
Abstract
Mesenchymal stem cell (MSC) differentiation is dramatically reduced after long-term in vitro culture, which limits their application. MSCs derived from induced pluripotent stem cells (iPSCs-MSCs) represent a novel source of MSCs. In this study, we investigated the therapeutic effect of iPSC-MSCs on diabetic mice. Streptozocin-induced diabetic mice transplanted with 400 islets alone or with 1×106 iPSC-MSCs were examined following rapamycin injection (0.1 mg/kg/day, i.p., from days 0 to 9) after transplantation. Our results showed that iPSC-MSCs combined with rapamycin significantly prolonged islet allograft survival in the diabetic mice; 50% of recipients exhibited long-term survival (>100 days). Histopathological analysis revealed that iPSC-MSCs combined with rapamycin preserved the graft effectively, inhibited inflammatory cell infiltration, and resulted in substantial release of insulin. Flow cytometry results showed that the proportion of CD4+ and CD8+ T cells was significantly reduced, and the number of T regulatory cells increased in the spleen and lymph nodes in the iPSC-MSCs combined with the rapamycin group compared with the rapamycin-alone group. Production of the Th1 proinflammatory cytokines interleukin-2 (IL-2) and interferon-γ was reduced, and secretion of the anti-inflammatory cytokines IL-10 and transforming growth factor-β was enhanced compared with the rapamycin group, as determined using enzyme-linked immunosorbent assays. Transwell separation significantly weakened the immunosuppressive effects of iPSC-MSCs on the proliferation of Con A-treated splenic T cells, which indicated that the combined treatment exerted immunosuppressive effects through cell–cell contact and regulation of cytokine production. Taken together, these findings highlight the potential application of iPSC-MSCs in islet transplantation.
Introduction
I
MSCs showed various degrees of efficacy in preclinical animal studies [27]; however, their limited accessibility is a major factor inhibiting their use in routine clinical treatment. Current methods to obtain MSCs from patients are invasive and labor intensive. Furthermore, MSCs have a limited capacity to expand in culture. Successive passages slow the proliferation rate, and MSCs progressively lose their multipotency and lack immunosuppressive activity. In addition, aging and age-related disorders significantly impair the survival and differentiation potential of BM-MSCs, thus limiting their therapeutic efficacy [28 –32]. Therefore, it is important to identify alternative sources of MSCs before they can be used as a mainstream treatment for organ transplantation. A breakthrough in the generation of human-induced pluripotent stem cells (iPSCs) from adult somatic cells offered the possibility of generating a high yield of MSCs [33 –35].
Several laboratories have found that iPSC-derived MSCs have the same in vitro and in vivo characteristics as MSCs derived from adult sources. Previous studies indicate that iPSC-MSCs grown on a calcium phosphate scaffold enhanced osteogenic differentiation and promoted bone regeneration [36 –38]. iPSC-MSCs could form mature mineralized structures that were histologically similar to mature bone, facilitating periodontal regeneration [39,40]. Transplanting iPSC-MSCs attenuated severe hindlimb ischemia and improved the hepatic function in mouse models [33,41,42]. These results suggested that iPSC-MSCs have high potential for tissue-engineering applications. In addition to their tissue repair ability, iPSC-MSCs also exhibit immunomodulatory properties [43 –45]. For example, iPSC-MSCs displayed long-lasting immunosuppressive properties toward natural killer cells by interfering in their activation, thus protecting target cells [44]. Human iPSC-MSCs exerted immunomodulatory effects on T-cell subsets in the peripheral blood from allergic rhinitis patients by modulating T-cell phenotypes toward Th2 suppression and inducing T regulatory cell (Treg) expansion [45]. iPSC-MSCs also prevented allergic airway inflammation in mice [43]. Therefore, iPSC-MSCs may be a novel source of tolerance induction, although their immunosuppressive activity in organ transplantation remains to be explored. The aim of this study was to assess the efficacy of iPSC-MSCs in combination with Rapa in islet transplantation immunosuppressive therapy in streptozocin (STZ)-induced diabetic mice.
Materials and Methods
Animals
Female BALB/c mice (8–12 weeks old) and C57BL/6 were purchased from SLAC Laboratory Animal Co., Ltd. (Shanghai, China) and used as graft donors and recipients, respectively. The care and handling of the animals were conducted in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. The protocol was approved by the Animal Studies Committee of Xiamen University, China.
Drugs
Rapamycin was purchased from LC Laboratories (LC Labs, Woburn, MA) and dissolved in PBS at 0.02 mg/mL for injection. The recipient mice received Rapa at doses of 0.1 mg/kg/day i.p. from days 0 to 9 after transplantation.
