
Abstract
Tissue engineering is a popular topic in peripheral nerve repair. Combining a nerve conduit with supporting adipose-derived cells could offer an opportunity to prevent time-consuming Schwann cell culture or the use of an autograft with its donor site morbidity and eventually improve clinical outcome. The aim of this study was to provide a broad overview over promising transplantable cells under equal experimental conditions over a long-term period. A 10-mm gap in the sciatic nerve of female Sprague-Dawley rats (7 groups of 7 animals, 8 weeks old) was bridged through a biodegradable fibrin conduit filled with rat adipose-derived stem cells (rASCs), differentiated rASCs (drASCs), human (h)ASCs from the superficial and deep abdominal layer, human stromal vascular fraction (SVF), or rat Schwann cells, respectively. As a control, we resutured a nerve segment as an autograft. Long-term evaluation was carried out after 12 weeks comprising walking track, morphometric, and MRI analyses. The sciatic functional index was calculated. Cross sections of the nerve, proximal, distal, and in between the two sutures, were analyzed for re-/myelination and axon count. Gastrocnemius muscle weights were compared. MRI proved biodegradation of the conduit. Differentiated rat ASCs performed significantly better than undifferentiated rASCs with less muscle atrophy and superior functional results. Superficial hASCs supported regeneration better than deep hASCs, in line with published in vitro data. The best regeneration potential was achieved by the drASC group when compared with other adipose tissue-derived cells. Considering the ease of procedure from harvesting to transplanting, we conclude that comparison of promising cells for nerve regeneration revealed that particularly differentiated ASCs could be a clinically translatable route toward new methods to enhance peripheral nerve repair.
Introduction
Background
P
Primary repair is limited to injuries in which there is little or no defect and in which, thus, a tension-free coaptation is possible. If a peripheral nerve injury is associated with a loss of substance, autologous nerve grafts are usually employed for bridging the gap [2]. Although seen as gold standard, there are remarkable disadvantages that go along with this treatment, that is, donor site loss of function, formation of potential painful neuromas, structural differences between donor and recipient grafts, and the limited graft material [3,4].
Next generation of nerve repair
Alternatively, nerve entubulation is often chosen as the method of treatment to act as a physical guide for regenerating nerve sprouts. Several natural and artificial materials, such as collagen, silicone, fibrin, or vein grafts, are used as such conduits [5 –8]. Ideally, the degradation kinetic of the conduit is to match the nerve regeneration rate. Our group introduced a bioresorbable nerve conduit constructed from fibrin glue, which shows good tissue acceptance without altering the tissue environment around the implantation site [9].
Yet, to bridge a gap between the proximal and distal nerve stump, not only the sheath is of importance but also different types of cells and stem cells, respectively, are being explored in terms of their support in nerve regeneration.
The ideal transplantable cell
The ideal transplantable cell should be easily accessible, proliferate in culture, and successfully integrate into host tissue with immunological tolerance [10]. With the aim of finding this ideal cell, numerous trials have been carried out in the past, introducing different types of cells and comparing one with another.
Schwann cells play an inalienable role in supporting axonal regrowth and migration following peripheral nervous system (PNS) injuries. They support neuronal survival, release bioactive factors, and affect blood vessel formation [11]. In lesioned nerves, Schwann cells form longitudinally oriented strands (bands of Büngner) that serve as a guiding rail for regrowing axons [12].
Unfortunately, next to the autografts, the administration of Schwann cells is also limited: To obtain a sufficient number of viable Schwann cells, 3–10 weeks of ex vivo expansion and purification of the Schwann cells are often needed [13]. The requirement for nerve donor material evokes additional morbidity and the time required to culture and expand the cells would delay treatment [14]. The enrichment with allogeneic cells could shorten this delicate time lag. However, immunosuppression is desirable in this case so as to avoid rejection of the allogeneic donor Schwann cells implanted [15].
In the past, we showed how differentiated ASCs could enhance neurite outgrowth in vitro [14]. In the following, the ability of these differentiated ASCs to enhance nerve regeneration in vivo could be confirmed [5,16].
We decided to focus on adipose tissue-derived cells due to the above-mentioned promising results combined with further advantages when compared with bone marrow-derived cells: The amount of ASCs is said to be approximately 500-fold greater than that found in marrow [17]. Moreover, ASCs have high proliferation rates, can be harvested through minimally invasive procedures, and are multipotent [18,19]. We previously showed that ASCs might be a clinically translatable route toward new methods to enhance peripheral nerve repair [5]. The utilization of human adult stem cells is an important step toward this goal. Here again, human bone marrow-derived stem cells (hBMSCs) were compared with human adipose-derived stem cells (hASCs): In a nude mice model of hind limb ischemia, hASCs exhibited greater proangiogenic activity than hBMSCs [20,21]. We were the first to show that undifferentiated hASCs in vitro also express a range of neurotrophic factors [22]. In particular, human adipocyte precursor cells from the subcutaneous adipose tissue region appear to be attractive since they are not only easily accessible but also proliferate faster when compared with those from the omental region [23]. What is more, the neurotrophic activity of hASCs isolated from deep and superficial layers of abdominal fat tissue (separated by the abdominal fascia) was analyzed separately to find out whether their innate capacities of nerve regeneration therapies might differ. Cells from the superficial layer proliferate significantly faster than those from the deep layer and promote neurite process outgrowth in vitro [22]. Furthermore, ASCs isolated from the superficial layer are relatively more resistant to apoptosis than those from the deep layer [24]. Santiago et al. found that the transplantation of hASCs following sciatic nerve transection promoted the formation of a more robust nerve and produced a decrease in muscle atrophy [25].
In addition to ASCs, the SVF also contains blood-derived cells, in particular erythrocytes and leukocytes [26]. Freshly isolated SVF can effectively induce new vessel formation through the dynamic reassembly of blood endothelial cells and could thus be applied to achieve therapeutic neovascularization [27].
Aim of this study
A comparison of the published results might be limited due to differing aspects, for example, cell isolation and differentiation protocols, number of cells introduced into the conduit, type of conduit, or examined time interval. To enable this comparison, the present work provides a broad overview over promising transplantable cells in peripheral nerve repair according to the current state of research, including rat adipose-derived stem cells (rASCs), differentiated rASCs (drASCs), rat Schwann cells, human (h-)ASCs from the superficial (hASCsup) and deep (hASCdeep) abdominal layer, as well as the human stromal vascular fraction (SVF), and, as a control, an autograft to bridge a 10-mm gap in the sciatic nerve of rats. Not only the diverse cell types are comprised, but they have also been examined long term (12 weeks) in a multimodal manner containing functional and morphometric, as well as MRI analyses.
Materials and Methods
Study design
The sciatic nerve injury model was used, creating a 10-mm gap in the left nerve of female Sprague-Dawley rats (7 groups of 7 animals, 8 weeks old) that was bridged through a biodegradable fibrin conduit by the microsurgical suture technique. 1 Mio of each of the following cells was introduced into the conduits: rASCs, differentiated rASCs (drASCs), rat Schwann cells, human (h-)ASCs from the superficial and deep abdominal layer, as well as human SVF. As a control, we resutured a 10-mm cut nerve segment backward as an autograft.
After 12 weeks, functional and histomorphometric analyses were performed. Gastrocnemius muscles were weighed. Furthermore, MRI evaluation of nerve growth, muscle atrophy, and degradation of the conduit was carried out.
Harvest and culture of the cells
For ASC isolation and differentiation of rASCs, established protocols from our group were used [14,16,28,29].
From the superficial or deep abdominal layer, hASCs were isolated from patients undergoing either dermolipectomy or liposuction for reconstructive or cosmetic reasons. All patients had signed an informed consent form. To isolate ASCs from rats, subcutaneous adipose tissue from the neck of adult 8-week-old female Sprague-Dawley rats was excised. The adipose tissue was cut into fine pieces and digested in 0.1% collagenase (Sigma; C1639) for 1 h at 37°C on the shaker. The enzymatic digestion reaction was stopped by the addition of Dulbecco's modified Eagle's medium: nutrient mixture F-12 (DMEM/F-12; Gibco, 31331-093) supplemented with 10% FBS. The cells were centrifuged, resuspended in PBS (Invitrogen; AM 9625), followed by filtration through a 70-μm cell strainer (BD Biosciences), before being centrifuged again. The cell pellet was resuspended in growth medium (DMEM/F-12 supplemented with 10% FBS and 1% penicillin/streptomycin). The culture dishes were transferred to an incubator at 37°C with 5% CO2. The medium was changed twice a week, and all of the experiments were carried out using cells that were between passages 2 and 4.
