
Abstract
Multipotent mesenchymal stromal cells (MSCs) are more and more frequently used to treat orthopedic injuries in horses. However, these cells are limited in their expandability and differentiation capacity. Recently, the first equine-induced pluripotent stem cell (iPSC) lines were reported by us [1]. In vitro differentiation of iPSCs into MSC-like cells is an attractive alternative to using MSCs derived from other sources, as a much larger quantity of patient-specific cells with broad differentiation potential could be generated. However, the differentiation capacity of iPSCs to MSCs and the potential for use in tissue engineering have yet to be explored. In this study, equine iPSCs were induced to differentiate into an MSC-like population. Upon induction, the iPSCs changed morphology toward spindle-shaped cells similar to MSCs. The ensuing iPSC-MSCs exhibited downregulation of pluripotency-associated genes and an upregulation of MSC-associated genes. In addition, the cells expressed the same surface markers as MSCs derived from equine umbilical cord blood. We then assessed the multilineage differentiation potential of iPSC-MSCs. Although chondrogenesis was not achieved after induction with transforming growth factor-beta 3 (TGFβ3) and/or bone morphogenic protein 4 (BMP-4) in 3D pellet culture, mineralization characteristic of osteogenesis and lipid droplet accumulation characteristic of adipogenesis were observed after chemical induction. We demonstrate a protocol for the derivation of MSC-like progenitor populations from equine iPS cells.
Introduction
M
In horses, several reports have been published on the treatment of orthopedic defects using MSCs derived from various sources, typically isolated from the bone marrow and adipose tissue [9]. Although these tissues provide a source of MSCs, collection is an invasive process and the capacity for proliferation and differentiation of these cells declines with the age of the horse [11,12]. In addition, MSC isolation and expansion from bone marrow and adipose tissue take weeks to reach therapeutic cell numbers. MSCs isolated from umbilical cord blood (CB-MSCs) of newborn foals can eliminate some of these issues. The cord blood is collected in a minimally invasive manner, and the cells are phenotypically young when compared to adipose- and bone marrow-derived MSCs from adult donors [13,14]. Cryopreservation allows CB-MSCs to be used as required. The cells can be banked for autologous and allogeneic use [15]. However, there are unique limitations to CB-MSC isolation, in particular failure to collect blood at the time of foaling [16]. Like MSCs from other sources, CB-MSCs are ultimately limited in their proliferation and differentiation potential in vitro. Therefore, there is a need for an MSC source that has greater expandability and high differentiation potential.
Embryonic stem cells are capable of proliferating indefinitely and are considered pluripotent. Equine embryonic-like stem cells have been reported, but no research groups have described equine embryonic stem cells that fulfill all criteria of pluripotency [17]. Therefore, focus has turned to generating equine-induced pluripotent stem cells (iPSCs) as a cell source that could be utilized for developmental studies and cell-based therapies in the horse [1,18,19]. Our equine iPSC lines [1] were generated from fetal fibroblasts using the piggyBac transposon transgene delivery system to transfect the reprogramming factors Oct4, Sox2, Klf4, and c-Myc, as well as green fluorescent protein (GFP) [20]. The cells display pluripotency characteristics: expression of hallmark pluripotency genes, the ability to self-renew, form embryoid bodies, and form teratomas containing all three embryonic germ layers in NOD/SCID mice [1].
Since iPSCs can theoretically be propagated indefinitely, a possible treatment would be to inject these cells directly into the affected individual (systemically or at the site of injury); similar to what has been done with MSCs. However, there is a substantial risk of teratoma formation through direct iPSC injection in wild-type and immunocompromised animals, both syngeneically and allogeneically [21]. The differentiation of iPSCs into specific tissue types before implantation is bound to decrease the risk of teratoma formation, as it reduces both the expandability and pluripotency of the iPSCs [21,22]. Nevertheless, directed differentiation of iPSCs into a homogenous population of differentiated cells is technically difficult and can result in immature and/or inferior tissue with regard to cell and extracellular matrix organization [23]. Therefore, differentiating iPSCs into an intermediate multipotent cell type, such as MSCs, may not only reduce or eliminate the risk of teratoma formation upon implantation but may also provide a more homogenous and mature cell type for tissue engineering [24]. Recently, successful differentiation of human and mouse iPSCs into MSC-like cells has been reported [25 –27] and shows that a high yield of MSCs can be generated with enhanced proliferation and differentiation capabilities compared to adult MSCs [27]. These MSC-like cells appear to incorporate the benefits of both iPSCs and MSCs, making them a promising new cell type for regenerative medicine.
