
Abstract
Human adult dental pulp tissue is a source of adult stem cells that have a potential to differentiate into various tissues, although the primary cell suspensions cultured from pulp tissue are mixtures of both stem cell and nonstem cell populations with heterogeneous phenotypes and various differentiation efficiencies. Therefore, cell surface protein markers on dental pulp stem cells are critical for detection and purification of stem cell populations. Yet, little is known about the cell surface molecules that are specifically associated with the undifferentiated and progenitor state of human adult dental pulp stem cells (hDPSCs). Presently, cell surface proteins expressed on hDPSCs were assessed by screening surface molecules specifically expressed on dentinogenic progenitors. Using a decoy immunization strategy, a set of monoclonal antibodies (MAbs) was generated against undifferentiated pulp progenitor cells. Forty-five hybridomas produced MAbs that interacted weakly, if at all, to differentiated pulp cells. Of these, 19 MAbs (18 IgG, 1 IgM) recognized surface molecules on undifferentiated hDPSCs. By multicolor flow cytometric analysis, 40%–60% of newly identified MAb-positive cells were demonstrated to be positive for the CD44 and CD90 mesenchymal markers. When MAb-positive cells were sorted from the heterogeneous pulp cell suspension, mineralization efficiency was increased three to five times compared with MAb-negative cells. The results suggest that the decoy immunization is an efficient method for isolation of MAbs against dentinogenic progenitors. These MAbs will be helpful for identification and enrichment of hDPSCs for efficient dentin regeneration.
Introduction
T
Materials and Methods
Cell culture
Discarded normal impacted third molars were collected from adult patients aged 20–25 years under guidelines approved by the IRB of the Dankook Dental Hospital. For human dental pulp cell culture, tooth was fractured and pulp tissue was separated from the crown and root. To primary cultures of periodontal ligament cells and gingival fibroblast, the periodontal ligament tissue and the spinous layer of gingival were collected, respectively. The pieces of tissue were digested in a solution of 3 mg/mL collagenase type I (Sigma-Aldrich) and 4 mg/mL dispase (Sigma-Aldrich) for 1 h at 37°C. Cell suspension was obtained by passing through a 70-μM strainer, and incubated in alpha-minimum essential medium (α-MEM) containing 20% fetal bovine serum (FBS) (Hyclone) and antibiotics at 37°C in 5% CO2. For differentiation into the mineralized phenotype, cells were incubated in α-MEM containing 10% FBS, 5 mM β-glycerophosphate (Sigma-Aldrich), 100 nM dexamethasone, and 100 μM ascorbic acid for 7–14 days. Commercial human bone marrow mesenchymal stem cells (Lonza, Inc.) were cultured in mesenchymal stem cell growth medium (MSCGM™; Lonza, Inc.) at 37°C in 5% CO2. For inhibition of N-linked glycosylation on cell surface, cells were treated with 1 μg/mL of tunicamycin (Sigma-Aldrich) for 24 h.
Production of MAbs against cell surface molecules of hDPSCs
Production of MAbs against cell surface molecules of hDPSCs were performed as previously mentioned with modification [20,21], and represented in Fig. 1A. Briefly, the right hind footpad and the left hind footpad of 10 female Balb/c mice (Koatech) were injected with differentiated and undifferentiated hDPSCs by turn, respectively (Fig. 1B). A total 1×106 cells/30 μL phosphate-buffered saline (PBS) (pH7.4) was injected in each shot without adjuvant. After total eight times of repeated alternate immunization, a lymphocyte suspension from the left popliteal lymph nodes, which are the injection sites of undifferentiated cells, was prepared and fused to FO myeloma cells (ATCC). Hybridomas were cultured in 96-well plates in Dulbecco's modified Eagle's medium supplemented with 20% FBS (HyClone)/HAT (Sigma-Aldrich). Animal work was approved by the Ethics review board of Dankook University and Sejong University. All animal procedures were performed as the guidelines from DKU-IACUC on the protection of animals used for scientific purposes.
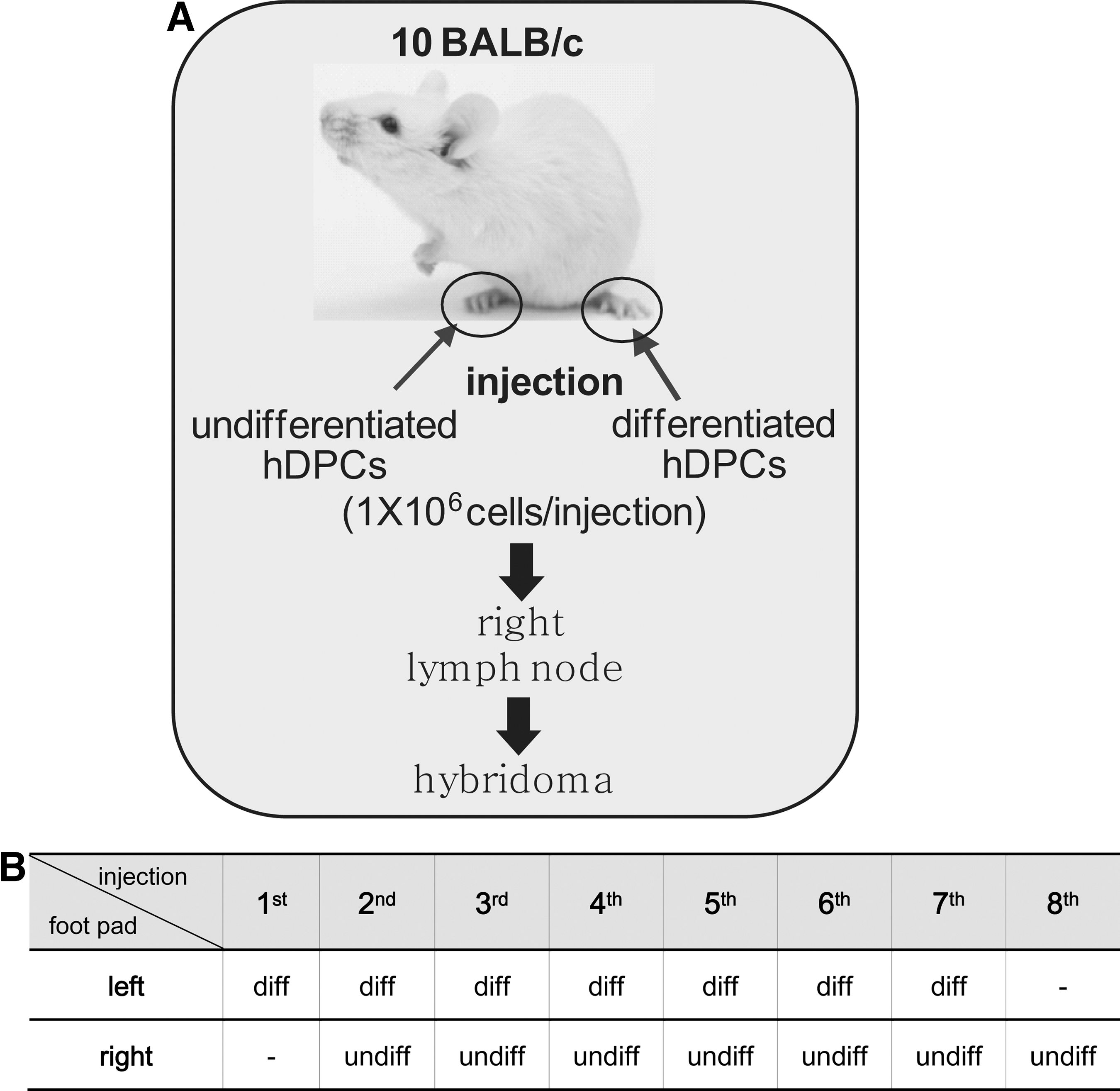
Schematic strategy of decoy immunization for immunomic screening of the undifferentiation-specific cell surface antibodies. Strategy of antibody screening
Immunophenotyping
