
Abstract
Hematopoietic cells represent an attractive starting cell type for induced pluripotent stem (iPS) cells induction, yet the molecular mechanisms in hematopoietic reprogramming are poorly defined. In this study, we showed that long-term hematopoietic stem cells are more amenable for iPS cells induction among several hematopoietic stem and progenitor cell (HSPC) populations, and that this is accompanied by an earlier induction of the transcriptional program that is involved in the promotion of macromolecule metabolism and cell proliferation. Notably, we identified multiple signaling pathways that exhibited distinct expression patterns in HSPCs compared to that of fibroblasts, which is the most commonly used model for probing the somatic reprogramming process. We further experimentally confirmed the differential requirements of the Wnt/β-catenin and transforming growth factor-beta (TGF-β) signaling pathways in these two cell types. These data demonstrate that hematopoietic cells have a cell-type specific transcriptional program and possess unique signaling requirements in the early phase of reprogramming.
Introduction
B
Although mouse embryonic fibroblasts (MEFs) are the most commonly used model system for investigating the molecular mechanism involved in iPS cell induction, multiple somatic cells with distinct tissue identity, including hematopoietic, neural, liver, and stomach cells, have been successfully reprogrammed to pluripotency [3 –5]. Specifically, cells with hematopoietic origin are regarded as an excellent alternative in the generation of iPS cells. While human skin fibroblasts are obtained via an invasive and nonsterile procedure and require 2–3 weeks of expansion before experimentation, hematopoietic cells, especially peripheral blood mononuclear cells, are easily accessible [6]. In addition, skin cells carry more somatic genomic variations due to environmental aggressions, such as ultraviolet irradiation, than cells from inside the body [7].
Understanding the molecular mechanisms involved in reprogramming will address fundamental questions such as cell identity and cell fate decision and improve reprogramming efficiency and quality. Recent studies suggest that reprogramming is a multiphase process [8,9]. The early phase involves inhibition of the original cell identity accompanied by events associated with proliferation, metabolism changes, and cytoskeleton organization. Subsequently, the cells will go through a process of stochastic activation of pluripotency markers and glycolysis (intermediate phase) and activation of the core pluripotency circuitry and silencing of transgenes (late phase) before they reach iPS cell maturation. Despite these insightful findings in MEF reprogramming, much less is known regarding the underlying mechanisms that occur when other cells are employed.
Considering the drastic differences in reprogramming efficiency among various cell types and cells at different differentiation stages [10,11], cell-specific molecular mechanisms may be in operation. In this study, we explored the early events in the reprogramming of hematopoietic stem and progenitor cells (HSPCs). Our results suggested commonalities between fibroblasts and hematopoietic cells and revealed distinct requirements of the Wnt/β-catenin and transforming growth factor-beta (TGF-β) signaling pathways in these two cell types.
Materials and Methods
Animals
Specific pathogen-free mice were housed in the animal facility of the National Institute of Biological Sciences. All studies adhered to procedures that were consistent with the National Institute of Biological Sciences Guide for the care and use of laboratory animals.
Induced pluripotent stem cell induction and all-iPS mice generation
Tet-on-inducible lentiviruses carrying cDNAs of the reprogramming factors were used for iPS induction. All-iPS mice were obtained through tetraploid embryo complementation as previously described [12].
Hematopoietic cell reprogramming
Long-term hematopoietic stem cells (LT-HSCs), short-term hematopoietic stem cells (ST-HSCs), myeloid progenitors (MPs), and lineage negative (Lin−) hematopoietic cells were cultured in iPS cell culture medium supplemented with 50 ng/mL stem cell factor (SCF), 20 ng/mL human thrombopoietin (TPO), 10 ng/mL mouse interleukin (IL)-3, 10 ng/mL mouse IL-6, 10 ng/mL mouse Flt-3 ligand, and penicillin/streptomycin (100×; Merk Millipore) in the presence of feeder cells. All cytokines were purchased from Peprotech, if not mentioned specifically. The Gsk-3β inhibitor (CHIR-99021; Selleck) and Tgf-β R1 kinase inhibitor A-83-01 (616454; Merck) were added to the culture medium the same time as iPS induction. A Leukocyte Alkaline Phosphatase Kit (Sigma) was employed for alkaline phosphatase (AP) staining according to the manufacturer's instructions.
RNA sequencing and expression analysis
The single-cell RNA-seq strategy [13] was employed to capture the transcriptomes of 25 hematopoietic cells. An RNA-seq library was prepared using the protocol of NEBNext Singleplex (#E7350) oligos from Illumina. All reads were aligned to the mouse reference genome mm9 from UCSC using TopHat [14]. The rates of unique maps ranged from 43.20% to 54.28% for 18 samples (Supplementary Table S1; Supplementary Data are available online at
Statistical analysis
Gene functional enrichment analysis was performed using DAVID (
Data access
The deep sequencing RNA data from this study have been deposited in the NCBI database and are accessible through GEO series accession number GSE54462 (
Detailed description could be found in Supplementary Experimental Procedures section.
