
Abstract
Tissue-engineered constructs are rendered useless without a functional vasculature owing to a lack of nutrients and oxygen. Cell-based approaches to reconstruct blood vessels can yield structures that mimic native vasculature and aid transplantation. Vascular derivatives of human induced pluripotent stem cells (hiPSCs) offer opportunities to generate patient-specific therapies and potentially provide unlimited amounts of vascular cells. To be used in engineered vascular constructs and confer therapeutic benefit, vascular derivatives must exhibit additional key properties, including extracellular matrix (ECM) production to confer structural integrity and growth factor production to facilitate integration. In this study, we examine the hypothesis that vascular cells derived from hiPSCs exhibit these critical properties to facilitate their use in engineered tissues. hiPSCs were codifferentiated toward early vascular cells (EVCs), a bicellular population of endothelial cells (ECs) and pericytes, under varying low-oxygen differentiation conditions; subsequently, ECs were isolated and passaged. We found that EVCs differentiated under low-oxygen conditions produced copious amounts of collagen IV and fibronectin as well as vascular endothelial growth factor and angiopoietin 2. EVCs differentiated under atmospheric conditions did not demonstrate such abundant ECM expression, but exhibited greater expression of angiopoietin 1. Isolated ECs could proliferate up to three passages while maintaining the EC marker vascular endothelial cadherin. Isolated ECs demonstrated an increased propensity to produce ECM compared with their EVC correlates and took on an arterial-like fate. These findings illustrate that hiPSC vascular derivates hold great potential for therapeutic use and should continue to be a preferred cell source for vascular construction.
Introduction
T
In our previous study, we examined the codifferentiation of ECs and pericytes in a bicellular population, termed early vascular cells (EVCs), from human pluripotent stem cells (hPSCs) [7]. EVCs were able to assemble into microvascular structures within synthetic hydrogels that could then integrate with host tissue upon in vivo implantation. To be used in engineered constructs and confer therapeutic benefit, transplanted vascular cells must exhibit several important properties. Physiologically, the extracellular matrix (ECM) maintains structure and support for tissues [8 –10]. Likewise in engineered constructs, ECM production from encapsulated cells is critical for structural integrity. Of particular importance to the vascular ECM are collagen I, collagen IV, fibronectin, and laminin, commonly found in the basement membrane [11,12].
Second, growth factors released from derived cells help to facilitate integration of the scaffold [13]. Vascular endothelial growth factor (VEGF) is perhaps the most potent molecular regulator in vascular regeneration and has been extensively studied in vascular repair [14]. Transplanted scaffolds encapsulating VEGF have been demonstrated to augment blood vessel infiltration from the host (reviewed in Zisch et al. [15]). Another angiogenic growth factor, angiopoietin 1 (Ang1), has been implicated in vessel maturation and stabilization and is commonly produced by perivascular cells [16]. Conversely, angiopoietin 2 (Ang2) is demonstrated to promote endothelial sprouting—and thereby less pericyte coverage and vessel maturation—and is commonly produced by ECs. Production of these growth factors by transplanted cells can aid in the integration of host tissue with the transplanted construct.
Third, a large number of cells must be available for transplantation; thus, cells must be able to be passaged and expanded in the lab to generate a sufficient number of cells. For vascular tissue engineering applications outside of microvascular reconstruction, such as rebuilding vessel grafts or larger vessels, expanded EVCs will be a less-relevant population because pericytes will likely grow to occupy the majority of the culture [17,18]; thus, understanding the propagation potential of derived ECs will be more broadly applicable to tissue engineering applications at large. Inherently, mature ECs possess a low-proliferation capacity [19]. Vascular cells derived from hPSCs, known for their limitless self-renewal capacity and ability to differentiate into all cell types, can be harnessed to overcome this limitation. Additionally, it is important to assess whether ECs derived from hPSCs may also be expanded to generate a large cell population as previous studies have documented limited expansion capabilities of hPSC derivatives owing to cellular senescence after a certain number of population doublings [20].
