
Abstract
Apoptosis of bone marrow-derived mesenchymal stem cells (BMMSCs) is an essential pathogenic factor of osteoporosis. Ginsenoside-Rb2 (Rb2), a 20(S)-protopanaxadiol glycoside extracted from ginseng, is a potent treatment for bone loss, which raises interest regarding the bone metabolism area. In the present study, we found that dose–response Rb2 inhibited high dosage of dexamethasone (Dex)-induced apoptosis in primary murine BMMSCs. Interestingly, Rb2 promoted GPR120 induction, which is the unsaturated long-chain fatty acid receptor. We further confirmed that GPR120-specific ShRNA reversed the inhibition of Rb2 on Dex-induced apoptosis by activating caspase-3 and reducing cell viability. In addition, Rb2 notably increased phosphorylated ERK1/2 levels and Ras kinase activity dependently through the GPR120. The ERK1/2 activity-specific inhibitor U0126 remarkably blocked the Rb2-induced antiapoptotic effect in response to Dex-induced apoptosis. Together, dose–response Rb2 protected BMMSCs against Dex-induced apoptosis dependently by inducing GPR120 promoted Ras-ERK1/2 signaling pathway. Therefore, in the prevalence of the abuse of Dex in the clinic, our findings suggest for the first time that Rb2 is not only a key to understand the link between Chinese medicine and the pathology of osteoporosis but also an underlying target for the treatment of bone complications in the foreseeable future.
Introduction
O
Dexamethasone (Dex) is a potent synthetic member of the glucocorticoid class of steroid drugs. It acts as an anti-inflammatory and immunosuppressant [6,7]. Much attention has been paid to the effect of Dex on MSC proliferation and differentiation [8,9]. However, there are data showing that the effect of Dex on MSCs depends on the dosage when performed [10]. It was reported that low dosage of Dex promoted cell viability and osteogenic differentiation of BMMSCs [9,11], whereas high dosage could lead to apoptosis and cell death of BMMSCs [12], which would undoubtedly cause osteoporosis sequentially. However, the accurate dosage of causing apoptosis in BMMSCs remains to be answered.
The root of Panax ginseng C.A. Meyer is considered a highly bioactive phytomedicinal remedy and is widely used in the traditional medicine of China, Korea, and Japan [13]. Ginsenosides, the major pharmacologically active compounds found in ginseng, appear to be responsible for most of the activities of ginseng, including antioxidative, antidiabetic, anti-inflammatory, and anticarcinogenic effects [14 –16]. Rb2 is the most abundant saponin contained in P. ginseng [17 –20] and is reported to inhibit angiogenesis and improve wound healing by enhancing epidermal cell proliferation [21,22]. In addition, we have demonstrated that Rb2 displayed antiosteoporosis effects through reducing oxidative damage and bone-resorbing cytokines during osteogenesis [23]. In the above study, we found that Rb2 could not only decrease oxidative damage but also promote the cell viability of MC3T3-E1 cells and protect from cell death. However, there are very few studies of linking Rb2 to cell apoptosis and proliferation. Therefore, in this study, we want to investigate the role of Rb2 in the antiapoptotic effect of BMMSCs under certain conditions and elucidate the underlying mechanism behind this magic compound.
GPR120, a G protein-coupled receptor, also named as FFAR4, can be activated by unsaturated long-chain fatty acid (FA) and perform its effect mainly on the modulation of secretion of insulin and glycometabolism [24]. Until recently, GPR120 was found to have a positive role in the proliferation and inhibiting serum deprivation-induced apoptosis of murine enteroendocrine cell line STC-1 [25]. It showed that GPR120 significantly rescued serum deprivation-induced apoptosis and protected from cell death through Erk1/2. In addition, our preliminary data also suggested that GPR120 was an essential factor to get involved in the proliferation and antiapoptotic process of BMMSCs. However, whether Ginsenoside-Rb2 might provide the potential antiapoptotic effects and whether these effects are produced by involving GPR120 in BMMSCs have not been established.
In the present study, we demonstrated the antiapoptotic effect of Rb2 and examined the role of GPR120 in BMMSC protection from Dex-induced apoptosis. Specifically, we found that by activating the Ras-ERK1/2 pathway through upregulating GPR120, Rb2 inhibits Dex-induced apoptosis in BMMSCs to a certain extent. These findings indicate that Ginsenoside-Rb2, currently as the treatment of type 2 diabetes, could have some bonus on the prevention of the abuse of Dex, which would lead to skeletal diseases such as osteoporosis in the foreseeable future.
Materials and Methods
Materials
The α-Modified minimal essential medium (α-MEM) was purchased from Thermo Scientific. Polystyrene culture dishes were obtained from Costar, and fetal bovine serum (FBS) was purchased from Gibco Life Technologies. Rb2 extracted from ginseng [molecular weight, 1,079; purity, >98.0%; dissolved in dimethyl sulfoxide (DMSO)] was purchased from Shanghai Tauto Biotech Co., Ltd. Anti-Erk1/2 (ab17942), anti-Akt (ab126811), anti-p-Akt (ab81283), anti-p38 (ab31828), anti-p-p38 (ab4822), anti-JNK (ab4821), anti-p-JNK (ab124956), and anti-Brdu (ab1893) antibodies were purchased from Abcam; anti-GPR120 (sc48203), anti-Ras-GRF1 (sc963), anti-GPR40 (sc32905), and anti-PPARγ (sc7273) were purchased from Santa Cruz; p-Erk1/2 (No. 4370) was purchased from Cell Signaling. TUG-891 (4601-50) and U0126 (1144/25) were purchased from R&D Systems. GW1100 (306974-70-9) was purchased from Apexbio Technology. Penicillin–streptomycin was purchased from Solarbio. The total RNA extraction kit was purchased from OMEGA. The PrimeScript™ RT reagent kit and SYBR® Premix Ex Taq™ were purchased from TaKaRa. Other reagents were of the highest commercial grade and purchased from Sigma Chemical.
