
Abstract
The immunomodulatory properties of mesenchymal stem/stromal cells (MSCs) make them an attractive therapeutic tool to treat chronic inflammatory diseases, such as rheumatoid arthritis or Crohn's disease. These indications are characterized by a chronic activation of immune cells that perpetuates the disease. In vitro, when adipose mesenchymal stem cells (ASCs) are cultured with T lymphocytes at the time of stimulation, their proliferation is inhibited. However, these experimental settings do not necessarily fit with what ASCs will face in inflammatory conditions in vivo, where ASCs will likely encounter and interact with already activated immune cells which might affect their immunomodulatory capacity. In most in vitro studies, MSCs have been cultured with peripheral blood mononuclear cells at the time of lymphocyte stimulation and information about the interaction between MSCs and prestimulated lymphocytes in vitro is scarce. Therefore, a better understanding of the capacity of MSCs to modulate the responses of preactivated immune cells is needed. In this study we focused on the effects of ASCs on prestimulated lymphocytes and systematically investigated the potential mechanisms involved. We report that prestimulation of T lymphocytes 48 h before the coculture with ASCs impairs the capacity of ASCs to inhibit proliferation. Preactivation of ASCs with interferon γ or the toll-like receptor ligand Poly I:C, but not other stimuli tested, enhanced the ability to inhibit the proliferation of 48 h-stimulated T lymphocytes. The inhibitory effect of ASCs was shown to be time dependent and mediated through the actual magnitude of tryptophan degradation by indoleamine 2,3-dioxygenase.
Introduction
I
The immunomodulatory properties of MSCs rely not only on cell-to-cell interactions [12,13], but also on effects mediated by a variety of soluble factors [14]. MSCs show such immunomodulatory properties in response to inflammatory mediators released from activated immune cells such as interferon (IFN)-γ, interleukin (IL)-1β, or tumor necrosis factor (TNF)-α [15,16]. Importantly, indirect paracrine effects by regulating tryptophan (Trp) metabolism through the enzymatic activity of indoleamine 2,3-dioxygenase (IDO) have been proven to be of special relevance for the immunomodulatory properties of MSCs [17 –19].
We have previously reported that the capacity of ASCs to inhibit T lymphocyte proliferation in vitro requires the induction of IDO through their activation by IFN-γ released from activated lymphocytes [20]. Moreover, the effect of IDO activity on T lymphocyte inhibition depended on the depletion of Trp rather than on the accumulation of its catabolic product, kynurenine (Kyn) [21].
In those studies, T lymphocyte stimulation was done simultaneously to the culture with ASCs, so ASCs were activated concurrently with the lymphocytes. However, these experimental settings do not necessarily fit with what ASCs will face in inflammatory conditions in vivo (for instance, in patients with rheumatoid arthritis or mice with collagen-induced arthritis) where ASCs will likely encounter and interact with already activated immune cells (ie, lymphocytes) which might in turn affect their immunomodulatory capacity.
Therefore, to understand the effect of ASCs in this inflammatory milieu, we investigated the capacity of ASCs to inhibit the proliferation of prestimulated T lymphocytes in vitro. In this study, we report that prestimulation of T lymphocytes impairs in a time-dependent manner the in vitro capacity of ASCs to inhibit their proliferation. Preactivation of ASCs with IFNγ or the Toll-like receptor (TLR) ligand Poly I:C, but not other stimuli tested, enhanced the ability to inhibit the proliferation of 48 h-stimulated T lymphocytes. The rescue effect of IFNγ and Poly I:C was mediated by IDO activity and the degradation of Trp.
Materials and Methods
Isolation and cell culture
Human samples were obtained after informed consent as approved by the Spanish Ethics Committee of reference for the site of tissue procurement (Clínica de la Luz Hospital). Human adipose tissue aspirates from healthy donors were washed twice with phosphate-buffered saline and digested with 0.075% collagenase (Type I; Invitrogen). The digested sample was washed with 10% fetal bovine serum (FBS), treated with 160 mM NH4Cl to eliminate the remaining erythrocytes, and suspended in culture medium [Dulbecco's modified Eagle's medium (DMEM) with 10% FBS].
Cells were seeded (2–3×104cells/cm2) in tissue culture flasks and expanded (37°C, 5% CO2) with change of culture medium every 3–4 days. Cells were transferred to a new flask (103 cells/cm2) when they reached 90% confluence. Cells were expanded up to duplication 12–14 and frozen. Experiments were performed with cells from two male and two female adult donors at population doublings 12–14. ASCs were thawed from the same cryobanks and seeded before each experiment. ASCs were defined according to the criteria of the International Society for Cellular Therapy [22]: being positive for HLA-I, CD73, CD90, and CD105 and negative for CD11b, CD14, CD31, CD34, and CD45.
Peripheral blood mononuclear cells (PBMCs) were isolated from buffy coats using Ficoll-Paque PLUS (GE Healthcare Biosciences AB) following the supplier's protocol. Briefly, blood samples were diluted with balanced salt solution and Ficoll was added to create a density gradient. After centrifugation, we gently collected the interface containing mononuclear cells. Purity was verified by flow cytometry. Buffy coats were provided by the National Transfusion Center of the Comunidad Autónoma of Madrid, Spain.
