
Abstract
Lymphedema is a medically irreversible condition for which currently conservative and surgical therapies are either ineffective or impractical. The potential use of progenitor and stem cell-based therapies has offered a paradigm that may provide alternative treatment options for lymphatic disorders. Moreover, basic research, preclinical studies, as well as clinical trials have evaluated the therapeutic potential of various cell therapies in the field of lymphatic regeneration medicine. Among the available cell approaches, mesenchymal stem cells (MSCs) seem to be the most promising candidate mainly due to their abundant sources and easy availability as well as evitable ethical and immunological issues confronted with embryonic stem cells and induced pluripotent stem cells. In this context, the purpose of this review is to summarize various cell-based therapies for lymphedema, along with strengths and weaknesses of these therapies in the clinical application for lymphedema treatment. Particularly, we will highlight the use of MSCs for lymphatic regeneration medicine. In addition, the future perspectives of MSCs in the field of lymphatic regeneration will be discussed.
Introduction
T
Lymphedema is a progressive and lifelong condition for which no curative treatment exists. To manage the complication, several conservative treatments have been proposed, such as pharmacotherapy, physiotherapy, and compassion garments [4], and, occasionally, surgical treatment options, including liposuction, lymph node transfer, and lymphaticolymphatic bypasses, also have been tried [5 –7]. However, these therapies are not very effective and it is also difficult to identify and preserve the lymphatic vessels even using microsurgical methods. In addition, with regard to lymph node transfer therapy, donor site morbidity is the major shortcoming. Furthermore, recent studies have demonstrated the molecular components involved in lymphatic development, which simultaneously points to a promising means of employing lymphatic-specific growth factors to alleviate tissue consequences of lymphatic vascular insufficiency [8 –12]. In this context, vascular endothelia growth factor C (VEGF-C) gene therapy via adenoviruses, naked plasmids, as well as direct application of recombinant VEGF-C were suggested to reduce edema in several preclinical animal models of lymphedema [13 –16]; however, several concerns still remain regarding these therapies, including finite half life of recombinant proteins, short effect of gene therapy, as well as blood vascular side effects and poorly functional lymphatic hyperplasia resulting from employment of high levels of VEGF-C [17,18].
Alternatively, cell-based therapy using progenitor and stem cells has provided a promising hope for lymphatic regeneration. Initial preclinical and clinical studies have evaluated the therapeutic potential of lymphatic endothelial progenitor cells (LEPCs) for regeneration of lymphatic vessels. Furthermore, recent basic research has revealed the potential of embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSCs) to differentiate toward lymphatic endothelial lineage. Meanwhile, scientific interest also shifts to mesenchymal stem cells (MSCs) approaches, as they are available from autologous donation; therefore, this eliminates ethical and immunological disputes (compared with ESCs) and other concerns associated with limited proliferative capacity (compared with LEPCs) as well as complicated manipulation procedures (compared with iPSCs). Besides, basic and preclinical studies have suggested that the use of autologous MSCs as cellular therapies for lymphatic regeneration is feasible and potentially safe and effective in clinical applications.
In this review, keeping in consideration a broad readership, first, we present basic information about the specific markers and transcription factors during embryonic lymphangiogenesis. The focus of this review is on the cell-based therapy for the growth of new lymphatic vessels. In this regard, we will give an overview of LEPCs, ESCs, and iPSCs therapy (Table 1); particularly, we will focus on the promise of MSCs for cell-based lymphatic regeneration therapy, by comparison to other cell approaches.
Phase 0, pre-clinical trials.
LEPCs, lymphatic endothelial progenitor cells; VEGF-C, vascular endothelia growth factor C; ESCs, embryonic stem cells; iPSCs, induced pluripotent stem cells; hiPSCs, human induced pluripotent stem cells; BM-MSCs, bone marrow-derived mesenchymal stem cells; ASCs, adipose-derived stem cells; MSCs, mesenchymal stem cells; VEGFR-3, vascular growth factor receptor-3.
Specific Markers and Transcription Factors During Embryonic Lymphangiogenesis
In mice, the development of lymphatic system starts from expression of prospero-related homeobox transcription factor (Prox-1), which is initially localized at a restricted subpopulation of endothelial cells in anterior cardinal veins at E10 [19]. In addition, studies of Prox-1-null embryos showed normal budding of vascular endothelial cells but failure to give rise to the lymphatic system, revealing the necessities of Prox-1 to specify lymphatic endothelial cell (LEC) phenotypes in embryonic lymphatic development [20]. In other words, Prox-1 is a critical marker determining the fate of lymphatic system. In addition to LECs, Prox-1 is also observed to be expressed in hepatocytes, cardiomyocytes, and neural tube [21,22]. At E10.5, high levels of vascular growth factor receptor-3 (VEGFR-3) expression was detected in the budding Prox-1 expressing cells, concomitant with decreased expression in arteries and veins, indicating a role of VEGFR-3 in blood vessel function [20]. In this regard, VEGFR-3 null mice displayed abnormal vascular remolding and maturation, accordingly, resulting in fluid accumulation and a disorganized cardiovacular system before the emergence of lymphatic vessel [23]. LYVE-1, a lymphatic endothelial HA receptor expressed on both the luminal and abluminal surfaces of lymphatic vessels, resembles the expression of Prox-1 at E10.5 [24]. However, the difference between the expression pattern of LYVE-1 and Prox-1 is that the expression of LYVE-1 is distributed uniformly in the cardinal veins, while that of Prox-1 is restricted into a subset of endothelial cells, suggesting different expression mechanisms of the two markers [20,25]. Despite high expression of LYVE-1 in lymphatic capillaries, it is downregulated in lymphatic collecting vessles [26]. However, the complete function of LYVE-1 is unknown, for mice deficient for LYVE-1 displayed normal lymphatic function, including trafficking of dendritic cells and resolution of skin inflammation [27]. Moreover, an investigation by Kulkarni also displayed the role that nuclear factor activated T-cells cytoplasmic calcineurin-dependent 1(NFATc1) play in the migration of endothelial cells and lymphatic patterning via the calceneurin-NFATc1 signaling [28]. Besides Prox-1 and NFATc1, podoplanin, a transmembrane glycoprotein, also contributes to the migration of endothelial cells, as podoplanin knockout mice displayed lymphedema and abnormal lymphatic function due to the impaired migration of LECs [29,30]. Palate receptor CLEC-2, aggregated by podoplanin, could induce the activation of the downstream targets spleen tyrosine kinase (Syk) and lymphocyte cytosolic protein 2 (SLP-76) in order to part lymphatic from blood vessels [31]. After separation from blood vessels, further development of lymphatic system requires postnatal remolding to form collecting vessels, which requires the formation of lymphatic valves and the recruitment of smooth muscle cells. It is noteworthy that ephrinb2 and transcription factor Foxc2, respectively, are involved in the remolding of collecting vessels [26,32].