Immunophenotyping experiment was performed as previously described with modification [9]. Cells were dissociated by using enzyme-free dissociation buffer (Millipore), harvested, and suspended in PBA (PBS containing 1% bovine serum albumin). The 1×105 cells/mL were treated with the proper momoclonal antibodies or CD antibodies conjugated phycoerythrin (PE). Anti-mouse IgG or -IgM was treated in this experiment as a negative control. For two-color flow cytometric analysis, cells incubated with appropriate MAb and FITC-conjuated anti-mouse IgG (Santa Cruz) were treated with biotinylated CD antibody and streptavidin-PE. After washing with PBA, cells were analyzed by flow cytometry in FACSCalibur system (BD Biosciences). The data were quantified by using Cell Quest. The values of mean fluorescence were calculated by using WinMDI 2.9 program.
Antibody isotyping
To characterize the immunoglobulin isotype of MAb, 96-well plate is coated by rat anti-mouse IgG1, -IgG2a, -IgG2b, -IgG3, -IgM, -IgA, -Igκ, and -Igλ. After blocking with PBS with 10% FBS, each hybridoma supernatant or purified MAbs was added. The reference antigen mixtures (BD Pharmingen) were used as positive control. Horseradish peroxidase-labeled rat anti-mouse immunoglobulins were added to each secondary antibody, and the isotyping signals were determined by optical density at 450 nm.
Immunohistochemistry
Cells were cultured on a cover-slip, and fixed in 4% paraformaldehyde at 4°C for 1 h. After blocking with 10% horse serum, cells were incubated with the primary antibody at 4°C overnight, followed by treatment with FITC-conjugated anti-mouse IgG. Nuclei were detected by staining with 4,6-diamidino-2-phenylindole (DAPI).
Biotinylation of antibody and intact cells
Purified MAbs were biotinylated with EZ-Link Sulfo-NHS-LC-Biotin Kit as described in the manufacturer's protocol (Thermo Scientific). Individual antibody dialyzed to 0.1 M NaHCO3 (pH 8.4) was incubated with biotin, followed by redialysis with PBS. For biotinylation of cell surface, cells were washed with ice-cold PBS (pH 7.4), and 1.0 mg Sulfo-NHS-LC-Biotin was added to 1×107 cells/mL in PBS.
Magnetic-activated cell sorting of MAb-positive cells
Human dental pulp cells (∼1×107 cells) were incubated with 1 μg of the biotinylated MAb at 4°C for 1 h, and after washing, were incubated with Streptavidin MicroBeads at 4°C for 1 h in dark. Cells labeled were isolated using magnetic-activated cell sorting (MACS) column system as described in manufacturer's guideline (Miltenyi Biotec). Cell viability was higher than 90% after sorting.
Alizarin red staining
After differentiation into a mineralizing phenotype, cells were fixed with 70% ethanol for 5 min at room temperature, and then allowed to dry. For staining, cells were treated with 2% Alizarin red S (pH 4.5), and washed thoroughly. To quantify mineralization, the deposits stained by Alizarin red were extracted with 10% acetic acid, and neutralized with 10% ammonium hydroxide, and the optical density was analyzed at a wavelength of 405 nm.
Quantitative real-time PCR
The expression levels of alkaline phosphatase, osteocalcin, osteonectin, dentin sialophosphoprotein (DSPP), and dentin matrix protein-1 (DMP-1) were analyzed by quantitative real-time PCR (qPCR). Briefly, total cellular RNA was isolated from cells harvested in triazole and used for synthesis of cDNA by using ReverTra Ace® qPCR RT kit (Toyobo Corporation). The qPCR was performed by using SsoFast Evagreen Supermix in CFX 384 (Bio-Rad) system. Primers used are listed in Table 1. The cycling parameters of qPCR were followed; 1 cycle for 1 min at 95°C, 45 cycles for 5 s at 95°C, and 1 cycle for 5 s at 58°C. After polymerase chain reaction (PCR), a dissociation curve was constructed in the range of 65°C–95°C. The glyceraldehyde 3-phophate dehydrogenase (GAPDH) used as an internal control to normalize the variability in expression levels. Relative quantity (ΔCq) for each gene is calculated with this formula: ΔCq=2[Cq(Min)−Cq(Sample)]. Cq(Min), average Cq for sample with the lowest average Cq for GOI; Cq(Sample), average Cq for sample; GOI, gene of interest. Normalized expression (ΔΔCq) is the relative quantity of each gene normalized to the quantities of GAPDH gene. The calculation for normalized expression is described in the following formula: ΔΔCq=ΔCq(Sample)/ΔCq(Ref).
GAPDH, glyceraldehyde 3-phophate dehydrogenase; DMP-1, dentin matrix protein-1; DSPP, dentin sialophosphoprotein.