Reagents and antibodies
5(6)-Carboxyfluorescein diacetate N-succinimidyl ester (CFSE),
IDO activity
IDO activity was measured by determining both Trp and Kyn concentrations on conditioned supernatants at different time points. A total of 100 μL of conditioned supernatants were mixed with 100 μL of 50 mM phosphate buffer and 25 μL of 2 M trichloroacetic acid. After centrifugation for 10 min at 15,600 g, 100 μL of the supernatant was analyzed by high-performance liquid chromatography in a Waters 717plus (Autosampler) using a Waters 1515 Isocratic Pump, a Waters 2487 Dual Absorbance Detector, and C18 4.6×50 mm column (Teknokroma Analítica S.A.) with 40 mM sodium citrate (pH 5.0)-1% acetonitrile as mobile phase at a flow rate of 1 mL/min. Trp and Kyn were detected at an absorbance of 280 and 360 nm, respectively.
IDO expression
ASCs were seeded in a six-well plate and were either left unstimulated or stimulated with IFN-γ (3 ng/mL) or Poly I:C (10 μg/mL). After 48 h of stimulation, cells were harvested and intracellularly stained with anti-IDO antibody using the kit for intracellular staining from eBioscience, following the manufacturer's instructions. A total of 10×103 events were acquired using a FACSCalibur (BD Bioscience). CellQuest Pro software was used for acquisition and analysis. CaliBRITE beads (BD Bioscience) were used before each assay to calibrate the cytometer.
IDO clones
ASCs overexpressing IDO (ASC-IDO+) or expressing a siRNA that downregulates IDO expression (ASC-IDOsi) were used in some experiments. These cells have been previously described [20]. Briefly, ASC-IDOsi cells were generated by cloning the following oligos: forward: ACTGGAACTGCCTCCTATT and reverse: AATGGAACTGCCTCCTATT into the pSUPER plasmid carrying the Pol III-dependent H1 promoter. Oligonucleotide sequences were validated at the RNAi Consortium and were purchased from Sigma-Aldrich. Then the H1-shRNA cassette was cloned in the pLVTHM plasmid using EcoRI-ClaI sites.
The IDO gene was amplified by polymerase chain reaction (forward: AACTCGAGATGCCACCATGGCACACGCTATGGAAAAC, reverse: ATGAATTCAATTAACCTTCCTTCAAAAGGGAT) and cloned in PRV-IRES-Neo using XhoI-EcoRI sites. Transfection and generation of lentiviral supernatants were performed using Polyethylenimine (PEI) (1 vol PEI:2 vol DNA). Viral supernatants were used for infection of ASCs.
Immunosuppression assay with ASCs
For CFSE labeling, PBMCs were washed extensively to remove FBS, resuspended in a 10 μM CFSE solution (107 PBMC per 200 μL of solution), and incubated with constant shaking (37°C, 10 min). Reaction was stopped with cold RPMI+10% FBS. Cells were washed and cultured overnight and one aliquot was used to set up and control FL-1 voltage for CFSE in the cytometer. After overnight resting, CFSE-labeled PBMC was left unstimulated or were activated with the Pan T Cell Activation Kit (microbeads coated with anti-CD3/CD2/CD28; Miltenyi Biotec) following the manufacturer's instructions.
PBMCs (106 cells/well) were cultured with ASCs (4×104 cells/well; ASC:PBMC ratio 1:25) at the time of stimulation or after 6, 24, or 48 h of stimulation in 24-well plates in a total volume of 1 mL of RPMI+10% FBS. The 1:25 ratio was chosen as it provided a high inhibitory effect, based on our previous studies [20]. Coculture was done in the PBMC-conditioned medium. After 120 h of stimulation, PBMCs were harvested, labeled with 7-AAD and anti-CD3 antibody, and cell proliferation of the CD3pos/7-AADneg population (viable CD3 T lymphocytes) was determined by flow cytometry according to loss of CFSE signal.
For experiments of rescue of the inhibitory effect of ASCs, PBMCs were stimulated as indicated above and, in parallel, ASCs were activated with different stimuli: IFN-γ (3, 0.3 or 0.03 ng/mL), TNF-α (20 ng/mL), IL-1β (20 ng/mL), LPS (10 μg/mL), Poly I:C (1 or 10 μg/mL), TGF-β (10 ng/mL), SDF-1α (150 ng/mL), and IL-17 (50 ng/mL). After 48 h, PBMCs were harvested, spun down (400 g, 5 min), and cocultured with ASCs (ASC:PBMC ratio 1:25) either in the 48 h-ASC-conditioned medium (containing the corresponding stimulus) or in fresh medium (containing the corresponding stimulus). This fresh medium was incubated in 24-well plates with the corresponding stimulus for 48 h in parallel to the preactivation of ASCs. Where indicated, ASC:PBMC ratios of 1:100 or 1:250 were used. Proliferation was determined at 120 h after PBMC stimulation.
In some experiments 200 μM Trp was added to the IFN-γ- or Poly I:C-activated ASC-conditioned medium at 48 h, right before the coculture with PBMCs. Trp was also added to the fresh medium of the control group (PBMC stimulated). Data were analyzed using FCS Express 4 software. CaliBRITE beads (BD Bioscience) were used before each assay to calibrate the cytometer.
Immunosuppression assay with ASC-conditioned supernatant
ASCs (or medium alone) were incubated with IFN-γ (3 ng/mL) for 48 h and supernatants were collected and spun down to remove cells or cell debris. Forty-eight hours-prestimulated PBMCs were cultured in the fresh medium with IFN-γ or in the conditioned medium from IFN-γ-preactivated ASCs. Proliferation was monitored as indicated above.
PBMC counts
PBMCs (106 cells) were stimulated as indicated above or were left unstimulated. After 48 h, cells were harvested and the total number of cells and viability were calculated using Nucleocounter YC-100 from ChemoMetec A/S, following the manufacturer's instructions.