Phenotypic analysis of iPSC-MSCs
iPSC-MSCs were kindly provided by Prof. Qi-Zhou Lian of the University of Hong Kong. The iPSC lines were prepared from iPSCs, which were reprogrammed from human fibroblast cells, and differentiated into MSCs according to a previously described protocol [33]. Briefly, MSCs were purified by sorting for CD105+/CD24− cells and maintained in a medium containing 90% knockout Dulbecco's Modified Eagle's Medium (Gibco, Invitrogen Corporation, Carlsbad, CA) supplemented with 10% serum replacement medium (Gibco) and basic fibroblast growth factor (10 ng/mL; Gibco). The morphology of iPSC-MSCs was very similar to BM-MSCs and they have the capacity to differentiate into osteoblasts, adipocytes, and chondroblasts [33]. Millipore's FlowCellect™ Human Mesenchymal Stem Cell Characterization Kit was used for the phenotypic analysis of iPSC-MSCs. iPSC-MSCs (passage 5–10) were resuspended in an assay buffer and then incubated with an antibody working cocktail solution for 30 min on ice in the dark. The antibody working cocktail solution contained anti-CD105/PE-, anti-CD90/FITC-, anti-CD73/APC-, and anti-CD14/CD34/CD45/PerPC-conjugated antibodies. Each fluorescence analysis included the appropriate FITC-, PE-, or cytochrome-conjugated isotype Ab controls. Cells were separated using flow cytometry (FACS Calibur; Becton-Dickinson, San Diego, CA) and not sorted. The use of iPSC-MSCs in this study was approved by the Ethics Committee of Xiamen University, China.
Chemical induction of diabetes
Diabetes was induced in female C57BL/6 mice by intraperitoneal injection of streptozocin (180–220 mg/kg; Sigma-Aldrich, St. Louis, MO) [46]. Blood glucose was measured using a FreeStyle glucose meter (Abbott, Alameda, CA), and diabetes onset was defined as two consecutive daily blood glucose measurements above 16.7 mmol/L.
Islet isolation, purification, and transplantation
BALB/c islets were isolated using the digestion method [47,48] with collagenase P (1 mg/mL; Roche, Basel, Switzerland). The pancreas was perfused through bile duct cannulation with 3 mL of 1 mg/mL collagenase P per mouse and then excised. Briefly, the pancreas was digested at 37°C–38°C for 20 min and then shaken vigorously in cold Hank's Balanced Salt Solution (HBSS) containing 10% fetal bovine serum (FBS; Shanghai ExCell Biology, Shanghai, China). The digested pancreatic tissues were filtered through a 200-μm mesh, washed thrice [49], and then purified using human mononuclear cells and granulocyte separation media Histopaque-10771 and Histopaque-11191 (Sigma-Aldrich). The islets were sorted manually under the microscope.
Four hundred BALB/c islets were used for a single transplantation. Islets were transplanted under the kidney capsule of diabetic C57BL/6 mice. Blood glucose levels were monitored in the recipient mice. Islet transplants were considered functional when two consecutive blood glucose measurements were <8 mmol/L, and graft rejection was defined as a blood glucose level of >11.1 mmol/L on 2 consecutive days. Body weight was recorded (daily) until complete graft rejection occurred.
Recipient therapy and experimental groups
Four hundred islets isolated from BALB/c mice with or without 1×106 iPSC-MSCs were transplanted into diabetic mice. The control group received islets alone. The Rapa group was treated with 0.1 mg/kg/day Rapa alone. The iPSC-MSCs+Rapa group received islets with iPSC-MSCs and was treated with 0.1 mg/kg/day Rapa. Rapa treatments were conducted from days 0 to 9.
Mixed lymphocyte reaction
Nylon wool columns (Wako, Osaka, Japan) were used to isolate T cells from the spleen of the recipient mice, which were used as responder cells. Spleen cells obtained from the BALB/c mice were used as stimulator cells. The responder cells (5×105 cells) were cultured in 96-well plates in the presence of stimulator cells (5×104 cells), pretreated with mitomycin C (40 μg/mL; Amresco, Solon, OH) in 200 μL RPMI 1640 supplemented with 10% FBS and 1% penicillin and streptomycin, and incubated at 37°C in a 5% CO2 humidified atmosphere for 72 h. Cell proliferation was measured using a bromodeoxyuridine (BrdU) cell proliferation assay kit (Roche Applied Science, Mannheim, Germany). The magnitude of the absorbance is proportional to the quantity of BrdU incorporated into cells, which is a direct indication of the cell proliferation rate. The optical density values were measured in an enzyme-linked immunosorbent assay (ELISA) reader (Model 680; BIO-RAD, Hercules, CA) at 450 nm (the reference wavelength was 690 nm). Measurements were performed in triplicate.
Transwell experiments
For the Transwell experiments, 24-well Transwell plates with a 4-μm-pore membrane (Costar, Corning, NY) were used to separate T cells from the iPSC-MSCs. T cells were isolated from the spleens of C57BL/6 mice using nylon wool columns (Wako). iPSC-MSCs were plated into the lower chamber at 5×104 cells/well, and 5 μg/mL Con A (Sigma-Aldrich)-stimulated T cells (5×105 cells/well) were cultured in the upper chamber of the Transwell insert. Cell culture media were supplemented with 2 ng/mL Rapa. After 3 days of coculture, T cells were harvested and placed in a 96-well plate at a concentration of 1×105 cells/well (n=6). Cell proliferation was measured using the BrdU cell proliferation assay kit (Roche Applied Science), as described above. Measurements were performed in triplicate.