A fourth consideration is that transplanted cells should be patient specific, in order to prevent an immune reaction [21,22]. Human induced PSCs (hiPSCs) exhibit the advantages of hPSCs and are reprogrammed directly from a patient's own cells. Thus, in the current study, we focus on vascular differentiation from the hiPSC line BC1, which was derived nonvirally and fully genetically sequenced, imparting translational importance [23,24].
In our previous study, we examined the derivation of EVCs from hPSCs [7]. Subsequently, we demonstrated the ability to obtain derived populations with different EC-to-pericyte ratios by allowing differentiation to proceed in various hypoxic, or low-oxygen (∼5% O2), environments [25]. Three classes of EVCs were derived and compared: control, primed, and continuous. Control EVCs were differentiated under 21% O2 (i.e., atmospheric) conditions and yielded ∼20% vascular endothelial cadherin (VEcad)–positive cells, which we used to denote the fraction of ECs present. Primed EVCs were differentiated under 5% O2 conditions for the first half of differentiation and 21% O2 conditions for the second half of differentiation and were composed of ∼39% ECs. Continuous EVCs were differentiated under 5% O2 conditions throughout the entire 12-day differentiation period and were made up of ∼44% ECs. EVCs derived under hypoxic conditions also exhibited EC phenotypes and functionalities as well as a preference toward arterial-like fates.
In the current study, we sought to interrogate our vascular derivatives for their potential to produce ECM proteins collagen I, collagen IV, fibronectin, and laminin as well as angiogenic growth factors VEGF, Ang1, and Ang2, and to be passaged, expanded, and characterized to yield clinically relevant quantities of cells. We anticipate that these findings will shed light on their therapeutic use and elucidate whether differences exist between EVCs derived from varying differentiation conditions.
Materials and Methods
hiPSC culture
hiPSC line, BC1 [23,24], was cultured as previously described [7]. Briefly, BC1 was grown on inactivated mouse embryonic feeder layers (Globalstem) in growth medium made up on 80% embryonic stem cell-Dulbecco's modified Eagle's medium (ES-DMEM/F12) (Globalstem) supplemented with 20% knockout serum replacement (Invitrogen) and 10 ng/mL basic fibroblast growth factor (Invitrogen). Cells were passaged every 4–6 days with type IV collagenase (Invitrogen) and media were changed daily.
hiPSC differentiation
hiPSC line BC1 was digested into a single-cell suspension with ethylenediaminetetraacetic acid (EDTA; Promega) and seeded onto collagen IV-coated dishes as described previously [7,25]. Cells were cultured in a differentiation medium composed of alpha minimum essential medium (alpha-MEM) (Invitrogen), 10% fetal bovine serum (FBS; Hyclone), and 0.1 mM β-mercaptoethanol and cultured for 6 days. On day 6, differentiated cells were collected and reseeded on collagen-type-IV-coated plates in EC growth media (ECGM; Promocell) supplemented with 2% FBS, 50 ng/mL VEGF, 10 μM SB43542 (Tocris), and 0.1% penicillin-streptomycin for another 6 days, at which point they were labeled EVCs. Media were changed every 3 days. To derive control EVCs, the aforementioned differentiation protocol took place under normoxic (21% O2) conditions. To derive primed EVCs, the differentiation occurred under 5% O2 conditions over the first 6 days and normoxic conditions over the second 6 days. To derive continuous EVCs, the differentiation took place under 5% O2 conditions for the entire 12 days, as previously described [25]. To observe ECM production capacity, the differentiation media were replaced with endothelial growth medium (EGM2) supplemented with 10% FBS for 48 h [26].
Immunofluorescence
Cells were prepared for immunofluorescence as previously described [26]. Briefly, cells were fixed with 3.7% formaldehyde; blocked in 1% bovine serum albumin; treated with 0.1% Triton-X (Sigma); incubated with the mouse anti-human collagen I (Abcam), mouse anti-human collagen IV (Santa Cruz Biotechnology), rabbit anti-human fibronectin (Santa Cruz Biotechnology), rabbit anti-human laminin (Abcam), or mouse anti-human Ki67 (Invitrogen), followed by the either anti-mouse FITC (Sigma) or anti-rabbit Alexa Fluor 488 conjugate (Molecular Probes) and Phalloidin-546 (Invitrogen); and counterstained with DAPI (Roche Diagnostics) to visualize cell nuclei. Samples were imaged using an Olympus BX60 microscope (Olympus).