T lymphocyte activation markers
PBMCs (106 cells) were stimulated as indicated above or were left unstimulated. After 48 h of stimulation, cells were harvested and stained with anti-CD3 and either anti-CD38 or anti-CD69 antibodies. A total of 10×103 events were acquired using a FACSCalibur (BD Bioscience). CellQuest Pro software was used for acquisition and analysis. CaliBRITE beads (BD Bioscience) were used before each assay to calibrate the cytometer.
Cytokine measurement
Concentrations of TNF-α, IL-10, piostaglandin E-2 (PGE-2), and human leukocyte antigen-G (HLA-G) in conditioned supernatants were determined by enzyme-linked immunosorbent assay (ELISA) at different time points following the manufacturer's instructions. ELISA for TNF-α was from ELISA Ready-SET-Go! (eBioscience), ELISA for IL-10 was from Affymetrix eBioscience, ELISA for sHLA-G (detecting HLA-G1/5) was from BioVendor Research and Diagnostic Products, and ELISA for PGE-2 was from Thermo Scientific.
Statistical analyses
Data represent mean±standard deviation. Statistical analyses were performed by applying the two-tailed Student's t-test. A value of P≤0.05 was considered statistically significant.
Results
The capacity of ASCs to inhibit proliferation depends on the duration of T lymphocyte prestimulation
To determine whether ASCs can inhibit the proliferation of prestimulated T lymphocytes, we stimulated PBMCs with microbeads coated with anti-CD2/CD3/CD28 antibodies for 6, 24, or 48 h. Then, PBMCs were either left alone or cultured with ASCs in the PBMC conditioned medium (to have an inflamed microenvironment and activate the ASCs). Proliferation of viable CD3 T cells was determined after 120 h of PBMC stimulation by flow cytometry.
Stimulated T lymphocytes proliferated up to generation seven (G7) (Fig. 1A). The capacity of ASCs to inhibit T cell proliferation varied depending on the length of T cell prestimulation. Thus, when ASCs were cultured with PBMCs at the time of stimulation or 6 h after stimulation, proliferation was inhibited after generation three (G3) and no differences were observed between the two time points of prestimulation. A reduction on the inhibitory effect of ASCs was observed when they were cultured with PBMC stimulated 24 h before (T lymphocytes proliferated up to G4). Strikingly, ASCs failed to inhibit T cell proliferation any longer when they were cultured with PBMCs prestimulated for 48 h.
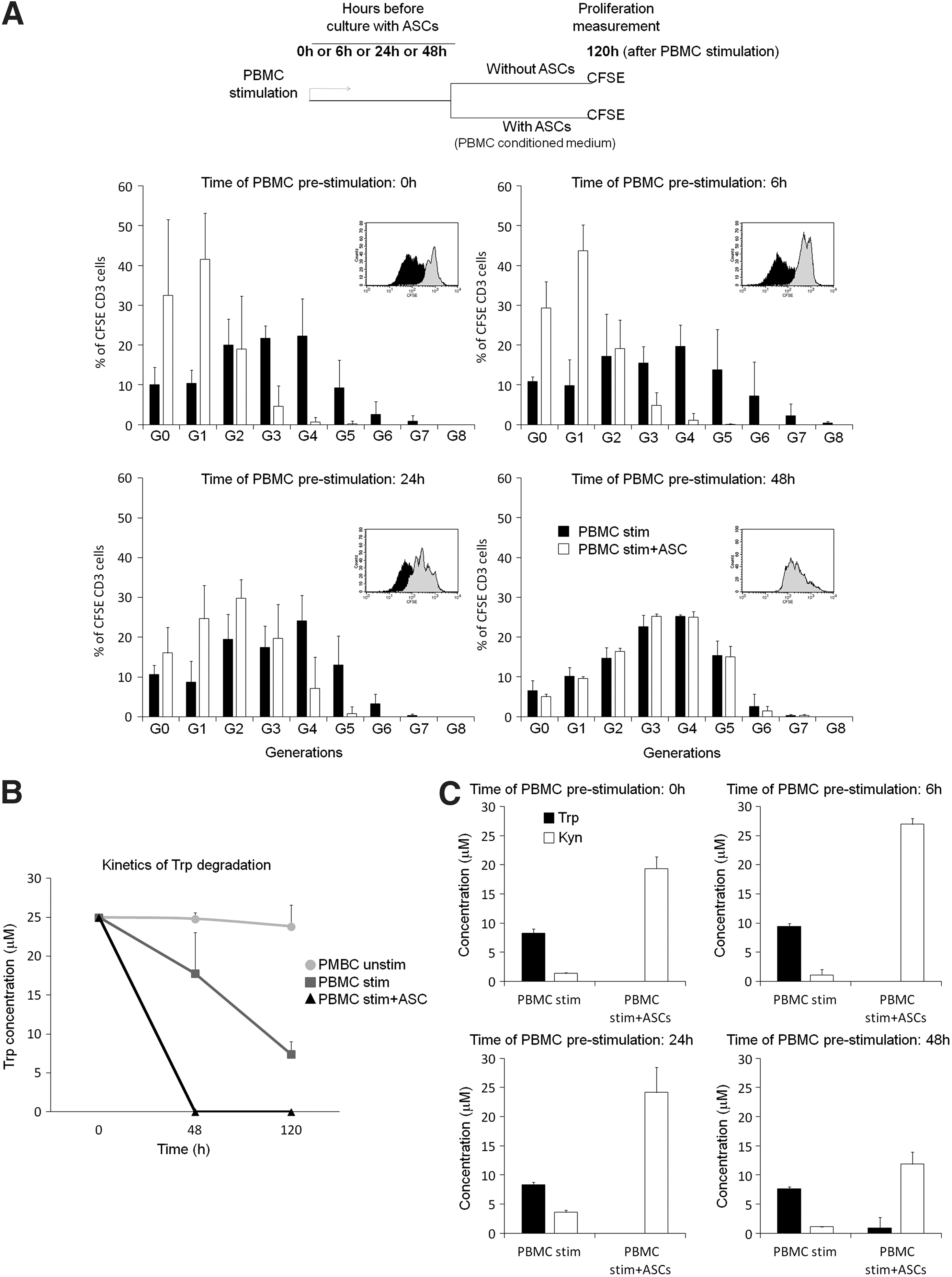
Effect of T lymphocyte prestimulation on the capacity of adipose mesenchymal stem cells (ASCs) to inhibit proliferation.