We hypothesized that equine iPSCs are a robust and highly expandable source of MSCs. We report on the differentiation of equine iPSCs into MSC-like progenitor cells under specific culture conditions. The resulting equine iPSC-MSCs are plastic adherent and display a fibroblastoid morphology that resembles MSCs from other sources. They show a downregulation of the pluripotency-associated endogenous genes, Oct4, Nanog, Sox2, and Klf4, an upregulation of the MSC-associated genes, Sox5, Sox6, Sox9, CD29, CD44, and CD90, and an upregulation of mesodermal lineage genes, BMP7 and SNAI3, under induction conditions. In addition, these cells express MSC surface markers CD90, CD29, and CD44 and did not express the markers CD4, CD8, CD73, CD105, MHC I, CD11a/18, CD45, and MHC II. Our equine iPSC-MSCs are capable of undergoing adipogenesis and osteogenesis, but not chondrogenesis in vitro.
Materials and Methods
iPSC culture and maintenance
Equine H3-B iPSCs were grown in Dulbecco's modified Eagle medium-high glucose (DMEM-HG, No. 11960–044; Invitrogen) supplemented with 2 mM GlutaMAX™ (No. 35050; Invitrogen), 0.1 mM nonessential amino acids (No. 11140; Invitrogen), 0.1 mM betamercaptoethanol (No. M7522; Sigma-Aldrich), 1 mM sodium pyruvate (No. 11360-070; Invitrogen), 50 U/mL penicillin/streptomycin (No. 15070; Invitrogen), 15% fetal bovine serum (FBS; HyClone), 1,000 U/mL leukemia inhibitory factor (No. ESGRO; Millipore), 10 ng/mL basic fibroblast growth factor (bFGF, No. 100-18B; PeproTech), and 1.5 μg/mL doxycycline (No. D9891; Sigma-Aldrich). The cells were plated on mitomycin-inactivated mouse embryonic feeder cells and cultured in a humidified incubator at 37°C and 5% CO2. They were passaged every 2–3 days by enzymatic digestion with TrypLE Select (No. 12563; Invitrogen) at a 1:4 ratio.
Derivation of an MSC-like population from equine iPSCs
A serial plating strategy was used to differentiate the iPSC line H3-B [1] into MSC-like cells (Fig. 1). To separate the iPSCs from their mouse embryonic fibroblast feeder layer, the cells were enzymatically digested as described above, centrifuged, and resuspended in the sorting medium containing phosphate-buffered saline (PBS), 25 mM HEPES, and 0.2% FBS. Fluorescent-activated cell sorting of GFP-positive iPSCs was performed using the MoFlo Astrios Cell Sorter (Beckman Coulter). GFP-positive cells were seeded onto gelatin-coated plates at 1 × 104 cells/cm2 in MSC induction media consisting of DMEM-HG, 10% FBS, 0.1 mM nonessential amino acids, 50 U/mL penicillin/streptomycin, 10 μM anaplastic lymphoma kinase (ALK) inhibitor SB431542 (Stemgent No. 04-0010), and 5 ng/mL bFGF. Cells were expanded to 80% confluency and then passaged onto a second gelatin-coated plate for one passage. Hereafter, the cells were passaged onto uncoated tissue culture plates and propagated for 3–11 passages. The cells gradually obtained a fibroblast-like spindle-shaped morphology. ALK inhibitor was removed from the MSC induction media after 10 days. Cells were routinely passaged every 3–4 days, at a 1:3 split ratio, until reaching the eighth passage. This process was reproduced thrice with H3-B iPSCs (n = 3). The resulting iPSC-MSCs were evaluated at an early passage (passage 3–4) and late passage (passage 6–8) by gene expression, flow cytometry, and functional differentiation.