Results
Reprogramming of HSPCs
To explore the early molecular events in the reprogramming of HSPCs, we employed a secondary reprogramming system in which reprogramming factors can be induced by doxycycline (Dox) in a genetically homogenous cell population [12,17]. Specifically, we obtained primary iPS cell lines by infecting mouse fibroblasts with Dox-controlled, Tet-on-inducible lentiviruses expressing Oct4, Sox2, Klf4, and c-Myc. All-iPS mice were generated by tetraploid embryo complementation using one of these cell lines, LiPS-37 [18]. Therefore, all tissues/cells of these mice carry the expression vector at identical genomic positions and can be induced to express the reprogramming factors by the addition of Dox (Fig. 1A).
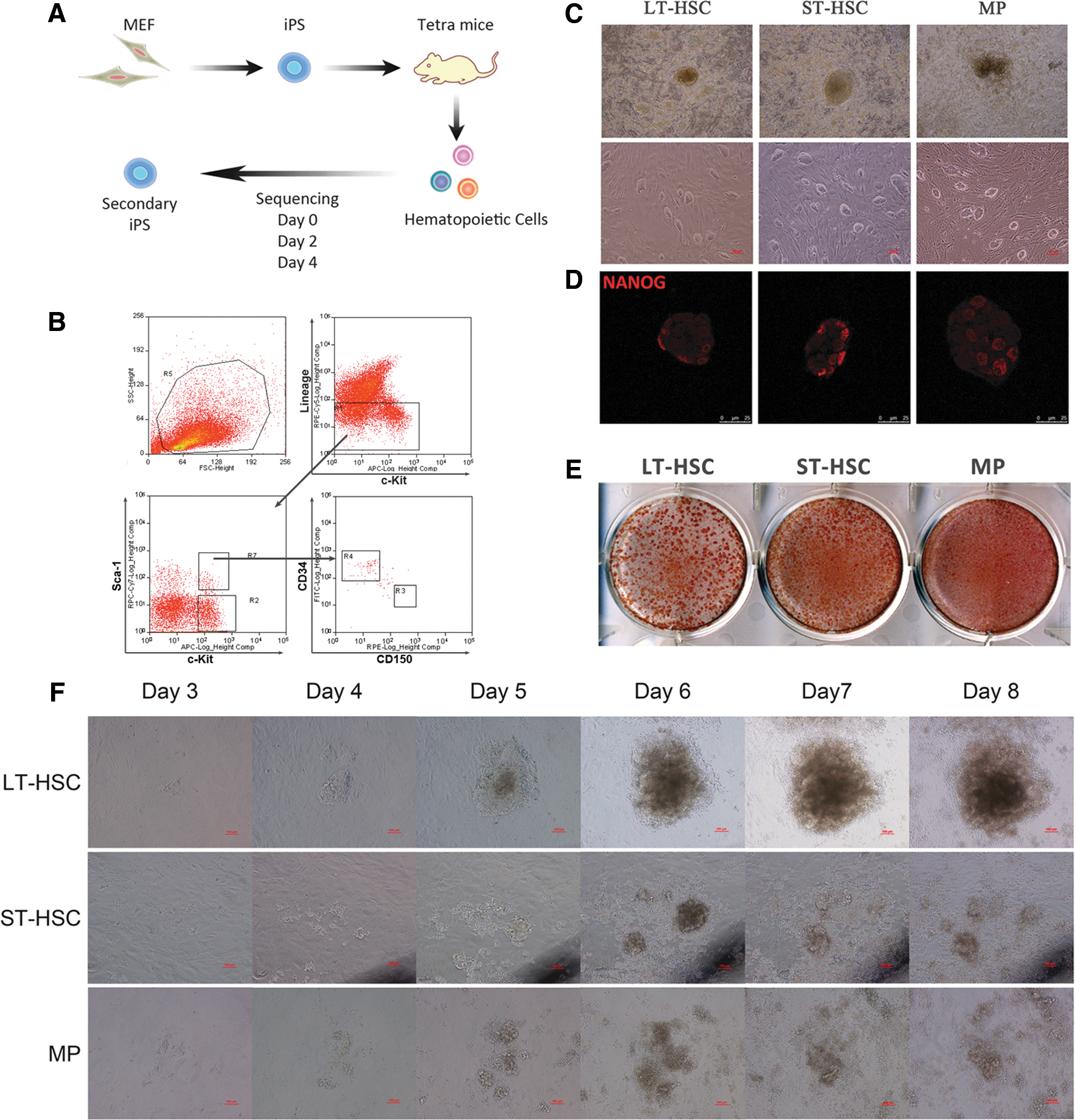
Reprogramming of hematopoietic stem and progenitor cells (HSPCs).