We have previously reported that, in our experimental conditions, Trp metabolism and IDO activity play a key role in the mechanism of inhibition of T cell proliferation by ASCs [20,21]. In fact, in our standard conditions, in which PBMCs and ASCs are cultured together at the time of PBMC stimulation, Trp is fully depleted already at 48h, whereas in stimulated PBMCs alone, the amount of Trp remains much higher (Fig. 1B).
To determine whether a diminishing of IDO activity could be implicated in the lower capacity of ASCs to inhibit proliferation of prestimulated lymphocytes, Trp and Kyn concentrations, were determined at 120 h under different conditions. In the presence of ASCs, Trp was virtually fully depleted in all the conditions regardless of the time of lymphocyte prestimulation and Kyn was highly accumulated in all the conditions, although to a lesser extent when PBMC:ASC coculture was done at 48 h (Fig. 1C), which is likely due to a higher consumption of Trp by the proliferating lymphocytes (Fig. 1B) and, therefore, lower levels of Trp available to be degraded by IDO activity.
These results suggested that the deficiency of ASCs to inhibit proliferation of 48 h-prestimulated lymphocytes is not due to a restriction on IDO activity, but rather to the kinetics of Trp degradation and the levels of Trp in the medium at 48 h.
We next hypothesized that the impairment of ASCs to inhibit the proliferation of 48 h-stimulated T lymphocytes could be a consequence of facing very actively proliferating lymphocytes at the time of coculture, which could make them difficult to stop.
To investigate this possibility, the activation and proliferation status of 48 h-stimulated T lymphocytes were determined. The activation status of T cells was determined by the expression levels of the activation markers CD38 and CD69 and by the release of TNF-α in unstimulated and stimulated PBMCs at 48 h. Our results indicated that after 48 h of stimulation, T lymphocytes were highly activated as indicated by the upregulation of CD38 and CD69 expression and the accumulation of TNF-α in the medium (Fig. 2A, B), but they were not actively proliferating as indicated by the CFSE staining and cell counts (Fig. 2C, D). These results suggested that the observed incapacity of ASCs to inhibit T lymphocyte proliferation is not related to their proliferation status.

Activation and proliferation status of stimulated T lymphocytes at 48 h.
Preactivation of ASCs with IFN-γ or Poly I:C restores their capacity to inhibit T cell proliferation
Proinflammatory cytokines (ie, IFN-γ, TNF-α, and IL-1β) and TLR ligands (LPS, Poly I:C) have been reported to modulate the immunoregulatory capacity of ASCs [15,16,23]. In addition to these stimuli, we found that TGF-β, SDF-1α, and IL-17 can also activate ASCs, as determined by their effects modulating the release of IL-6, IL-8, monocyte chemoattract ant protein-1, or vascular endothelial growth factor by ASCs (data not shown).
To determine whether preactivation of ASCs with these cytokines or TLR ligands could enhance their capacity to inhibit the proliferation of 48 h prestimulated T lymphocytes, PBMCs were stimulated and, in parallel, ASCs were activated with IFN-γ (3 ng/mL), TNF-α (20 ng/mL), IL-1β (20 ng/mL), TGF-β (10 ng/mL), SDF-1α (150 ng/mL), IL-17 (50 ng/mL), LPS (10 μg/mL), or Poly I:C (10 μg/mL). After 48 h, PBMCs and preactivated ASCs were cocultured together, either in the ASC-conditioned medium or in fresh medium. As a control, PBMCs were cultured with non-preactivated ASCs.
Preactivation with IFN-γ and, to a lesser extent, Poly I:C (but not with TNF-α, IL-1β, TGF-β, SDF-1α, IL-17, or LPS) allowed ASCs to inhibit proliferation of 48 h-prestimulated lymphocytes (Fig. 3A and data not shown). This inhibitory effect was only observed when coculture was done in the conditioned medium from preactivated ASCs, but not when PBMCs and preactivated ASCs were cultured in fresh medium (data not shown).