For the transdifferentiation of rASCs into drASCs, the growth medium was removed from subconfluent rASC cultures at passage 2 and replaced with a medium supplemented with 1 mM β-mercaptoethanol (Sigma-Aldrich; M7522) for 24 h. Cells were then washed and fresh medium supplemented with 35 ng/mL all-trans-retinoic acid (Sigma; R2625) was added. A further 72 h later, cells were washed and the medium replaced with differentiation medium; cell growth medium supplemented with 5 ng/mL platelet-derived growth factor (PDGF; PeproTech Ltd., 100-157 13A), 10 ng/mL basic fibroblast growth factor (bFGF; PeproTech Ltd., 100-18B), 14 μM forskolin (Sigma; F6886), and 252 ng/mL neuregulin-1 (R&D Systems; 377-HB-50). Cells were incubated for 2 weeks under these conditions with fresh medium added approximately every 72 h.
For the isolation of human SVF, floating adipocytes and supernatant were removed after centrifugation from the SVF pellet, washed, and resuspended in sterilized PBS. The single-cell suspension was filtered through a 70-μm cell strainer. The total cell number was assessed by crystal violet stain (Sigma-Aldrich Chemie GmbH) and the nucleated cells were counted in a Neubauer chamber (Marienfeld).
For the isolation, expansion, and characterization of rat Schwann cells in vitro, rat Schwann cells were isolated as described previously by our group [16]. To harvest Schwann cells, sciatic nerves from adult female Sprague-Dawley rats were exposed, removed, and kept in DMEM plus Glutamax containing 1% penicillin–streptomycin (DMEM; Invitrogen, 21063029). Nerves were then dissected in trunks, desheathed, and finally cut in 1-mm segments under a microscope. The segments were then plated in a Petri dish with growth medium supplemented by 14 μM forskolin and 40 ng/mL neuregulin-1 (Schwann cell growth medium). Segments were incubated for 2 weeks at 37°C with 5% CO2 and fresh medium was added approximately every 72 h. After 2 weeks, the medium was aspirated and 0.125% collagenase type IV (Invitrogen; 17104-019) and 117 U/mg dispase (Invitrogen; 17105-041) were added to the Petri dish. After 24 h of incubation, the cell suspension was filtered through a 70-μm cell strainer and centrifuged at 900 rpm for 5 min to obtain the cell pellet. Finally, the cell pellet was resuspended in Schwann cell growth medium, seeded into a six-well Primaria plate (BD Bioscience; 353803), and incubated under the same conditions. The following day, the medium was replaced and cells were left to proliferate. At confluence, the Schwann cells were purified by an antibody–complement method to eradicate remaining fibroblasts by initially resuspending the Schwann cells in mouse anti-rat Thy 1.1 antibody (1:500; Serotec). Rabbit complement was then added to the cells, followed by centrifugation at 600 g for 5 min. The supernatant was aspirated and the cell pellet suspended in Schwann cell growth medium. Cells were then plated and incubated in 5% CO2.
Cell characterization
The multipotent nature of the ASCs was tested by ensuring their ability to differentiate along various lineages, as previously described [14].
Fluorescence-activated cell sorting (FACS) and immunocytochemistry using stem cell or lineage-specific markers, respectively, for each cell type were carried out: Cytofluorimetry analysis of SVF/ASCs for standard markers [CD105, CD90, and CD73 (mesenchymal markers), CD31 and CD34 (endothelial markers), and the pan-hematopoietic marker, CD45] was performed as described previously [28,30]. The suspensions were incubated for 30 min at 4°C with fluorochrome-conjugated antibodies against the indicated proteins or an isotype control. The antibodies used were obtained from Becton, Dickinson and Company (
Cell suspension
Cells were counted as 1 Mio pellets suspended in a fibrinogen/culture medium solution (1:10) since former in vitro studies of our group had shown significantly better adhesion to and distribution throughout the conduit when cells were seeded in fibrin compared with delivery in growth medium alone [31].
Fibrin conduit
The fibrin conduit was prepared from a two-compound fibrin glue (Tisseel® Kit VH 1.0; Baxter SA). Tisseel contains fibrinogen, 70–110 mg mL−1; plasma fibronectin, 2–9 mg mL−1; factor XIII, 10–50 U mL−1; plasminogen, 40–120 μg mL−1; aprotinin solution 3,000 KIU mL−1; thrombin 4 IUmL−1; and calcium chloride, 40 mM. Fibrin glue was dispensed in a mold prepared from silicone around a stainless steel core and pressed into shape for 5 min. This allows the generation of uniform conduits measuring 14 mm in length, with a 2-mm lumen and 1 mm wall thickness designed to bridge a 10-mm gap in the sciatic nerve of 8-week-old Sprague-Dawley rats.
Animals
All experiments were performed on adult female Sprague-Dawley rats (Harlan). All studies were approved by the local veterinary physician (No. 2455). All surgical procedures were performed under general anesthesia with isoflurane 3%. Routinely, buprenorphine (Temgesic®, 2 mL Amp, 0.3 mg/mL, 0.1–0.2 mg/kg body weight) was administered before surgery. Isolation of cells from rats was performed following euthanasia by CO2 according to the local veterinary commission.
Surgical procedure
A sciatic nerve lesion creating a 10-mm gap was used. The left sciatic nerve was approached dorsally using a gently spreading technique of the gluteus muscle. Once identified, the nerve was divided with microsurgical scissors at a distance of 10 mm from its emergence of the sacral notch. After resection, the nerve was replaced by the tissue-engineered fibrin conduit or autograft, respectively. Surgical implantation of the structures was undertaken with an operating microscope (Zeiss), and the nerve ends were fixed to the conduit/graft by a single epineural suture (9/0 Nylon, Ethicon): Proximal and distal nerve stumps were inserted 2 mm into the nerve conduits, thus leaving a 10-mm gap. Muscles and fascia layers were closed with single resorbable stitches (5/0 Vicryl, Ethicon), and the skin was closed by using a continuously running suture (5/0 Prolen, Ethicon).
Functional evaluation
The sciatic functional index (SFI) was measured with a walking track analysis 12 weeks postoperatively using a functional evaluation tool, the CatWalk, for automated quantitative gait analysis (Noldus Information Technology). Three repeated measurements under constant conditions were conducted in every animal to capture the steadiness of gait. Recording the animals' footprints, followed by analysis of their walking pattern, and calculation of the SFI are well-established and commonly used methods for the assessment of motor nerve recovery after sciatic nerve injury [32,33].
Measurements included print length on both the experimental and the normal sides (EPL, NPL), toe spread between the first and fifth digits on both sides (ETS, NTS), and the distance between the middle of the second and fourth toes on both sides (EIT, NIT). The formula used to calculate SFI was as follows:
SFI=−38.8(EPL–NPL)/NPL+109.5 (ETS-NTS)/NTS+13.3 (EIT–NIT)/NIT−8.8
An SFI of 0 is normal. An SFI of −100 indicates total impairment, such as would result from a complete transection of the sciatic nerve [34,35].
MRI scanning
After euthanasia with CO2 12 weeks postoperatively, animals were examined on a 3T human whole body scanner (Magnetom Verio, Siemens) using an 8-channel wrist coil. After scout images in three directions had been taken, we performed imaging perpendicular to the calves and to the thighs using T1-weighted (TE 25 ms, TR 586 ms, 0.2×0.3 mm in-plane resolution, 3 mm slice thickness) and T2 STIR (short-T inversion recovery) sequences (TE 25 ms, TR 4000 ms, TI 220 ms, 0.3×0.3 mm in-plane resolution, 3 mm slice thickness), as well as sagittal images of the thigh using T2-weighted (TE 71 ms, TR 4250 ms, 0.2×0.2 mm, 1 mm slice thickness) and T2 STIR sequences (TE 25 ms, TR 4000 ms, TI 220 ms, 0.3×0.3 mm in-plane resolution, 1 mm slice thickness).
Image evaluation
Images obtained 12 weeks after surgery were evaluated by a radiologist with 5 years of postresidency experience in neuromuscular imaging using open-source imaging software (OsiriX, Pixmeo).
Evaluation of nerve regrowth was performed on sagittal and axial images of the thighs. The conduit was identified on all images where possible as well as the nerve growth cone, as indicated by a change in T2 hyperintensity inside the conduit: distance of the growth cone was measured to the proximal and distal end of the visible conduit. In addition, all images were analyzed qualitatively for signs of inflammation (regional or local T2 STIR hyperintensity and T1 hypointensity) as well as for signs of hematoma and postoperative change.
To quantify muscle atrophy, regions of interest (ROIs) were drawn on the axial images encompassing all muscles of the calves, but excluding the tibia bone. Muscle volume was calculated separately for each leg.