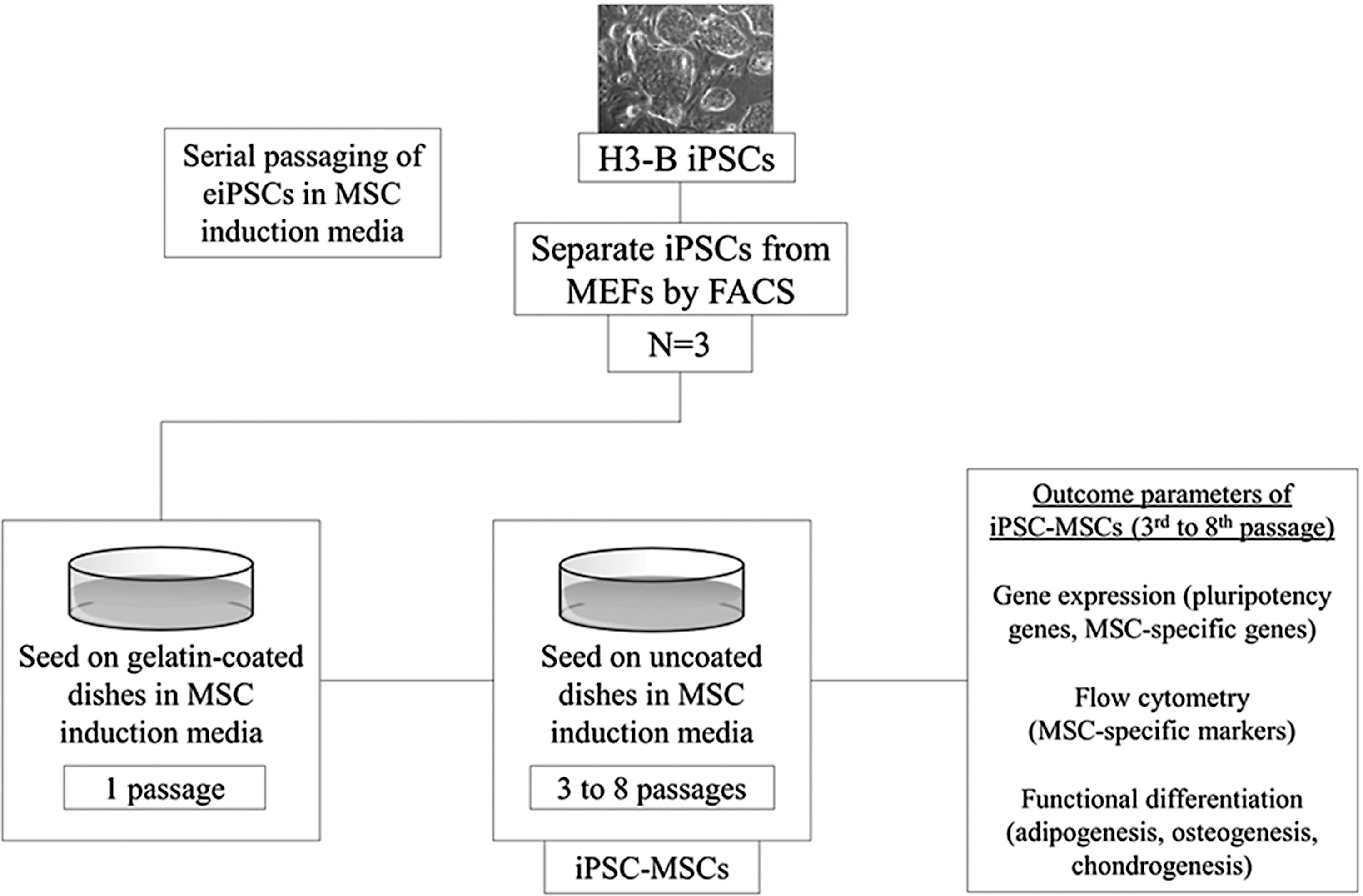
Study design on the derivation and subsequent evaluation of iPSC-MSCs from equine iPSCs. iPSC, induced pluripotent stem cell; MSCs, mesenchymal stromal cell.
Quantitative real-time PCR analyses
Total RNA was harvested from iPSC-MSCs using the Qiagen RNeasy Kit (Qiagen), and 1 μg of total RNA was reverse transcribed using the QuantiTect Reverse Transcription Kit (Qiagen). The reaction was performed at 42°C for 15 min. The cDNA was subsequently analyzed by qPCR using LuminoCt SYBR Green qPCR ReadyMix (Sigma-Aldrich) using the CFX384 Touch™ Real-Time polymerase chain reaction detection system (Bio-Rad). For all samples, a negative control was included that did not contain cDNA. All reactions were completed in triplicate. Values are expressed as 2^DeltaDeltaCt, with DeltaDeltaCt defined as the difference in crossing threshold (Ct) values between experimental and control samples, as described using β-actin and Hprt1 as internal standards. Fold difference was compared to the H3-B iPSC parent line. Oligonucleotide primers are listed in Table 1. Oct4, Sox2, and Klf4 primer pairs do not amplify mouse transcripts from the inserted transgenes [1].
qRT-PCR, quantitative real time-PCR.