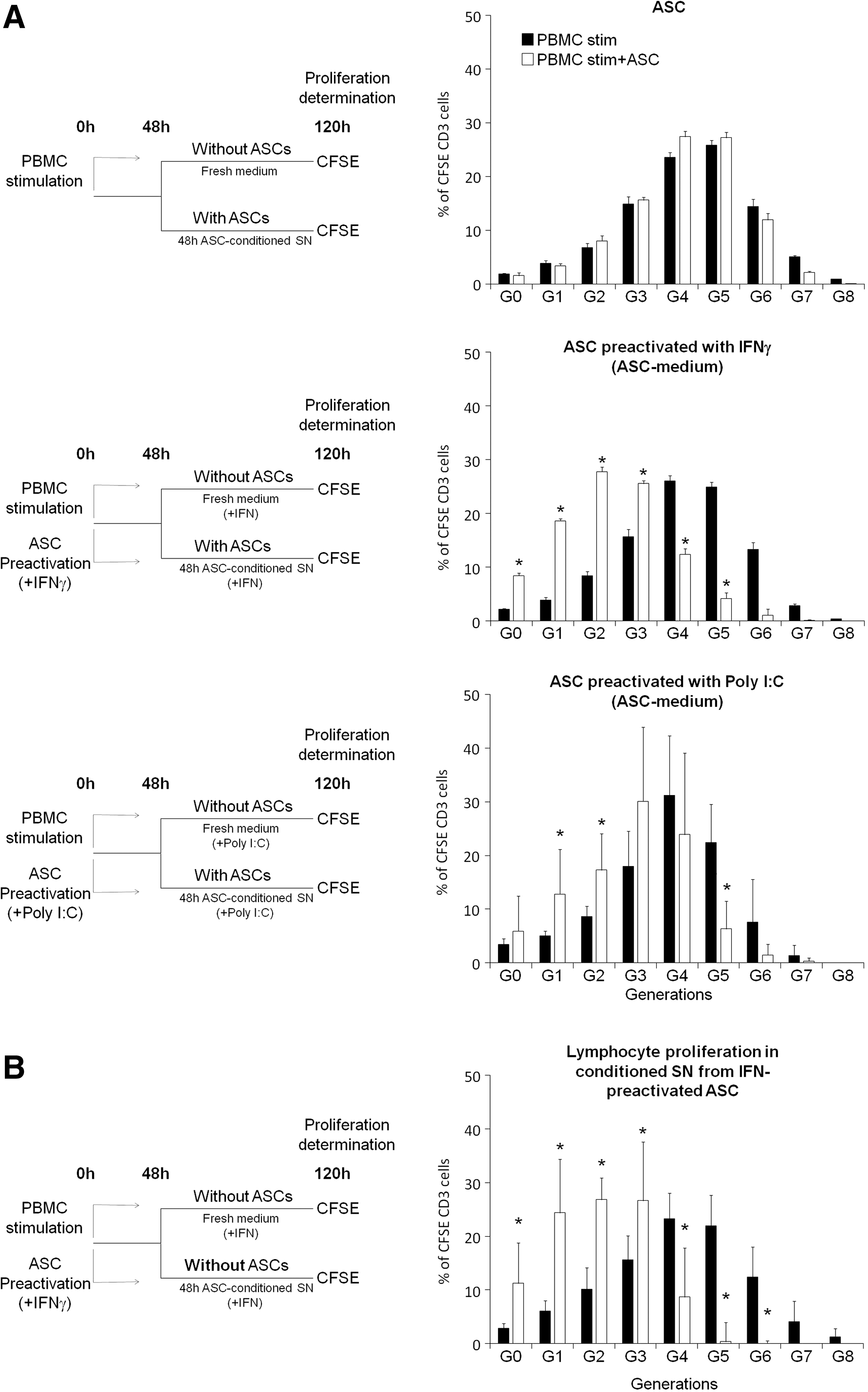
Preactivation with interferon (IFN)-γ or Poly I:C restores the capacity of ASCs to inhibit proliferation of 48 h-prestimulated lymphocytes.
To further confirm the importance of the conditioned supernatant, 48 h-prestimulated PBMCs were cultured only with conditioned supernatant from ASCs preactivated 48 h with IFNγ. As shown in Fig. 3B, the supernatant from IFN-preactivated ASCs was sufficient enough to inhibit the proliferation of 48 h-prestimulated lymphocytes.
We next investigated whether IFN-γ and Poly I:C effects were dose dependent or ASC:PBMC ratio dependent. Therefore, we carried out experiments using 0.3 or 0.03 ng/mL of IFN-γ and 1 μg/mL of Poly I:C, at a ratio 1:25 (ASC:PBMC). As shown in Fig. 4A, 0.3 ng/mL of IFN-γ had similar recovery effect than 3 ng/mL, but at 0.03 ng/mL the inhibitory effect of ASCs was very low. When 1 μg/mL of Poly I:C was used, ASCs had no longer effect on the proliferation of T lymphocytes (data not shown).
