verification of the role of Rbm46 in regulating trophectoderm specification by trophectoderm-specific knocking out the Rbm46 gene in mice.
Regulation of trophectoderm specification is rather complicated and involves many trophectoderm factors, including Tead4, Cdx2, Gata3, and Eomes. [7]. Whether or not Rbm46 can also interact with other trophectoderm factors is still unclear. In addition, it would be interesting to see which of the transcription factors and signaling pathways regulates Rbm46 expression. Nonetheless, our study has found Rbm46 to be the first RBP that plays a crucial role in, but is not required for, trophectoderm specification through post-transcriptional regulation of Cdx2 mRNA in early mouse embryos.
Footnotes
Acknowledgments
We thank Ying Jin for providing the E14Tg2a mouse ESC lines and the pPyCAGIP vector; Wengong Wang, Linheng Li, and Sheng Ding for their comments; and David Stenger, Byron Lee, and Jenny Wu for editing the article. We also thank Xiaochen Kou, Yanhong Zhao, and Hui Li for performing embryo injections and preparing embryos samples. Additionally, we thank Li Wang for supporting teratoma formation experiment, Yinan Liu and Xiaoyan Zhang for performing FACS, and Lei Zhai for providing siRbm46 constructs. This work was supported by the International S&T Cooperation Program (grant no. 2011DFA31040) and the 863 Program (grant no. 2011AA020106) from the Ministry of Science and Technology of China, the New Drug Innovation of China (grant no. 2011ZX09102-010-02), a grant from Strategic Priority Research Program of the Chinese Academy of Sciences (grant no. XDA01040000), and a special grant from Shanghai Bureau of Science and Technology (grant no. 12DZ1910900).
Author Disclosure Statement
The authors declare no competing financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.