Muscle atrophy—gastrocnemius muscle weight ratio
Rats were euthanized 12 weeks postsurgery in a CO2 chamber. Left (experimental) and right (control) gastrocnemius muscles were excised and weighed with an analytical balance (Mettler Toledo). For each rat, a gastrocnemius muscle weight ratio (E/C) based on the wet muscle weight of the experimental leg (left leg) versus the control leg (right leg) was calculated according to the following formula: E/C=weight experimental muscle/weight control muscle.
Cross sections proximal, of the newly formed nerves, and distal to the injury site
The sciatic nerves of the operated left sites were explanted over a length from 5 mm proximally to the proximal suture to 5 mm distally to the distal suture containing the medial part where the conduit had been sutured in. The explants were cut into three pieces, that is, medial through the former gap as well as proximal and distal to the lesion.
The regenerated nerve tissue was fixated with glutaraldehyde 2.5% for 2 h, followed by 0.01 M PBS overnight, before postfixation in 1% OsO4 for 2 h. After dehydration, the tissue was embedded using durcupan and semithin (2 μm) cross sections were cut with glass knives (Ultramicrotome Ultracut E, Reichert-Jung), mounted on uncoated glass slides, and finally stained with paraphenylenediamine 1% for 2 h to further enhance the myelin staining.
Histomorphometric analysis
For the morphometry analysis, pictures of the cross sections were taken using a light microscope (Olympus BX43) with a 10× and 40× magnification objective, respectively. Working with the 40× magnification, squares of 100×100 μm of all three localizations of each single nerve were analyzed using ImageJ 1.46 d as follows: The diameters of the axons as well as the total fiber diameters were measured manually always using the horizontal orientation and thus adjusting noncircular fibers. The average axonal diameter (AD), fiber diameter (FD), and myelin thickness were assessed. The axonal area (π AD/2)2 and the fiber area (π FD/2)2 were obtained, assuming the circularity of the nerve fiber area [36]. The myelinated area was considered as the difference between the fiber area and axonal area. Finally, the myelinated area per mm2 could be calculated and the myelinated axons per area were counted.
Morphometric analysis was carried out by an observer blinded to the experimental groups and localizations of cross sections (proximal, medial, distal). Blinding was achieved by numbering the nerve samples. Decoding took place after analysis was completed.
Statistical analysis
The SFI and re-/myelination, as well as the myelinated axon counts, were analyzed by linear mixed-effects models (MEMs) to account for the hierarchical structure of the data, that is, three repeated measurements of the SFI (under constant conditions) and three re-/myelination measurements and myelinated axon counts (proximal, medial, and distal) per individual rat. The MEM for the SFI included a fixed factor for the seven cell types and a random factor for an individual rat. In the MEM for re-/myelination (in %), we additionally included the proximal myelination, which should not be affected by the treatment as a fixed covariate to adjust for baseline differences in myelination, and a fixed factor for the remaining two locations of measurement (medial vs. distal). Likewise, proximal myelinated axon counts were included as a covariate in the MEM for myelinated axon counts. Because myelinated axon counts are count data, we used a generalized linear mixed-effects model (GLMM) with Poisson error distribution as a sensitivity analysis. Because the results were in line with the MEM and because the normality assumption for the MEM was not violated, we present the results of the MEM for the ease of interpretation. Muscle weight of the operated left side was analyzed by a general linear model, using cell type as a factor and muscle weight of the unoperated right side (baseline) as a covariate.
Six specific comparisons (contrasts) were a priori defined for the factor cell type: (1) differentiated versus undifferentiated rat ASCs: drASCs versus rASCs, (2) superficial versus deep abdominal layer human ASCs: hASCsup versus hASCdeep, (3) human ASCs versus Schwann cells: hASCsup/hASCdeep versus Schwann cells, (4) rASCs versus Schwann cells, (5) drASCs versus Schwann cells, and (6) human SVF cells versus Schwann cells.
For each contrast, an effect size was estimated along with a 95% confidence interval (CI), which corresponds to a significance level of 5%, and compared with the null hypothesis of no effect by means of a t-test.
All statistical analyses were done with R (Version 3.1.0).
Results
Animals and tissue processing
All animals survived the surgical procedure and recovered well from anesthesia. Except insignificant autotomy of the experimental foot, no surgical complications occurred (significant was considered as complete loss of or damage to multiple phalanges and/or the presence of open wounds, which would have led to premature euthanasia of the animal).
Sciatic functional index
Figure 1 shows the SFI by the cell group.
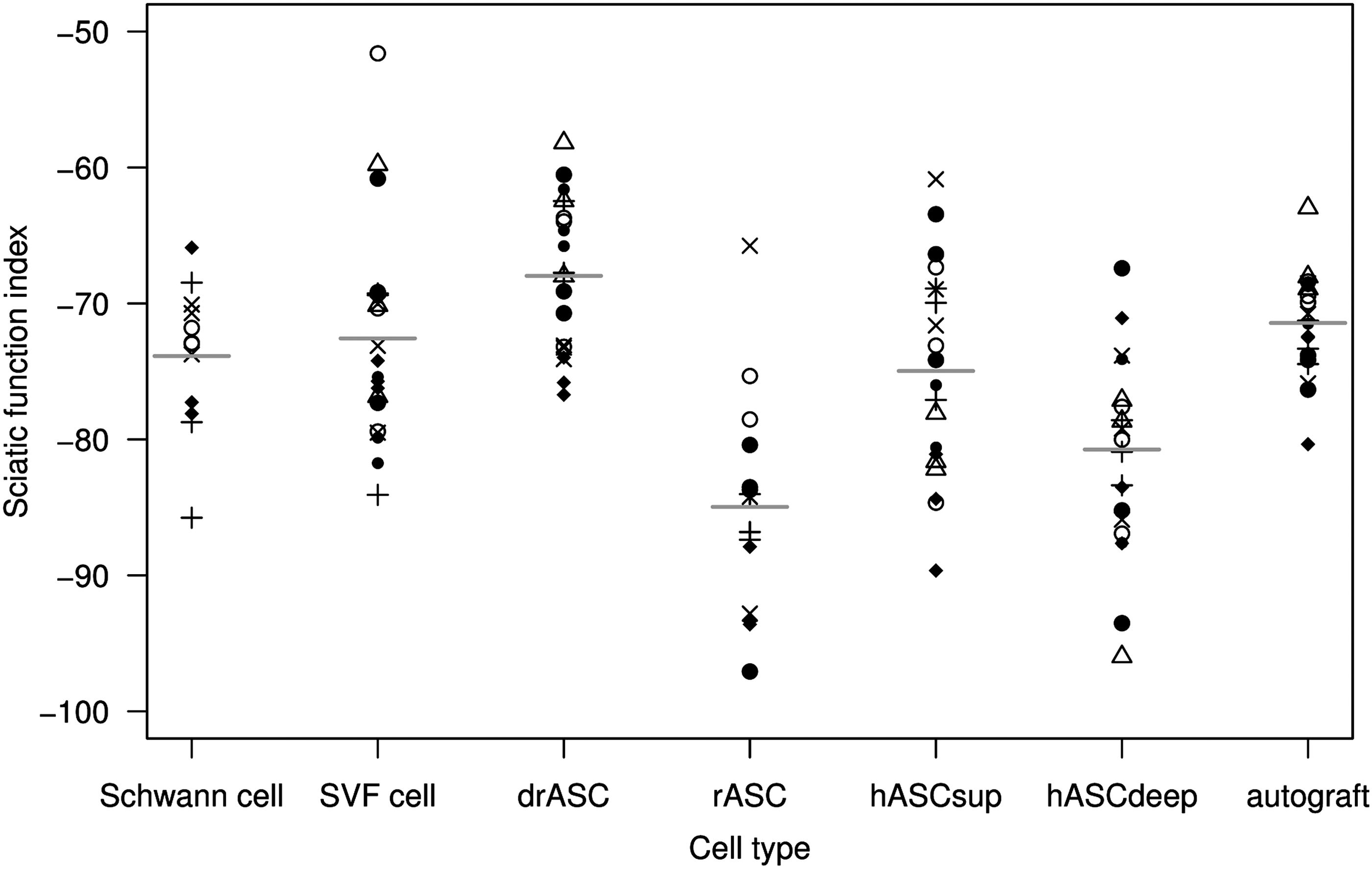
The sciatic functional index after 12 weeks by cell type group. Different symbols within a group distinguish rat individuals, but the same symbol in another group identifies a different individual since symbols are reused. The broken horizontal line indicates the mean value for the autograft, and short horizontal lines indicate mean values per cell type group.