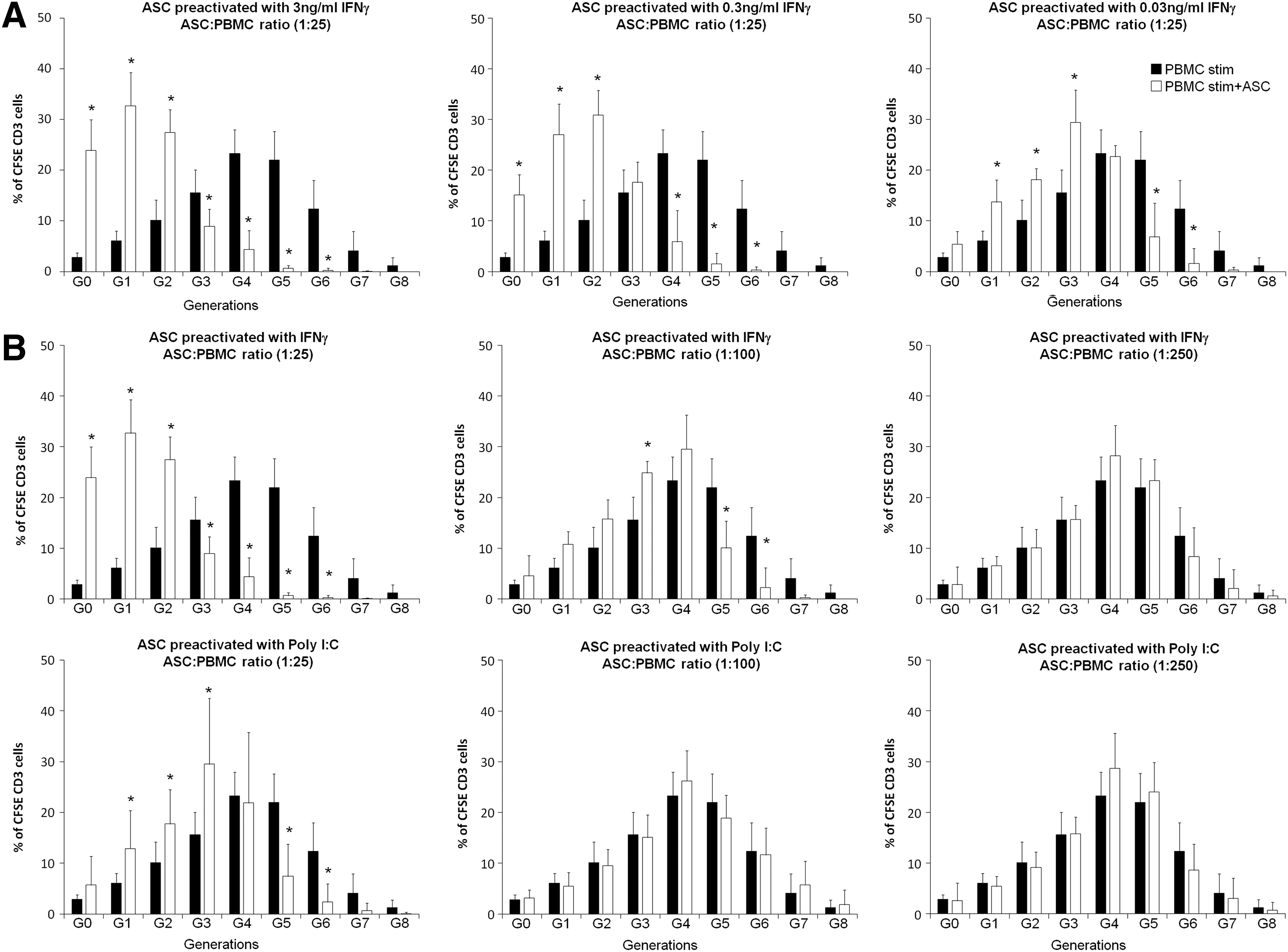
Dose and ASC:PBMC ratio dependency of IFN-γ and Poly I:C effects.
We then investigated whether IFN-γ and Poly I:C effects were ASC:PBMC ratio dependent. To do so, 48 h-prestimulated lymphocytes were cultured with IFN-γ (3 ng/mL) or Poly I:C (10 μg/mL) preactivated ASCs at ASC:PBMC ratios of 1:100 or 1:250 and the capacity to restore the inhibitory effect of ASCs was analyzed. As shown in Fig. 4B, the effect of IFN-γ was drastically reduced at the ratio 1:100 and completely lost at 1:250. The effect of Poly I:C was fully lost at 1:100 ratio.
Altogether, these results indicated that preactivation of ASCs with IFN-γ, and to a lesser extent Poly I:C, acts as an enhancer of the immunomodulatory capacity of ASCs and restores the capacity of ASCs to inhibit the proliferation of 48 h-prestimulated lymphocytes, through a mechanism that required the conditioning of the medium.
IDO activity mediates the ability of IFN-γ and Poly I:C to restore the capacity of ASCs to inhibit T lymphocyte proliferation
To determine whether the induction of IDO activity was involved in the effects of IFN-γ and Poly I:C, we measured the concentrations of Trp and Kyn in the conditioned supernatant from ASCs preactivated with IFN-γ or Poly I:C for 48 h.
As shown in Fig. 5A, while IFN-γ stimulation led to a complete depletion of Trp and high accumulation of Kyn, Poly I:C stimulation resulted only in a partial conversion of Trp into Kyn. These effects in Trp degradation correlated with the intracellular expression levels of IDO in IFN-γ or Poly I:C preactivated ASCs, because IFN-γ induced a higher expression of IDO compared to Poly I:C (mean fluorescence intensity IFN-γ: 56.58 vs. 26.56 of Poly I:C, Fig. 5B).