To determine whether secondary iPS cells can be obtained from HSPCs, we isolated several hematopoietic stem cell and progenitor populations based on the surface markers LSK, CD34 [19,20], and assisted by CD150 [21] (Fig. 1B). While LT-HSCs (Lin−Sca-1+c-Kit+CD34−CD150+) reside at the top of the hematopoietic hierarchy and have long-term populating capability, ST-HSCs (Lin−Sca-1+c-Kit+CD34+CD150−) and MPs (Lin−Sca-1−c-Kit+) are more limited in their potential to sustain hematopoiesis.
All three populations were cultured in iPS cell culture medium supplemented with Dox and the same cytokine cocktail consisting of SCF, TPO, IL-3, IL-6, and Flt-3 ligand for 2 weeks. After picking and expanding single colonies, all three HSPC populations gave rise to iPS colonies and stable iPS lines (Fig. 1C), and immunofluorescence staining showed the Nanog expression in these iPS cells (Fig. 1D).
Overall, more than 100 AP+ colonies could be derived from 2,000 starting cells (Fig. 1E). Interestingly, the iPS colonies that were derived from LT-HSCs were obviously larger than those from ST-HSCs and MPs (Fig. 1E), suggesting that these colonies might have formed at an earlier time. In fact, LT-HSCs became adherent to the plate, congregated the earliest, and gave rise to compact colonies as early as day 4 after Dox addition, whereas colony formation of ST-HSCs and MPs required an additional 1–2 days to take place (Fig. 1F and Supplementary Fig. S2). These results suggest that LT-HSCs reprogram faster than ST-HSCs and MPs.
Biological processes involved in early reprogramming of different hematopoietic cells
Based on the observed faster reprogramming in LT-HSCs, we hypothesized that LT-HSCs may have an earlier induction of favorable molecular events that lead to pluripotency. Therefore, we performed single cell RNA-seq [13] to capture the transcriptome from small number of hematopoietic cells.
We identified the transcriptional changes 2 and 4 days after Dox induction on each of the HSPC populations (Supplementary Fig. S3 and Supplementary Experimental Procedures section). Only about 11% and 11.5% of differentially expressed genes were common in three hematopoietic cells after 2 and 4 days induction respectively, while about 60% of the differentially expressed genes had cell specificity (Fig. 2A).
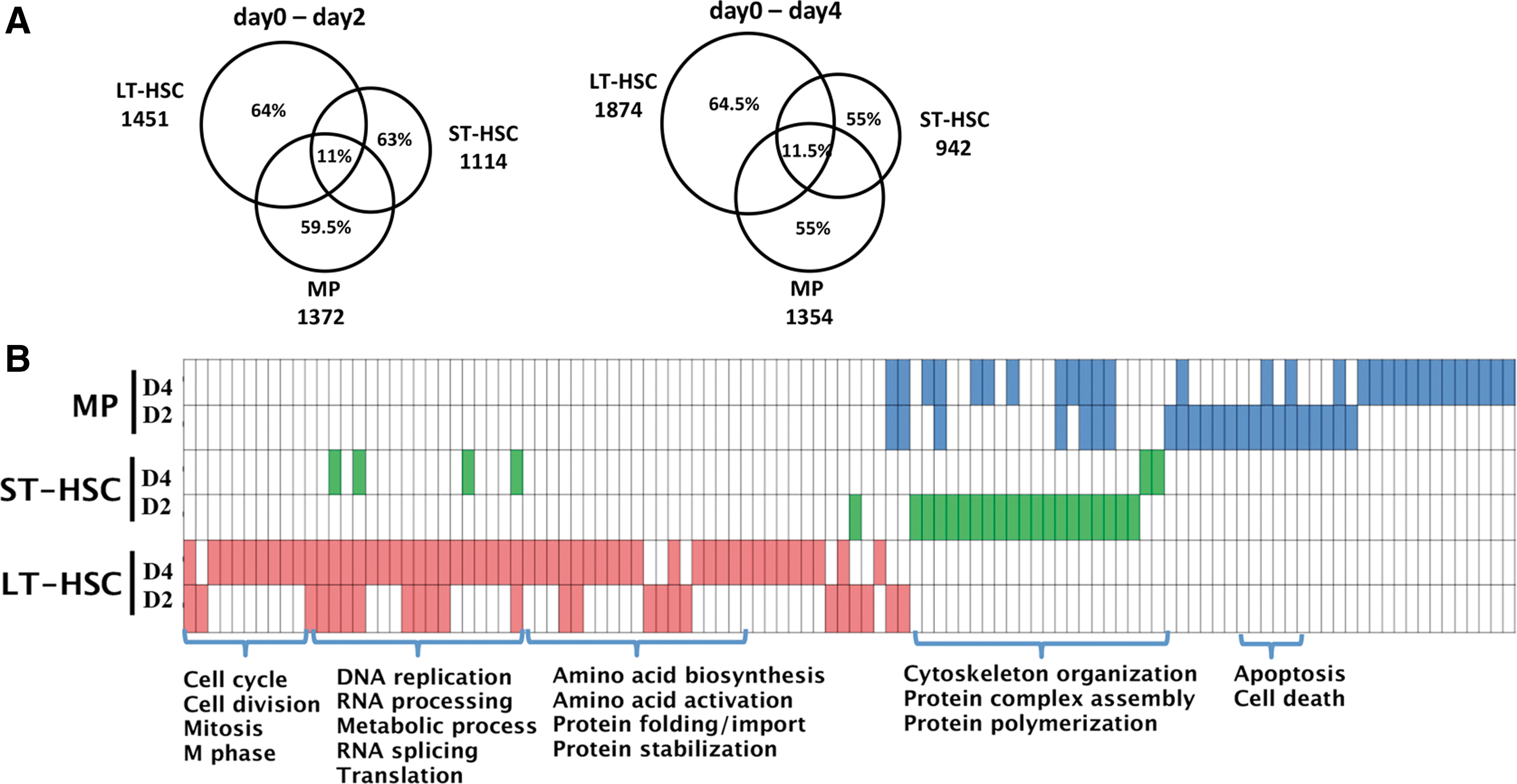
Expression profile analysis of early reprogramming of different HSPCs.