Flow cytometry analysis
Recipient splenic lymphocytes and T cells (isolated using nylon wool columns) were prepared in 100 μL PBS per 1×106 cells. The splenic lymphocyte cells were incubated with PE-Cy5-anti-CD4 (GK1.5), FITC-anti-CD8 (53-6.7), and their isotype controls (purchased from BioLegend, San Diego, CA) at 4°C for 30 min. T cells from lymph nodes were incubated with FITC anti-CD4 (RM4-5) and PE anti-Foxp3 (FJK-16s) (purchased from eBioscience, San Diego, CA) at 4°C for 30 min. Conjugated isotype antibodies were used as negative controls. The stained cells were detected on a FACScan flow cytometer (Partec Co., Munster, Germany), and the data were analyzed using FlowJo software (Tree Star, Inc., Ashland, OR).
The secretions of interleukin-2 (IL-2), IL-10, and interferon-γ (IFN-γ) into the recipient sera were detected using a Cytometric Bead Array™ (CBA; BD Biosciences, San Jose, CA), according to the manufacturer's instructions. The stained samples were detected on a BD FACS Aria Cell Sorter (BD Biosciences), and the data were analyzed using FlowJo software and FCAP Array software (BD Biosciences).
Histopathological analysis
Kidney islet grafts were removed from recipient mice at day 12 post-transplantation, fixed in 4% paraformaldehyde fixative (Beijing Solarbio Science & Technology Co., Ltd., Beijing, China), and embedded in paraffin. Grafts were cut into 5-μm sections, stained with hematoxylin and eosin (H&E), and examined by a transplant pathologist who was blinded to treatment modality. Representative specimens (n=36) from all treatment modalities were ranked from 1 to 36 (from least to most) for overall rejection/inflammation, with the median in each group presented [47].
Immunohistochemistry
At day 12 after islet graft, the islet grafts were removed for pathological examination, fixed in zinc fixative (Biolegend), and embedded in paraffin. The paraffin tissues were cut into 5-μm sections, deparaffinized in xylene, hydrated through graded ethanol series, and immersed in absolute methanol, which contained 0.3% hydrogen peroxide, for 10 min to block the endogenous peroxidase activity. Sections were incubated with nonimmune goat serum for 20 min to prevent nonspecific binding and then with the primary Insulin Rabbit mAb (1:100; Cell Signaling Technology, Boston, MA) diluted in PBS for 1 h. The sections were incubated with Polymer Helper for 15 min, with poly-HRP anti-rabbit IgG for 30 min, and with the peroxidase substrate diaminobenzidine for 1 min. Slides were counterstained with hematoxylin. The slides were examined under a microscope and evaluated in a blinded manner.
Quantitative real-time reverse transcription PCR analysis
Kidney islet grafts were removed from recipient mice at day 12 post-transplantation, and the mRNA was extracted using TRIzol (Life Technologies, Carlsbad, CA). Reverse transcription and quantitative real-time polymerase chain reaction (qRT-PCR) were performed using commercially available reagents (Toyobo, Osaka, Japan). The StepOne Real-Time PCR System (ABI, Foster City, CA) was used to detect IL-2, IFN-γ, IL-10, transforming growth factor (TGF-β), and Foxp3. β-Actin served as the control. Calculation of the relative expression was performed using the 2−ΔΔCT method. The following primer sequences were used for qRT-PCR:
β-actin: forward 5′-CATCCGTAAAGACCTCTATGCC AAC-3′
and reverse 5′-ATGGAGCCACCGATCCACA-3′;
IFN-γ: forward 5′-CGGCACAGTCATTGAAAGCCTA-3′
and reverse 5′-GTTGCTGATGGCCTGATTGTC-3′;
IL-2: forward 5′-GGAGCAGCTGTTGATGGACCTAC-3′
and reverse 5′-AATCCAGAACATGCCGCAGAG-3′;
IL-10: forward 5′-GACCAGCTGGACAACATACTGC TAA-3′
and reverse 5′-GATAAGGCTTGGCAACCCAAGTAA-3′;
TGF-β: forward 5′-GACCAGCTGGACAACATACTGC TAA-3′
and reverse 5′-GATAAGGCTTGGCAACCCAAGTAA-3′;
Foxp-3: forward 5′-CAGCTCTGCTGGCGAAAGTG-3′
and reverse 5′-TCGTCTGAAGGCAGAGTCAGGA-3′.
Enzyme-linked immunosorbent assay
Supernatants from the mixed lymphocyte reaction (MLR) after a 72-h incubation and the sera of recipient mice were collected and frozen at −20°C. ELISAs were performed using commercially available kits (NeoBioscience Technology Co., Ltd., Beijing, China) to detect the secretion levels of IL-2, IFN-γ, IL-10, and TGF-β. The process was conducted according to the manufacturer's instruction. Each reaction was carried out in triplicate.
Statistical analyses
The median survival times of the four groups were calculated and compared using the Kaplan–Meier method. Data from MLR, FACS, ELISA, and CBA experiments were analyzed by one-way analysis of variance (ANOVA) and expressed as the mean±standard deviation. A Bonferroni correction was calculated and applied because multiple comparisons were made during the analysis. A P value<0.05 was considered statistically significant; P<0.01 and P<0.001 indicated highly significant differences. All analyses were performed using the GraphPad Prism® (GraphPad Software, Inc., San Diego, CA) software.