Cell culture
BMMSCs were isolated from C57BL/6 mice as previously described [26], and cells were characterized using MSC minimal criteria [27]. Briefly, the bone marrow was flushed out of long bones of mice and plated as a cell suspension (0.5×106 cells cm−2) in the α-MEM supplemented with 15% FBS, 2 mM glutamine, 100 U/mL penicillin, and 100 μg/mL streptomycin. Culture dishes were incubated at 37°C in a humidified atmosphere of 95% air and 5% CO2. The culture medium was replaced every other day. Homogenous BMMSCs were obtained through three generations. The experimental procedures in this study were approved by the Ethics Committee for Animal Experimentation of the Fourth Military Medical University (permission code 2010C00843).
Western blot analysis
Cells were lysed in 1% OG buffer (20 mM Tris–HCl, pH 8.0, 150 mM NaCl, 1% OG, 1 mM EDTA, 10 mg/mL leupeptin, 2 mg/mL aprotinin, and 1 mM phenylmethanesulfonyl fluoride). A BCA Protein Assay Kit (Pierce Biotechnology) was then used to determine the total protein density, and equal amounts of protein were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a polyvinylidene fluoride microporous membrane (Millipore). After being blocked with 5% nonfat milk, the membrane was incubated for 2 h at room temperature with the designated antibody. Images were analyzed by Scion Image software.
Reverse transcriptase–polymerase chain reaction
Total RNA was purified from cells using Trizol® (Invitrogen). Reverse transcription–polymerase chain reaction (RT-PCR) was performed, and results were analyzed as previously described [28]. All RT-PCR experiments were performed in triplicate, and the primers sequences were as follows: GPR120 (Fwd) CCAGTGTTGCTGGAGAAATC and (Rvs) TGATGCCTTGGTGATCTGTA; β-actin (Fwd) CTGGCACCACACCTTCTACA and (Rvs) GGTACGACCAGAGGCATACA.
MTT assay
Cell viability was determined by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. After treatment with Dex for 24 h, cultures were washed with phosphate-buffered saline (PBS). MTT (0.5 mg/mL) was then added to each well and the mixture was incubated for 4 h at 37°C. The culture medium was then replaced with an equal volume of DMSO to dissolve formazan crystals. After the mixture was shaken at room temperature for 30 min, absorbance of each well was determined at 550 nm.
TUNEL assay
Apoptosis in BMMSCs were detected using the DeadEnd Fluorometric TUNEL System (Promega), which end-labels the DNA from the fragmented apoptotic cells using a modified TUNEL staining according to the manufacturer's instructions. Briefly, samples were fixed with 4% methanol-free formaldehyde solution, rinsed with PBS twice, then end-labeled the fragmented DNA of the apoptotic cells using the fluorescein-12-dUTP, and covered slides in DAPI to stain nuclei. Samples were immediately analyzed under a fluorescence microscope using a standard fluorescein filter set (Olympus) to view the green fluorescence of apoptotic cells at 520 nm and blue DAPI-stained nuclei at 460 nm.
Detection of caspase-3 activity
The caspase-3 activity was measured spectrophotometrically through the detection of pNA cleavage by caspase-3-specific substrates. These experiments were completed using a caspase-3 assay kit (Beyotime). After the cell lysates were incubated with Ac-DEVD-pNA for 2 h at 37°C, the samples were read at 405 nm.
Transfection of lentiviral vectors with shRNA for GPR120
The pGMLV-GFP-vshRNA-GPR120 was constructed (pGMLV, a shRNAi Vector; Shanghai Genomeditech Co. Ltd). In the present study, we constructed three vshRNA-GPR120 lentiviral vectors (pGMLV-GFP-shRNA-GPR120) to silence the expression of GPR120 in murine BMMSCs (BMMSCs-vshRNA-GPR120). The three shRNA-targeting sequences for GPR120 were as follows: (1) 5′-GCACCCACTTCCCTTTCTTCT-3′; (2) 5′-GCTCTTCTACGTGATGACAAT-3′; and (3) 5′-GGACCAGGAAATTCCGATTTG-3′. The sequence for control shRNA was 5′-TTCTCCGAACGTGTCACGT-3′. Stably transfectant clones were characterized by RT-PCR, western blot, and immunoblotting.
Annexin V/propidium iodide double staining
The number of apoptotic cells was determined by Annexin V/propidium iodide (PI) double staining. Cells were exposed to 1 μM Dex in the presence or absence of Rb2 for 24 h and then incubated with FITC-conjugated Annexin V in a binding buffer (0.01 M HEPES, 0.14 M NaCl, and 2.5 mM CaCl2, pH 7.4) for 30 min at 37°C in the dark. After incubation, the cells were washed, resuspended in 200 mL PBS with 1% fetal calf serum, and additionally incubated with 10 mL of 1 mg/mL PI solution. The Annexin V-positive cells were detected using a FACSCalibur flow cytometer (BD Biosciences), and the results were analyzed using CellQuest software (BD Biosciences). Annexin V-FITC conjugates were detected with the FL1 channel of the FACSCalibur machine. PI was read on the FL2 channel.
Statistical analysis
Data are expressed as mean±SD of multiple repeats of the same experiment (n=5). The data for these measurements were analyzed by one-way analysis of variance (ANOVA) with subsequent post hoc multiple comparisons by Dunnett's test. Statistically significant values were defined as P<0.05.