Results
Generation of a set of cell surface-specific MAb against undifferentiated hDPSCs
hDPSCs were primarily cultured from dental pulp tissues extracted from third molars. Cells in passage number 3–5 were pooled and used for further study, since the dentinogenic potential peaks during these passages [9]. Differentiation of hDPSCs was induced by a 14-day treatment with dexamethasone, ascorbic acid, and β-glycerophosphate, and the undifferentiated state of the cells was confirmed by immunophenotyping using general MSC surface markers. CD44, CD90, and CD146 were highly expressed on the cell surface of undifferentiated hDPSCs (Fig. 2A, a–c in upper panels), and the expression of STRO-1 was slightly increased in undifferentiated cells (Fig. 2A, d in upper panels). After induction of differentiation for 14 days, these markers were dramatically downregulated (Fig. 2A, a–d in lower panels). CD24 and CD106, which are dental apical papilla stem cell representative surface markers [31,32] were not found on the surface of both undifferentiated and differentiated hDPSCs (Fig. 2e, f). For screening of the cell surface markers expressed specifically in undifferentiated hDPSCs, we generated a set of hybridoma secreting MAbs that recognized the cell surface of undifferentiated hDPSCs, but not differentiated cells, using an immunomic approach (Fig. 1) [20,21]. Briefly, differentiated intact hDPSCs were injected into the left hind footpads of 10 BALB/c mice as a decoy immunogen, followed 3 days later by immunization with undifferentiated hDPSCs in the right hind footpads of these mice. After eight times of repeated alternate immunization (Fig. 1B), the lymph nodes were removed from the right hind popliteal and the lymphocytes were fused to myeloma cells to generate hybridomas. A total of 245 hybridomas were primarily selected. To confirm whether the MAbs were specific for the undifferentiated cells, cell surface binding was investigated by flow cytometry. Twenty-six of the MAbs bound to differentiated hDPSCs more avidly than to undifferentiated cells (data not shown). Nineteen of the MAbs appeared to be more specific to the undifferentiated hDPSCs; of these 19, 7 clones bound to differentiated hDPSCs much more weakly than to undifferentiated cells.
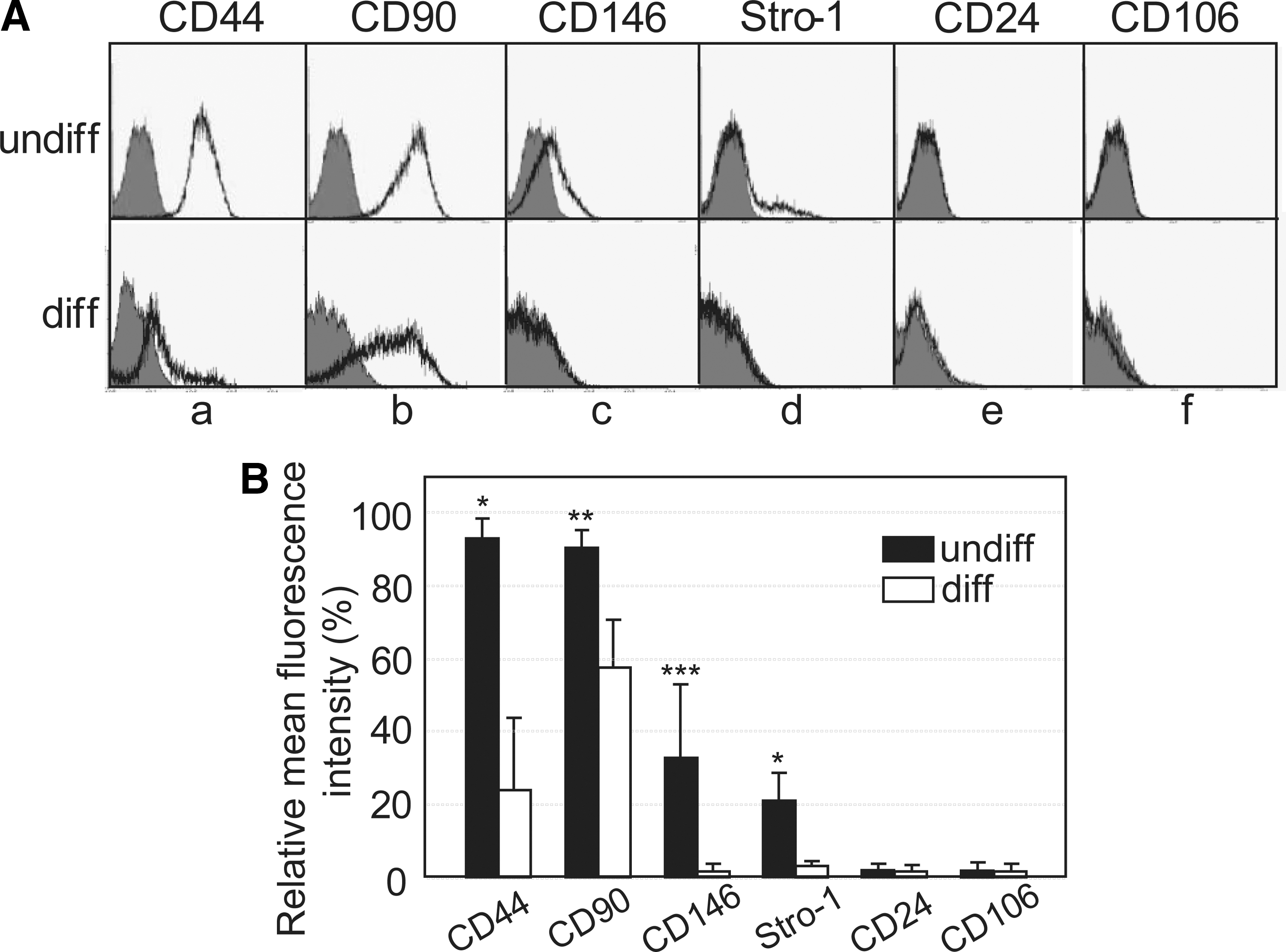
Expression of mesenchymal stem cell markers on the cell surface of the undifferentiated and differentiated human adult dental pulp stem cells (hDPSCs).
The binding affinities of the 3G12, 2C12, 1D12, 5B9, 6L4, 3A2, and 1G9 MAbs to the undifferentiated cell surface are depicted in Fig. 3A (panels 1–7). Twelve of 19 antibodies did not bind to differentiated cells, and they specifically interacted with the undifferentiated cell surface (Fig. 3A, panels 8–19).