Results
Flow cytometry analysis of iPSC-MSC surface antigens
iPSC-MSCs exhibited a spindle-shaped morphology and their identity was confirmed by flow cytometry. The results showed that the percentage of CD73, CD90, and CD105 triple-positive cells and the percentage of CD14, CD34, and CD45 triple-negative cells were 86.94±0.87 and 3.29±2.83, respectively, where the latter showed no significant difference compared with the isotype control (Fig. 1). The results showed that iPSC-MSCs expressed the cell surface marker characteristic of MSCs (CD73, CD90, and CD105) and were negative for markers typically absent on MSCs (CD14, CD34, and CD45). These results suggest that iPS-MSCs display morphological characteristics of adult MSCs.
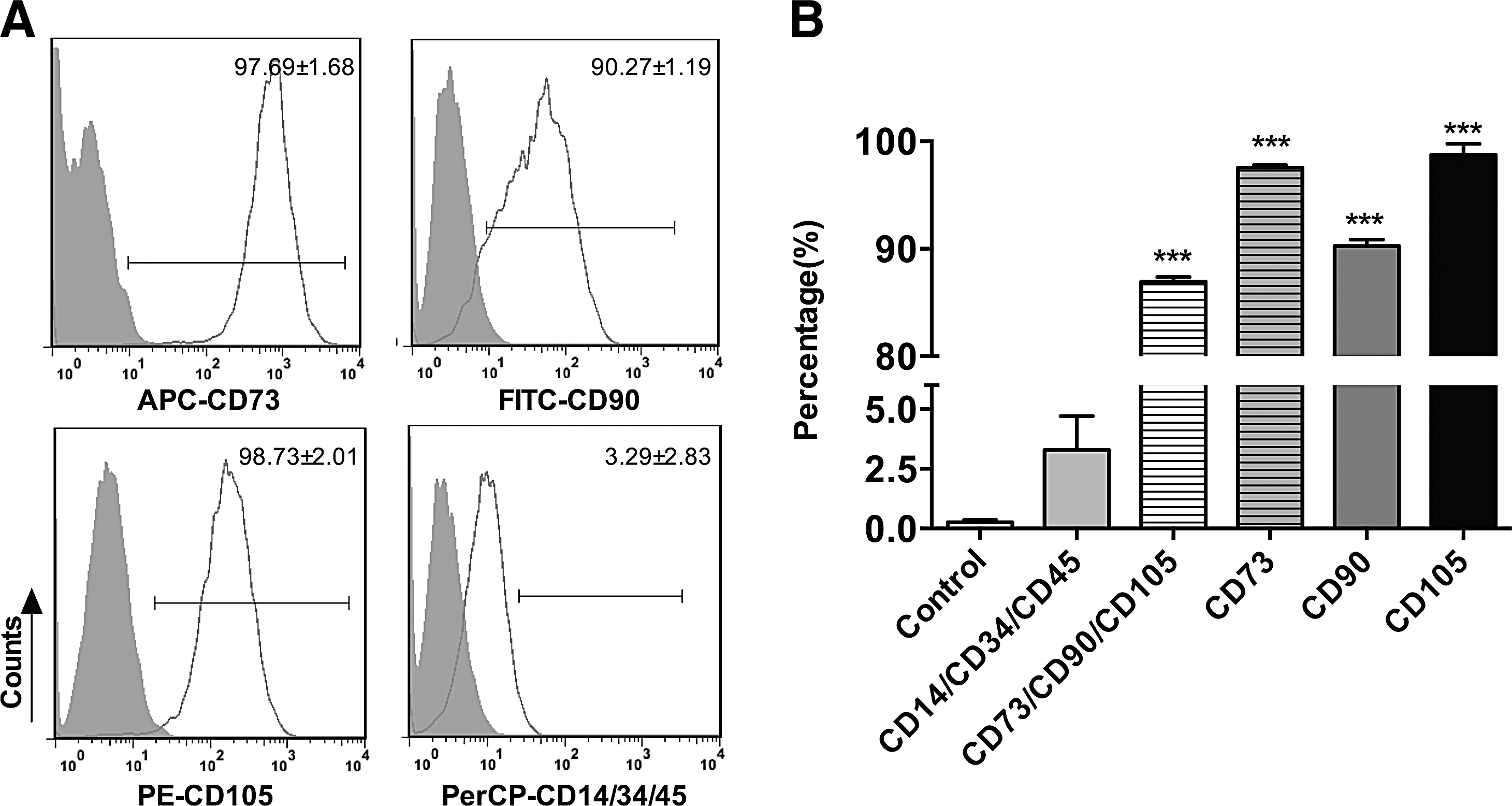
Cell surface antigens for iPSC-MSCs analyzed by flow cytometry.
Effect of iPSC-MSCs on islet allograft survival
We investigated the effect of iPSC-MSCs on islet allograft survival in STZ-induced diabetic mice by cotransplantation into recipient mice by kidney subcapsular injection (Fig. 2A). The survival times of the iPSC-MSCs and Rapa groups were 14 and 18 days, respectively, both of which were significantly prolonged compared with 12 days in the controls (P<0.05, iPSC-MSC group; P<0.001, Rapa group). iPSC-MSCs combined with Rapa prolonged survival time, compared with Rapa alone (P<0.001), and induced immune tolerance in 50% of the recipients. Blood glucose values (measured every 3 days) remained normal in Rapa-treated iPSC-MSCs, until the mice were nephrectomized at day 102 post-transplantation (Fig. 2B).