Flow cytometry
iPSC-MSCs and CB-MSCs (1 cell line at passage 3) were expanded and subsequently harvested using ACCUMAX (Stem Cell Technologies) and washed with flow buffer (PBS, 5 mM EDTA, 1% horse serum, and 0.1% sodium azide). The following primary antibodies were used to stain the cell suspensions (150,000–200,000 cells): mouse anti-human CD73 (Abcam), mouse anti-human CD90 (VMRD), mouse anti-human CD105 (ABD Serotec), mouse anti-horse CD44 (ABD Serotec), fluorescein isothiocyanate (FITC) mouse anti-human CD29 (Beckman Coulter), mouse anti-human CD45 (VMRD), mouse anti-horse CD4 (ABD Serotec), mouse anti-horse CD8 (ABD Serotec), mouse anti-horse CD11a/18 (ABD Serotec), mouse anti-horse MHC I (ABD Serotec), and mouse anti-horse MHC II (ABD Serotec). Rat anti-mouse IgM-FITC and goat anti-mouse IgG1-FITC were used as secondary antibodies. Specificity of all antibodies has been validated in our laboratory using positive controls for the target species of the antibody (horse or human) and with three negative controls: isotype-negative control, secondary antibody, and unstained control [28]. Ten thousand events were acquired for each experiment on BD FACScan (BD Biosciences) using CellQuest (Becton Dickinson) software, and analysis was conducted using FlowJo (Tree Star).
Osteogenesis assay
Osteogenic differentiation was induced as previously described [2]. In brief, passage 4 iPSC-MSCs were seeded in six-well plates at a density of ∼3,000 cells/cm2 and grown until 90%–100% confluency. To induce osteogenesis, cells were cultured for 10 days in osteogenic media containing Dulbecco's modified Eagle medium-low glucose (DMEM-LG), 10% FBS, 1%
Adipogenesis assay
Induction of adipogenesis was performed as previously described [29]. Passage 4 iPSC-MSCs were seeded at a density of ∼2.1 × 104 cells/mL in six-well plates. Adipogenesis was carried out in adipogenic induction media, containing DMEM-LG, 15% rabbit serum, 500 μM IBMX, 1%
Chondrogenesis assay
Passage 6 and 11 iPSC-MSCs were induced to undergo chondrogenesis in pellet culture as previously described [30]. Approximately 2.5 × 105 iPSC-MSCs were resuspended in 200 μL of chondrogenic induction media, containing high-glucose DMEM (DMEM-HG), 200 mM GlutaMAX, 100 mM sodium pyruvate, 1% ABAM, 0.1 mM dexamethasone, 100 mg/mL ascorbic acid-2 phosphate, 40 mg/mL proline, 1× insulin, transferrin, and selenious acid (ITS) premix (No. 515 00-056; Life Technologies), and 10 ng/mL transforming growth factor-beta 3 (TGFβ3, No. 243-B3; R&D Systems). The resuspended cells were plated in V-bottom polypropylene 96-well plates (No. MPG-651201; Phenix) and centrifuged to pellet the cells at the bottom of the wells. Bone morphogenic protein 4 (BMP-4, No. E12512; Roche) was added alone or in conjunction with TGFβ3 in various final concentrations (10, 50, and 100 ng/mL) to the chondrogenic media in a subset of pellets. BMP-4 was removed from the media after 1 week of pellet culture, as previously described [11]. Pellets were incubated at 37°C in 5% CO2 and 21% O2 for 2–4 weeks. Media were changed every 2–3 days.
Statistical analysis
GraphPad Prism 6 Software was used to perform statistical analysis on qRT-PCR data. One-way ANOVA, followed by Tukey's post hoc test, was used to determine expression differences between groups. P-values <0.05 were considered to be statistically significant.
Results
Derivation of iPSC-MSCs from equine iPSCs
Derivation of MSCs from iPSCs has been done indirectly through the formation of embryoid bodies and subsequent selection of adherent outgrowths, or directly through the addition of specific differentiation media [26,31,32]. Since attempts at propagating adherent cells from embryoid bodies were unsuccessful (data not shown), we opted to explore the latter method and include serial passaging onto uncoated Petri dishes to select for a more homogenous population of MSC-like cells. After 8 days on gelatin in MSC induction media, the iPSCs lost their colony forming phenotype and acquired a mixed cuboidal and spindle-like morphology (Fig. 2a, b). Upon subsequent passaging onto uncoated Petri dishes, the spindle-like morphology predominated (Fig. 2c) and eventually the majority of the differentiated population maintained this MSC-like phenotype (Fig. 2d).