Results
Ginsenoside-Rb2 inhibits Dex-induced apoptosis
Previous studies have shown that exposure of the cultures to Dex causes BMMSCs to apoptosis [12]. However, the accurate dosage for Dex to cause apoptosis of BMMSCs is not clearly defined. Therefore, we first cultured BMMSCs with different concentrations of Dex, and we found that 10−6 M is the concentration of causing apoptosis of BMMSCs (Supplementary Fig. S1 Supplementary Data are available online at
To find out the role and function of Ginsenoside-Rb2 on the process of Dex-induced apoptosis, cells were cultured with 10−6 M Dex in the presence or absence with increasing concentrations of Rb2 for 24 h. Figure 1 is the chemical structure of Ginsenoside-Rb2 (from National Center of Biotechnology Information, Pubchem CID: 5458674). As is shown in Fig. 2, pretreatment with increasing concentrations of Rb2 remarkably reversed the survival rate inhibited by Dex and decreased the percentage of apoptotic cell and interestingly, the effects also plateaued at 10−6 M (P<0.05, Fig. 2A–C). These results suggested that Rb2 could inhibit Dex-induced cell apoptosis and cell death. What is more, the caspase-3 activity further indicated that Rb2 could inhibit Dex-induced apoptosis and followed a dose–response manner (Fig. 2D). To determine whether the cell death was apoptotic like, TUNEL assay was performed. Because the responses to 10−5 and 10−6 M Rb2 were virtually identical, 10−6 M of Rb2 was thereafter employed. The statistic data showed that the percentage of apoptotic cells increased after Dex treatment. However, pretreated with 10−6 M of Rb2 reduced the percentage of apoptotic cells induced by Dex (P<0.05, Fig. 2E, F).
Chemical structure of Ginsenoside-Rb2 (from the National Center of Biotechnology Information, Pubchem CID: 5458674).
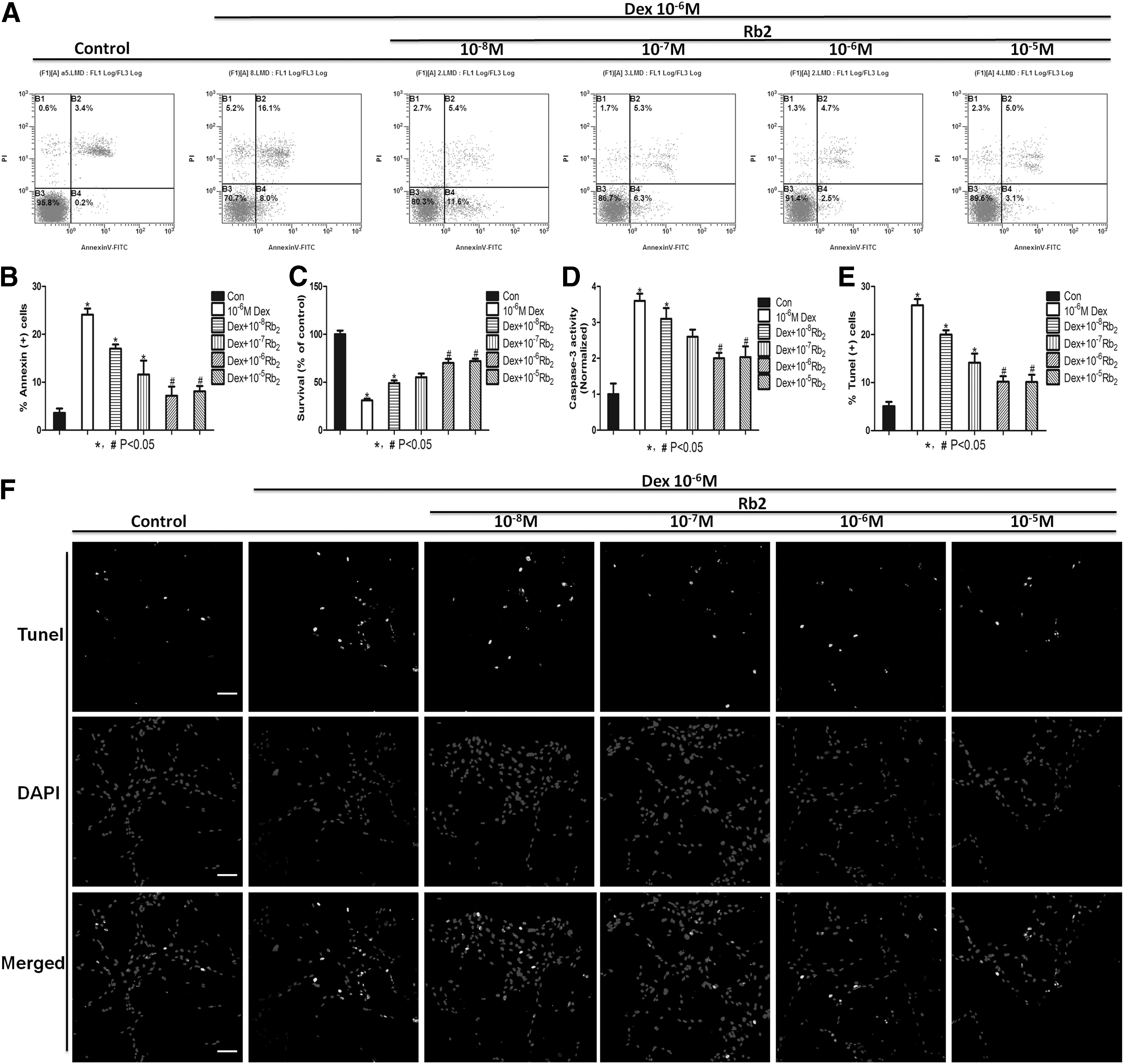
Ginsenoside-Rb2 inhibits Dex-induced apoptosis. BMMSCs were treated with Dex (10−6 M) in the absence or presence of Rb2 for 24 h.