Survival of islet allografts treated with iPSC-MSCs and Rapa. Islet graft survival and blood glucose levels in C57BL/6 mice with different treatments are shown, respectively
Effect of iPSC-MSCs on the inflammatory response of STZ-induced diabetic mice following islet transplantation
To investigate the effect of iPSC-MSCs on grafts in diabetic mice, grafts were dissociated from recipient mice at day 12 post-transplantation and processed for histological analysis (Fig. 3A). Grafts from control mice exhibited islet damage and little insulin secretion. Grafts from iPSC-MSCs or Rapa-treated mice showed less islet damage, more insulin secretion, and fewer infiltrating inflammatory cells. iPSC-MSCs+Rapa treatment preserved the graft most effectively and resulted in a substantial release of insulin. The overall ranking of rejection/inflammation is shown in Fig. 3B. The rankings given for the combined treatment group were significantly lower in terms of rejection/inflammation than those for the Rapa group (P<0.01). We next examined the expressions of inflammatory cytokines in graft and sera using qRT-PCR, ELISA, and CBA flow cytometry (Fig. 3C, D). IL-2 and IFN-γ in the iPSC-MSCs+Rapa treatment group were downregulated at both the mRNA and protein levels compared with the Rapa group. These results demonstrated that iPSC-MSCs and Rapa have synergistic effects on the expressions of inflammatory cytokines.

Preservation of islet graft and infiltration of inflammatory cells at 12 days post-transplantation.
Effect of iPSC-MSCs on CD4+ and CD8+ ratios in STZ-induced diabetic mice
Flow cytometry was used to investigate the effect of iPSC-MSCs+Rapa treatment on CD4+ and CD8+ T lymphocytes at day 12 post-transplantation. Although iPSC-MSCs or Rapa alone effectively decreased the number of CD4+ and CD8+ T lymphocytes, iPSC-MSCs combined with Rapa showed the best inhibitory effect on the proliferation of CD4+ and CD8+ T lymphocytes, whether in the spleen or in lymph nodes (Fig. 4B).

Proportion of CD4+ and CD8+ T lymphocytes in the spleen and lymph nodes at 12 days post-transplantation.
Protective effect of iPSC-MSCs+Rapa on islet allografts through Tregs induction
The proportion of splenic and lymph node Tregs was examined by flow cytometry (Fig. 5A). Although the results showed that both iPSC-MSCs and Rapa induce Tregs compared with the controls, iPSC-MSCs+Rapa treatment induced a larger numbers of Tregs compared with the Rapa group (Fig. 5B). We next examined IL-10, TGF-β, and Foxp3 expression in the islet grafts and sera of the recipient mice. iPSC-MSCs+Rapa treatment increased serum IL-10 and TGF-β concentrations (Fig. 5C, D). Furthermore, IL-10, TGF-β, and Foxp3 mRNA levels in the graft also increased compared with the Rapa group (Fig. 5E–G).