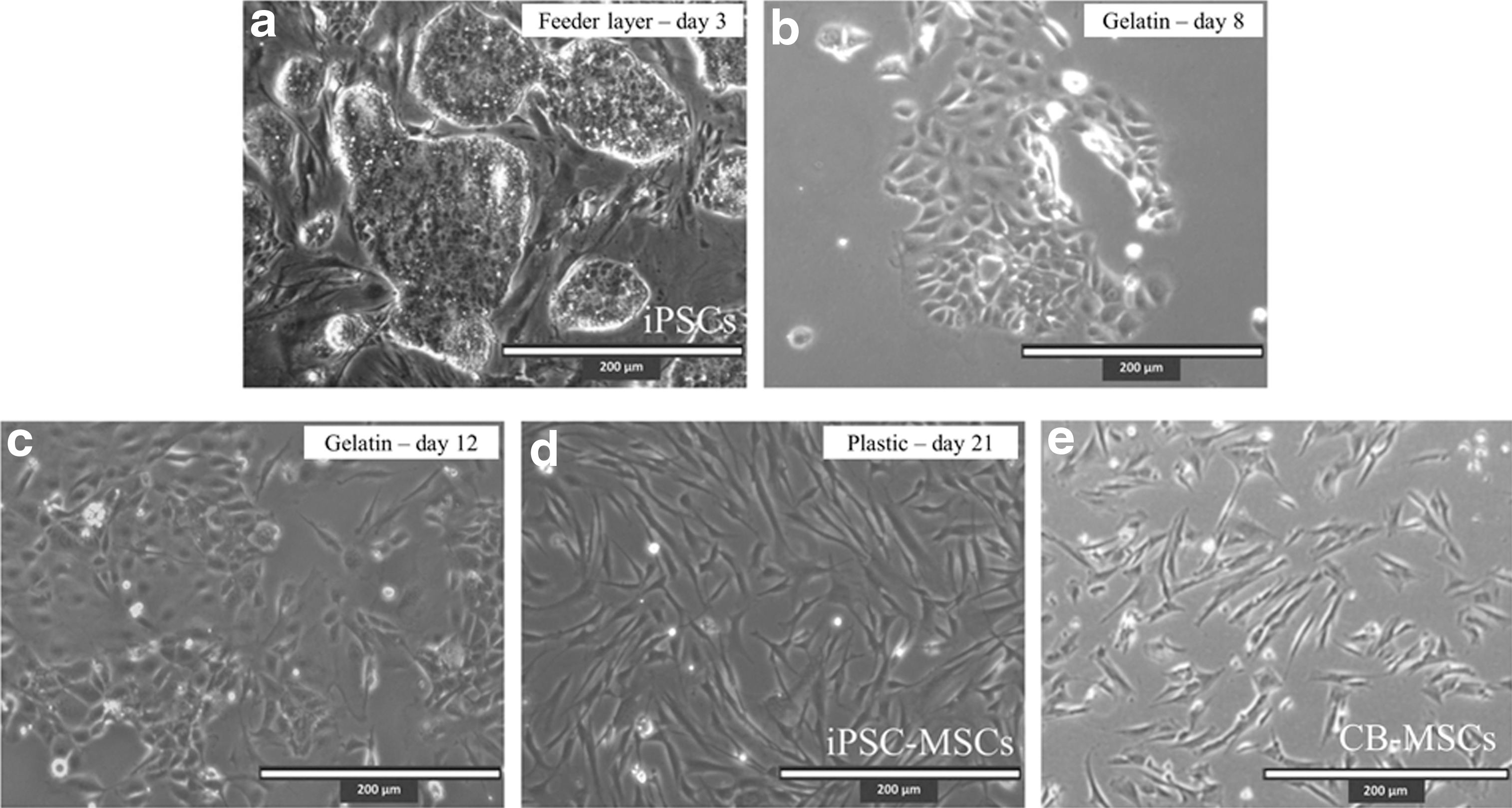
Derivation of an MSC-like population from equine iPSCs. iPSCs maintained in iPSC media form ES-like colonies
Gene expression of iPSC-MSCs
To further characterize and classify iPSC-MSCs, gene expression analyses were conducted. Gene expression of equine pluripotency markers (Oct4, Sox2, Nanog, and Klf4) in three separately derived iPSC-MSC lines was compared to their parent H3-B iPSC line. In both early passage (passage 3–4) and late passage (passage 6–8) cells, iPSC-MSCs showed consistent downregulation of expression in each gene relative to H3-B iPSC (Fig. 3a). In addition, iPSC-MSCs displayed significant upregulation of MSC-associated genes Sox5, Sox6, Sox9, and CD90 as well as an upregulation of mesodermal lineage genes BMP7 and SNAI3 relative to H3-B iPSC (Fig. 3b). In addition, Sox5, Sox6, Sox9, and CD44 and CD90 transcription levels approached that of CB-MSCs.