The protective effect of Rb2 on Dex-induced apoptosis is dependent on GPR120 (FFAR4)
To further expand the underlying mechanism of the antiapoptotic effect of Rb2 on BMMSCs, we performed the mechanistic experiments. At the initial stage of our experiment, we coincidently found that Rb2 could gradually upregulate the mRNA and protein level by increasing concentrations (Fig. 3A, B). To examine whether the protective effect of Rb2 on Dex-induced apoptosis is dependent on GPR120, we used ShRNA to knock down GPR120 in BMMSCs. GPR120 ShRNA led to a marked reduction of GPR120 protein and mRNA levels (Fig. 3C, D). When the cells were transfected with GPR120 ShRNA, Rb2 treatments did not decrease the percentage of apoptotic cell and increase the survival rate compared with the corresponding control cells (P<0.05, Fig. 4A, B). To further validate this finding, we examined the activities of caspase-3 in the condition that GPR120 induction was inhibited with GPR120 ShRNA. The results showed that Rb2 treatments decreased the activities of caspase-3 following control ShRNA treatments. However, Rb2 treatments were not able to inhibit the activities of caspase-3 following GPR120 ShRNA treatments (P<0.05, Fig. 4C). TUNEL assay also suggested that Rb2 treatments could decrease the percentage of apoptotic cells following control ShRNA treatments, whereas could not reduce the percentage of apoptotic cells following GPR120 ShRNA treatments (P<0.05, Fig. 4D, E). These results further suggested that Rb2 protects BMMSCs from Dex-induced apoptosis by promoting GPR120 induction.
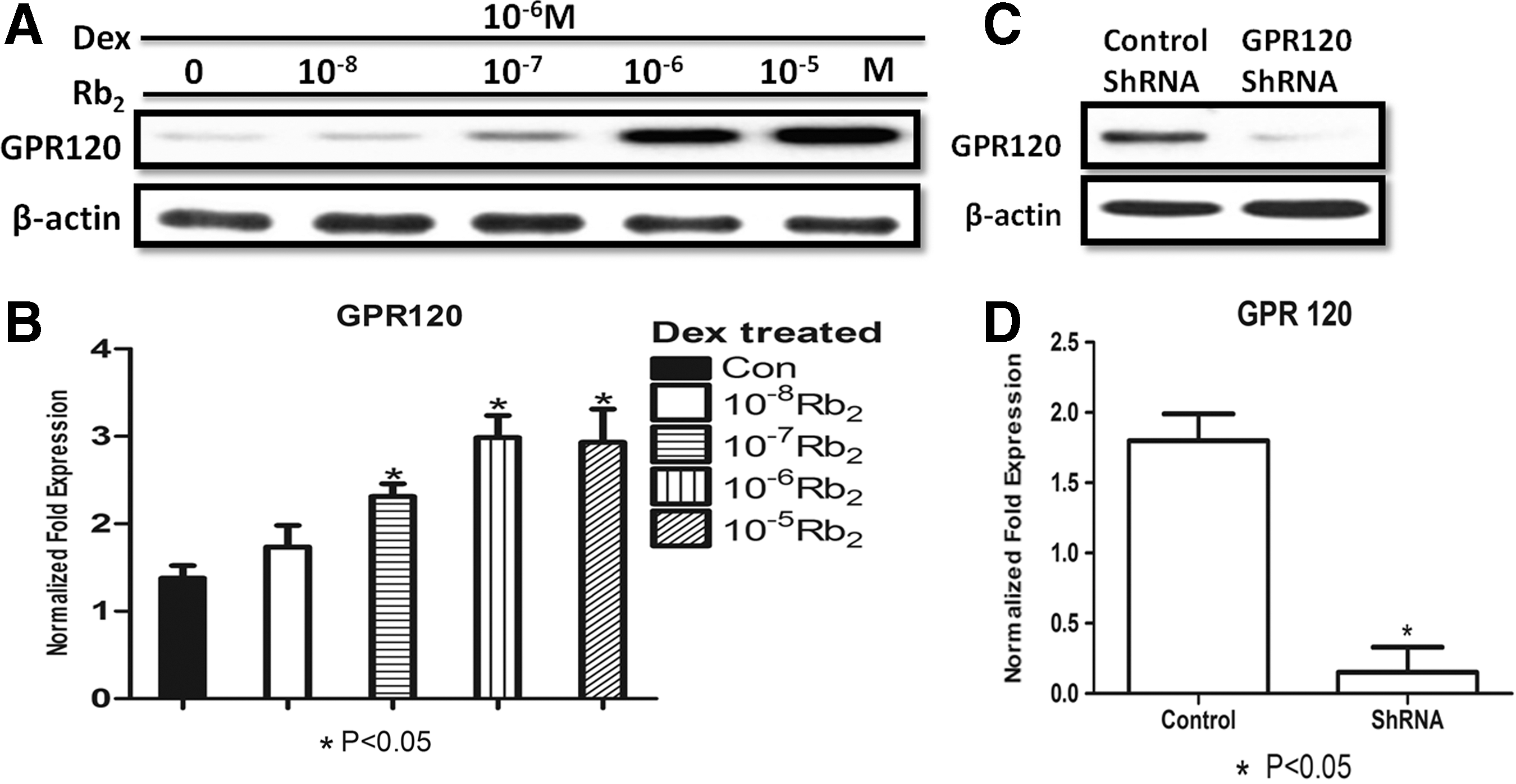
Dose–response Rb2 induces GPR120 expression.