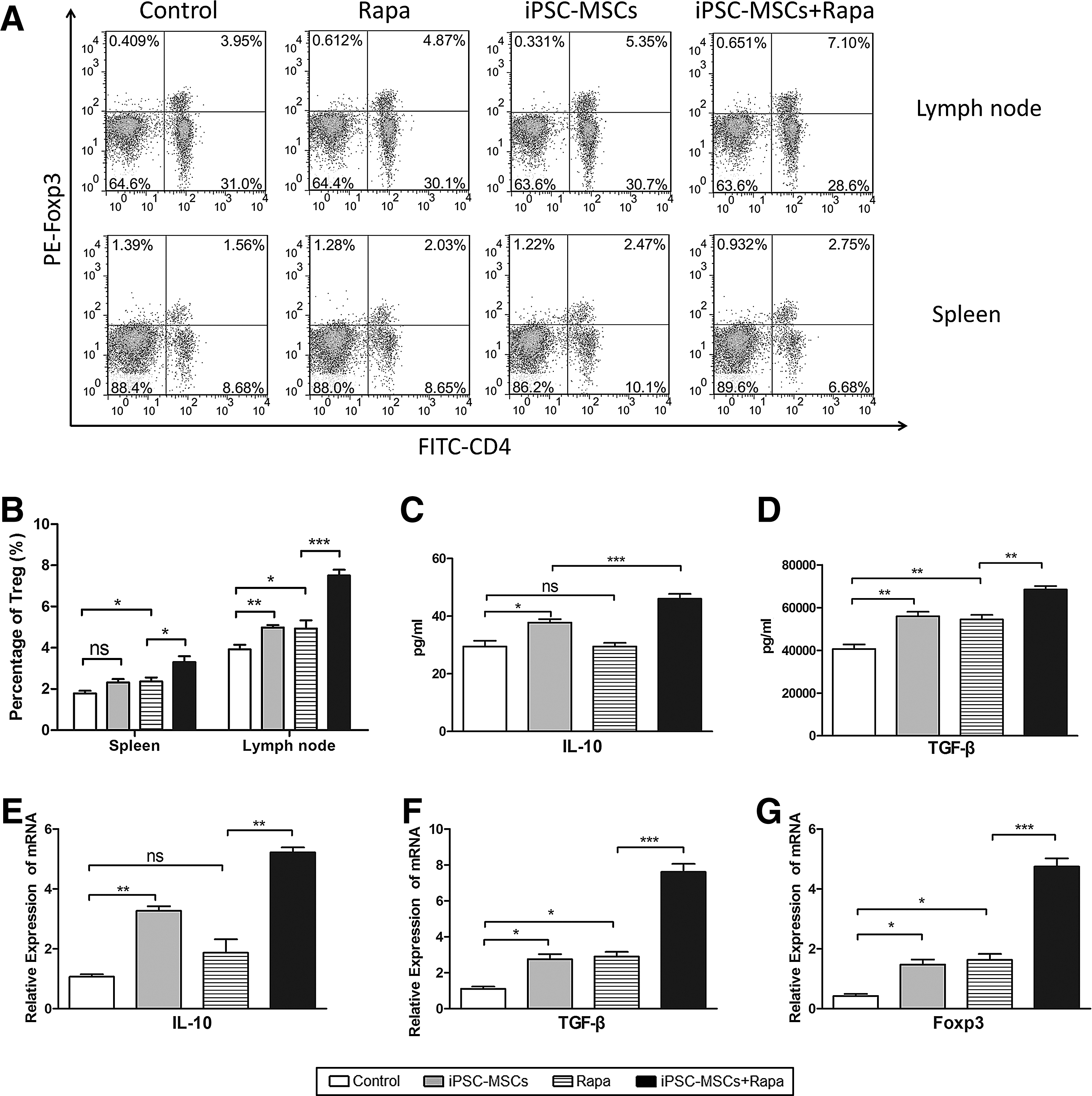
Protective effect of iPSC-MSCs+Rapa on islet allografts through T regulatory cell induction.
Immunosuppressive mechanisms of iPSC-MSCs+Rapa on T-lymphocyte proliferation and Treg production
T lymphocytes from the four groups were cocultured with mitomycin C-treated splenic lymphocytes from BALB/c mice in a 96-well plate for 3 days. The MLR test results indicated that recipient splenic T cells from combination-treated mice showed a reduced proliferative response when stimulated with mitomycin C-treated donor splenocytes compared with the Rapa group (P<0.01, Fig. 6A). In addition, IL-2, IFN-γ, IL-10, and TGF-β supernatant levels were determined using ELISA. iPSC-MSCs+Rapa treatment downregulated IL-2 and IFN-γ expression and upregulated IL-10 and TGF-β levels compared with the Rapa group (P<0.05, Fig. 6B–D). These results suggested that iPSC-MSCs+Rapa treatment suppressed Th1 function and increased Treg production.
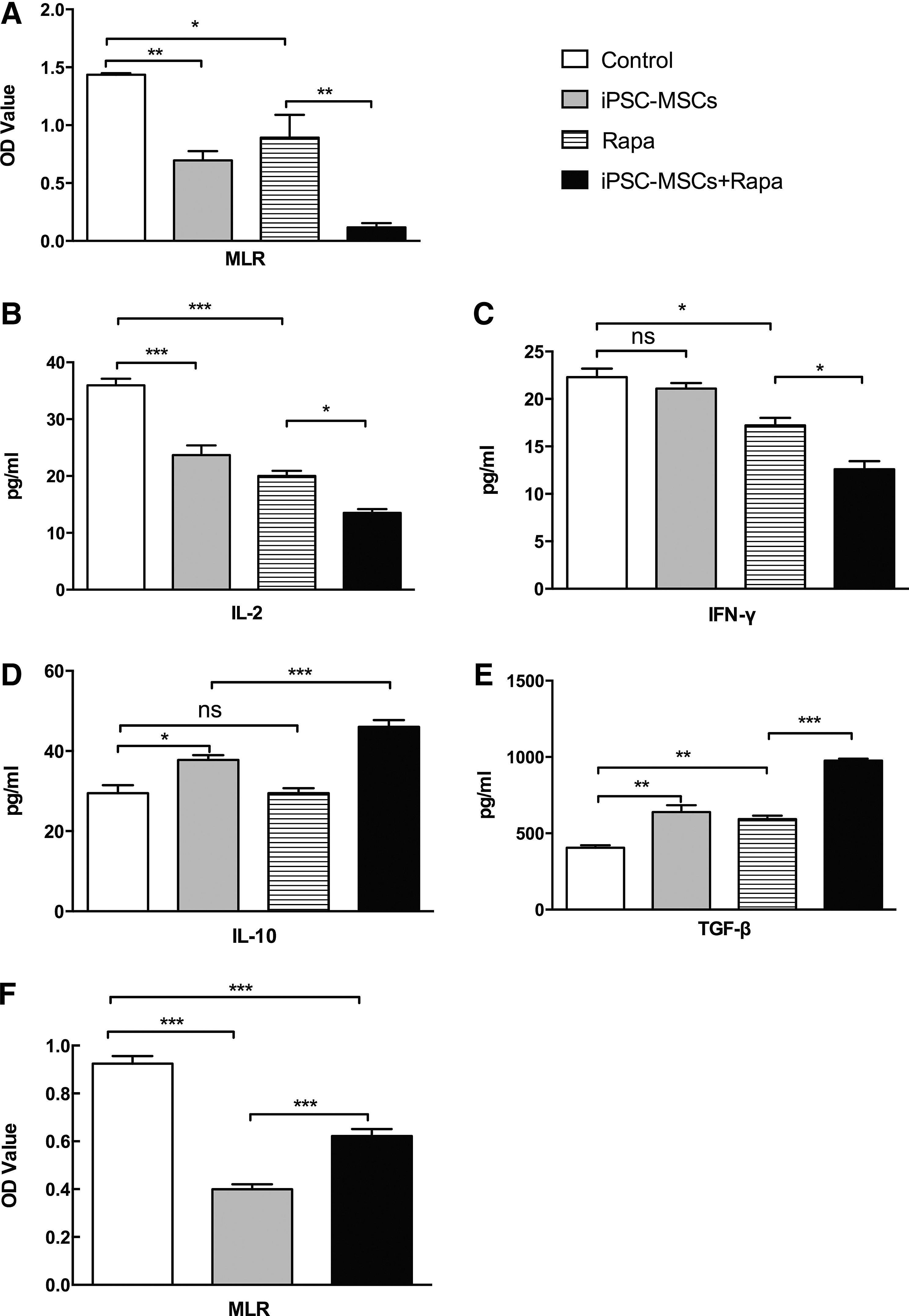
Immunosuppressive effect of iPSC-MSCs+Rapa on T-lymphocyte proliferation.
To evaluate the molecular mechanisms underlying these immunomodulatory properties, we further examined the possible role of cell contact in the modulation of T-cell proliferation by iPSC-MSCs+Rapa using Transwell experiments. As shown in Fig. 6F, iPSC-MSCs+Rapa significantly decreased the number of Con A-treated splenic T cells from normal mice. Moreover, Transwell separation significantly weakened the immunomodulatory effects of iPSC-MSCs+Rapa on Con A-treated splenic T cells. These findings suggested that cell contact could, at least partially, interfere with the immunomodulatory effects on lymphocyte proliferation under iPSC-MSCs+Rapa treatment.
Discussion
MSCs have previously demonstrated their capacity to facilitate regeneration and regulate immune responses in a range of animal models; however, major factors related to life span and tumorigenicity limit their widespread use in a clinical setting [28 –32]. Recent reports have described MSC-like cells derived from iPSCs [33 –35] with a greater proliferative capacity, lower immunogenicity, and greater immunoregulatory function compared with primary MSC cultures [33,43,45]. In addition, these iPSC-MSCs did not exhibit the tumorigenic properties associated with iPSCs [50,51], implying that iPSC-MSCs may be a safer MSC source. This study investigated the effects of iPSC-MSCs on islet allografts without using BM-MSC treatment as a