Table 1 shows the effect size estimates from the linear mixed-effects model for the six predefined comparisons.
CI, confidence interval; drASC, differentiated rat adipose-derived stem cell; SVF, stromal vascular fraction.
Rats whose nerve had been repaired with drASCs had a higher mean SFI than those with undifferentiated rASCs (mean difference between drASCs and rASCs was 16.99, 95% CI=[11.15, 22.83]) and an even higher mean SFI than rats with an autograft. Superficial abdominal layer human ASCs (hASCsup) had a higher mean SFI than deep (hASCdeep) abdominal layer human ASCs (mean difference between hASCsup and hASCdeep 5.77, 95% CI=[0.44, 11.11]). While human abdominal layer ASCs (hASCsup/hASCdeep) and human SVF cells did not differ from rat Schwann cells in SFI, rASCs had a lower mean SFI and drASCs had a higher mean SFI than rat Schwann cells (mean difference −11.10, 95% CI=[−17.79, −4.41] and 5.90, 95% CI=[−0.36, 12.15], respectively).
MRI
MRI revealed gastrocnemius muscle atrophy of the operated side across all groups. Fibrin conduits were barely discernible after 12 weeks on T2-weighted and STIR images, indicating biodegradability. Figure 2 shows exemplary cross and sagittal sections in T1 and T2, as well as in short-T inversion recovery (STIR)-weighted contrast of an animal 12 weeks after complete neurotomy and nerve gap repair using a fibrin conduit in comparison with imaging 2 weeks postoperatively.
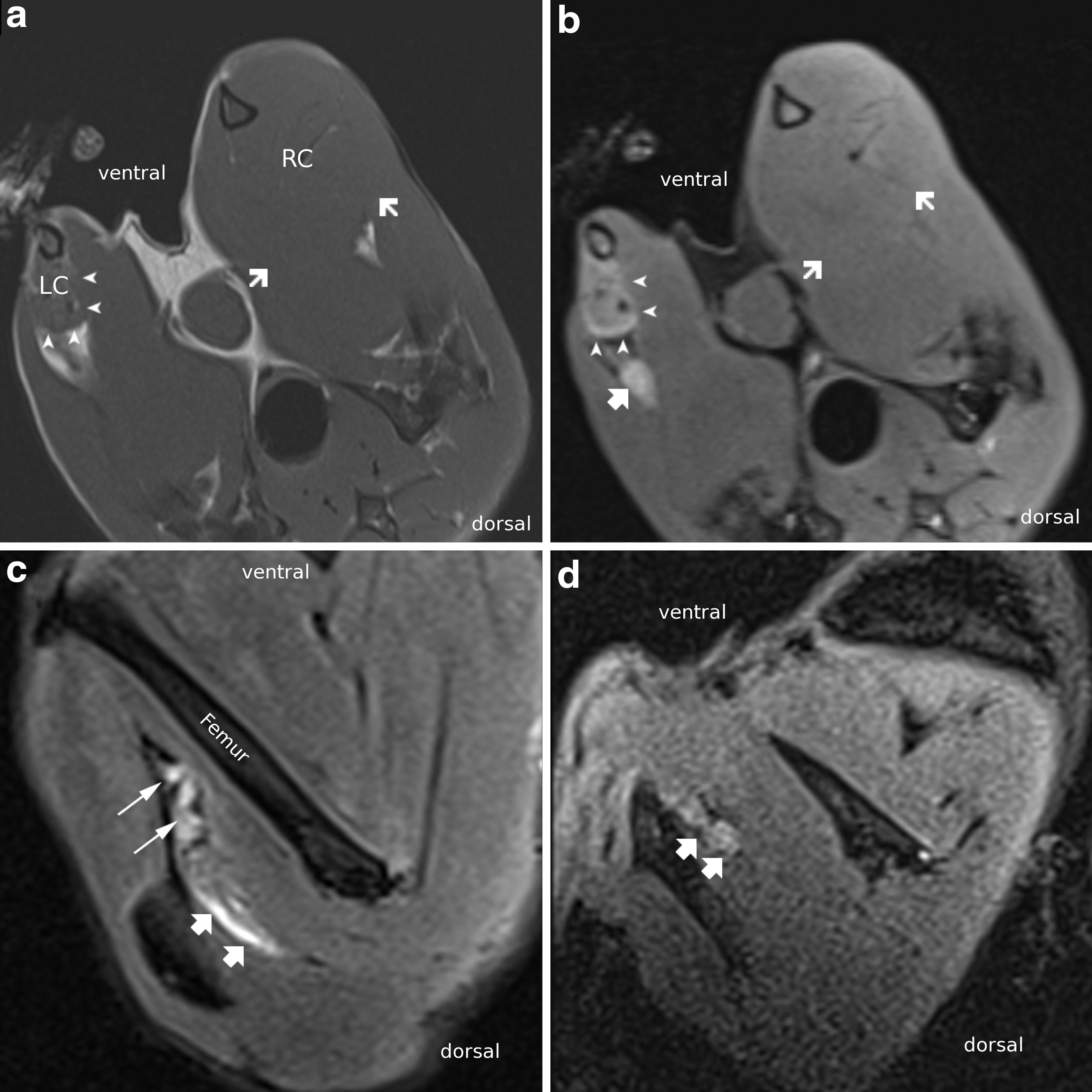
Cross-sectional scans of the calves in one animal 12 weeks after nerve suture in T1-weighted
Gastrocnemius muscle weights
Following complete neurotomy of the sciatic nerve, the gastrocnemius muscle undergoes atrophy due to the lacking innervation (Fig. 3). The reduction of muscle weight was quantified by weighing the excised gastrocnemius muscles of the experimental (E) and control (C) sides and calculating a ratio of their weight (E/C).
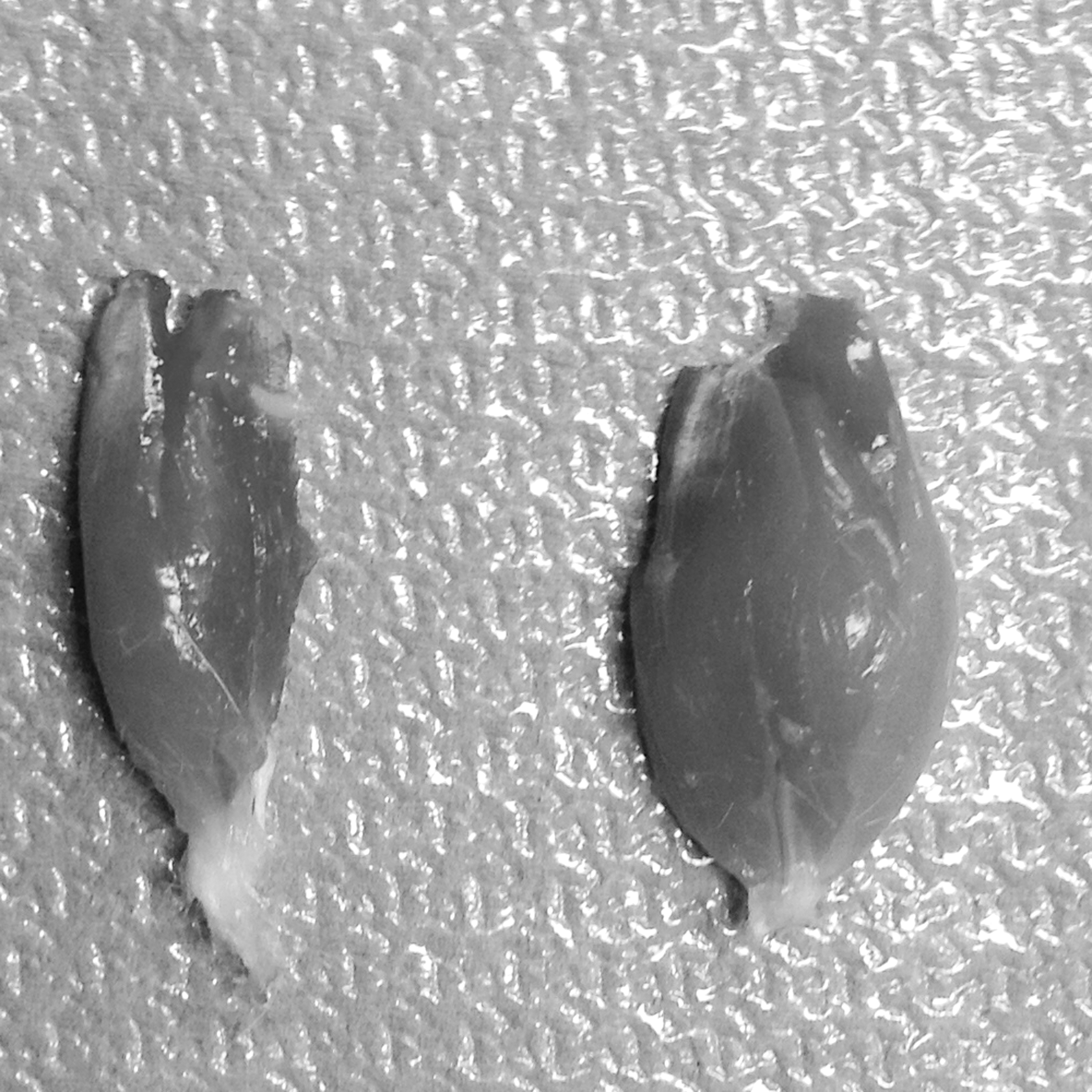
Gastrocnemius muscle atrophy 12 weeks after total neurotomy of the left sciatic nerve (experimental muscle: left; contralateral control muscle: right).