Differentially upregulated genes in Dox-induced LT-HSCs at days 2 and 4 were enriched for cell cycle, RNA/DNA processing, mitochondria, metabolism and biosynthetic processes, which are known molecular pathways that facilitate reprogramming (Fig. 2B and Supplementary Fig. S4 and Supplementary Experimental Procedures section). These results corroborate with earlier findings that molecular events related to proliferation are implemented at the earliest stages of reprogramming [9]. In contrast, these proliferation- and metabolism-related biological processes were more limited in Dox-treated ST-HSCs and MPs (Fig. 2B and Supplementary Fig. S4). ST-HSCs exhibited increased expression of genes related to filament depolymerization and protein complex assembly, while MPs exhibited significant induction of gene expression in organelle organization, cellular development, cell death, and apoptosis (Fig. 2B and Supplementary Fig. S5), which is a reported barrier of reprogramming [22].
Together, these data suggest that though LT-HSCs were the most quiescent cells among these cell types but responded quickly to the induction in early stage, and all three HSPC populations exhibited various degrees of metabolic reorganization in setting the stage for reprogramming.
Silencing of hematopoietic tissue-specific genes, but not the mesenchymal-to-epithelial transition, was an early feature of HSPC reprogramming
The mesenchymal-to-epithelial transition (MET) was identified as a hallmark in the initiation phase of MEF reprogramming and is characterized by downregulation of mesenchymal genes and induction of epithelial-associated genes [23,24]. To explore whether MET also occur as early event in HSPC reprogramming, we first examined the levels of expression of the key mesenchymal regulators and fibroblast markers Snai1, Slug, Zeb1, Zeb2, Cdh2, and Thy1. While high-level expression of all six genes were significantly downregulated as early as day 2 during fibroblast reprogramming, we could barely detect expression of the same set of genes in uninduced, as well as 2- and 4-day Dox-treated HSPCs (Fig. 3A). Considering the mesenchymal property of fibroblasts, we reasoned that this MET phenomenon emphasizes the suppression of the expression of genes specific to the starting cell type of reprogramming.
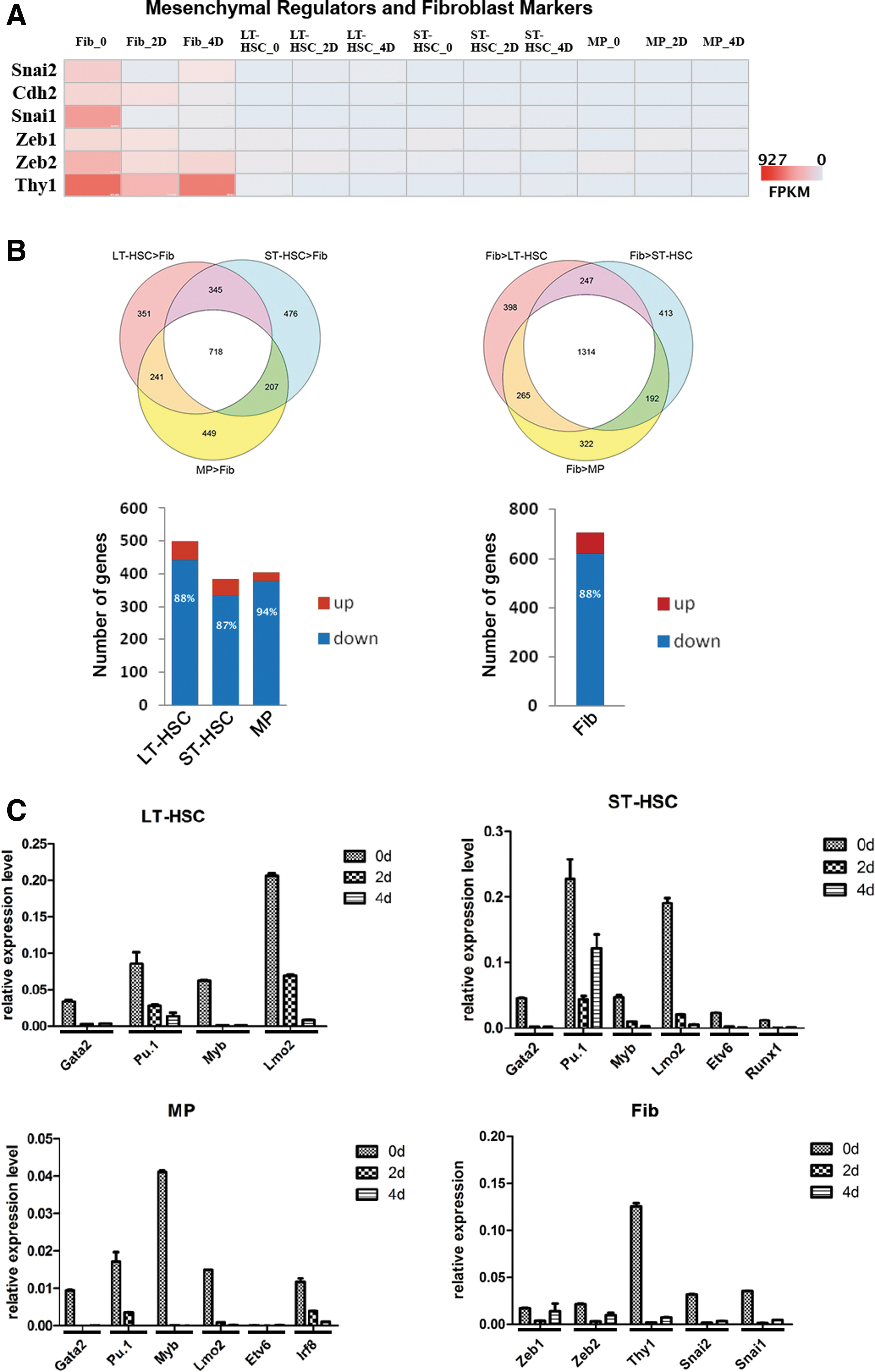
Silencing of somatic cell-specific genes in reprogramming.