Real-time quantitative reverse transcription–polymerase chain reaction
Two-step reverse transcription–polymerase chain reaction (RT-PCR) was performed on the same cDNA amount for all samples as previously described in accordance with Applied Biosystems manufacturer instructions [26]. Collagen I, collagen IV, fibronectin, laminin, VEGF, Ang1, Ang2, VEcad, ephrinB2, EphB4, Nrp1, Nrp2, and GAPDH (endogenous control) primer sets (Applied Biosystems) were assessed using the comparative computerized tomography method (Applied Biosystems) to calculate the amplification differences between samples. The values for experiments were averaged and graphed with standard error of the mean.
EC maturation and passaging
VEcad-positive cells were isolated from EVC cultures via magnetic activated cell sorting (MACS) as previously described [7]. Briefly, the three classes of EVCs were collected through digestion with MACS buffer, made up of 0.5 M EDTA and 1% BSA in phosphate-buffered saline (PBS); incubated with 10 μL anti-human, PE-conjugated VEcad (BD) in MACS buffer on ice, followed by 20 μL anti-PE microbeads (Miltenyi Biotec) in 80 μL MACS buffer; and washed twice. Resuspended cells were separated based on VEcad expression using an MS MACS separation column (Miltenyi Biotec). VEcad enrichment was confirmed by flow cytometry. Sorted cells were cultured on collagen-type-IV-coated dishes in ECGM supplemented with 50 ng/mL VEGF, 10 μM SB431542, and 0.1% penicillin-streptomycin. Media were changed every other day. For passaging, cells were grown to confluence (∼6–7 days), washed with PBS, digested with TrpLE (Invitrogen), collected, and recultured at 15,000 cells/cm2. To quantify fold expansion, we divided the total number of cells by the number of cells initially seeded at each passage.
Flow cytometry
Flow cytometry was performed as previously described [26]. Briefly, cells were incubated with PE-VEcad or PE-IgG. Forward–side scatter plots were used to exclude dead cells. User guide instructions were followed to complete the flow cytometry analysis via Cyflogic v1.2.
Proliferation quantification
Cell proliferation was quantified as previously described [27]. Briefly, at least 10 different fields of view were imaged at 20× for each triplicate sample. The ratio of Ki67-positive cells to total number of cells was quantified to obtain the percentage of replicating cells.
Arteriovenous fate determination
Arteriovenous fate was calculated as previously described [25]. Briefly, quantitative real-time RT-PCR was performed on derivatives and control endothelial colony-forming cells (ECFCs), human umbilical arterial ECs (HUAECs), and human umbilical venous ECs (HUVECs). All data were normalized to expression by ECFCs. The ratio of ephrinB2 to EphB4 and Nrp1 to Nrp2 was calculated. The natural log of the ratio was taken and graphed versus cell type. This form of analysis yields positive values for arterial cells (e.g., HUAECs) and negative values for venous cells (e.g., HUVECs).
Statistical analysis
All analyses were performed in triplicate samples for n=3 at least. Real-time RT-PCR was also performed on triplicate samples (n=3) with triplicate readings. Two-way ANOVA tests were performed with a Bonferroni post-test (GraphPad Prism 4.02). Significance levels were set at *P<0.05, **P<0.01, and ***P<0.001. All graphical data were reported.
Results
ECM production from EVCs
When we evaluated the production of ECM proteins collagen I, collagen IV, fibronectin, and laminin, we could observe production of all tested proteins except for collagen I in all three types of EVCs (Fig. 1A and Supplementary Fig. S1; Supplementary Data are available online at

Extracellular matrix (ECM) production by early vascular cells (EVCs). Comparison of control, primed, and continuous EVCs for
Growth factor production by EVCs
An important tissue engineering consideration is the production of growth factors to recruit cells to the implanted construct in order to facilitate tissue integration. We evaluated our vascular derivatives for the production of angiogenic growth factors VEGF, Ang1, and Ang2 (Fig. 2). We found that VEGF expression was increased in continuous EVCs compared with primed EVCs, which exhibited more VEGF expression than control EVCs. We suspect that this difference may be due to the nature in which the cells were differentiated, as many previous studies identify VEGF as a downstream product of hypoxic environments [28]. Ang1 expression was greatest in control EVCs compared with primed or continuous EVCs. Conversely, Ang2 followed an opposite trend; expression was greatest in continuous EVCs, followed by primed EVCs and then control EVCs.