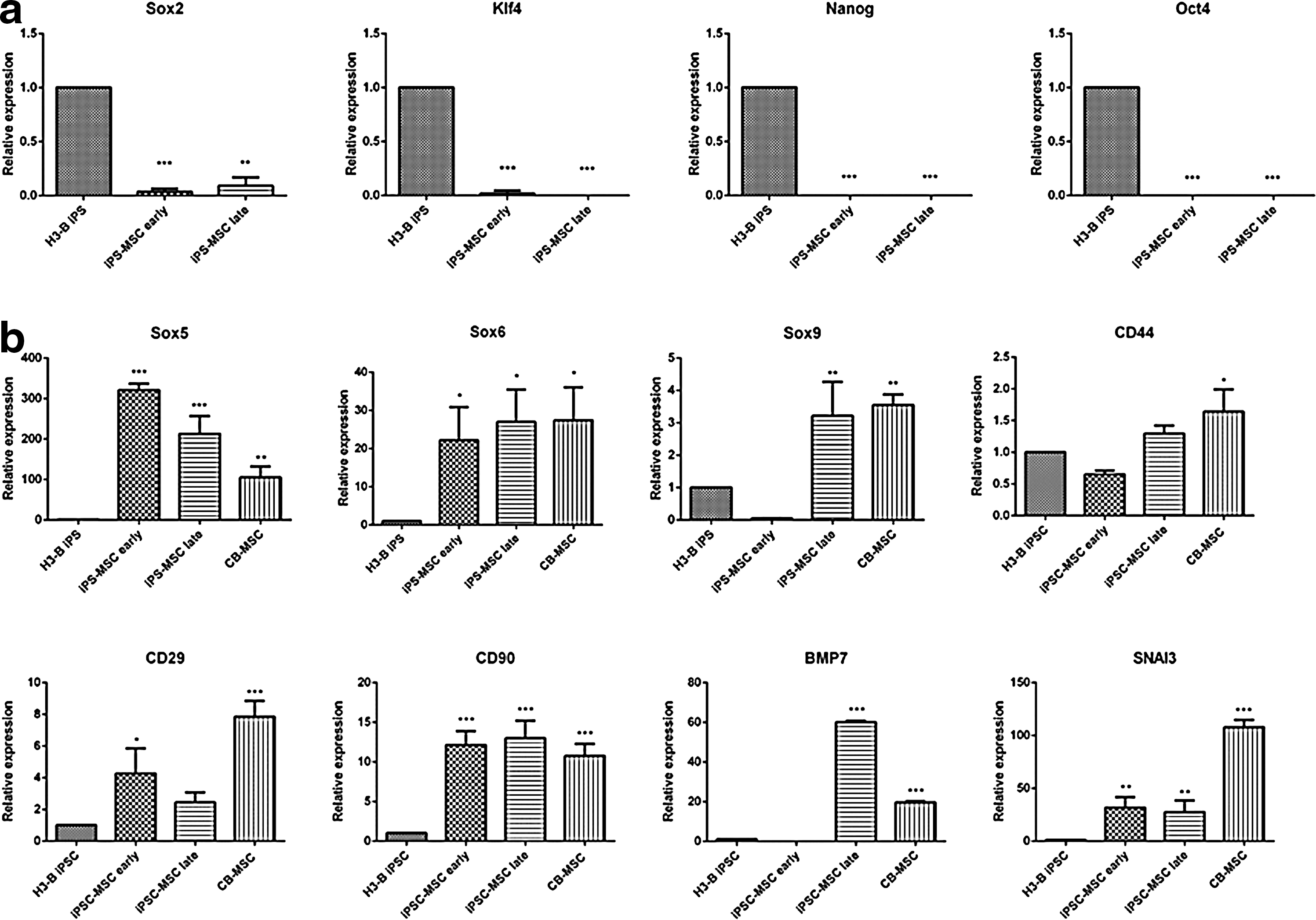
Gene expression analysis of pluripotency and mesenchymal/mesodermal-related genes by qRT-PCR.
Flow cytometry analyses and derivation efficiency of iPSC-MSCs
Another criterion for the classification of MSCs is expression of a certain panel of surface markers [33]. As equine MSCs lack a standardized set of surface markers associated with MSCs of various sources, our laboratory has established surface antigen expression for MSCs derived from cord blood [28]. Upon flow cytometry analysis, we observed that iPSC-MSCs display a similar immunophenotype as CB-MSCs, namely, positive expression for CD29, CD44, and CD90 and low or absent expression of CD4, CD8, CD105, CD45, CD73, CD11a/18, MHC I, and MHC II (Fig. 4a, b). Of cells that remained plastic adherent throughout the MSC induction process, 74.4% of iPSC-MSCs expressed CD90, 97.5% expressed CD29, and 97.7% expressed CD44 (Fig. 4). As these values are nearly identical to the percent-positive cells in CB-MSCs (CD90: 76.6%; CD29: 97.0%; and CD44: 90.9%; Fig. 5), MSC derivation efficiency of adherent iPS cells using our serial plating method is close to 100%.