Figure 4 shows the relative muscle weight on the operated left side as a percentage of the muscle weight on the unoperated right side.

Relative muscle weight on the operated left body side (E) as a percentage of the muscle weight on the unoperated right side (C) after 12 weeks. The broken horizontal line indicates the mean value for the autograft, and short horizontal lines indicate mean values per cell type group. Each point represents one rat individual.
Table 2 shows the estimates from the linear model.
Muscle weight on the unoperated right side (baseline) was positively associated with muscle weight on the operated left side. The estimate of 0.42 (95% CI=[0.19, 0.65]) means that an increase in muscle weight on the unoperated right side by 1 g was associated with an increase in muscle weight on the operated left side by 0.42 g. Adjusting for differences in muscle weight on the unoperated right side, rats whose left nerve had been repaired with drASCs had a higher relative muscle weight than those with rASCs (Fig. 4). All other contrasts were nonsignificant. Rats with an autograft had the largest relative muscle weights.
Histomorphometric analysis
We analyzed light microscopy images of osmium-stained semithin slices (2 μm) of sciatic nerve sections 12 weeks after total neurotomy (Fig. 5). On the sciatic nerve samples proximal to the conduit, the sections are densely populated by myelinated axons. On the samples taken within the medial part between the two sutures where the conduit used to be, myelinated and nonmyelinated axons of varying caliber appear. Samples taken distal to the bridged area show reduced axon caliber with respect to the proximal domains. The frequency at which the smaller caliber axons are encountered in the distal domains is greater than the proximal aspects of the tissue, typical of the early stages of nerve regeneration [37]. The density of myelinated axons in the medial and distal parts is reduced when compared with the proximal area in all groups except for the Schwann cell group and the autograft.
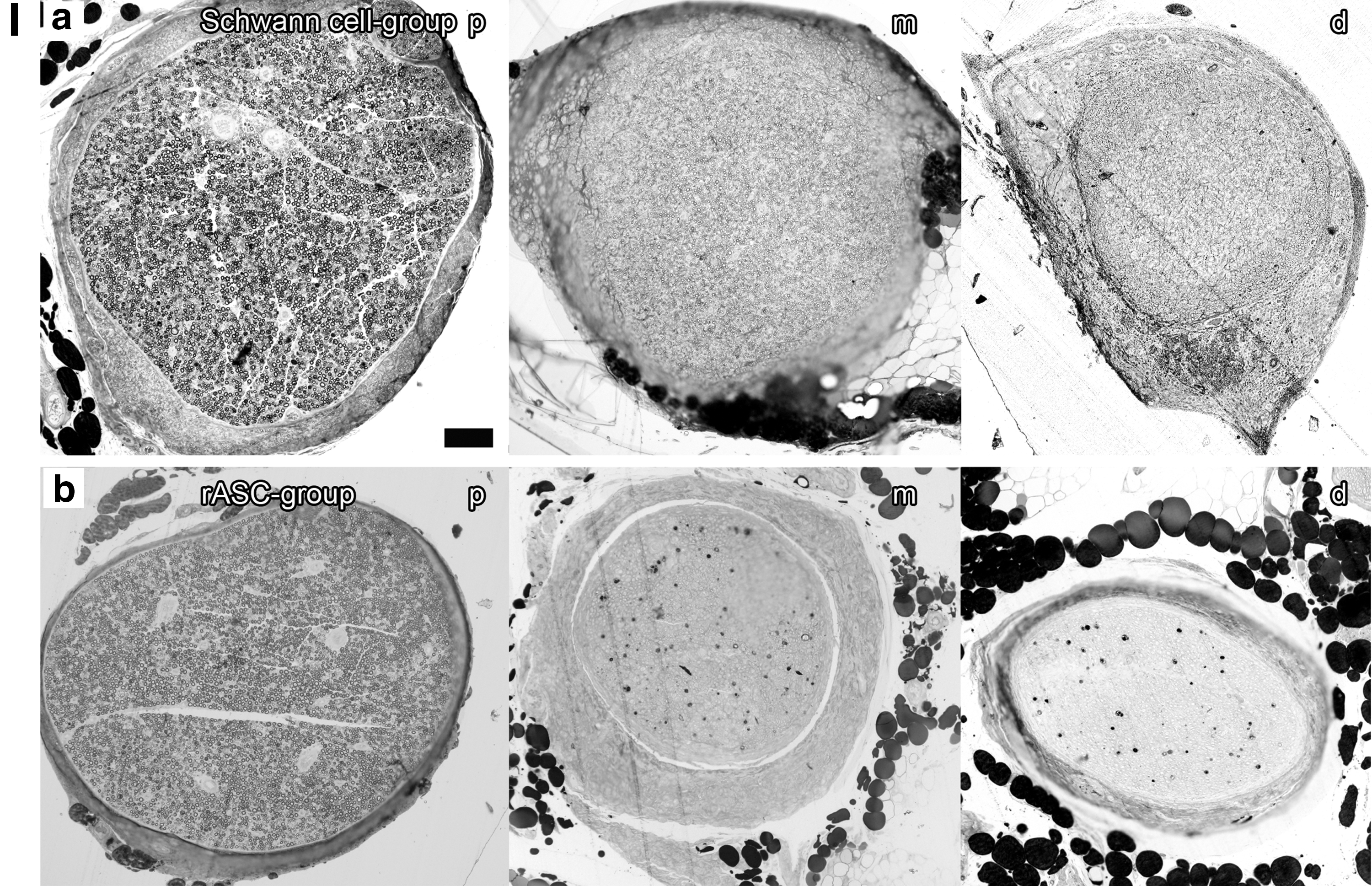
Light microscopy: Osmium-stained semithin slices (2 μm) of sciatic nerve sections 12 weeks after total neurotomy;
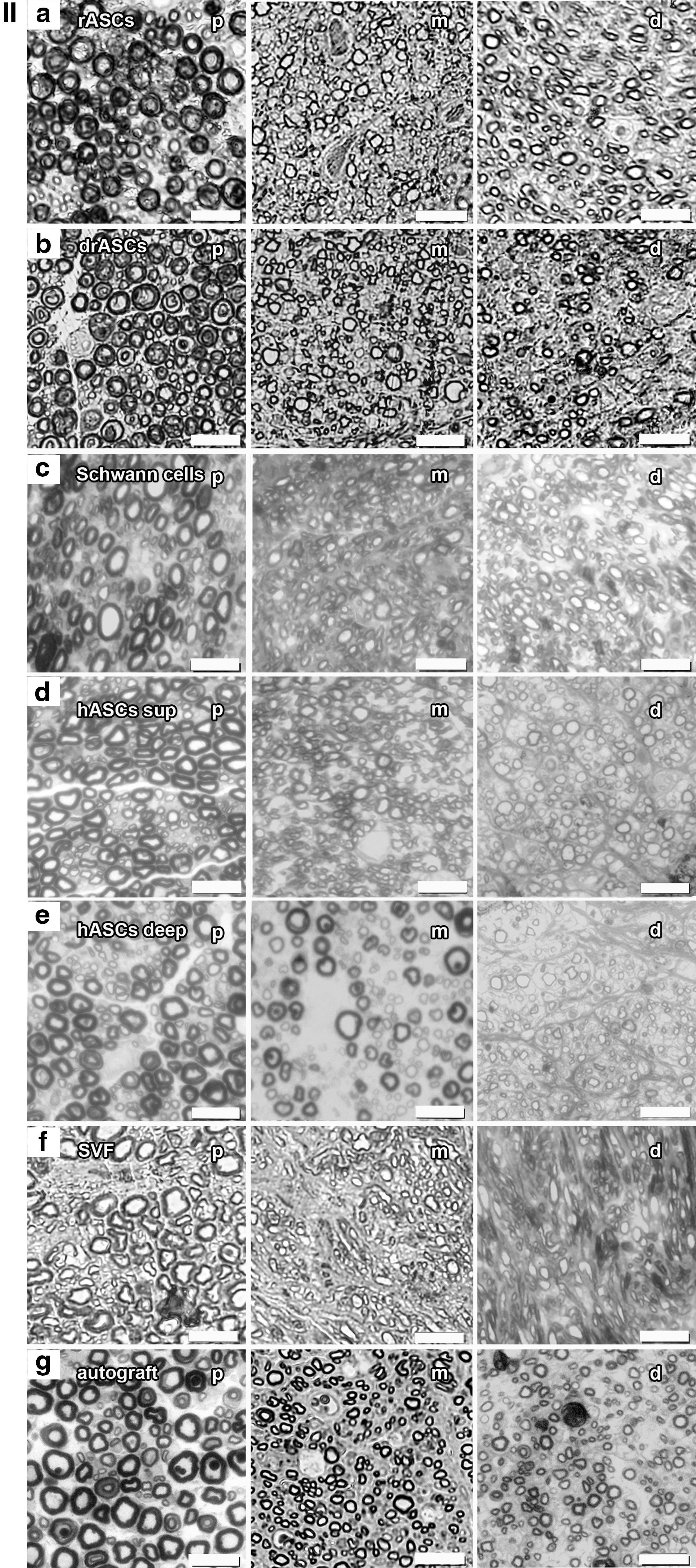
Figure 6 shows the re-/myelination and Fig. 7 shows the myelinated axon counts by the cell group and by the location of measurement.