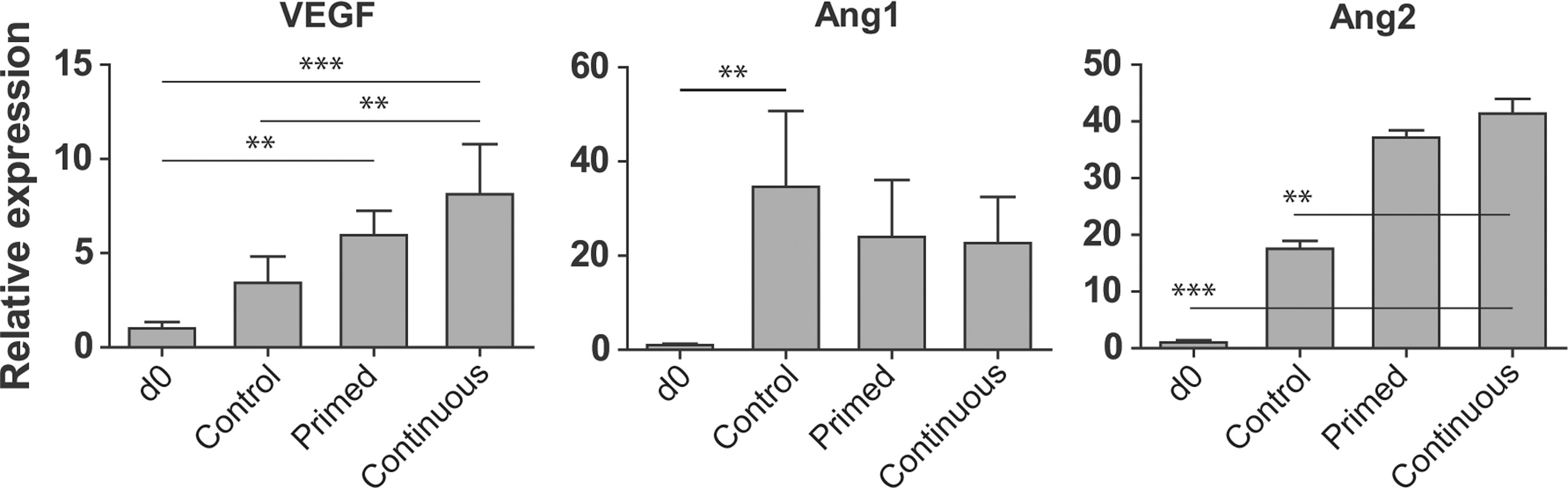
Growth factor production by EVCs. Quantitative real-time RT-PCR analysis of vascular endothelial growth factor (VEGF), angiopoietin 1 (Ang1), and Ang2 production by EVCs. **P<0.01, and ***P<0.001.