Flow cytometry analysis of one representative iPSC-MSC line for expression of common surface antigens. Dot plots generated from flow cytometry analysis show percent-positive cells (FITC positive) after gating around the isotype control population (top row: IgM secondary antibody; bottom row: IgG secondary antibody). The unstained control is shown as being gated around the IgG-negative control. FITC, fluorescein isothiocyanate.

Flow cytometry analysis of one representative CB-MSC line for expression of common surface antigens. Dot plots generated from flow cytometry analysis show percent-positive cells (FITC positive) after gating around the isotype control population (top row: IgM secondary antibody; bottom row: IgG secondary antibody). The unstained control is shown as being gated around the IgG-negative control.
Characterization of functional differentiation of iPSC-MSCs
To elucidate the multilineage differentiation potency of iPSC-MSCs, we performed adipogenic, osteogenic, and chondrogenic induction assays in vitro. After 10 days in osteogenic induction media, von Kossa staining and Alizarin Red staining revealed that the iPSC-MSCs acquired mineralization after induction, indicative of early osteogenesis (Fig. 6). Furthermore, after 14 days in adipogenic induction media, morphology changes and lipid droplets were observed in the iPSC-MSCs following Oil Red O staining (Fig. 6). The results obtained were consistent with what was observed in CB-MSCs induced for osteogenesis and adipogenesis for the same time period (Supplementary Fig. S1; Supplementary Data are available online at

In vitro adipogenesis and osteogenesis assays of iPSC-MSCs (200× magnification).
Discussion
Our aim in this study was to determine whether equine iPSCs could be successfully differentiated into MSC-like cells that exhibit properties similar to MSCs from other sources, such as umbilical cord blood. We report that the iPSC line H3-B [1] can be reproducibly differentiated into a progenitor cell type that meets many criteria of MSC: adherence to and proliferation on plastic, expression of certain surface markers, and multilineage differentiation potential. Our iPSC-MSCs display a significant reduction in gene expression of the pluripotency genes Oct4, Klf4, Sox2, and Nanog while upregulating MSC-associated and mesodermal lineage genes Sox5, Sox6, Sox9, CD44, CD90, BMP7, and SNAI3.
Several different techniques have been employed to derive MSCs from iPSCs. These include culture of adherent outgrowths from embryoid bodies [32,34,35] and culture of iPSCs on various surfaces such as fibrillar collagen [36 –38] or through cell selection [27]. Serial plating or direct plating is a comparatively simple method to direct the differentiation of iPSCs to MSCs [26,39,40]. Our method involved simultaneously seeding equine iPSCs onto gelatin-coated plates in MSC induction media, and this was shown to be sufficient in directing their differentiation into the flattened fibroblastic morphology typical of MSCs. A homogenous population of cells resulted after only 2–3 passages on uncoated Petri dishes in MSC induction media; this technique selected for cells that were plastic adherent, as any floating cells were lost when changing media. MSC induction media were found to be required for the differentiation of plastic-adherent cells through the serial passaging method; simply removing doxycycline from the iPS culture media was not sufficient in inducing the cells to differentiate into MSC-like cells (data not shown).
Gene expression analysis confirmed that pluripotency-associated genes Oct4, Sox2, Nanog, and Klf4 were downregulated in iPSC-MSCs upon differentiation. This suggests a loss of pluripotency, which was correlated with an upregulation of MSC-associated genes Sox5, Sox6, Sox9, CD29, and CD90 and an upregulation of mesodermal lineage markers BMP7 and SNAI3. Sox5 and Sox6 are transcription factors involved in the osteochondroprogenitor lineage program and act upstream of Sox9 [41]. The Sox9 transcription factor is the key regulator in chondrogenesis, initiating the expression of cartilage-specific extracellular matrix genes such as Col2a1 [42]. Sox9 is required for the commitment of chondroprogenitors, mesenchymal condensation, and proper chondrocyte proliferation, differentiation, and maturation [43]. Our data revealed that although Sox5 and Sox6 were upregulated in iPSC-MSCs in both early and late passages, Sox9 expression was only upregulated in late passage cells (Fig. 3b). This variable expression of the Sox trio of transcription factors has been observed before in iPSC-MSCs, although chondrogenesis was observed in these cell lines [26].
BMP7 is a member of the TGFβ superfamily and is an early mesodermal lineage marker [44,45]. It has been found to be expressed at numerous sites throughout development. Late passage iPSC-MSCs show a distinct upregulation of BMP7 over H3-B iPSC, although no expression was detected in early passage iPSC-MSCs (Fig. 3b). SNAI3 is a zinc finger protein, a member of the Snail protein family, and is also an early mesodermal marker. Snail proteins function primarily as transcriptional repressors and are capable of inducting epithelial–mesenchymal transition [46]. SNAI3 was upregulated in both early and late passage iPSC-MSCs (Fig. 3b).