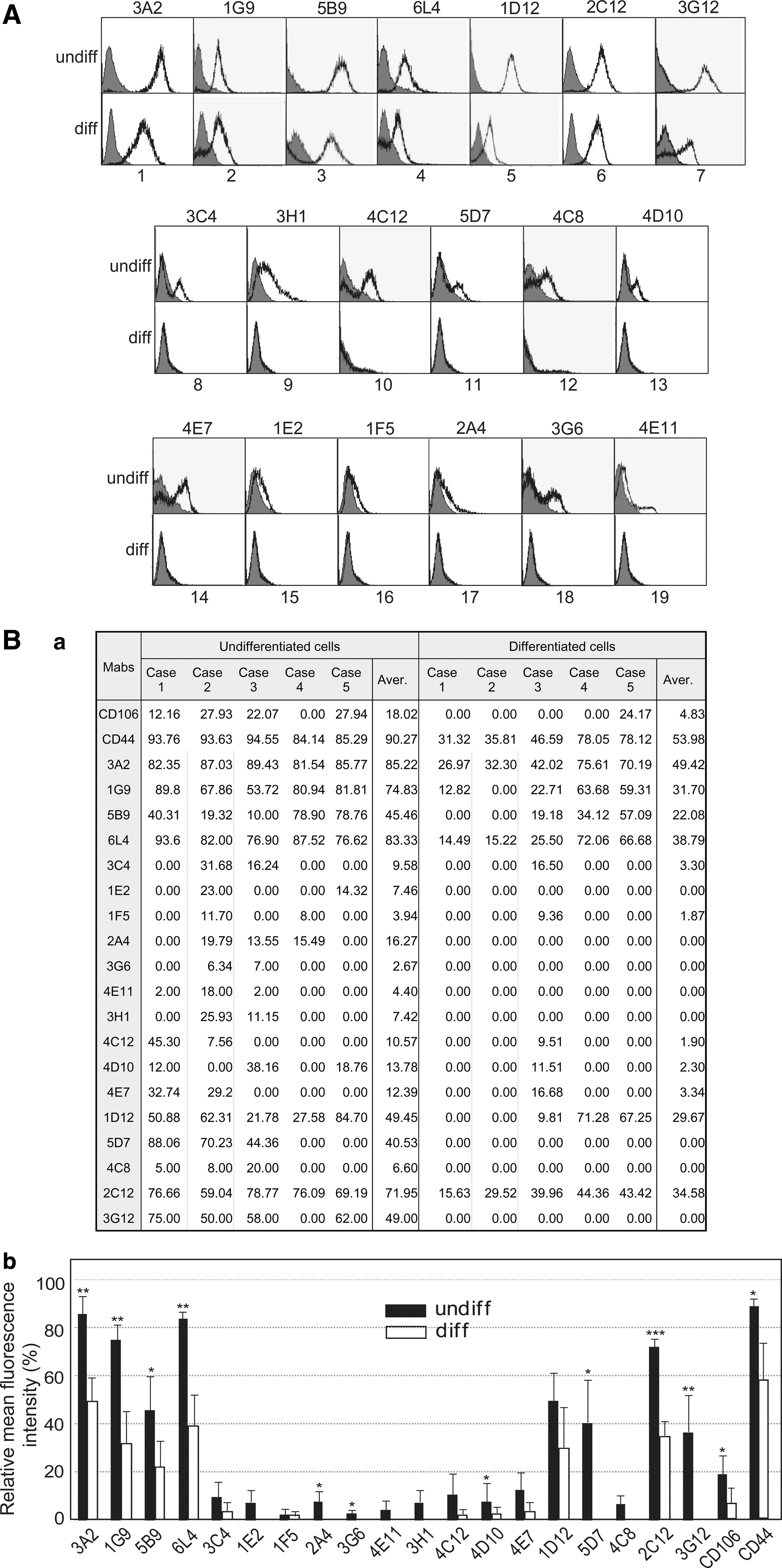
Undifferentiation-specific binding of 19 monoclonal antibodies (MAbs) on the cell surface of hDPSCs.
To investigate whether MAb binding specificity to the undifferentiated cell surface was consistent, hDPSCs cultured from pulp tissues of five different third molars were analyzed by flow cytometry. Most of the 19 MAbs bound to the surface of hDPSCs cultured from the individual cases, although differences in binding intensity were evident among the five pulp cell cultures (Fig. 3B, undifferentiated cells are shown in panel a). As expected, cell surface binding intensities were dramatically decreased in all five individual hDPSC cultures under the differentiated condition (Fig. 3B, differentiated cells are shown in panel a). Cell surface interaction of the 1E2, 1F5, 3G6, and 4D10 MAbs was individually relatively weak or was not detected in several culture cases of hDPSCs (Fig. 3B, a). As positive control, two MSC surface markers, CD44 and CD106, were consistently interacted strongly on the undifferentiated hDPSCs from the five individual culture (Fig. 3B, a). The statistical values of mean fluorescences from five individual cases were graphed, and the specificity for undifferentiation of the 19 MAbs was reaffirmed (Fig. 3B, b). Antibody isotyping analysis demonstrated that more than 96% of MAbs from hybridoma supernatant were IgG. The immunoglobulin subclasses of 19 MAbs are shown in Fig. 4; 18 were IgGs with the remaining MAb being IgM clone 4C8. Clone number 1G9 and 6L4 of the 18 IgG clones were the IgG2b type and 3G12 clone was the IgG3 type. All antibodies from the 19 hybridoma clones contained the kappa light chain (Fig. 4).
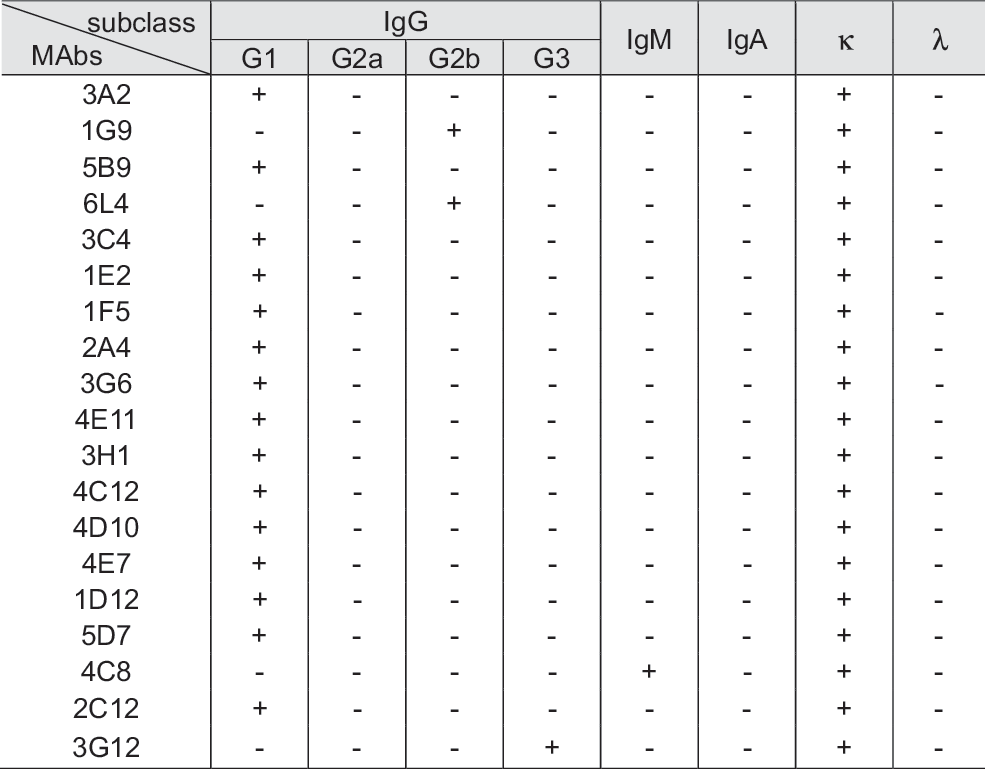
The isotypes of mouse MAbs (IgG1, IgG2a, IgG2b, IgG3, IgM, IgA, and kappa/lambda) derived from hybridoma supernatant of the representative 19 clones. Mouse IgG subtypes of each mAb are defined as (+). (-)s mean IgG subtype-negative of mAbs.