control. Rapa is used frequently in islet transplantation; however, recent research showed evidence of Rapa toxicity in islet transplantation in clinical studies. For example, Rapa exerts dual effects on the islet endothelium by inhibiting angiogenesis and downregulating receptors that are involved in lymphocyte adhesion and activation [52]. Furthermore, Rapa also inhibits the revascularization of isolated pancreatic islets [53] and has significant detrimental effects on peripheral insulin resistance and β-cell function and survival [54]. Rapa at ≥0.5 mg/kg had detrimental effects on islet engraftment, while lowering the concentration to 0.1 mg/kg did not affect engraftment when tested for preventing rejection in the full mismatch allogeneic transplant BALB/c to the C57BL/6 model [55]. This result suggested that the detrimental effects of Rapa were dose dependent. Therefore, 0.1 mg/kg Rapa was chosen for use in our study. However, we showed that Rapa alone was inefficient in preventing rejection, which was consistent with published studies [55]. Thus, a combination of 0.1 mg/kg Rapa with iPSC-MSCs was used to suppress immune rejection of islet allografts in this study.
Our results showed that iPSC-MSCs had the morphological characteristics of adult MSCs. Previous published experimental studies suggested that MSCs can, under certain conditions, induce allograft tolerance together with immunomodulatory drugs [9,10,14,19]. In our study, iPSC-MSCs+Rapa treatment effectively prolonged islet allograft survival time and even induced immune tolerance in 50% of the recipients, which was consistent with results using adult MSCs with 2 mg/kg/day Rapa treatment in heart allografts [9]. Our results demonstrated that the combined treatment had synergistic effects.
Adult MSCs isolated from various sources (adipose tissue and Wharton's jelly) have been reported to equally suppress proliferation of CD4+ and CD8+ T-cell subsets in a dose-dependent manner [56]. Furthermore, adult MSCs inhibit Th1 and IFN-γ secretion in vitro [57,58]. Using a rat model of STZ-induced diabetes, adult MSCs significantly improved glycemic control and reduced inflammatory cell infiltration in either allogeneic or syngeneic pancreatic islet transplantation [59]. In this study, iPSC-MSCs+Rapa treatment effectively decreased the proportion of splenic and lymph node CD4+ and CD8+ T lymphocytes in vivo and strongly inhibited T-cell proliferation. We also found that IL-2 and IFN-γ expression was downregulated. These results suggested that iPSC-MSCs+Rapa treatment reduces Th1 inflammatory cytokines and may suppress the Th1 response.