Expansion of isolated ECs
Few previous studies have examined whether ECs derived from hPSCs are able to be expanded and propagated in culture to obtain a clinically relevant quantity of ECs [29], although this is an important consideration for therapeutic use. Our previous study demonstrated that under low-oxygen tension, hPSCs preferentially differentiated toward EC fate [25]. To determine whether our derived cells could be passaged, we sorted and cultured VEcad-positive cells (i.e., ECs) from all three classes of EVCs, labeling the derived ECs according to their differentiation procedure and assessing whether the derivation procedure influences the propagation potential of derived ECs. After each passage, cells were reseeded at identical cell densities of ∼15,000 cells/cm2. We found that after two passages, derived ECs exhibited similar growth rates of approximately a three- to fourfold increase over the initial cell seeding density regardless of EVC type (Fig. 3A). However, at passages three and four, we observed that control ECs exhibited a significantly decreased proliferation capacity compared with continuous ECs. ECs derived via the continuous 5% O2 method consistently exhibited the greatest propagation capacity at each serial passage, followed by primed ECs and control ECs, although the propagation capacity gradually diminished over subsequent passages. To confirm proliferative potential of the EC derivates, ECs at passage 4 were stained with proliferation marker, Ki67 (Fig. 3B). Quantification of Ki67 expression revealed the greatest proliferation in continuous ECs, followed by primed ECs, and then control ECs, corroborating our data obtained via serial passaging. Upon evaluation of VEcad expression during serial passaging, we detected that, at early passages, VEcad was enriched in all derived EC populations, which exhibited >90% positive VEcad expression (Fig. 3C). At passage 4, however, cells appeared to lose their VEcad expression, dropping to levels between 30% and 70% VEcad positive.

Expansion of isolated endothelial cells (ECs). Isolated ECs were serially passaged and assessed for
To assess how many times the isolated ECs could be passaged in their differentiation medium, we performed quantitative real-time RT-PCR on cells between passages 1 and 4. We found that VEcad expression significantly dropped at passage 4, but stayed at similar levels between passages 1 and 3 (Fig. 3D). These passaging data suggest that our isolated ECs could be expanded for three passages, ensuring EC phenotype. Extrapolating from these data, we quantified how many ECs could theoretically be obtained up to passage 3 per input hiPSC at day 0, taking into account VEcad+ yield on EVCs, and cell growth expansion data per EC type. This analysis revealed that we could obtain ∼24 ECs per input hiPSC on day 0 using the continuous differentiation protocol (Fig. 3E); ∼11 and 4 ECs could be derived per input hiPSC using the primed or control differentiation protocol, respectively.
Characterization of derived ECs
We sought to examine whether isolated and expanded ECs differ in ECM production and arteriovenous fate from EVCs (Fig. 4 and Supplementary Fig. S2). As observed in EVCs, collagen I was not produced by any of the tested ECs at passage 2. Collagen IV appeared to be produced in a fibrous manner in all cultures; interestingly, production of collagen IV by all derived ECs appeared to be greater than that by EVCs (from Fig. 1A). Fibronectin produced by primed and continuous ECs formed fibrous networks throughout the cultures, whereas fibronectin produced by control ECs appeared localized to the cells. As seen with EVCs, laminin appeared perinuclearly around the cells (Fig. 4A). We next compared ECM production between derived ECs at passages 1 and 4 to elucidate whether this property varies with continued passaging (Fig. 4B). We found that collagen IV expression did not change drastically over the passages; intriguingly, the expression levels were higher than those seen from EVCs (from Fig. 1B). Similarly, fibronectin expression did not vary between passages (Fig. 4B), but exhibited much higher expression levels compared with EVCs (from Fig. 1B). Laminin was the only protein detected to decrease over serial passages; however, this difference was not significant. Laminin expression levels were comparable to those seem from EVCs in Fig. 1B.