Binding affinity of a set of cell surface-specific MAbs on other human dental stem cells and bone marrow mesenchymal stem cells
Both periodontal ligament and gingival tissues in addition to dental pulp tissue are good sources of multipotent adult stem cells [33 –35]. To investigate the potentiality of the antigenic molecules recognized by these antibodies as proper markers of adult stem cells and progenitor cells, human periodontal ligament cells, human gingival fibroblasts (hGFs), and bone marrow mesenchymal stem cells (BMMSCs) were analyzed by antibody cell surface binding with the 19 MAbs. Stro-1 strongly interacted with the cell surface of hGFs, but less avidly with the surfaces of human periodontal ligament stem cells (hPDLSCs) and BMMSCs (Fig. 5, Stro-1). All tested cells were positive to five MAbs (3A2, 1G9, 6L4, 1D12, and 2C12) and the representative MSC markers CD44 and Stro-1. The 3C4, 3G6, and 3G12 MAbs bound specifically only to the surface of hDPSCs, while the 1E2, 2A4, and 4E11 MAbs interacted with the cell surface of hPDLSCs and hDPSCs (Figs. 3 and 5). Fourteen of 19 MAbs did not interact with the surface of BMMSCs (Fig. 5).

Cell surface binding of 19 MAbs in human gingival fibroblast (hGF), human periodontal ligament stem cells (hPDLSC), and human bone marrow mesenchymal stem cells (hBMMSCs). Based on FACS data, the mean fluorescence of each antibody–cell surface interaction was quantified and defined as (+). ++ +, over 60%; ++ , 30%–60%; +, below 30%; −, absent.
Preliminary identification of surface antigen molecules recognized by MAbs
Cell surface binding of MAbs was confirmed by immunocytochemistry. Because this article was written for the purpose of introducing a screening strategy of the cell surface markers of undifferentiated hDPSC, not for showing individual marker characterization, we actually chose eight representative MAbs, and performed further antigenic characterization in making match. As representative examples, the antigenic molecules recognized by the 3A2, 1D12, 5D7, and 2C12 MAbs, and CD44 were localized on cell membrane/cytoskeletal regions, whereas CD90, 1G9, 5B9, 6L4, and 4C12 antigens were partially localized in both the intracellular region and surface of the cells (Fig. 6A). To identify the cell surface antigens recognized by the MAbs, intact undifferentiated hDPSCs were biotinylated during cell culture and total labeled surface molecules were subjected to immunoprecipitation. The products of immunoprecipitation were analyzed by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and immunoblot analysis was done using horseradish peroxidase-conjugated streptavidin. Identification of molecular weight of antigens recognized by the representative eight MAbs is shown in Fig. 6B. Under the normal culture condition, the surface antigen molecules recognized by six MAbs (1D12, 1G9, 5D7, 2C12, 4C12, and 6L4) were detected as more than 180 kDa by SDS-PAGE with reduction of disulfide bonding (Fig. 6B, lanes 2–6 and 11 in a). In addition to these large molecular weight species, a constant 20-kDa molecule was detected from immunoprecipitants produced by 1D12 and 5D7 MAbs (Fig. 6B, lanes 2 and 4 in lower panel in a). The weight is not a precise determination, since lower molecular weights cannot be estimated under this condition. When cells were treated with tunicamycin during culture to inhibit N-linked glycosylation of whole cell surface proteins, the common signal of the 180 kDa molecules purified by several antibodies disappeared (Fig. 6B, lanes 3, 6 and 11 in b) or decreased (Fig. 6B, lanes 2, 4 and 5 in b). Additionally, the 20 kDa molecules became stronger or newly detected (Fig. 6B, lanes 2, 4, and 5 in lower panel in b). When cells were treated with tunicamycin, overall molecular weights of antigenic molecules obtained from immunoprecipitants became clearer and more specific according to MAb used. The 180 kDa molecules recognized by the 1G9 and 6L4 MAbs disappeared, and species ∼48 and 55 kDa were clearly detected upon tunicamycin treatment (Fig. 6B, lanes 3 and 11 in a and b). Additionally, a broad range of molecules between 45 and 55 kDa recognized by the 6L4 MAb were clearly detected as two bands (Fig. 6B, lane 11 in upper panel in b). The 5B9 MAb recognized two clear molecules between 100 and 135 kDa in size under the normal culture condition (Fig. 6B, lane 9 in a), whereas the molecular weights of these molecules were shifted down to between 40 and 48 kDa, when the glycosylation was inhibited (Fig. 6B, lane 9 in b). A 75 kDa surface molecule recognized by the 3A2 MAb was shifted down and detected with a molecular weight of ∼50 kD when cells were treated with tunicamycin (Fig. 6B, lanes 10 in a and b). The antigenic molecules of ∼180 kDa isolated by the 1D12, 5D7, and 4C12 MAbs were shifted down to 75 kDa by tunicamycin treatment (Fig. 6B, lanes 2, 4, and 5 in a and b). These data suggested that most of the cell surface molecules recognized by MAbs on undifferentiated hDPSCs possibly are modified by glycosylation. The general MSC markers, CD44, CD90, and STRO-1, were expressed on the cell surface of both undifferentiated and differentiated hDPSCs. Their expressions were dramatically decreased in the differentiated cells (Fig. 2, a–d in A and B) [9]. The molecular weights of CD44, CD90, and STRO-1 were known as 80–95, 25–37, and 75 kDa, respectively. In addition, CD73 (63–71 kDa) was reported to be expressed in hDPSCs in an expression pattern similar to other markers [35]. To verify whether the surface molecules recognized by MAbs were coincident with the these MSC markers, the immunoprecipitants produced using these MAbs were subjected to immunoblot analysis with CD44, CD73, CD90, and Stro-1 antibodies, and each showed a similar molecular weight range with surface molecules. The representative surface molecules that interacted with the 3A2, 1G9, 5B9, 6L4, and 2C12 MAbs did not show any cross-reactivity with CD44, CD73, CD90, and Stro-1 (data not shown).