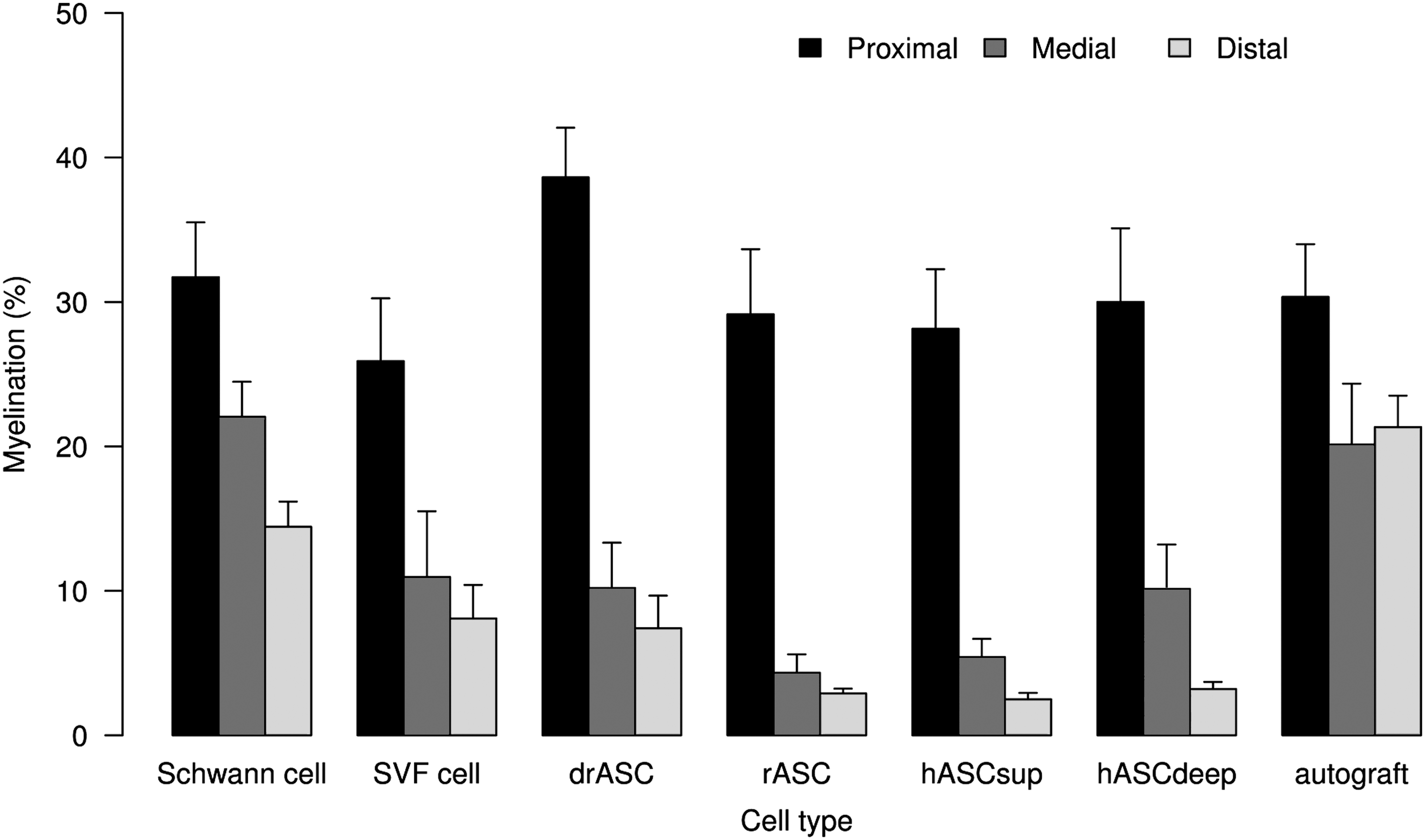
Re-/Myelination (%) after 12 weeks in cross sections, proximal, medial, and distal of the gap by cell type group.
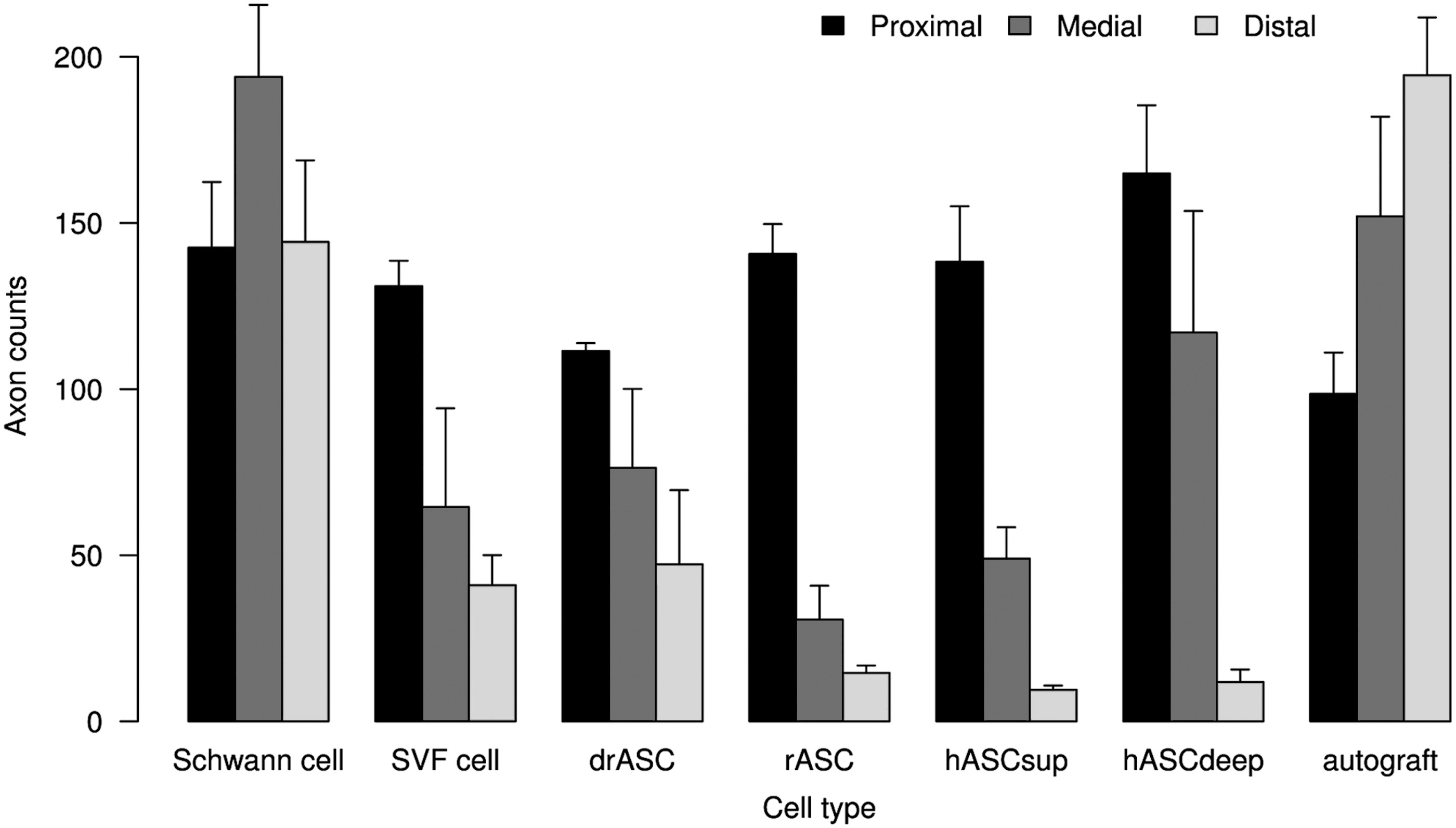
Myelinated axon counts (per 100 μm times 100 μm area) after 12 weeks in cross sections, proximal, medial, and distal of the gap by cell type group.