Besides specific markers, transcription factors, such as sex determining region Y box 18 (SOX18) and orphan nuclear receptor (COUP-TFII), are also suggested to be involved in the process of lymphendothelial differentiation. SOX18, an SRY-related transcription factor, is a molecular switch inducing Prox-1 expression in venous LEC progenitors. At 9 days post coitum (d.p.c), SOX18 is expressed in a subset of endothelial cells, which is before that of Prox-1. Later, cells expressing SOX18 coupled with Prox-1 migrate to form the primary lymphatic sacs [33,34]. In addition, it has been proposed that nuclear hormone receptor COUP-TFII not only contributes to the activation of PROX-1 in embryonic veins through a direct interaction but also associates with the maintenance of Prox-1 expression during the early stage of LEC differentiation [34,35]. Moreover, mir-181 was demonstrated to function as a negative regulator of lymphatic vascular lineage-specific differentiation through inhibition of the translation of gene Prox-1, so was the mir-31 [36,37].
LEPCs-Based Therapy
Putative cell sources of LECs during embryonic and postnatal lymphangiogenesis
Historically, lymphatic vessels are suggested to generate through proliferation and migration of pre-existing LECs. However, recent findings have not only demonstrated lymphanvasculogenesis as a new way of contributing to lymphatic vessel formation, which is not restricted to the definition of embryonic neolymphangiogenesis, but also refer to the postnatal lymphangiogenesis operated by progenitor cells or stem cells. With regard to embryonic lymphangiogensis, the controversy has continued for a long time, mainly focusing on whether lymphatic vessels grow solely through budding and sprouting from the established LECs, or existence of certain progenitor cells may contribute to the developing lymphatic vessels. In early 1909, Sabin proposed that venous endothelial cells sprout to form lymph sacs from which the entire lymphatic system generates and then spreads to surrounding tissues and organs [38]. Later, Srinivasan et al. validated the venous origin proposal by using lineage tracing technology, which revealed that Prox-1 expressing LEC progenitors, other than hematopoietic cells, was the unique, or at least the main source of LECs contributing to lymphatic vessel formation [39]. However, Buttler et al. held an opposite point based on the result that Prox-1 and LYVE-1 positive mesenchymal cells in dermis and mediastinum could integrate into the growing murine lymphatic vessels, indicating the dual origin of embryonic lymphatic development and confirming the differentiation capacity of mesenchymal cell toward lymphatic lineage [40]. Interestingly, avian lymphatics are suggested to possess dual origin. Exactly speaking, avian lymphatic system is fused by two components, one of which is superficial and dermal lymphatics derived from lymphangioblasts [41]. Thus, fundamental research aiming at revealing the mechanisms of embryonic lymphangiogenesis is anticipated. Nevertheless, conclusions can be drawn from works cited earlier that certain cells derived independently from venous endothelial cells might exist and play a crucial role in lymphvasculogenesis. In fact, a pool of endothelial progenitor cells (EPCs) derived from bone marrow (BM) are involved in the postnatal vasculogenesis, which are characterized by their unspecific surface markers and cobblestone-like morphology [42 –44]. In addition, various well-established animal models, including hindlimb, myocardial ischemia, and corneal neovascularization, have proved the capability of bone marrow-derived or ex vivo expanded EPCs to restore blood flow, increase capillary density, and enhance the perfusion [45 –47]. However, the initial findings asserting the existence of LEPCs derived from human fetal liver and cord blood were presented by Salven et al., who defined a distinct population of CD133+CD34+ cells along with the expression of VEGFR-3 as a kind of progenitor cells, which could express a fraction of vascular and lymphatic endothelial specific markers in in vitro culture, covering VE-cadherin, LYVE-1, and podoplanin [48]. Similarly, endothelial cells derived from human cord blood CD133+CD34+ progenitors share similar phenotypes with LECs under incubation with growth factors, though merely with a few portion of cells, they express LYVE-1 and a lacked expression of podoplanin, Prox-1, and VEGFR-3 [49]. Intriguingly, under conditioned culture, CD34+VEGFR-3+ progenitor cells isolated from human cord blood were shown to possess the ability to differentiate into lymphatic-specific phenotypes, such as LYVE-1 and Prox-1. Moreover, CD34+VEGFR-3+ cells were further defined as endothelial progenitors through forming tube-like vessels in three-dimensional (3D) collagen or Matrigel [50]. Furthermore, LEPCs were demonstrated to integrate into newly formed lymphatic vessels in various animal models. In this regard, Religa et al. observed incorporation of EGFP+VEGFR-3+ cells into the newly formed lymphatics through transplanting these purified cell populations into FGF-2-induced cornea models. Simultaneously, BM-derived EGFP+ cells were also detected in mice receiving transplanted tumor cells, indicating that LEPCs may contribute to lymphatic regeneration [51]. What is more, podoplanin expressing cells isolated from BM mononuclear cells and cultured with VEGF-A and VEGF-C were demonstrated to