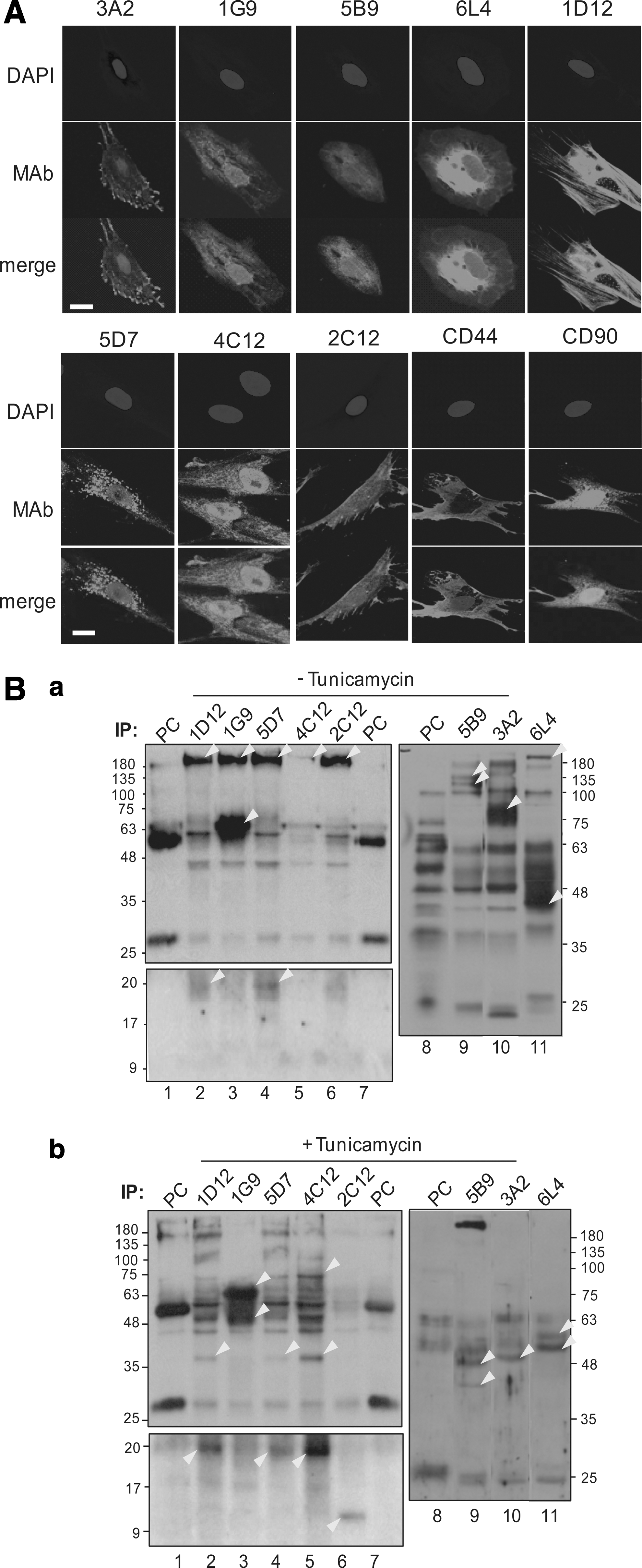
Characterization of cell surface molecules recognized by representative MAbs.
Expressions of the undifferentiated specific cell surface molecules are relevant to MSC-specific markers on hDPSCs
To investigate whether the expression of surface molecules recognized by 19 MAbs in the undifferentiated cells were related to other MSC markers, two-color flow cytometry was performed in undifferentiated hDPSCs. As representative examples, 1G9, 1D12, 3A2, 5B9, and 6L4 MAbs were detected by treatment with fluorescein isothiocynate-conjugated anti-mouse secondary antibody. To avoid cross-reactivity among the mouse primary and secondary antibodies used in this experiment, anti-CD44, anti-CD90, and anti-CD73 mouse antibodies were labeled using biotin and were detected by streptavidin conjugated with PE. As a result, 79.42%, 58.64%, and 75.94% of 1G9-positive hDPSCs co-expressed CD44, CD90, and CD73 on cell surfaces, respectively (Fig. 7, panels 2–4 in b). In addition, 80.77%, 46.08%, and 75.55% of cells were double-positive with 1D12 antibody and CD44, CD90, or CD73, respectively (Fig. 7, panels 2–4 in c). Most of the 3A2-, 6L4-, 4C12-, 2C12-, 5D7-, and 6L4-positive cells were also positive for CD44, CD90, and CD73 (Fig. 7, panels 2–4 in d and f–i). The expression of 5B9 antigen on the cell surface was relevant to the expression of three CD markers (Fig. 7, panels 2–4 in e), although cell population of double positive were relatively low (<40%) in case of the expression of CD90 (Fig. 7, panel 3 in e).