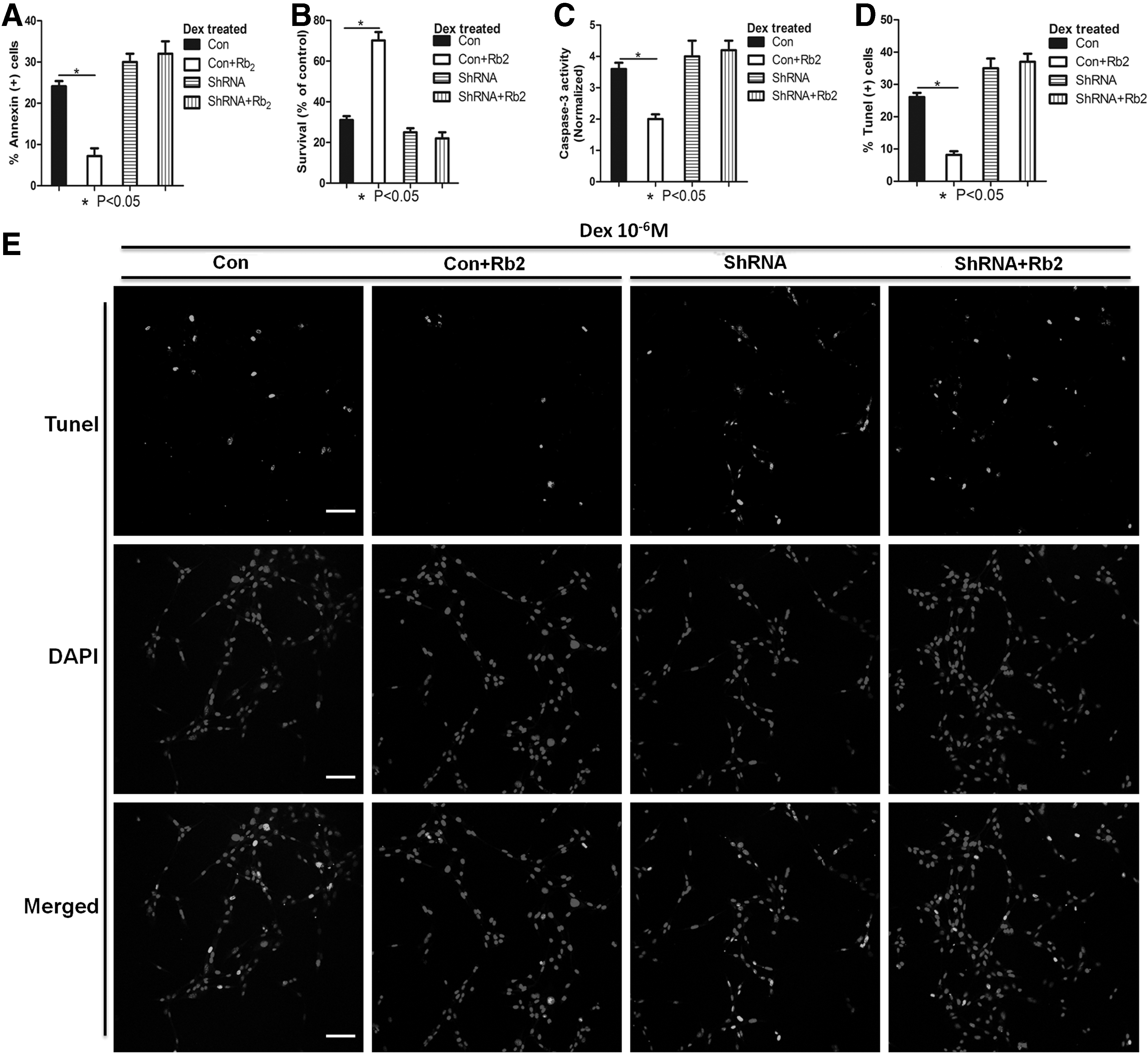
The protective effect of Rb2 on Dex-induced apoptosis is dependent on GPR120 (FFAR4). Control or GPR120 ShRNA-treated cells were treated with Dex (10−6 M) in the absence or presence of Rb2 (10−6 M) for 24 h.
Rb2 activates Ras-ERK1/2 signaling pathway through GPR120 induction in response to Dex-induced apoptosis in BMMSCs
Recent study has indicated that GPR120 could exhibit the antiapoptotic effect [25], however, the downstream of GPR120 target on apoptosis has not been established. Therefore, we tested the phosphorylation and expression levels of Erk1/2, Akt, P38, and JNK, which were previously proven to take part in the antiapoptotic process [29 –33]. Cells from the control or GPR120 ShRNA group were pretreated with 10−6 M of Dex in the presence or absence with 10−6 M of Rb2. Although the activation of GPR120 by Rb2 did not alter the phosphorylation and expression levels of Akt, P38, and JNK, Rb2 markedly increased the phosphorylated ERK1/2 levels in BMMSCs after Dex treatment, and compared with control ShRNA-treated BMMSCs, the phosphorylated ERK1/2 levels of cells treated with GPR120 ShRNA were much lower and could not be upregulated by Rb2 (Fig. 5A). In addition, we observed that addition of Rb2 markedly promoted the Ras kinase activity after Dex treatment (Fig. 5B). These data indicated that Rb2 may activate the Ras-ERK1/2 signaling pathway of BMMSCs dependently through GPR120 in Dex-induced apoptosis.