Flow cytometry of MSC markers (CD90, CD29, and CD44) predicted a homogenous population of differentiated MSC-like cells successfully derived from iPSCs. This indicates that the efficiency of derivation among plastic-adherent cells is close to 100% (Fig. 4). We chose this efficiency assessment method as it provides the most accurate evaluation of the percentage of cells with an MSC phenotype. However, it must be recognized that this is not a true measure of derivation efficiency from undifferentiated iPSCs to MSCs. This is because of the nature of the serial plating method, where some iPSCs were lost during media replacement due to lack of adherent properties. Efficiency of MSC generation is difficult to establish, and reports on iPSC-MSC or embryonic cell conversion often do not include efficiency data [22,26,47 –49]. However, the flow cytometry results show that the cells that remained attached throughout iPSC-MSC conversion are homogenous and phenotypically similar to CB-MSCs.
Our iPSC-MSCs successfully underwent adipogenesis and osteogenesis after chemical induction, as determined by morphology and staining assays (Fig. 6). Analysis of the expression of adipogenesis-associated genes LPL and PPARγ did not yield significant results in induced iPSC-MSCs or CB-MSCs (Supplementary Fig. S2). We believe this to be a power issue rather than an indication of minimal adipogenic differentiation, as clear morphological changes and lipid vacuole formation are seen in induced iPSC-MSCs and CB-MSCs (Fig. 6 and Supplementary Fig. S1).
Intriguingly, iPSC-MSC response to chemical chondrogenic induction in 3D pellet culture was lower than expected. This may be associated with the low expression of Sox9 in early passage cells; although Sox9 expression increases in late passage cells, the increased passage number may correlate with decreased chondrogenic potential as is seen in MSCs [50]. Other studies have demonstrated the benefit of overexpressing Sox9 to enhance chondrogenesis [43,51,52]. It may be that the use of only one equine iPSC line reflects donor-to-donor variation in the chondrogenic potency of MSCs from adult tissues, a known phenomenon [11,53,54]. iPSC-MSCs generated from other equine iPSC lines may possess chondrogenic potency under the described conditions. Several unsuccessful attempts at direct differentiation of iPSCs to chondrocytes were attempted as well; these included embryoid body formation, pellet culture of iPSCs, membrane culture of iPSCs alone, and membrane culture of iPSCs cocultured with bovine chondrocytes (data not shown). This work exemplifies the caveat that established mouse and human protocols for differentiation cannot simply be recapitulated in the horse. This report is a first step toward development of equine specific protocols for iPS cell fate conversion toward MSCs.
In conclusion, the present study provides an important look at the capacity of horse iPSCs to differentiate into an MSC-like progenitor population. Using a simplified serial plating technique, we demonstrate that MSC-like cells with properties similar to CB-MSCs can be derived from equine iPSCs. Although this technique is reproducible in one equine iPSC line, the general robustness of the protocol should be tested using a larger number of unrelated iPSC lines. Testing several iPSC lines would also help answer whether the lack of chondrogenic potential is a general feature of equine iPSC-MSCs or the iPSC line used in this study.
Footnotes
Acknowledgments
The authors sincerely thank Malgosia Kownacka for cell culture reagent and feeder preparation and David Lee for assistance with the differentiation assays and intellectual input. They also thank Laurence Tessier and Betty-Anne McBey for help with flow cytometry and Carmon Co for technical assistance. This project was partially supported by funds to T.G.K. from the Danish Agency for Technology, Production and Innovation (FTP/274-09-0024), Equine Guelph (EG/2012-12), the Department of Biomedical Sciences, and the Dean's Office at the Ontario Veterinary College. Further components of this work were supported by grants awarded to A.N. from the Ontario Research Fund Global Leadership Round in Genomics and Life Sciences grants (GL2), from the Canadian Stem Cell Network [9/5254 (TR3)], and from the Canadian Institutes of Health Research (CIHR MOP102575). A.N. is a Tier 1 Canada Research Chair in Stem Cells and Regeneration.
This work was presented at the International Society for Stem Cell Research Annual Meeting, Vancouver, BC, 2014.
Author Disclosure Statement
T.G.K. acts in a volunteer capacity as a nonexecutive Director, Scientific Affairs (ex officio) of eQcell therapies Inc., Aurora, Ontario, Canada, a company for which T.G.K.'s research laboratory provides equine stem cell isolation and storage services. T.G.K. holds a minor noncontrolling share in eQcell therapies Inc., S.I.M.L., K.N., H.-K.S., R.A.K., and A.N. declare no competing interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.