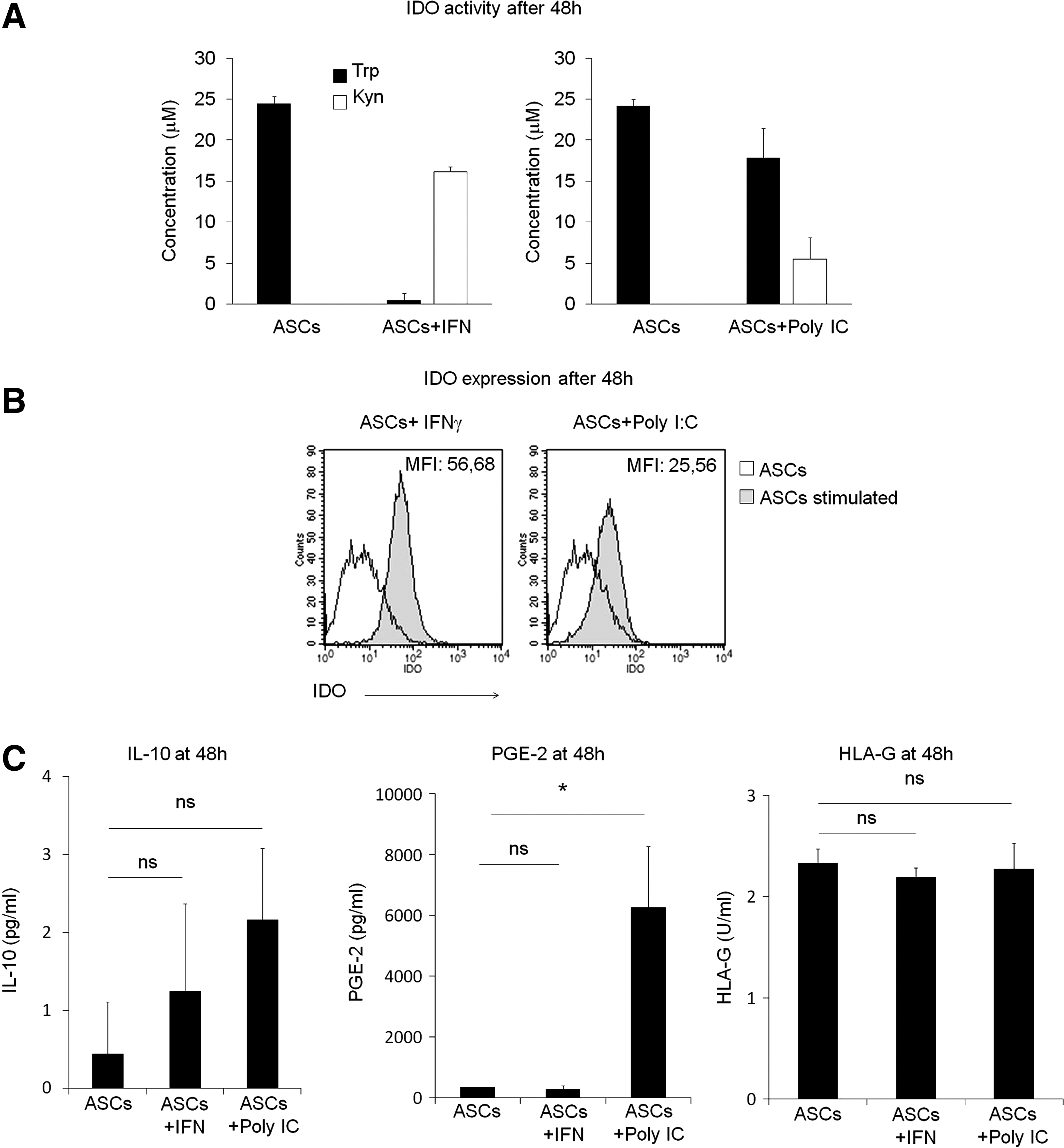
Indoleamine 2,3-dioxygenase (IDO) expression and activity correlates with the restoring effects of IFN-γ and Poly I:C.
In addition, the levels of IL-10, PGE-2, and HLA-G, immunomodulatory molecules also involved in the inhibitory effects of MSCs, were measured in these conditioned supernatants. As shown in Fig. 5C, IFN-γ did not significantly affect the expression of any of these molecules. Activation with Poly I:C did not affect the expression of IL-10 and HLA-G, but did increase the release of PGE-2 (Fig. 5C).
Our results suggested that Trp depletion by IDO activity might mediate the ability of IFN-γ and Poly I:C to restore the antiproliferative effects of ASCs, at least in our experimental settings. To confirm this possibility, Trp (200 μM) was added to the conditioned media at 48 h, right before the coculture of IFN-γ or Poly I:C preactivated ASCs with prestimulated PBMCs. As shown in Fig. 6A, Trp supplementation abolished the inhibitory effect of both IFN-γ and Poly I:C preactivation.
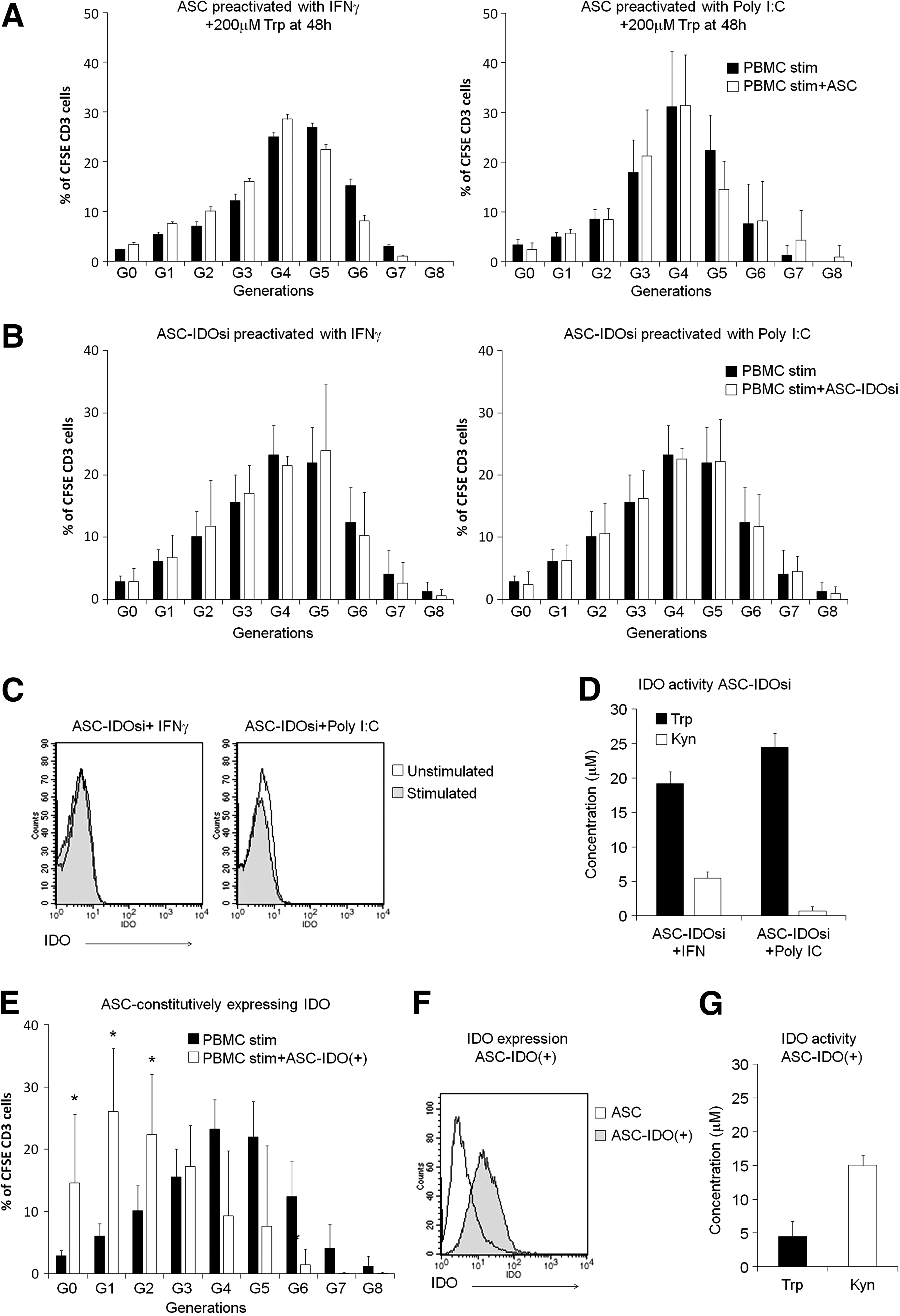
IDO activity mediates the effect of IFN-γ and Poly I:C.
The requirement of IDO activity was further confirmed by using ASCs that either express a siRNA that knock down IDO expression (ASC-IDOsi) or that constitutively express the protein without any activation (ASC-IDO+) [20].
When ASC-IDOsi was preactivated with either IFN-γ or Poly I:C, T cell proliferation could not be inhibited (Fig. 6B). As expected, after 48 h of stimulation IDO expression and activity were highly inhibited (Fig. 6C, D).
Notably, when prestimulated PBMCs were cocultured with ASC-IDO+ (not preactivated) T lymphocyte proliferation was inhibited. This effect was only observed when coculture was done in the ASC-conditioned medium (Fig. 6E), but not in fresh medium (data not shown). As expected, IDO expression and activity were high in these cells at the time of coculture and levels of Trp were already very low (Fig. 6F, G).
Altogether, these results confirm that Trp depletion through the induction of IDO activity plays an important role in the capacity of ASCs to inhibit T lymphocyte proliferation.
Discussion
The immunomodulatory properties of MSCs make them an attractive therapeutic tool to treat chronic inflammatory diseases. These indications are characterized by a chronic activation of immune cells that perpetuates the disease [1,2]. When MSCs are infused in patients with an inflammatory disease they will likely encounter and interact with immune cells that are already in an activated mode. Therefore, a better understanding of the capacity of MSCs to modulate the responses of preactivated immune cells is needed.