Therefore, we tested whether the expression of hematopoietic-specific genes were specifically downregulated in the process of HSPC reprogramming. To define hematopoietic/HSPC-specific genes, we compared the expression profile of each of the HSPC populations with that of fibroblast cells obtained from the skin of all-iPS mice in the absence of Dox treatment (day 0) (Supplementary Fig. S3 and Supplementary Experimental Procedures section).
Compared to fibroblasts, 718 highly expressed genes were shared by the three HSPC cell populations (P<0.001 and fold change (FC) >3, Fig. 3B, upper left). Approximately 90% of the differentially expressed, HSPC-specific genes (see Supplementary Experimental Procedures section) were downregulated after Dox treatment (Fig. 3B, lower left and Supplementary Fig. S6). These genes were enriched in the immune response, leukocyte activation, and leukocyte differentiation (Supplementary Table S2). Consistent with the RNA-seq data, the expression of hematopoietic key regulators, such as Gata2, PU.1, Myb, and Lmo2 were markedly decreased after the induction of reprogramming, as detected by real-time PCR (Fig. 3C).
Compared to HSPCs, 1,314 genes were highly expressed in fibroblasts (fibroblast-specific genes, Fig. 3B, upper right), and 88% of the differentially expressed, fibroblast-specific genes after Dox-treatment were downregulated and mostly enriched in cell adhesion, biological adhesion, and extracellular matrix, among others (Fig. 3B, lower right and Supplementary Fig. S6 and Supplementary Table S2). Together, these results demonstrated that downregulation of tissue-specific genes was a common event in the initial phase of reprogramming of fibroblasts and hematopoietic cells.
Differential requirements of Wnt/β-catenin and TGF-β signaling in reprogramming of HSPCs and fibroblasts
The identification of critical signaling events in reprogramming provides biological insights into this complex process and offers attractive targets when using small molecules to improve iPS induction. To search for molecular pathways that are specifically involved in the early phase of iPS induction of HSPCs, we identified differentially expressed genes after 2 days of Dox treatment in HSPCs and fibroblasts, respectively (Fig. 4A and Supplementary Fig. S3 and Supplementary Experimental Procedures section).