possess lymphatic endothelial differentiation capacity. In addition, cultured-isolated pod+ cells were also suggested to incorporate into newly formed lymphatic vessels within corneal, wound-healing, and tumor models. Furthermore, under lymphangiogenic incubation, the number of pod+ cells were increased in BM and peripheral blood (PB), simultaneously with the increased expression of LYVE-1 and VEGFR-3 in PB, which indicates that pod+ cells derived from BM were capable of differentiating more lymphatic and less progenitor-like cells through mobilizing into PB under supplemental stimuli. Besides, pod+ cells were also demonstrated to possess the potential ability to drive lymphangiogenesis by producing lymphangiogenic growth factors, including VEGF-C, ANG1, and bFGF. In addition, isolated pod+ cells also displayed expression of hematopoietic stem cell markers sca-1, c-kit, and flk-1, indicating the potential of hematopoietic origin [52]. As EPCs, which are demonstrated to be safely and efficiently used in randomized clinical trials for the treatment of myocardial infarction [53], a recent pilot study containing patients with lymphedema secondary to breast cancer surgery to axillary lymphadenectomy was performed to contrast autologous cell transplantation therapy with compression sleeve therapy (CST). Compared with CST, patients receiving an injection of CD34+ cells derived from BM reduced the volume of edema as well as associated symptoms of pain and decreased sensitivity, supporting the application of cell therapy in clinical treatment for lymphedema [54].
What is more, terminally differentiated cells of myeloid lineage, such as macrophages, have long been demonstrated to promote lymphangiogenesis through secretion of paracrine mediators, including VEGF-C, VEGF-D, and VEGF-A. However, recently, macrophages are suggested to function as a source of LEPCs to promote postnatal lymphangiogenesis via a direct differentiation manner, chiefly associated with chronic inflammation and tumor progression. For instance, CD11b+CD11c− macrophages, along with the expression of Prox-1 and LYVE-1, not only elicited the formation of lymphatic vessels in an inflamed corneal model but also contributed to the formation of tube-like structures in vitro. Besides, depletion of macrophages resulted in suppression of neo-lymphangiogenesis in cornea model [55]. In addition, by taking advantage of genetic lineage tracing strategy, GFP-tagged BM derived cells were observed to incorporate into the lymphatic vessels surrounding tumors [56]. However, contrary points were provided by Gordon et al. that lymphatic vasculature arises independently of myeloid cell lineage, for which triple expression of GFP, Prox-1, and LYVE-1 macrophages were not found during embryonic genesis using lineage tracing data, excluding the potential of macrophage as LEPCs. Meanwhile, macrophages were suggested to restrain vessel caliber by regulating the proliferation of LEC proliferation [57]. Nevertheless, macrophage-derived LEPCs (M-LEPCs) are frequently detected in inflammatory lymphagiogenesis, arguing for the potential of M-LEPCs in lymphatic regeneration [58].
Limitations of LEPCs-based therapy for lymphatic regeneration
Collectively, except macrophage-LEPCs, circulating BM derived LEPCs mainly refer to those possessing both the endothelial and hematopoietic markers. Under supplemental stimuli, such as trauma and inflammation, LEPCs mobilize to the foci and participate in the newly formed lymphatics through directly differentiating into LECs or secreting growth factors to promote lymphangiogenesis, accompanying with increased endothelial-specific markers and declined hemapoietic markers. Thus, similar to EPCs, LEPCs are monopotent and roughly regarded as stem cells for their unipotent capacity to give rise to lymphatic endothelial-lineage cells as well as to its limited proliferative ability. Despite the potential of lymphendothelial differentiation ability and effective application in various preclinical animal models and clinical trials, however, considering that LEPCs express phenotypes which overlap those of the hematopoietic stem cells, there is concern that much time is needed to collect, isolate, and proliferate the progenitor cells. Besides, poor availability of LEPCs in peripheral blood raises another challenge for its application in lymphedema treatment. For example, only 0.1%–0.3% CD34+VEGFR-3+ cells were isolated from healthy adult blood [48]. Likewise, only a mean of 3–10×109 bone marrow mononuclear cells (BM-MNCs) can be isolated from one patient [59]. Besides, it remains unclear whether the newly formed lymphatic vessels are mainly induced through direct differentiation from LEPCs or an indirect paracrine manner in both preclinical and clinical studies. Therefore, it seems less feasible to apply LEPCs as a treatment option for lymphedema treatment. Thus, refined isolation methods as well as more precisely defined LEPC phenotypes continue to be investigated in order to employ LEPCs in the field of therapeutic lymphangiogenesis, which, in turn, requires a better understanding of expressed specific cell markers during the development of stepwise LECs differentiation process. Furthermore, associated long-term clinical cases are also needed to observe adverse reactions of LEPCs therapy. Meanwhile, limitations of LEPCs therapy for lymphedema treatment also require the application of stem cell-based therapy.