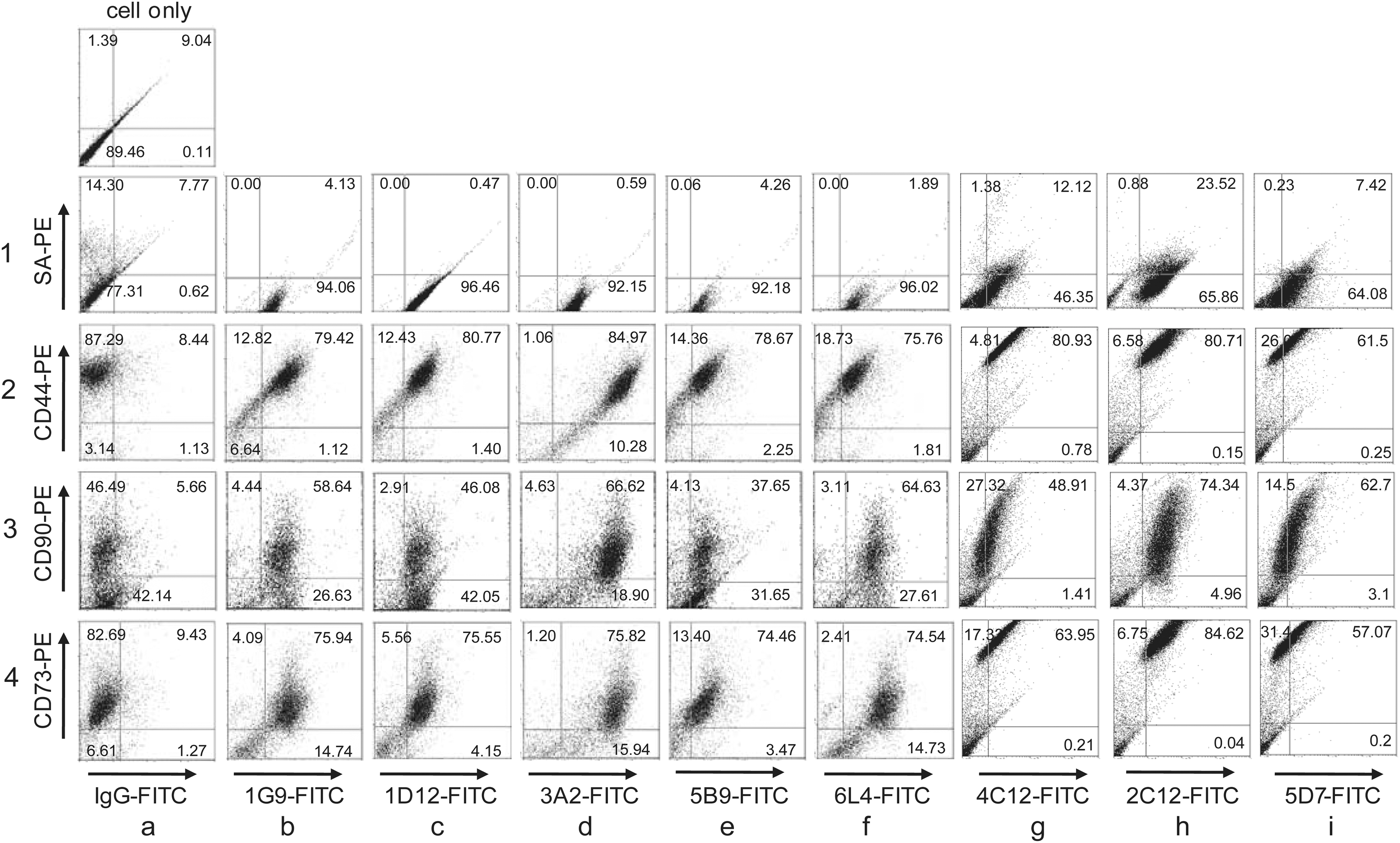
Expression correlation between antigenic molecules specific for the representative MAbs and hMSC-specific markers. The hDPSCs were incubated with one of the indicated MAbs, 1G9, 1D12, 3A2, 5B9, 6L4, 4C12, 2C12, or 5D7 and FITC-conjugated IgG antibody. Then, biotin-labeled CD44, CD73, or CD90 antibody and phycoerythrin (PE)-conjugated streptavidin were treated. Values in each quadrant indicated the percentage of positive cells on each antibody. Cell only, cells incubated without antibodies;
Mineralization efficiency was increased in MAb-positive hDPSCs
To investigate the differentiation power of MAb-positive hDPSCs, human dental pulp cells were separated by MACS. Two representative MAbs, 5B9 and 1G9, were biotinylated for the cell sorting, and cells bound to the labeled antibody were separated using streptavidin-conjugated magnetic beads. The MAb-positive cells were highly mineralized by treating with ingredients for induction of mineralization for 14 days (Fig. 8A, Ab-positive in upper panel). The mineralizations of 5B9-positive and 1G9-positive cells increased 5.1 times and 3.5 times as much as antibody-negative cell were mineralized, respectively (Fig. 8, α-5B9 and α-1G9 in A and B). Although the magnitude of differentiation of the sorted cells with antibodies compared with the unsorted cells was not so much increased (Fig. 8B, gray bars), the mineralized particles were highly concentrated in the sorted cell culture and the mineralization intensity were stronger than that of the unsorted cells (Fig. 8A, total). Again, the mineralization in the unsorted cells occurred in the whole surface of cells, even its intensity was weaker than that in the sorted cells (Fig. 5A, Ab-positive and total in α-5B9 and α-1G9). In addition to the increment of mineralization in antibody-positive cells, the expression of bone markers such as alkaline phosphatase, osteocalcin, and osteonectin during differentiation was increased in 1G9- and 5B9-positive cells (Fig. 8C, gray bars in graphs a–c). In addition to bone markers, the transcript levels of DSPP and DMP-1 were increased in 1G9- and 5B9-positive cells under the differentiating condition (Fig. 8C, gray bars in graphs d and e). These results indicated that the set of surface antibodies constructed from the undifferentiated hDPSCs can be useful for enrichment of DPSCs from heterogeneous cell populations for pulp tissue regeneration.