Table 3 shows the estimates from the linear mixed-effects model. Myelination of the proximal cross section (baseline) was marginally (P<0.1) positively associated with re-/myelination medial or distal. The estimate of 0.15 (95% CI=[0.03, 0.32]) means that an increase in proximal myelination by 1% was associated with an average increase in medial or distal re-/myelination by 0.15%. Medial re-/myelination was generally higher than distal remyelination (by 3.77% on average). Adjusting for differences in proximal myelination and for medial or distal location of measurement, re-/myelination with rat Schwann cells was superior to re-/myelination with human abdominal layer ASCs (hASCsup/hASCdeep) by 12.87 (95% CI=[7.18, 18.56])%, to undifferentiated rASCs by 13.84 (95% CI=[6.94, 20.74])%, to drASCs by 10.43 (95% CI=[4.06, 16.81])%, and to human SVF cells by 7.87 (95% CI=[0.48, 15.26])% (all contrasts significant).
Table 4 shows the fixed-effects estimates from the linear mixed-effects model. Proximal axon counts were not associated with medial or distal axon counts. Medial axon counts were generally higher than distal axon counts. The estimate of 34.25 (95% CI=[9.00, 59.49]) means that medial axon counts were on average by 34.25 higher than distal axon counts. Adjusting for differences in proximal axon counts, axon counts (medial and distal) with rat Schwann cells were superior to axon counts with human abdominal layer ASCs (superficial or deep) by 125.89 (95% CI=[72.60, 179.20]), to undifferentiated rat ASCs by 146.56 (95% CI=[83.19, 209.95]), to differentiated rat ASCs by 107.76 (95% CI=[47.57, 167.97]), and to human SVF cells by 116.54 (95% CI=[48.43, 184.68]) (all contrasts significant).
In both tables, we can see that there was neither a difference between hASCsup and hASCdeep nor between drASCs and undifferentiated rASCs.
Discussion
In this study, we could show that adipose tissue-derived cells are promising for nerve repair. Moreover, differentiation of adipose stem cells toward Schwann cell-like cells makes them, regarding the ease of procedure from harvesting to transplanting and finally the better functional results, even superior to Schwann cells.
ASCs were chosen in our trial not only due to their abundance, ease of harvest, high proliferation rates, and multipotency but also for therapeutic reasons, namely that they can be used in allogeneic transplantation [18,19,38].
Immunogenicity
ASCs in a rat model of allogeneic transplantation to immunocompetent hosts were shown to have a nonimmunogenic and suppressive profile. It could be concluded that the immune system does not negatively affect the ability of allogeneic ASCs to mediate tissue generation in vivo [39,40].
Sempere et al. examined human MSCs: the group showed results that provided mechanisms that may be involved with the in vivo MSC-mediated induction of tolerance, which to their point of view could be used therapeutically for reducing graft rejection [41]. Xie et al. found that anti-inflammatory proteins, which are induced from xenografted hASCs, are capable of responding to host inflammatory signals [42]. In a former study of our group, it was decided to perform xenogeneic transplantation experiments as a model of maximal HLA mismatch since no allogeneic model is available for human cells due to ethical reasons [43]. From a clinical point of view, no severe adverse events were observed during the course of the experiment related to the transplantation of human ASCs using an immunocompetent host with a functional immune system [44]. This observation goes in line with recent published studies, which describe the absence of immunological rejection toward xenograft stem cells [43]. What is more, in a xenotransplantation model, the systemic administration of hASCs to normal immunocompetent mice turned out to be safe and high doses were not toxic. Moreover, the effects of hASCs are more potent than those elicited by murine stem cells, what might be due to the already known relevant immunomodulatory potential of hASCs [45]. Indeed, in our study, the results achieved with hASCs did not differ significantly from those of rASCs with, yet, a tendency toward a better functional outcome, evident in the walking track analysis. This might suggest the absence of immunological interactions compromising nerve repair between the donor human cells and the host rat cells. Thus, we feel confirmed using our xenogeneic model and seeing the impact of differentiation itself shown in this study that in future studies, it will be interesting to analyze the impact of differentiation of human ASCs.
Human SVF filled conduits were used in one group of our animals. The SVF has the advantage of reduction in the interval from tissue collection until cell injection and simplicity of laboratory procedure, especially where a traumatic injury is dealt with [46]. It was presumed that neurotrophic activity of vascular cells attracted to the injury site by transplanted ASCs might be responsible for a long-term effect of those cells [47]. Based on this notion, it could be reasoned that SVF already containing such vascular cells might even be more effective in the long term. However, SVF cell treatment did not lead to differing results when compared with the other human cell lines. An explanation could be that better blood supply might have been counterbalanced by immunological interactions, which were likely evoked by nonimmunoprivileged cells, such as erythrocytes and leukocytes, composing the SVF.
Not only is the type of cell of interest but also in which way it generates the desired results. Different theories exist concerning the probable mode of action due to which ASCs enhance nerve regeneration:
Differentiation
In our study, differentiated rat ASCs were compared with undifferentiated adipose-derived stem cells. It was assumed that differentiation toward a Schwann cell-like phenotype was indispensable for nerve repair support.
In earlier studies, it was described that rat ASCs enhance neurite outgrowth, but only after treatment with various stimulating factors so that they were resembling Schwann cells [14,48]. Lopatina et al. demonstrated that the ability of mouse ASCs to stimulate the growth of nerve sprouts depends on brain-derived growth factor (BDNF) secretion. However, changes in BDNF secretion were only observed in ASCs incubated in differentiation medium [47]. Tomita et al. were able to show that differentiated human ASCs (dhASCs) secreted BDNF/nerve growth factor (NGF) at a comparable level and glial cell-derived neurotrophic factor (GDNF) at a level even higher than human Schwann cells, whereas undifferentiated human ASCs (uhASCs) secreted low levels of these neurotrophic factors. In coculture with NG108-15 neuronal cells, both dhASCs and human Schwann cells significantly increased the percentage of cells with neurites, the neurite length, and the number of neurites per neuron, whereas uhASCs increased only the percentage of cells with neurites [49]. The abovementioned goes in line with our results showing significantly better functional results and considerably less muscle atrophy in the drASC group when compared with the rASC group, which strongly supports the important role of in vitro differentiation before implantation (Tables 1 and 2).
Paracrine effects
Paracrine effects of ASCs means that they exert their effects by the release of factors, which stimulate tissue recovery at many potential levels, including stimulation of resident stem cells, remodeling of the extracellular matrix, and stimulation of blood vessel formation [45]. Erba et al. reasoned that any regenerative effect of transplanted ASCs is more likely to be mediated by an initial boost of released growth factors and/or by an indirect effect on endogenous Schwann cell activity since, in their trial, there were no significant quantities of viable cells 14 days after transplantation [50]. According to Hsiao et al., human adult MSCs support the engineering of functional tissue constructs by secreting angiogenic and cytoprotective factors to influence cell survival and vascularization [51]. ASCs were found to induce the local production of GDNF by Schwann cells and so their positive effectiveness was classified as a bystander effect since ASCs in culture do not produce detectable levels of GDNF [52]. There is evidence that the positive effects of adult stem cells on nerve repair are largely due to the fact that these appear to provide a protective microenvironment [45,50,53]. We have to state that indeed paracrine factors may have played a role in our trial and may also have influenced the different analyses leading to partly divergent results.