General properties of stem cells
The stem cell-based therapy has been the hot spot for regeneration medicine for years. In the light of lymphatic vessel regeneration, the administration of stem cells may hamper the occurrence of lymphedema in donor site. It is widely accepted that stem cells are characterized by their capacity for self-renewal and multilineage differentiation capacity. In addition, based on the ability of differentiating into various cell types, stem cells are divided into totipotent, pluripotent, and multipotent types. To be exact, totipotent stem cells refer to zygote and blastomeres that are capable of differentiating into cells of all lineages, including extraembryonic tissue. However, pluripotent stem cells are restricted to ESCs and iPSCs, which possess the ability of differentiating into all cell lineages of the adult body. However, compared with totipotent and pluripotent stem cells, multipotent stem cells are merely sufficient to give rise to cells of a given germ layer, which are also regarded as adult stem cells. BM mesenchymal stem cells, adipose-derived stem cells (ASCs), and hemapoietic stem cells are typical multipotent stem cells [60]. A detailed presentation of stem cells used in therapeutic lymphagiogenesis is described next.
ESCs and iPSCs-Based Therapy
ESCs in lymphatic regeneration
In 1981, Martin first derived mouse ESCs (mESCs) from inner mass cells of the blastocyst. In vitro culture, ESCs showed potential to differentiate into cells of all three embryonic germ layers, including endoderm, mesoderm, and ectoderm. Besides, the stem properties of ESCs were further confirmed through transferring them into a mouse blastocyst from which a chimeric mouse containing tissues of all three germ layers was generated [61]. The pluripotency of human ESCs (hESCs) is suggested to resemble that of mESCs. For instance, when cultured under appropriate conditions, hESCs are able to initiate multilineage differentiation programs of a variety of cell types, including cardiomyocytes, epithelial cells, and neural cells. In addition, by virtue of expressing high levels of telomerase, hESCs exhibit tremendous proliferative capacity in an undifferentiated state. Similar to mESCs, the pluripotency of hESCs is verified by injecting them into immune-deficient mice to generate teratomas for regeneration medicine. In fact, both hESCs and mESCs have been demonstrated to own the ability to form derivatives composed of cells expressing gene profiles of three germ layers [62,63]. Accordingly, the establishment of ESCs, undoubtedly, opens an avenue into vascular vessels in ES-derived embryonic bodies (EBs) and may function as an efficient source of cells for vasculature regeneration [64 –69]. In addition, implantation of ESCs or ESCs-derived lineage-committed cells to animal models resulted in regional regeneration of vascular endothelial cells, or even cardiomyocytes and smooth muscle cells [42,65,70,71], implying clinical promise for ischemic and heart disease. However, the first step toward stem cell-based therapy for therapeutic lymphangiogenesis also starts from the administration of ESCs. In this regard, EBs are provided as a useful tool to observe lymphatic vessel formation in vitro, which also mimics the process of lymphatic development in mouse embryos. Distinct from blood vessel formation, the development of LECs had not been detected till 14 days after EBs formation [72,73]. In these cases, addition of lymphangiogenic factors is required for the lymphatic endothelial differentiation in EBs. VEGF-A or VEGF-C, not bFGF, were indentified to potently promote the differentiation of EBs into lymphatic vessels. Besides, Prox-1, LYVE-1, and CD31 expressing cells in the central area of EBs were found close to blood vessels, supporting the vein origin of embryonic lymphatic development [72]. Consecutively, studies from Kreuger et al. reconciled those of Liersch, compared with individual treatment with VEGF-A, which displayed a marked induction of vascular vessel sprouts but a relatively poor effect on LYVE-1 expression, a network of LYVE-1, podoplanin, Prox-1, and VEGFR-3 positive lymphatic vessel structures was induced in EBs through a synthetic effect of VEGF-A and VEGF-C stimulation. Besides, scattered LYVE-1+CD45+ cells were detected distal from the vascular vessels, indicating that they may serve as lymphatic endothelial progenitors developing independently of vascular endothelial cells, which, in turn, supports dual origin of lymphatic development [74]. In addition, VEGFR-2 cells derived from ESCs succeed in differentiating into LECs on OP9 cells, mainly through stimulation by VEGF-C and ANG1 secreted by OP9 [75]. Besides VEGF-C, EBs incubated with epidermal growth factor (EGF) were also demonstrated to lead to the formation of lymphatic vessel-like structures coexpressing CD31 and LYVE-1, strongly supporting the role of EGF in therapeutic lymphangiogenesis. Meanwhile, the lymphangiogenic effects were significantly reduced when supplied with EGFR inhibitors [11]. Taken together, EBs are capable of differentiating into lymphatic vessels under conditioned culture, indicating a therapeutic promise for lymphedema treatment.
Benefits and drawbacks of ESCs-based therapy
Compared with adult stem cells, ESCs possess greater pluripotentiality and proliferative capacity, making ESCs an appealing candidate for the treatment of lymphatic disease. However, ethical debate and immunological barrier should be taken into consideration when applying ESCs in clinical use. In order to overcome immune rejection in recipients, large doses of immunosuppressive drugs are manipulated to reduce the immunogenicity of ESCs, inevitably leading to various complications, such as wound healing, infection, and drug-associated toxicity [63]. Although nuclear transfer technology may potentially be provided to efficiently match the genetic profile with that of the recipients through reprogramming transferred somatic nuclear to cytoplasm of enucleated unfertilized oocytes, scarce source of human oocytes, time-consuming transfer process, as well as controversial ethical issues, undoubtedly, make it practically unreachable to put nuclear transfer into effect. The multi-lineage differentiation ability of ESCs, on the other hand, may hinder its broad application in regeneration medicine for which differentiation from ESCs to certain cell types is barely warranted despite conditioned culture with specific growth factors. Of importance, the heterogeneous property of ESCs may lead to the formation of ES-derived tumors, as a small portion of undifferentiated stem cells may possibly result in tumor formation [61,62]. During embryonic development, lymphatic endothelial cells (ECs) are thought to arise from ECs of cardinal vein and, therefore, acquire lymphatic phenotypes. Accordingly, application of specific ESCs-derived fully differentiated LECs or vascular progenitor cells may be a better choice for regeneration. In these circumstances, purified ES-derived cells, such as VEGFR-2+ cells derived from ESCs, are imperatively required to effectively generate the differentiation program to LECs. However, one should be guaranteed that pluripotent properties of derivates from ESCs are close to those of their natural counterparts. Though mESCs-derived cardiomyocytes and neurons exhibit similar physiological function and phenotypes with those of their natural counterparts, it remains to be further investigated whether ESCs-derived LECs or vascular progenitors would serve as functional cell types for therapeutic lymphangiogenesis in animal models as well as for clinical application [76,77]. Hence, identification of ESCs-derived cells and advanced cell-sorting methods are required to be developed.