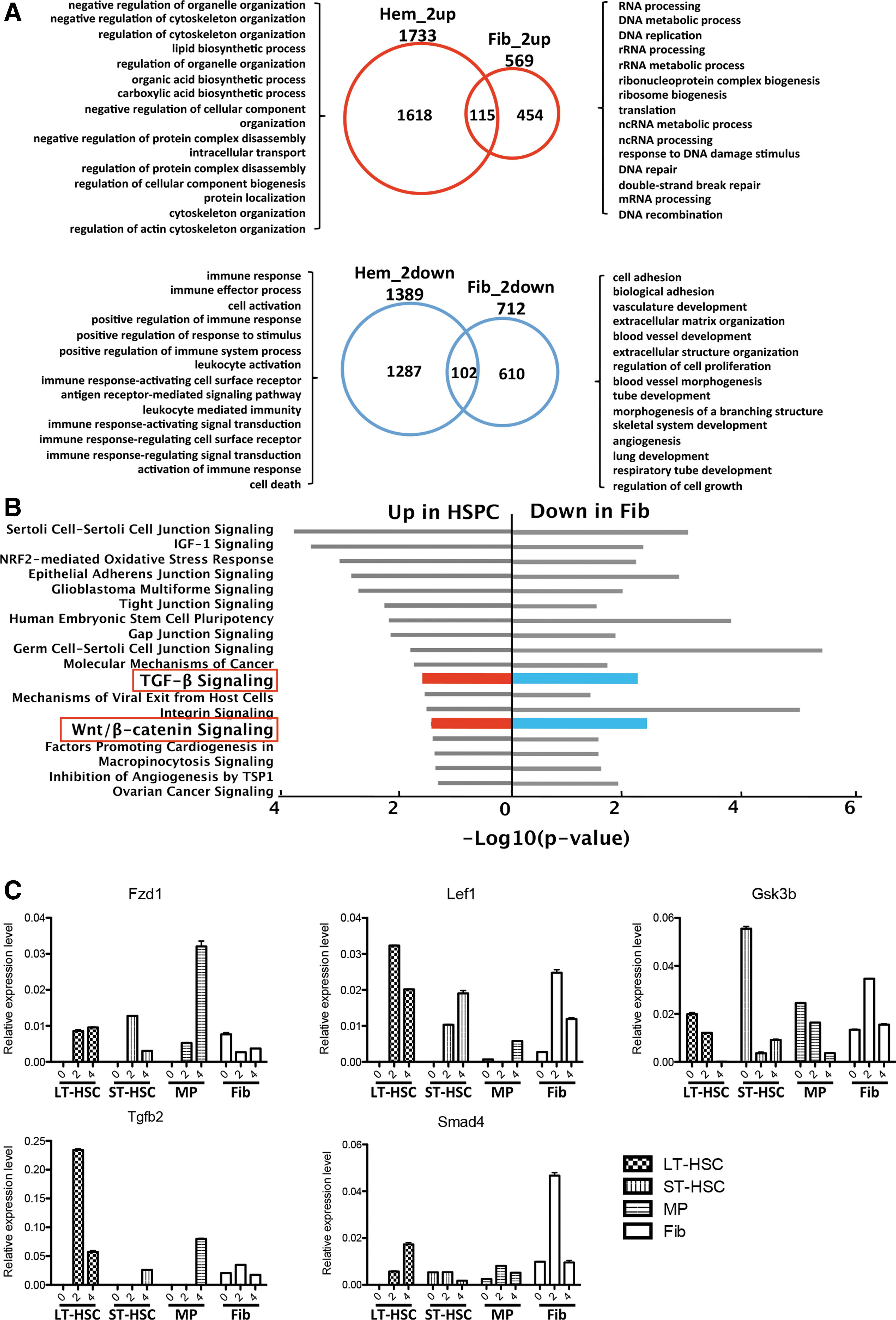
Wnt/β-catenin and transforming growth factor-beta (TGF-β) signaling profile in HSPCs and fibroblasts reprogramming.
Using signaling pathway enrichment analysis (IPA), we found several pathways that were upregulated in HSPCs but downregulated in fibroblasts (Fig. 4B and Supplementary Table S3). Among these pathways, Wnt/β-catenin and TGF-β have been previously shown to be important signaling pathways in stem cells and reprogramming. The expression changes during hematopoietic cells reprogramming were confirmed by real-time PCR of key genes, including Fzd1, Lef1, and Gsk3b in the Wnt pathway and Tgfb2 and Smad4 in the TGF-β pathway (Fig. 4C).
We further performed functional tests to determine the effects of these two pathways in the reprogramming of HSPCs (Lin− hematopoietic cells) and fibroblasts (see Supplementary Experimental Procedures section). Consistent with previous findings [25,26], the inhibition of TGF-β signaling via A-83-01, a small molecule that specifically inhibits TGF-β type I receptor ALKs (activin receptor-like kinases), remarkably promoted reprogramming in fibroblasts (FC=2.73, P<0.05) (Fig. 5A, left). This result is consistent with the notion that TGF-β signaling could induce the mesenchymal phenotype [27], which is a barrier to MEF reprogramming. In sharp contrast, A-83-01 minimally affected iPS induction in HSPCs (FC=0.71, P<0.05) (Fig. 5B, right), demonstrating the differential requirements of TGF-β signaling in hematopoietic cells and fibroblasts.