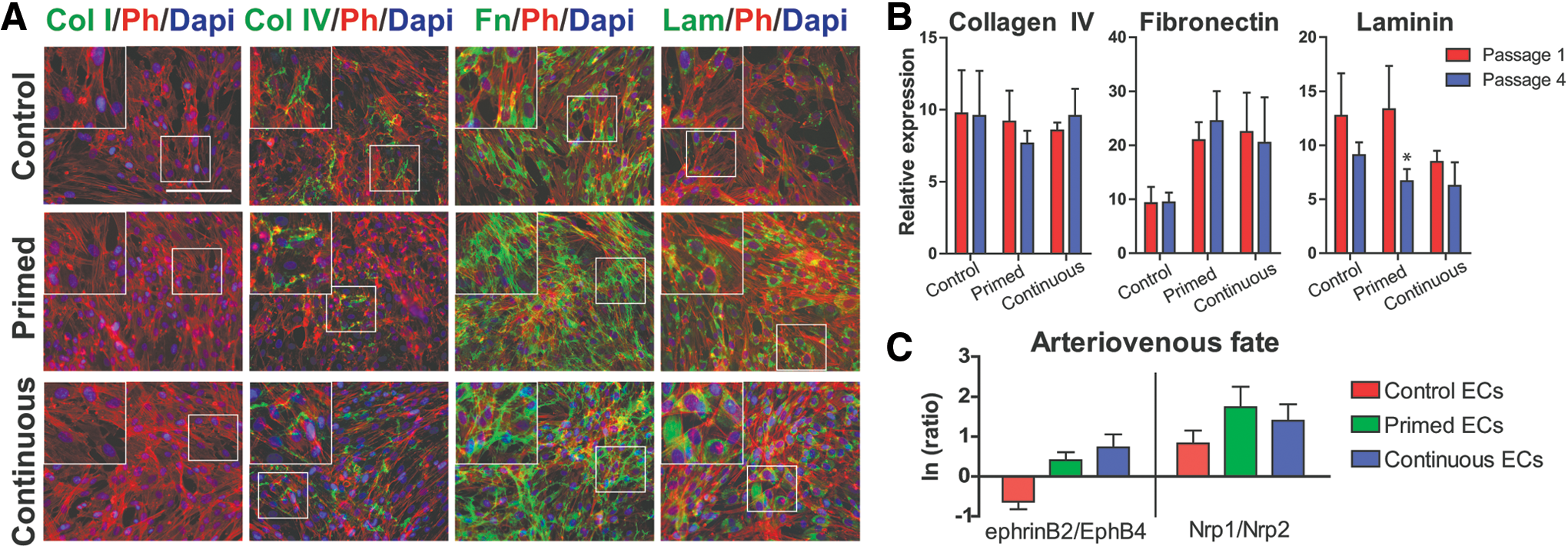
ECM production and arteriovenous fate of derived ECs. Comparison of control, primed, and continuous ECs for
Our previous work revealed that primed and continuous EVCs took on an arterial-like fate, whereas control EVCs did not demonstrate a preference toward arterial or venous fate [25]. To further characterize our EC derivatives, we assessed arteriovenous marker expression at passage 2 (Fig. 4C). As we previously described [25], we assessed our derivatives for the expression of arterial markers, ephrinB2 and Nrp1, and venous markers, EphB4 and Nrp2. We took the natural log of the ratio of ephrinB2 to EphB4 and Nrp1 to Nrp2, yielding positive values for cells exhibiting an arterial phenotype and negative values for cells exhibiting venous phenotype. Isolated primed or continuous ECs more closely resembled an arterial fate, whereas control ECs did not demonstrate a clear fate specification.
Discussion
In this study, we uncover important tissue engineering properties of our hiPSC vascular derivatives, namely, ECM and growth factor production, and propagation potential. Vascular derivatives, including both EVCs and derived ECs, obtained under low-oxygen differentiation conditions demonstrated a greater propensity to produce ECM proteins collagen IV, fibronectin, and laminin. In our previous studies, we also document a correlation between hypoxic conditions and ECM production [26,30]; hypoxic exposure promoted ECM deposition from mature ECs and yielded a more fibrous ECM ultrastructure from cocultures of fibroblasts and breast cancer cells. In both our previous studies and current study, we could not observe collagen I deposition from any types of ECs tested in atmospheric conditions; however, 5% O2 conditions stimulated collagen I production by ECs harvested from human umbilical vein or artery [26]. Collagen IV and fibronectin production by the three classes of EVCs was markedly less than that by derived ECs. In our previous study, we compared ECM production by mature ECs (HUVECs and HUAECs) and an adult stem cell population, ECFCs, and found that only ECFCs were able to deposit ECM proteins collagen IV, fibronectin, and laminin; production by mature ECs was sparse [26]. Comparing these results to our current work, we note that our isolated and derived ECs are able to produce copious amounts of these ECM proteins and to a greater extent than the mature ECs in our previous study. ECFCs had appeared to reorganize their ECM into a web-like network; whereas, in our current work, most instances of collagen IV and fibronectin appeared as a fibrous mesh network without evidence of organization. Of all our tested cell types, continuous ECs produced fibronectin in more of a web-like fashion, reminiscent of fibronectin production by ECFCs. These observations impart another advantage to using ECs derived and isolated from hPSCs over mature ECs.