iPSCs in lymphatic regeneration
As a previous work suggested, embryonic, fetal, and adult-derived cell populations can be reconstructed to product embryos of various mammalian species by using nuclear transfer techniques, implying the presence of factors that are capable of activating the reprogramming process [78 –81]. In this context, Takahashi and Yamanaka reprogram mouse embryonic and adult fibroblasts into ESC-like pluripotent stem cells via viral transduction of transcriptional factors, including Oct4, Sox2, c-Myc, and Klf4 [82]. Successively, Takahashi et al. and Yu et al. respectively, reversed human fibroblasts to a pluripotent state by using Oct4, Sox2, c-Myc, and Klf4 factors and Oct4, Sox2, Nanog, and Lin28 factors [83,84]. These pluripotent stem cells, denoted iPSCs, also share some similarities with ESCs; for example, mouse iPSCs (miPSCs) were demonstrated to express key ESC genes and generate teratomas in vivo as well as form chimeras after transplantation into blastocysts. Likewise, human iPSCs (hiPSCs) are also suggested to differentiate into multiple cell types in vitro, including neurons, cardiomyocytes, hepatocytes, and cells [85 –87]. Based on multi-lineage differentiation background of iPSCs, there is no doubt that iPSCs may represent an attractive option for regenerative medicine. Actually, induction and isolation of endothelial cells from human and miPSCs have been previously suggested by Narazaki et al. and Taura et al. [88,89]. Moreover, the therapeutic potential of hiPSCs-derived ECs has been effectively assessed in mouse model of scleroderma and peripheral arterial disease, in which enhanced capillary density and improved reperfusion were observed [90,91]. hiPSC-ECs purified based on CD31 expression exhibit functional heterogeneity in nature, displaying arterial, venous, and lymphatic phenotypes. With regard to lymphatic endothelial differentiation, hiPSC-ECs, cultured with the use of supplemental VEGF-C and ANG1, showed a marked increase in the expression of LYVE-1 and podoplanin. In this context, hiPSC-ECs might be of utility to regenerate lymphatic vessels for the treatment of lymphedema [92].
Benefits and drawbacks of iPSCs-based therapy
Taken together, iPSCs may be an appropriate alternative for ESCs without immunological barriers that constrain the exploitation of ESCs-based therapy. Above all, compared with ESCs, the most important contribution of iPSCs to clinical use may be their easily accessible tissue sources, which circumvents the use of ESCs and avoids dealing with nuclear transfer technology. However, several hurdles still need to be overcome before putting iPSCs into clinical practice, including intrinsic predisposition to teratoma formation, viral integration into host cells after factors delivery, poor reprogramming efficiency, and dysfunctional regenerative capacity with acquired abnormalities. Although several approaches, such as using chemicals and proteins to facilitate reprogramming technology, finding more efficient cell types, and a suitable environment to improve iPSCs generation, have been taken to enhance reprogramming efficiency, these methods have not yet solved all the problems currently faced in the clinical use of iPSCs. Of importance, for lymphatic vessel regeneration, such as ESCs-based therapy, guiding iPSCs to terminally differentiated LECs or vascular lineage cells may mostly favor the therapeutic application. Thus, searching for efficient selection markers of iPSCs-derived cells and refined sorting methods is also required for iPSCs-based therapy [93]. What is more, it is noteworthy that further studies of ESCs and iPSCs in animal models as well as in clinical trials need to be exploited in the field of lymphatic vascular regeneration.