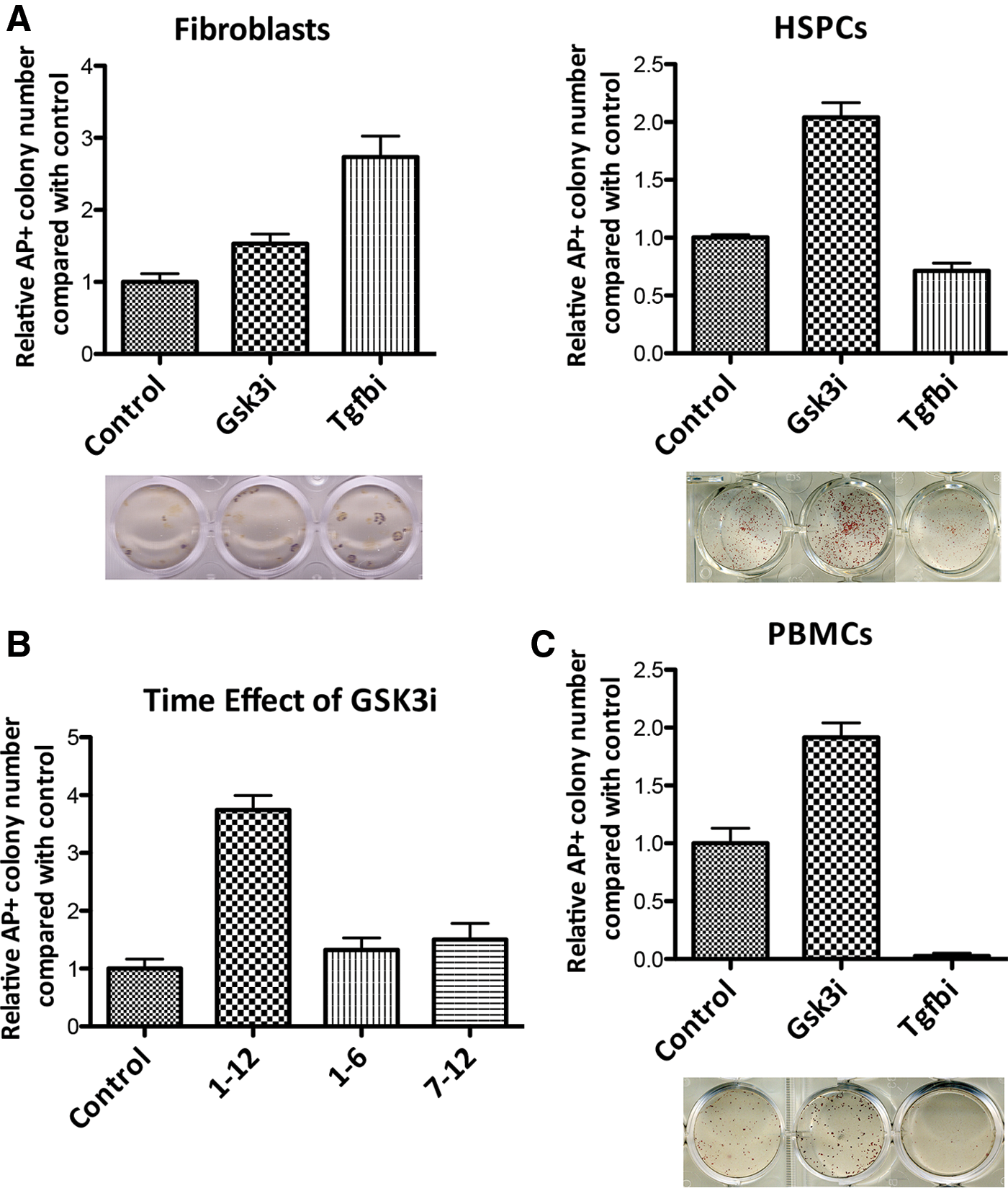
Wnt/β-catenin and TGF-β signaling profile in HSPCs and fibroblasts reprogramming.
To explore whether the different effects of TGF-β inhibitor on reprogramming are depending on different starting contexts, we investigated the initial expression levels of TGF-β pathway genes in HSPCs and fibroblasts with RNA-seq data. The expression level of TGF-β pathway genes in starting cells of fibroblasts was much higher than that of HSPCs (P<0.01, Supplementary Fig. S7). This expression difference tended to be eliminated after reprogramming (P=0.6, Supplementary Fig. S7).
Furthermore, we found that Alk4 (Acvr1b), one of the targets of A-83-01, showed significantly lower expression in hematopoietic cells than in fibroblasts (Supplementary Fig. S8). Thus, differential expression of Alk4 may render hematopoietic cells unresponsive to the inhibitor. Interestingly, c-Myc, which is a key gene known to significantly impact TGF-β signaling, was expressed at a higher level in hematopoietic cells than in fibroblasts, as shown by our own RNA-seq data (Supplementary Fig. S9) and an independent public dataset (GSE36290) [28]. This result is supportive of earlier findings in which a TGF-β inhibitor much more significantly promoted reprogramming in the absence than in the presence of c-Myc [25,26].