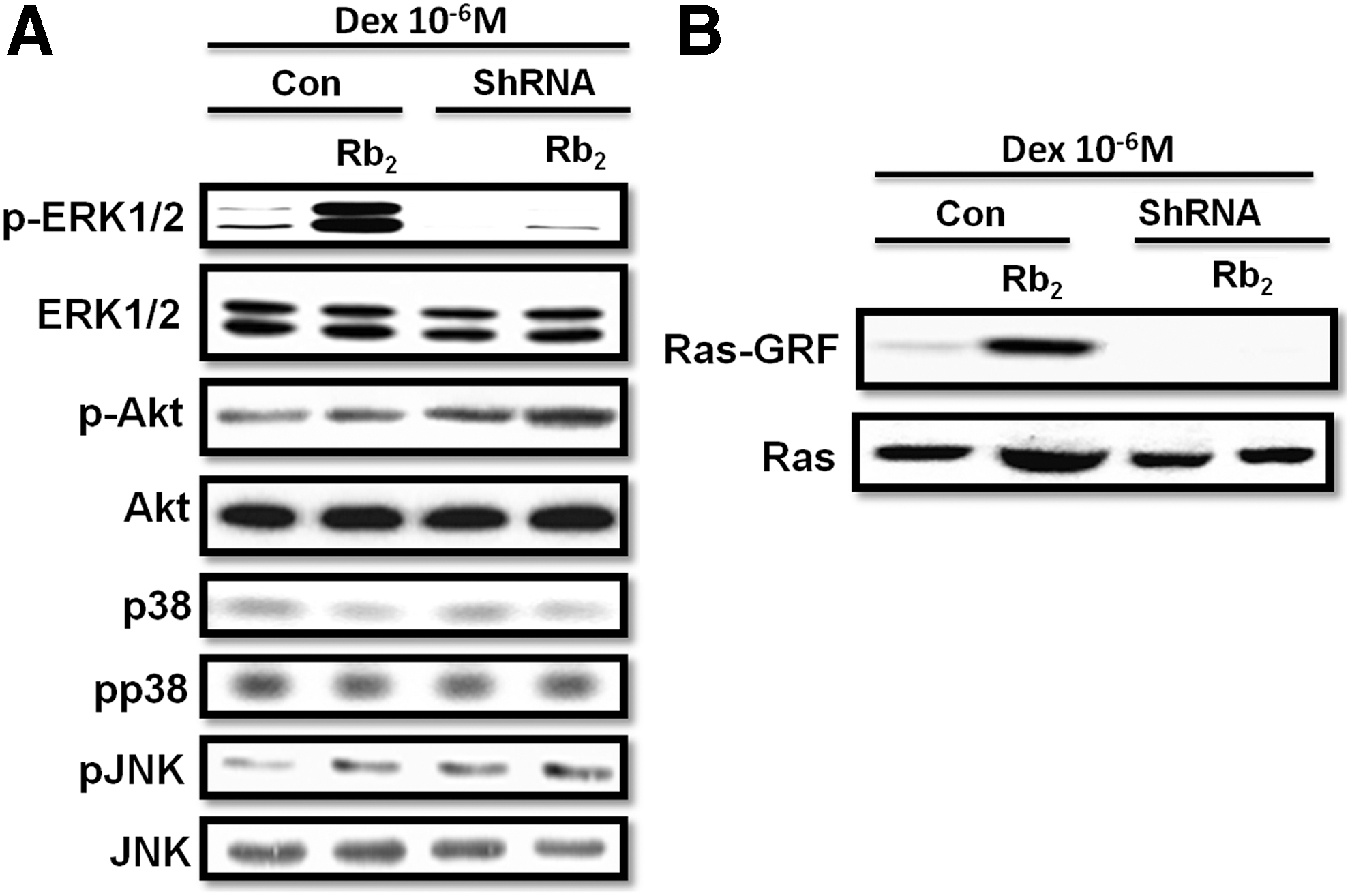
Rb2 activates the Ras-ERK1/2 signaling pathway through GPR120 induction in response to Dex-induced apoptosis in BMMSCs. Control or GPR120 ShRNA-treated cells were treated with Dex (10−6 M) in the absence or presence of Rb2 (10−6 M) for 24 h.
The antiapoptotic effect of Rb2 by inducing GPR120 is blocked by Erk1/2 inhibitor
To examine whether the antiapoptotic effect of GPR120 upregulated by Rb2 was dependent on the Ras-ERK1/2 signaling pathway, the ERK1/2 activity-specific inhibitor U0126 was used. We observed that the addition of ERK1/2 inhibitor U0126 significantly blocked the inhibitory effect of Rb2 on the percentage of apoptotic cell caused by Dex (Fig. 6A). Moreover, we confirmed that the addition of ERK1/2 inhibitor blocked the promotive effect of Rb2 on survival rate in BMMSCs compared with corresponding control cells (P<0.05, Fig. 6B). What is more, we further demonstrated that the supplementation of ERK1/2 inhibitor promoted the inhibitory effect of Rb2 on caspase-3 activity (P<0.05, Fig. 6C). These results all suggested that the Ras-ERK1/2 signaling pathway is dependently involved in the antiapoptotic effect of Rb2 and worked downstream of GPR120.
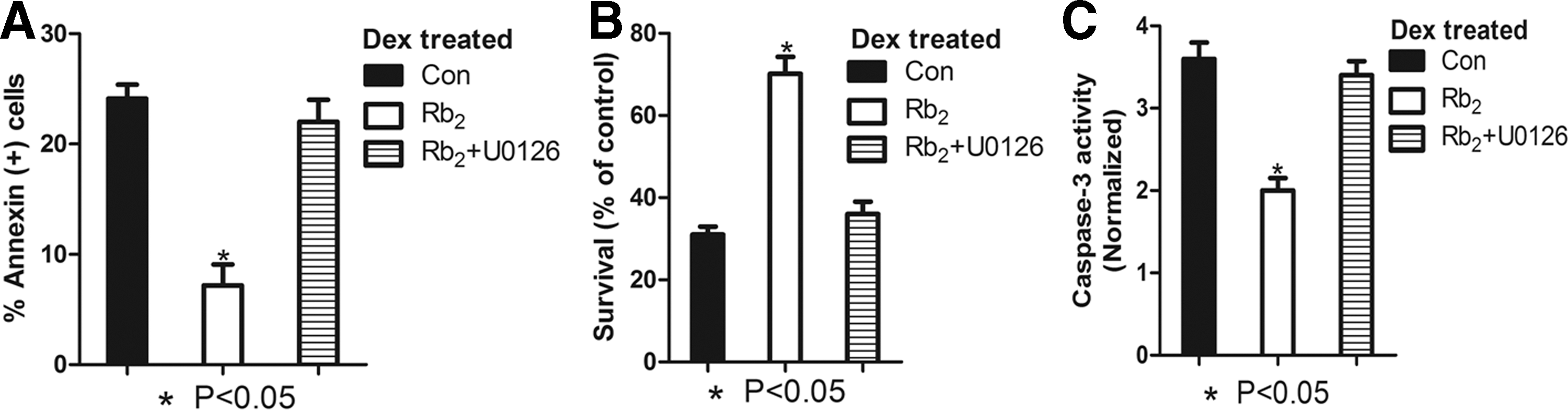
The antiapoptotic effect of Rb2 by inducing GPR120 is blocked by Erk1/2 inhibitor. The cells were pretreated with ERK1/2 inhibitor U0126 (10−7 M) and then incubated with Dex (10−6 M) in the absence or presence of Rb2 (10−6 M) for 24 h.