MSCs-Based Therapy
Application of MSCs in lymphatic regeneration
MSCs, non-hematopoietic form of postnatal adult stem cells, reside in a number of adult tissues, including BM [94], adipose tissue [95], skin [96], peripheral blood [97], umbilical cord blood (UCB) [98], and dental pulp [99,100], as well as in periodontal ligament [101]. Currently, MSCs are regarded as an attractive candidate for tissue regeneration because of their self-renewal capacity and ability to differentiate into various lineages of mesenchymal tissues [102]. Positive surface markers, such as CD90, STRO-1, CD73, CD105, and CD146 as well as negative markers CD34, CD45, and CD14 are demonstrated to be used in attempts to indentify putative MSCs [103]. BM was the first source demonstrated to contain MSCs [104]. Contrast to UCB-MSCs, which have poor isolation efficiency [105], BM is the main source for the clinical application of MSCs. Under controlled in vitro conditions, bone marrow-derived mesenchymal stem cells (BM-MSCs) have the potential to differentiate into osteogenic, chondrogenic, adipogenic, myosupportive stroma, myogenic, and neurogenic lineages [103]. Besides, endothelial differentiation of BM-MSCs was achieved through cultivation with VEGF, which is characterized by an increased expression of vascular endothelial markers and acquired ability to form capillary-like structures in vitro [106,107]. In addition, potential angiogenic function of BM-MSCs was further determined by implanting cells into isogenic animal models, in which implanted Matrigel-embedded BM-MSCs elicited a robust angiogenic response and resulted in an enhanced vessel density. By taking advantage of retroviral gene making of LacZ, the authors observed that only 10% donor-derived LacZ+ BM-MSCs differentiated into CD31+ and VEGF+ endothelial cells via direct contribution, while the majority of neo-generated blood vessels (BVs) arose from host-derived angiogenic response potentially induced by paracrine effect of BM-MSCs in vivo [108]. Furthermore, the endothelial differentiation capacity of BM-MSCs also shed light on its potential to correct lymphatic dysfunction in human disorders. Recently, Conrad et al. demonstrated that murine BM-MSCs and human peripheral blood-derived MSCs are able to acquire a lymphatic phenotype when exposed to purified recombinant VEGF-C or supernatant from human dermal microvascular endothelial cells (HMVECs) and LECs. Besides, functional expression of chemokine receptor CCR7 on MSCs lines may indicate a paracrine effect of MSCs in recruiting lymphatic endothelial precursors in vivo. Ultimately, an injection of BM-MSCs into a lymphedema model leads to a significant reduction in edema formation and enhanced restoration of lymphatic drainage, indicating clinical value of BM-MSCs in patients with lymphedema [109]. Whether direct differentiation or paracrine effect of BM-MSCs contributes to the formation of lymphatic vasculature, however, remains unclear.
In addition to BM-MSCs, multipotent stem cells within adipose tissue, termed ASCs, seem to be another ideal population of stem cells for regeneration medicine. Besides, compared with BM-MSCs, ASCs seem more promising due to its infinite amount of accessible tissue sources and ease of harvest with little donor site morbidity and patient discomfort. What is more, ASCs not only possess the ability to differentiate along multiple lineage pathways but also exhibit stable and robust proliferation capacity with more than 100 population doublings (PDs) [110]; in contrast, BM-MSCs are only limited to a growth potential of 30–50 PDs after ex vivo expansion [111]. Vitro studies, preclinical animal models, and recent clinical trials have demonstrated the endothelial capacity of ASCs as well as its clinical implication in wound-healing and ischemia applications [112 –114]. It is also noteworthy that the contribution of implanted ASCs to the formation of vasculature in ischemic models is mainly achieved via cytokine-dependant paracrine actions, for ASCs were demonstrated to secrete high levels of angiogenic growth factors [115,116]. With regard to the role of ASCs in lymphatic regeneration, in response to VEGF-C stimulation, ASCs displayed a significant expression of Prox-1, VEGF-C, and VEGF-A, accompanied with a reduced expression of ASCs stem cell markers [117]. Besides, implantation of ASCs in a lymphedema model significantly augmented lymphangiogenesis in a paracrine effect via secreting lymphangiogenic growth factor VEGF-C [118]. Compared with the temporal nature and durability of recombinant VEGF-C, a continuous release of growth factors by ASCs is necessary for a sustained and enhanced lymphangiogenic effect. Collectively, it is speculated that newly formed lymphatic vessels initiated by ASCs are not only through direct differentiation but also through paracrine factor-mediated residual lymphangiogenesis. To further assess the clinical implication of ASCs, 3D tissue-engineering scaffolds were developed to mimic an extracellular matrix-like microenvironment for stem cells to attach, migrate, proliferate, as well as deliver growth factors and provide molecularly tailored biofunctions. Meanwhile, 3D scaffolds allow a spatial control of stem cell differentiation, tissue formation, and, ultimately, direct application in vivo conditions [119]. Actually, dextran and gelatin-based hydrogels have been employed to serve as delivery frameworks to release a sustained dose of growth factors and also appear promising for vascular, cartilage, and cardiac tissue regeneration [120 –122]. Gelatin hydrogel containing VEGF-C has been efficiently applied in lymphatic vessel regeneration as determined by its ability to encapsulate human adipose-derived stem cells (hADSCs) and significantly enhance vessel density as well as reduce dermal edema depth when put into the injury site of a lymphedema mouse model. Expectably, hADSCs stimulated by activator VEGF-C also differentiate toward a lymphatic endothelial lineage, as co-localization of hADSCs and LYVE-1 staining was observed in implantation site [123]. Matrigel is derived from a murine sarcoma cell line and composed of a wide array of components, some of which may contribute to lymphendothelial differentiation. A previous work has demonstrated that blood endothelial cells undergoing tubulogenesis in Matrigel assays exhibit reversibly altered gene conversion to lymphatic phenotypes [124]. Likewise, Matrigel plugs embedded with VEGF-C pre-treatmented ASCs led to an increased number of lymphatic vessels surrounding Matrigels after being implanted subcutaneously, mainly initiated by direct transdifferentiation of ASCs, which is supported by identification of donor ASCs co-expressing GFP and LECs markers podoplanin [117]. Besides, Shimizu et al. observed that unstimulated ASCs injected subcutaneously elicited robust lymphangiogenesis mainly via cytokines secreted by donor ASCs, whereas only a few GFP-positive donor cells were found in the implanted area [118].