Furthermore, the enhancement of the Wnt pathway by inhibition of Gsk3β (CHIR-99021), a strong negative regulator of the Wnt pathway, significantly increased the efficiency of the AP+ iPS colonies generated from HSPCs (FC=2.04, P<0.05) (Fig. 5A, right). Addition of the Gsk3β inhibitor in the early (1–6 day) or late (7–12 day) phase of the reprogramming had only moderate impact (Fig. 5B), suggesting that Gsk3β inhibition is required during the entire course of reprogramming to promote hematopoietic cells reprogramming. Consistent with the finding in HSPCs, enhanced Wnt signaling via inhibition of Gsk3β significantly facilitated reprogramming of monocytes isolated from peripheral blood, a convenient source for clinical applications of iPS technology (FC=1.91, P<0.05) (Fig. 5C). However, treatment of fibroblasts with the Gsk3β inhibitor had a moderate effect on iPS colony formation (FC=1.53, P<0.05) (Fig. 5A, left). These results identified Wnt as a strong stimulator in the early phase of reprogramming for HSPCs and in mature monocytes.
Discussion
The induction of iPS cells is a low-efficiency, time-consuming process, and its molecular mechanism is not well understood. While progress has been made regarding molecular insights into MEF reprogramming, less is known about the underlying mechanisms when other somatic tissues, including HSPCs, are reprogrammed to pluripotency. Our study is the first to examine the early events in HSPC reprogramming. Similar to the downregulation of mesenchymal-associated genes in fibroblasts, the repression of hematopoiesis-associated genes was featured in hematopoietic reprogramming. Significantly increased gene expression that was associated with cell cycle, RNA processing, mitochondria, and metabolism was observed when HSPCs with different self-renewal potentials were induced to form iPS cells.
Our study also provided evidence that LT-HSCs, when compared with several HSPC populations, were more amenable to reprogramming with higher iPS cell induction velocity, and this was accompanied by an earlier transcriptional induction of metabolic and proliferative genes. This result is consistent with the recent finding that nonstochastic reprogramming was observed in hematopoietic cells [29].
An earlier study also suggested that HSPCs are easier to reprogram than differentiated cells and that hematopoietic progenitor cells showed higher reprogramming efficiency than HSCs [11]. It is worth noting that the iPS induction efficiency is traditionally presented simply as the ratio of the number of iPS colonies to the number of starting cells. For adherent cells, the progenies of single-starting cells will give rise to a single colony. However, for floating cells, all progenies of single-starting cells have the possibility of giving rise to independent colonies before they become adherent to the plate. Therefore, this counting method may be largely influenced by cell proliferation and the time that is required for the cells to become adherent and iPS cells. This might also explain why hematopoietic progenitor cells, which are more active in proliferation, had a slower reprogramming pace, yet showed higher reprogramming efficiency than HSCs [11].
The Wnt/β-catenin pathway has been shown to participate in somatic reprogramming. In the absence of c-Myc [22] or Sox2 [30], Wnt signaling can facilitate the generation of iPS cells. However, in the presence of all four factors, Wnt stimulation had modest to no effect on the reprogramming of MEFs to pluripotency [22,31]. These reports are consistent with our finding that showed that the activation of the Wnt pathway by a Gsk3β inhibitor had only a modest impact on the reprogramming of fibroblasts (Fig. 5A, left). In contrast, inhibition of Gsk3β significantly promoted the induction of iPS cells from hematopoietic cells (Fig. 5A, right).
Interestingly, a recent study demonstrated a biphasic role of Wnt signaling, in which the key Wnt signaling genes Tcf3 and Tcf4 played a different role in the early and late stages of somatic MEF reprogramming [31]. In our gene expression analysis, we also noticed a stronger transcriptional induction of these genes during early HSPC reprogramming compared with that of fibroblasts (data not shown). These findings warrant further investigation on how the cell type-specific expression of key genes influences the distinct requirement of Wnt signaling.
TGF-β signaling can actively maintain the mesenchymal phenotype in fibroblasts and is a well-known inducer of the EMT, the reverse process of MET. Consistently, TGF-β inhibition enhanced reprogramming of MEFs by inducing the MET [24,32]. In support of these earlier findings, our data showed that the TGF-β inhibitor A-83-01 increased the efficiency of iPS colony formation from fibroblasts. However, this inhibitor showed no positive effects in HSPC reprogramming. This difference may result from different starting contexts, as we showed, HSPCs exhibited a completely opposite expression pattern of the key genes that are known to positively affect MEF reprogramming, including low-expressed mesenchymal genes and key TGF-β signaling genes such as Alk4 and a higher level of expression of c-Myc. These data suggest that the cell type-specific context, which is defined by the unique expression pattern of key genes in the starting cells, may have a strong impact on the signaling requirement in the early phase of reprogramming.
Footnotes
Acknowledgments
We thank Professor Rudolf Jaenisch for generously supplying the lentivirus vectors containing the cDNAs of the four transcription factors, Zhenyu Ju and Tao Cheng for help with the hematopoietic cell sorting strategy, and Fuchou Tang for the introduction of single-cell transcriptome analysis. This project was supported by the National Natural Science Foundation of China (grants 31371512, 81322029, and 81300438) and the “Strategic Priority Research Program” of the Chinese Academy of Sciences XDA01010305.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.