Discussion
In this study, we have demonstrated that Ginsenoside-Rb2, a 20(S)-protopanaxadiol glycoside extracted from ginseng, inhibits Dex-induced apoptosis through GPR120-induced Erk1/2 activation in BMMSCs. We found that dose–response Rb2 markedly reversed the apoptotic effect of Dex and improved the cell viability of BMMSCs dependently through the GPR120/Ras/Erk1/2 cascade.
Ginsenosides have been used as a traditional medicine with various therapeutic effects. They are generally separated into two groups: protopanaxadiol (PPDs; Rb1, Rb2, Rb3, Rc, and Rd) and protopanaxatriol (PPTs; Rg1, Re, Rf, and Rg2) [34]. Previous studies of the structure–activity relationship of ginsenosides have shown that the activities of the PPD compounds are greater than those of the PPT compounds. In this regard, Rb2 was used in this study and shown that it inhibited the apoptotic effect caused by high dosage of Dex. However, there have been no reports on whether Rb2 exhibited the antiapoptotic effect. Indeed, Sohn and colleagues reported that the protopanaxadiol ginsenosides, Rb (1) and Rb (2), and their intestinal bacterial metabolite, 20-O-beta-D-glucopyranosyl-20(S)-protopanaxadiol (M1), and the protopanaxatriol ginsenoside, Rg (1), and its intestinal bacterial metabolite, 20(S)-protopanaxatriol, triggered apoptosis in activated rat hepatic stellate cells through caspase-3 activation, which could have an anticancer effect [35]. Moreover, Yang and colleagues indicated that the profiles of Panax notoginseng saponin extract, the major bioactive ingredients in P. notoginseng (Burk.) F.H. Chen, exhibited anticancer effects such as antioxidant, antiproliferative, and proapoptotic activities in the human colon cancer cell line LoVo [36]. However, no report has ever demonstrated the effect of Rb2 alone on the antiapoptotic effect. Choi once reported that Rb2 enhanced epidermal cell proliferation by upregulating the expressions of proliferation-related factors [22]. What is more, our previous study has indicated that Rb2 possessed the antiosteoporosis effect and improved the cell viability treated by H2O2 [23]. Regarding these findings, we performed in vitro experiments to investigate the role and function of Rb2 alone on the apoptotic effect.
Dex is a potent synthetic member of the glucocorticoid class of steroid drugs and is widely used as an anti-inflammatory and immunosuppressant [6,7]. It was reported that a low dosage of Dex induced osteogenesis and displayed the antiapoptotic effect of BMMSCs, while a high dosage of Dex pushed BMMSCs to undergo adipogenesis and induced cells to apoptosis. Due to the abuse of this drug in clinic, discovering a new solution to relieve the side effect of Dex and meanwhile to inhibit bone loss will undoubtedly bring benefits to all patients who suffer skeletal diseases such as osteoporosis. However, until recently, the dosage of Dex to cause apoptosis has reached no final conclusion. Therefore, we used increasing concentrations of Dex to figure out the accurate dosage for it to cause apoptosis of BMMSCs. We found that high dosage of Dex markedly induced apoptosis of BMMSCs and the effect plateaued at 10−6 M (Supplementary Fig. S1). According to what has been discussed above, we used Rb2 to the culture system to further expand its role in Dex-induced apoptosis of BMMSCs. Several apoptotic markers indicated that the antiapoptotic effect of increasing concentrations of Rb2 significantly inhibited Dex-induced apoptosis and followed a dose-dependent manner. Indeed, several studies have mentioned the proliferative, anticancer, and antiosteoporosis effect of Rb2 [35,36]. However, no report has ever demonstrated the effect of Rb2 alone on the antiapoptotic effect of BMMSCs. Therefore, this is the first study to show the role of dose–response Rb2 on the antiapoptotic effect of BMMSCs.
Caspases are the primary modulators of apoptosis. Caspase-12 triggers the activation of caspase-3, caspase-7, and caspase-9 in a cytochrome c-independent manner [37]. Caspase-3 is of particular interest as it appears to be a common downstream apoptosis factor. It exists as a proenzyme in most cells, including BMMSCs, and is processed and activated by caspase-8 and caspase-9 to the heterodimeric form [38]. In this study, we found that Rb2 was shown to attenuate Dex-induced activation of caspase-3, which further confirmed that Rb2 could protect BMMSCs from Dex-induced apoptosis that was mediated by caspases.