More than a decade ago, Gronthos and colleagues suggested the existence of stem cells derived from dental pulp, designated human dental pulp stem cells (hDPSCs) [99], which are superior to other types of MSCs, as they can be easily isolated from extracted adult teeth and the procedure for harvesting hDPSCs is less painful and invasive compared with BM-MSCs. hDPSCs also possess high proliferative capacity with a PD of more than 120, which is almost a three-fold difference with that of BM-MSCs [103]. Moreover, hDPSCs isolated from dental pulps with irreversible pulpitis still possess stem-like properties and multilineage differentiation capacities [125]. Furthermore, hDPSCs can be induced to numerous cell types in vitro, including odontoblasts [126], osteoblasts, adipocytes [127], and neuronal cells [128]. It is noteworthy that the multilineage differentiation capacity could still be retained even after cryopreservation [129]. Dental-derived stem cells have been demonstrated to promote angiogenesis via two distinct fashions, one of which is direct differentiating into LECs and then contributing to newly formed lymphatic vessels. In this regard, the endothelial differentiation of hDPSCs and stem cells from exfoliated deciduous teeth, through stimulation of VEGF-A, was assessed by the formation of capillary-like structures and significant expression of endothelial markers [130,131]. On the other hand, angiogenic factors secreted by hDPSCs also participate in the formation of lymphatic vessels through promoting homing, migration, and proliferation of LECs from donor tissue [132,133]. In these cases, it is speculated that dental-derived stem cells, especially hDPSCs, might have a great clinical potential for lymphatic regeneration, either through direct differentiation toward lymphendothelial lineage or through a paracrine effect by secreting lymphangiogenic cytokines (Fig. 1). Thus, investigations in this field are still anticipated.
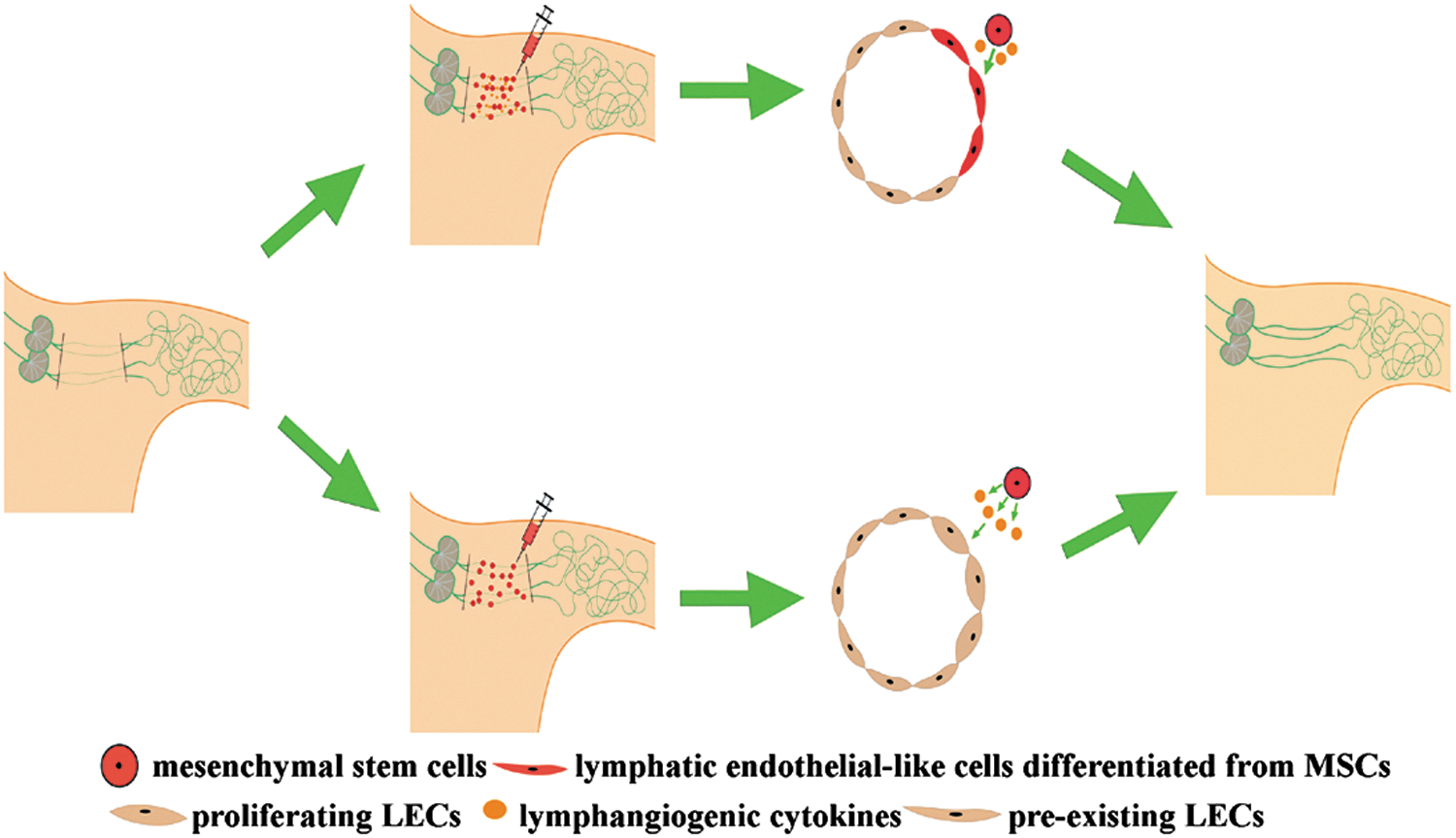
Schematic diagram demonstrating the mechanisms of applying mesenchymal stem cells (MSCs) to reconstruct lymphatic vasculature in an axillary lymphedema model. As shown in the figure, two approaches are potentially suggested to be involved in the lymphatic vessel regeneration. On the one hand, injection of lymphangiogenic cytokines and MSCs leads to lymphatic vessel formation, mainly through direct lymphendothelial differentiation from MSCs under the stimulation of growth factors (shown in the upper approach). On the other hand, implantation of MSCs alone also augments robust lymphangiogenesis in a paracrine manner via secreting lymphangiogenic cytokines (shown in the lower approach). LECs, lymphatic endothelial cells.