Biodegradable conduits
Good tissue acceptance was proven for the fibrin conduits in former studies of our group. Moreover, biodegradation kinetics turned out to be convenient [5,9]. In a recent study comparing four different biomaterial scaffolds (PCL, PLGA, fibrin, and collagen) for cord blood cell expansion, the results strongly suggest fibrin scaffolds with stromal (MSCs) support: The use of these scaffolds resulted in higher cell proliferation rates, increased adhesion, accelerated cell migration, and engraftment efficiencies [54]. Fibrin scaffolds that were used to deliver human MSCs led to a strikingly diminished invasion by host cells, suggesting that fibrin per se acts as a barrier against cellular invasion from the host surrounding tissue [55]. In this study, MRI imaging, 12 weeks after implantation, proved biodegradation of the fibrin conduit since it was only minimally discernible in T2-weighted contrast (Fig. 2).
Multimodal comparison
Sciatic functional index
Functional evaluation in our study (Fig. 1) revealed a higher mean SFI in the superficial than in the deep hASC group. This is in line with prior studies, which were able to show that cells from the superficial layer proliferate significantly faster and are relatively more resistant to apoptosis than those from the deep layer [22,24]. We could, moreover, find that rats whose nerve had been repaired with drASCs had a higher mean SFI than those with undifferentiated rASCs and an even higher mean SFI than rats treated with rat Schwann cells (Table 1). Since our a priori defined specific comparisons did not include the comparison between drASCs and autograft, statistical significance cannot be ascertained here, but, interestingly, the group treated with drASCs showed an even higher mean SFI than the autograft group regarding the absolute values (Fig. 1).
In addition, Orbay et al., when conducting a walking track analysis 6 months postoperatively, revealed even better improvement in the drASC group than in the nerve graft group. Their results showed significant improvement in the rASC group when compared to the empty silicon tube or a silicon tube filled with collagen gel and even the nerve graft group, whereas rASCs in our trial had a lower mean SFI when compared with the Schwann cell group. Orbay et al. assumed that a nerve graft does not integrate into the gap and just acts as a guide for newly sprouting axons, which led them to the conclusion that an empty conduit should perform the same scaffold function as a nerve graft. In contrast, cells introduced into a conduit are secreting stimulatory factors and thus enhance regeneration [2]. Unfortunately, their study did not comprise a Schwann cell group, although besides poor practicability of harvest and culture, Schwann cells appear to be the ideal cell line since PNS myelin is formed by Schwann cells and they therefore constitute the perfect comparison in the given context.
MRI
Results from the MRI evaluation indicate that nerve regrowth can be measured on a clinical 3-T scanner using FDA-approved equipment. While image contrast and resolution could be improved using dedicated high-field (eg, 7 Tesla) animal scanning facilities, our results could be directly translated into clinical practice as nerve conduits for palmar nerve transplant are of similar size to the ones used in this study and are within the field of view of the wrist coil.
While T2 hyperintensity can be caused by multiple mechanisms, for example, postoperative inflammation or foreign body reaction, we attribute the linear signal change as seen in Fig. 2 to the conduit itself as there is only minimal expansion of less than 1 mm into the surrounding tissue. In addition, we found circular signal increase on axial images with tram track-like bilinear signal on sagittal images on early imaging after nerve transplantation, while some animals showed clear postoperative changes in the subcutaneous regions of operative access (data not shown) [44]. Initial T2 signal increase of the distal nerve crush denervation and loss of signal after nerve repair have been described in animal models [56].
Gastrocnemius muscle weights
The results of the walking track analysis were partly consistent with the gastrocnemius muscle weights: Rats treated with drASCs had a higher relative muscle weight than those with rASCs (Fig. 4).
However, the animals of the autograft group by far had the largest relative muscle weights (46.97% when compared with the unoperated side), followed by the drASC group (29.51% when compared with the unoperated side). So, here we have to state inconsistency since, regarding the SFI, the drASC group visibly reached the autograft results.
Histomorphometry
Histomorphometric analysis (Fig. 5) showed generally higher medial than distal re-/myelination, which should reflect the nerve sprouts growing from the proximal toward the distal nerve stump accompanied by their sheaths. Re-/Myelination in the Schwann cell group was significantly superior to re-/myelination in all other cell groups (Table 3) and so was the number of myelinated fibers (Table 4). The myelinated axon counts in the medial and distal domains even exceeded that of the proximal section in the Schwann cell as well as in the autograft group. This can be interpreted as premature regeneration: Collateral myelinated fibers, so-called regenerating units, are sprouting from the proximal axon stump's growth cone toward the distal part of the nerve.
Thus, each unit may be said to be the anatomical expression of the regenerative activities of a single myelinated axon and the fiber count is therefore higher than the number of appendant neurons [57,58].
Lopatina et al. compared undifferentiated and differentiated mouse and human ASCs and showed that these express numerous genes of neurotrophins required for myelination. It was demonstrated that the ability of ASCs to upregulate nerve sprout growth even correlates with their production of BDNF, which in turn was upregulated by culturing the cells in neural differentiation medium before transplantation [47].
Regarding our data, there was no significant difference in re-/myelination or number of myelinated axons between cells that had been cultured in neural differentiation medium and undifferentiated rat ASCs.
Nor was there a difference between deep and superficial human abdominal layer ASCs, although the latter in our previous in vitro study were shown to promote neurite process outgrowth superiorly [22].
In another former study, our group compared drASCs and drBMSCs, as well as rat Schwann cells; autografts and empty fibrin conduits served as controls. No significant differences with regard to the myelination could be shown within the cross sections of the distal stumps. Overall, the best results here as well were achieved in the Schwann cell group and similar outcomes were found in the drASC and drBMSC groups.
Nonetheless, based on the above-mentioned applicability of drASCs, it was concluded and summarized that these are the most promising cells for future clinical use [5].
Incongruity within the modes of examination
We have to state that despite the superior functional results, myelination in the drASC group was not significantly better and even muscle atrophy was less in the autograft group when compared with the drASC group. Partial incongruity within the modes of examination could also be found in a study of Liu et al., which revealed a significantly higher SFI in the ASC group than in the autograft group. In their histological observation and immunohistochemistry in contrast, where they examined the morphology and distribution patterns of nerve fibers, similar results were observed in the ASC nerve conduits and the autografts [10].
Scholz et al. observed a 4-month period and found that the autograft group had the best SFI, followed by the group with tissue-engineered nerve construct with differentiated hASCs and renewal of differentiation medium at days 14 and 28, which was not significantly inferior. Regarding the gastrocnemius muscle weight per contra, the latter group showed significantly less atrophy when compared with all other groups, including the autograft [59]. The observed period of time may play an important role regarding the dynamic visible at the different time points.
Focusing on the re-/myelination, drASCs are able to express the myelin proteins found in the PNS, thus there is evidence that these cells are morphologically and functionally similar to Schwann cells. Nevertheless, although electron microscopy showed the alignment of differentiated cells along the neurites, it was not possible to demonstrate the formation of myelin around axons in a coculture model. The lack of myelin formation was justified with the short time of coculturing [60]. Apparently, also in the long-term, there are other mechanisms than myelination that improve the functional outcome such as secretion of soluble factors and contact-mediated mechanisms such as the observed cell bridging and cell towing [61,62].
In part, it is known that these features differ between undifferentiated and differentiated MSCs. This could account for the good muscle weight of the drASC group when compared with the other cell groups and the superior SFI, despite the lack of significantly better re-/myelination.
Conclusion
Having previously shown the gain of cell assistance in fibrin conduits in a nerve gap model for peripheral nerve repair, this study went one step further: It provides a broad overview by comparing a variety of promising cell lines according to the actual state of research in a multimodal manner under equal experimental conditions over a 12-week period. Conducting functional and morphometric, as well as MRI analyses, we were able to show that the benefit of adipose tissue-derived adult stem cells can be enhanced by in vitro differentiation into Schwann cell-like cells before transplantation and thus even exceed the functional results of Schwann cells. In future studies aiming for human applications, we will consider these findings in rodents and therefore we would also like to analyze the impact of differentiation of human ASCs before implantation. Finally, it will be our goal to replace the autograft with its donor site morbidity.
Ethical conduct of research
The authors state that they have obtained appropriate institutional review board approval or have followed the principles outlined in the Declaration of Helsinki for all human or animal experimental investigations. In addition, for investigations involving human subjects, informed consent has been obtained from the participants involved.
Footnotes
Acknowledgments
The authors would like to thank Prof. Markus Tolnay and Prof. Stephan Frank for their support throughout the study, as well as Dr. Romano Kasper, Katerina Ismene Kappos, and Miriam Lorenz for organizational workup. This work was supported by the Swiss National Science Foundation (SNF grant #31003A_133076/1) and the Gottfried und Julia Bangerter-Rhyner-Stiftung, as well as by the Department of Surgery, University Hospital of Basel, the Nachwuchsförderungsfonds of the University of Basel, and the Freie Akademische Gesellschaft Basel.
Author Disclosure Statement
No competing financial interests exist.