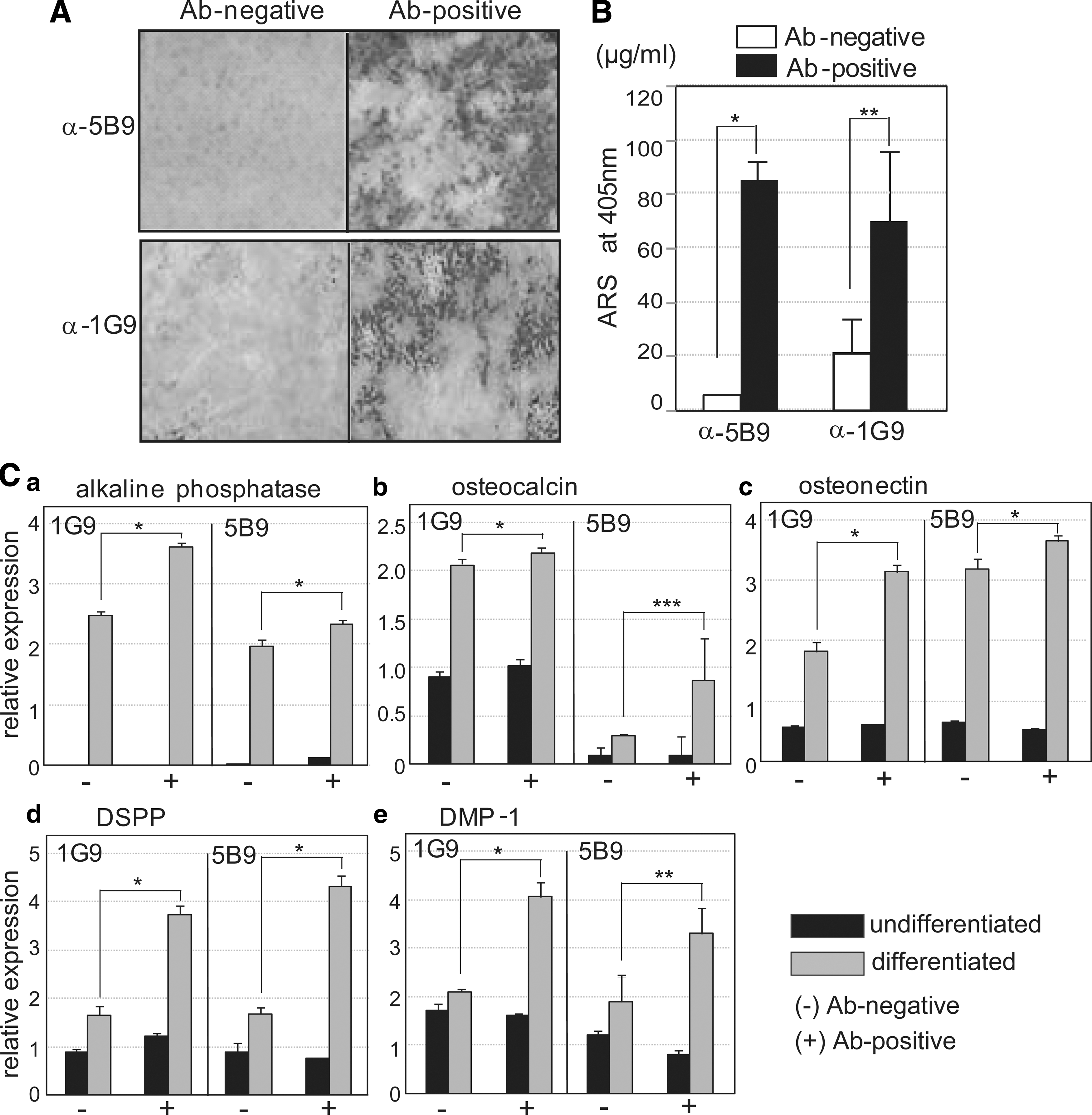
Osteogenic potentials were increased in the antibody-positive hDPSCs. To separate the MAb 5B9 or 1G9-positive cells from total hPDSC population, the magnetic-activated cell sorting analysis was performed. The antibody-positive cells (Ab-positive) and the remnant cells (Ab-negative) were cultured and differentiated. The mineralization by Alizarin-red staining was detected under the microscope
Discussion
For successful pulp/dentin regeneration, it is essential to isolate and to enrich the desired stem cells, which become odontoblasts and construct dentin in vivo. DPSCs are similar to BMMSCs in aspects including fibroblastic adherent morphology, multipotent capacity to differentiate, and the repair ability of tissues. However, the odontogenic potential of DPSCs and BMMSCs are not the same, although MSCs have been used to be a source of dentinogenic cells. Dental pulp cells are heterogeneous and show a mixed population of stem cells, progenitors, and nonstem cells. Moreover, unlike MSCs originating from bone marrow and other sources, DPSCs display an ectomesenchymal feature, which plays a critical role in the formation of the hard and soft tissues of the head and neck, such as teeth and bone. Although there is a difference between MSCs and DPSCs, the latter have been considered as MSC-like cells. Therefore, cell surface markers that have been used to identify MSCs are used for the isolation of DPSCs; these markers include CD44, CD73, CD90, CD146, and CD166 and STRO-1. In a practical manner, however, these positive markers, except for STRO-1, are not specific for undifferentiated stem cells/progenitors. These markers still interact with a large population of highly differentiated cells. In addition, the use of these markers for regeneration of pulp/dentin is controversial; whereas STRO-1+/CD146+ DPSCs form dentin/pulp-like structure [36], a subpopulation of CD31−/CD146− pulp cells also regenerate pulp tissue [37]. Moreover, c-kit+/CD34+/STRO-1+ dental pulp cells were reported as being multipotent stem cells, although c-kit+ and CD34+ are hematopoietic stem cell markers [38]. Under this situation, having markers for bona fide dental pulp stem cells would be important for tracking them during development, postnatal growth, and aging, and for applicative tissue engineering. In this report, to find more promising surface markers specific for undifferentiated DPSCs, we performed decoy immunization using intact human dental pulp cells. Although proteomic approaches, such as non gel-based and two-dimensional electrophoresis coupled with mass spectrometry, are well established for comparative investigation of cell surface molecules of various stem cells [39 –41], the decoy immunization approach has the advantage of using MAbs for direct recognition of surface molecules of undifferentiated DPSCs. In this report, we show that 19 MAbs obtained from 245 hybridomas specifically bound to undifferentiated hDPSCs. Because the stemness of human dental pulp primary culture might vary according to each individual patient and to the culture condition, the mean values of five individual cases were analyzed statistically. Indeed, the specificity for undifferentiation of the 19 MAbs was reaffirmed in five cases (Fig. 3B, b). In addition to usefulness of this set of MAbs for further molecular and biochemical studies on undifferentiated hDPSC-specific surface molecules and for isolation of the undifferentiated stem cells from of pulp primary culture, some of the MAbs were also useful for isolation of hPDLSCs and BMMSCs (Figs. 3 and 5). Most of all, the surface expressions of the presently identified antigenic molecules are relevant to the expression of MSC markers (Fig. 7), and they suggested that these surface antibodies might be applied for combinational treatment with known stem cell markers. By using MACS system, representative MAbs-positive hDPSCs increased mineralization potential (Fig. 8), indicated that this set of surface antibodies constructed from the undifferentiated hPDSCs can be useful for enrichment of DPSCs from heterogeneous cell populations for pulp tissue regeneration. In future, characterization and functional analysis of surface antigenic molecules may be very important for understanding renewal and differentiation mechanisms of hDPSCs. Ultimately, our study suggests that hDPSCs purified by these antibodies may be a better adult stem cell resource for pulp/dentin regeneration and tooth repair.
Footnotes
Acknowledgments
We are grateful to Dr. S. Shi at University of Pennsylvania for his critical reading and for having a discussion on this article. This work was supported by Bio & Medical Technology Development Program (2015M3A9C6029130) and Basic Science Research Program (NRF-2013R1A1A3007213) funded through the National Research Foundation of Korea.
Author Disclosure Statement
No competing financial interests exist.