Promise and perils of MSCs-based therapy
Taken together, MSCs, whether harvested from BM, adipose, or dental pulp, share a number of traits, making them appealing in the field of lymphatic regeneration, including (1) MSCs are isolated from patients in which the cells are ultimately to be used, circumventing the application of immunosuppression regimes as well as the ethical issues associated with ESCs. (2) As mentioned earlier, MSCs possess the capacity of unlimited self-renewal as well as the potential to differentiate toward a variety of cell lineages both in vitro and in vivo. (3) In contrast to iPSCs, the procedure for harvesting MSCs are easy and the tissue sources are more accessible. (4) Recently, studies have demonstrated the immunosuppressive effects of MSCs, the mechanisms of which mainly lie in the ability of MSCs to down-modulate immune response initiated by immune cells (dendritic cells, T cells, B cells, and NK cells) in vitro and escape immune surveillance in vivo [134]. Of importance, the immunomodulatory potential of MSCs may make up the scarce and time constraint of autogeneic MSCs, leading to a broad administration of allogeneic MSCs that have been suggested to be well tolerated by recipients. Furthermore, MSCs with versatile growth and differentiation potential are ideal candidates for use in regenerative medicine and are currently making their way into clinical trials. As for hMSCs, case studies and clinical trials have demonstrated the success of local or systemic employing of autologously and allogeneicly ex vivo expanded MSCs to initiate for the treatment of several diseases, including bone and cartilage defects, myocardial defect, osteogenesis imperfecta, peripheral ischemia in lower extremities, and autoimmune diseases. Besides, hMSCs are also thought to enhance hematopoietic stem cells (HSCs) engraftment in the course of BM transplantation, to function as vehicles for cancer gene therapy and to treat graft-versus-host disease (GVHD) [135,136]. Similarly, autologous and/or allogeneic ASCs are envisaged to be employed for the clinical treatment of several diseases such as Fistula, limb ischemia, GVHD, liver cirrhosis, osseous and cartilage defects, and bone and breast reconstruction [137]. As mentioned, MSCs can provide therapeutic benefits for lymphatic vessel formation through direct differentiation toward lymphatic lineage [117] and secretion of cytokines [118] to initiate migration, homing, and proliferation of donor cells. Nevertheless, the potential of MSCs therapy has not been exploited in clinical trials for lymphatic regeneration. Thus, well-planned clinical studies are needed to apply MSCs in actual clinical settings outside the laboratory.
However, several hurdles involved in biological characteristics of MSCs need to be solved before fully implementing clinical trials using MSCs-based therapy. First, the limited proliferation capacity of MSCs resulting from replicative senescence may partly hinder its clinical application, as a large body of cells is required to ensure the success of a therapy. By contrast, ES and iPSCs are potentially immortal and they are an unlimited source for tissue regeneration. Classical media used for isolating and expanding MSCs are typically supplemented with fetal bovine serum or human platelet lysate (HPL), raising concerns with regard to unidentified zoonoses, immunological reactions by biomolecules, and complicated procedures for obtaining large amounts of HPL and autologous serum [138]. Recently, serum-free media have been successfully designed to promote the production of MSCs while maintaining its differentiation and immunosuppressive capacity [139]. However, considering donor-to-donor variability in MSCs proliferation kinetics, well-defined serum-free media adjusting to the biological properties of donor MSCs are imperatively needed for clinical application in order to fully maximize the true effects of MSCs therapy in clinical trials. Another concern is whether transplanted ex vivo expanded MSCs may undergo spontaneous transformation, and, more seriously, promote the growth of a latent tumor. Generally, hMSCs in long-term culture has not been considered to harbor the risk of malignant transformation [140]. So far, there have been no clinical reports suggesting tumor formation caused by MSCs therapy [141]. Even so, based on the speculations from studies performed on murine MSCs (mMSCs) which demonstrated that cultured mMSCs led to malignant transformation with changed phenotypes, the potential risk for spontaneous transformation with hMSCs therapy is still highly to be noted [142]. Thus, for each disease type and biological properties of donor MSCs, optimal timing of therapy administration, dose, and passage numbers of donor cells, optimized cell bioprocessing protocols need to be further decided. In addition, with regard to lymphatic regeneration, randomized, controlled, and multicenter clinical trials are urgently needed to optimize the MSCs-based therapy.
Conclusions and Future Perspectives
Progenitor and stem cell-based therapies have gradually gained a great deal of attention for clinical treatment of lymphatic disorders, given the differentiation potential and proliferative capacity of these cell populations. Of the available cell approaches, MSCs seem to be the most promising candidate partly due to their abundance and ease of availability with less donor site morbidity as well as avoidable ethical and immunological barrier. Though its multipotency is limited compared with ESCs and iPSCs, a variety of studies have evaluated its clinical potential in overcoming the current limitation in the field of lymphatic regeneration.
However, as mentioned earlier, much remains to be done to harness the cells for clinical purposes. On the one hand, the limited preclinical studies require large animal models and clinical trials for further evaluation and optimization in order to successfully apply MSCs therapy for lymphedema treatment for which no therapeutic strategies currently exist. On the other hand, the key molecular events and signaling pathways involved in the initiation of stem cell differentiation toward a lymphatic lineage, however, still remain unclear. Thus, integration of basic scientific understanding of mechanisms involved in lymphendothelial differentiation to clinical treatments will inevitably enhance the early promise to mature cell therapies from hope to reality.
Footnotes
Acknowledgments
This study was supported by the National Science Foundation of China (81072216) and Sichuan Provincial Science and Technology Department Foundation of China (2014JY0076).
Author Disclosure Statement
No competing financial interests exist.