Quantitative RT-PCR assessment of growth factors reflects production by the entire cellular populations, which inherently have varying ratios of producing cells. Our studies demonstrate that control EVCs harbor a greater propensity to produce Ang1 whereas primed and continuous EVCs tend toward Ang2 production. These results are in agreement with numerous previous studies that connected Ang1 production to pericytes and Ang2 production by ECs [16]. In our EVC populations, ECs make up ∼20%, 39%, and 44% of the population in control, primed, and continuous EVCs, respectively. The remainder of the population is primarily composed of derived pericytes; thus, it is not surprising that control EVCs, which are composed of more pericytes than the other two populations, demonstrated the greatest expression of Ang1. Correspondingly, the populations made up of more ECs demonstrated greater Ang2 expression.
Intriguingly, VEGF production is typically associated with pericytes; yet, our populations made up of larger fractions of ECs (i.e., primed and continuous EVCs) exhibited greater VEGF production. We suspect that this may be due to the fact that hypoxia acts upstream of VEGF activity and thus influences VEGF production in our derivatives [28]. Increased VEGF production is indeed an advantage to use these EVC populations as, when transplanted, this property can help recruit host vasculature and tissue. As demonstrated in our previous study, EVCs derived under continuous or primed hypoxic condition were also able to form cord-like structures faster than control EVCs [25]. In light of our new findings, it may be that VEGF produced by the hypoxic-derived cells plays role in augmenting this functionality.
Propagation of isolated ECs is useful for rebuilding larger, more complex, vascular structures. Upon serial passaging of our derived ECs, we found that ECs derived under hypoxic conditions exhibited the greatest fold expansion. Even after four passages, continuous ECs could grow threefold. We hypothesize that this differential potential may be due to the hypoxic nature of the differentiation conditions; indeed, previous studies have demonstrated increased proliferation under hypoxia during angiogenesis [31]. Here, we extend that notion to encompass cells even previously treated with hypoxia retain a slightly increased proliferation potential. We speculate that these differences in proliferation potential may be due to the differentiation status of the populations as cell viability is not compromised as we have previously shown [25]. However, upon assessment of EC potential after serial passages, we observed that sorted and matured ECs lost some VEcad expression by passage 4, warranting future work that will examine means to retain endothelial potential over numerous passages via media or culture optimization. These findings are in agreement with a previous study, which also demonstrated loss of vascular EC marker expression from mature, fully differentiated ECs (i.e., human umbilical vein ECs) and their subsequent transdifferentiation toward lymphatic ECs in vitro [32]. Moreover, previous studies report that PSC-derived ECs lose their endothelial potential over serial passaging, perhaps owing to the notion that these derivatives may still exhibit some plasticity [33]. These discrepancies highlight a major challenge in the field of vascular tissue engineering to retain endothelial potential over serial passages. Future work in this area focused on specific culture conditions or leveraging certain signaling pathways may help to overcome this drawback.
Finally, the addition of hypoxic conditions to our EVC differentiation scheme and the subsequent passaging of isolated ECs up to passage 3 allowed us to obtain over 20 derived ECs per input hiPSC. The theoretical number of continuous ECs derived was much greater than those for primed or control ECs, again suggesting that a continuous hypoxic differentiation condition stimulates EC propagation, resulting in larger numbers of ECs for transplantation.
Collectively, we demonstrate that our vascular derivatives hold great potential to improve tissue engineering strategies through ECM deposition, growth factor production, and EC propagation; further studies focused on lengthening the maintenance of isolated ECs in culture could yield even greater quantities of hiPSC ECs.
Footnotes
Acknowledgments
The authors thank Dr. Linzhao Cheng for providing hiPSC line. We gratefully acknowledge support for this work by predoctoral award from NIH grant F31HL112644 (to S.K.), NIH grants R01HL107938 and U54CA143868, and National Science Foundation grant 1054415 (to S.G.).
Author Disclosure Statement
The authors disclose no conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.