GPR120, as a G protein-coupled receptor, can be activated by unsaturated long-chain FA and perform its effect mainly on the modulation of secretion of insulin and glycometabolism [24]. Recently, GPR120 was reported to take a positive part in the proliferation and inhibit serum deprivation-induced apoptosis of murine enteroendocrine cell line STC-1 [25]. Moreover, some studies indicated that GPR120 has a great anti-inflammatory effect of macrophages [39 –41] and capable of activating the Ca2+ signaling response to FAs [42 –44]. Until now, Koren et al. suggested that omega-3 FA mediated GPR120 at early age to accelerate bone growth partially by increasing proliferation of osteocytes, indicating that GPR120 may take an important part in the process of proliferation of bone-related cells. In our preliminary data, we found that high dosage of TUG891, the highly selective agonist of GPR120, significantly promote cell viability and increase proliferation of BMMSCs. Moreover, to further figure out the accurate role of GPR120 on Dex-induced apoptosis, we used TUG-891 in the presence of 10−6 M Dex to find out whether the activation of GPR120 could have the antiapoptotic effect in BMMSCs. The result showed that TUG-891 could markedly inhibit the apoptotic effect of Dex and improve the viability of BMMSCs (Supplementary Fig. S2). By showing this, we further tested whether GPR120 could be activated by Rb2 to exert its antiapoptotic effect. Western blot showed that increasing concentrations of Rb2 could upregulate the expression level of GPR120. To further expand the role of GPR120 on the antiapoptotic effect of Rb2, we established GPR120 ShRNA to knock down this target. Annexin V/PI double staining, MTT assay, caspase-3 activity, and TUNEL assay all suggested that the dose–response Rb2 inhibits Dex-induced apoptosis and improves the survival rate dependently through GPR120, which further proved the antiapoptotic effect of this target on BMMSCs.
Recently, several groups reported that five orphan receptors, GPR40, GPR41, GPR43, GPR84, and GPR120, can be activated by free FAs (FFAs). Short-chain FAs are specific agonists for GPR41 and GPR43 [45], and medium-chain FAs for GPR84 [46]. Long-chain FAs can activate GPR40 [47] and GPR120 [48]. However, the role of these FFAs on apoptosis has not been clearly defined. GPR40 and GPR120 share some same agonists, such as GW9508. Therefore, to avoid GPR40 play a role in the antiapoptotic effect of Dex-induced apoptosis of BMMSCs, we tested the protein and mRNA level of GPR40 under the treatment of Rb2. The result showed that increasing concentrations of Rb2 could not activate GPR40 (Supplementary Fig. S3). To further figure out the relevant relationship between Rb2 and GPR40, the antagonist of GPR40-GW1100 was employed in the presence of Rb2 and Dex. However, downregulation of GPR40 by GW1100 has no effect on antiapoptosis by increasing concentrations of Rb2, which further indicated that it was GPR120, not GPR40, to be the mediator to modulate the antiapoptosis of Rb2 (Supplementary Fig. S4).
Tsujimoto and colleagues [25] once suggested that GPR120 inhibited serum-deprived-induced apoptosis of STC-1 cells through the activation of Erk1/2 and Akt. Therefore, to further explore the downstream target of GPR120 on its apoptotic effect in our experimental system, several canonical signaling targets were tested under the condition of Dex pretreatment, for instance, total and phosphorous Erk, Akt, P38, and JNK. The result showed that total and phosphorous Erk1/2 were markedly activated by GPR120 in the presence of Rb2 and Dex and with the knockdown of GPR120, Rb2 could not induce the activation of Erk1/2 in the presence of Dex-induced apoptosis, which further explained the essential role of GPR120 in the upstream of Erk1/2. We further tested the expression level of Ras and Ras-GRF. Consistent with the result of Erk1/2, GPR120 could also modulate the activation of Ras by the same tendency, which further indicated that Ras/Erk1/2 was the downstream target of GPR120 on its apoptotic effect. To further validate whether the antiapoptotic effect of Rb2 by inducing GPR120 was through the activation of Erk1/2, we used the Erk1/2-specific inhibitor U0126 to demonstrate this issue. Several apoptotic markers showed that U0126 significantly blocked the promotive effect of GPR120, which was activated by Rb2 and decreased the survival rate of BMMSCs. These results further confirmed that dose–response Rb2 induced GPR120 to activate Ras-ERK1/2 signal cascades to inhibit Dex-induced apoptosis.
Until now, it is still not clearly defined how Rb2 activates GPR120 to exhibit the antiapoptotic effect because previous views considered that the activation of GPR120 needs the ligand such as long-chain unsaturated FA or small molecular synthetic agonist. Nevertheless, our results showed that increasing concentrations of Rb2 could upregulate both the protein and mRNA expression level of GPR120, and with the knock down of GPR120, Rb2 could not exhibit its antiapoptotic effect, which strongly indicated that Rb2 has the ability to upregulate GPR120 directly or indirectly. Honestly, because of no direct evidence regarding the promoter of murine GPR120 until now, we will find out the promoter that Rb2 targeting on GPR120 and figure out more specific mechanism of Rb2 and GPR120 on the antiapoptotic effect on BMMSCs or other bone-related cells such as osteocyte, osteoblast and osteocyte in our following work.
In summary, our present results support that dose–response Rb2 protects BMMSCs from Dex-induced apoptosis through promoting the induction of GPR120 to activate the Ras-ERK1/2 cascade. Dex-induced apoptosis acts as a causative agent of osteoporosis. Until now, Ginsenoside-Rb2 has been suggested to be a potential opportunity and target to treat type 2 diabetes and obesity-induced inflammation in the future. However, in the prevalence of age- and metabolic-related disorders, our findings highlight the essential role of Rb2 in mediating apoptosis and proliferation of BMMSCs and inspire us that Rb2 and its downstream may be the new target and measure to combat skeletal diseases in the foreseeable future.
Footnotes
Acknowledgments
This work was supported by the Ministry of Science and Technology of China (2011CB964703), the National High Technology Research and Development Program 863 (2012AA020502), the National Natural Science Foundation of China (30901504), and the Program for Changjiang Scholars and Innovative Research Team in University (IRT13051). No benefits in any form have been or will be received from a commercial party directly or indirectly by the authors of this article.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.