Several studies have shown that adult MSCs alone, or combined with immunosuppressive drugs, induce allograft immune tolerance through Treg induction in vivo [8,9,17]. Berman et al. first reported that infusions of donor or third-party MSCs reversed rejection episodes and prolonged islet function, associated with increased numbers of Tregs in peripheral blood [60]. We hypothesized that iPSC-MSCs act like adult MSCs to induce Tregs. TGF-β is the perpetrator of immune suppression through regulatory T cells [61]. Although the in vitro dependency of Treg suppression on TGF-β is compelling, immune suppression mediated by Treg in vivo clearly requires TGF-β, because administration of an antibody against TGF-β blocked protection from colitis [62]. Moreover, in a type 1 diabetes model, CD8+ T cells bearing a dominant-negative TβRII transgene were incapable of responding to Treg suppression, resulting in diabetes progression [63]. Thus, we examined TGF-β expression in the islet grafts and sera of the recipient mice. The results showed that iPSC-MSCs+Rapa treatment increased sera TGF-β concentrations and graft TGF-β and Foxp3 mRNA levels, compared with Rapa treatment; this result supported our hypothesis.
Although a number of studies have revealed the immunosuppressive effects of MSCs, the mechanisms that modulate this process have not been fully explained. Generally, contact-dependent mechanisms and soluble factors, including 2,3-dioxygenase, prostaglandin-E2, nitric oxide, TGF-β, and hepatocyte growth factor, are thought to collaborate to induce MSC-mediated immunosuppressive effects [64,65]. Moreover, IL-10 has been reported to be involved in MSC-mediated immune regulation. Studies have demonstrated that addition of MSCs to MLRs increases IL-10 expression, while adding a neutralizing IL-10 antibody to MLRs results in recovery of the MLR response in long-term surviving splenocytes. This suggested that IL-10 mediates MSC suppressive capacity in autologous MSC+CsA-treated rats [17]. In this study, we used ELISA and qRT-PCR to measure IL-10 expression levels. Our results showed that iPSC-MSCs alone, or in the combination with Rapa, increased IL-10 secretion in sera and IL-10 mRNA levels in the graft, whereas Rapa alone did not. Further research is required to determine whether IL-10 mediates the immunosuppressive effect of iPSC-MSCs+Rapa on T cells. In addition, Transwell separation significantly weakened the immunosuppressive effects of iPSC-MSCs on the proliferation of Con A-treated splenic T cells, which indicated that the combined treatment exerts its immunosuppressive effects through cell–cell contact and the regulation of cytokine production.
It is worth mentioning that MSCs from the umbilical cord matrix, adipose tissue, and BM exhibit different capability to suppress peripheral blood B, natural killer, and T cells [66]. MSCs derived from the umbilical cord Wharton's Jelly displayed the most prominent immunosuppressive effects on phytohemagglutinin-induced T-cell proliferation, compared with MSCs derived from BM, adipose tissue, and the placenta [67]. Equine MSCs from solid tissue-derived sources, including the adipose tissue and umbilical cord tissue, inhibited T-cell proliferation by inducing lymphocyte apoptosis, while MSCs from BM and cord blood induced lymphocyte cell cycle arrest [68]. These studies showed that MSCs from different tissue sources possess different immunomodulatory effects and modulate immune cell function through overlapping and unique mechanisms [66 –68]. Thus, there may be differences in the immunosuppressive effects and mechanisms between iPSC-MSCs and MSCs from other sources, which require further study.
In conclusion, we have demonstrated that iPSC-MSCs combined with low-dose Rapa reduced the production of Th1 proinflammatory cytokines and significantly prolonged islet graft survival compared with iPSC-MSCs or Rapa alone. Furthermore, iPSC-MSCs alone, or combined with low-dose Rapa, induced IL-10 production in vivo and in vitro, which might have resulted from the immunomodulatory effects of iPSC-MSCs. In addition, iPSC-MSCs combined with low-dose Rapa significantly induced anti-inflammatory cytokines and Treg proliferation. The synergistic immunomodulatory effects of iPSC-MSCs and low-dose Rapa in islet transplantation suggest a promising strategy for preventing transplant rejection. This research also provides a preliminary experimental basis for applying MSCs not only in clinical islets but also in other solid organ transplants. However, there are still several questions that remain to be answered. Most importantly, the origin of the cell sources and the long-term effects of iPSC-MSCs need to be investigated to validate their safety and effectiveness in vivo. Finally, a more detailed understanding of iPSC-MSC functions should be determined in a transplant model.
Footnotes
Acknowledgments
This work was supported by grants from the Major State Scientific Research Program of China (no. 2012CBA01303), the National Natural Science Foundation of China (nos. 81302546, 31271038), the Xiamen Science and Technology Key Program Grant (no. 3502Z20100006), and partly from the Hong Kong Research Grant Council Collaborative Research Fund (HKU3/CRF/11R).
Author Disclosure Statement
No competing financial interests exist.