In most in vitro studies, MSCs have been cultured with PBMCs at the time of lymphocyte stimulation and information about the interaction between MSCs and prestimulated lymphocytes in vitro is scarce.
In this study, to reproduce an inflammatory environmental condition such as ASCs could encounter in patients, we investigated the capacity of ASCs to inhibit the proliferation of prestimulated T lymphocytes. To our knowledge, this is the first study that focuses on the effects of ASCs on prestimulated lymphocytes and systematically investigated the potential mechanisms involved. Our study is limited to lymphocytes and the interaction with other prestimulated immune cells (macrophages, dendritic cells, B lymphocytes, and NK cells) should also be investigated.
Our results indicate that the capacity of ASCs to inhibit T cell proliferation depends on the duration of T cell prestimulation. Thus, when ASCs were cultured with T lymphocytes prestimulated for 48 h, ASCs could not inhibit T cell proliferation.
Previously other authors reported already some results using prestimulated lymphocytes and MSCs. In contrast to our results, Yang et al. reported that mouse MSCs were able to suppress proliferation of prestimulated mouse splenocytes when cocultured after 36 h of mixed lymphocyte reaction [24]. These results are in apparent contradiction to those presented here, which could be explained by differences in the experimental settings: mouse MSCs and splenocytes versus human ASCs and PBMCs, different stimulus to trigger lymphocyte proliferation (mixed lymphocyte reaction vs. anti-CD2/CD3/CD28 beads), and time of MSC:lymphocyte coculture (36 h vs. 48 h).
On the other hand Valencic et al. reported that coculture of human Wharton's jelly MSCs with PBMCs prestimulated 24 h with phytohaemagglutinin (PHA) failed to inhibit PBMC proliferation [25]. When MSCs were preactivated with conditioned medium from stimulated PBMCs, the inhibitory effect of MSCs was restored. The difference with our study (impairment of MSC inhibitory effect after 24 h vs. 48 h), might be due to the use of different MSCs (Wharton's jelly vs. ASCs) and/or different stimulus to activate PBMCs (PHA vs. anti-CD3/CD2/CD28). Neither of these studies, however, further investigated the mechanisms underlying these observations.
We found that T lymphocytes were activated after 48 h of stimulation, but we did not yet observe proliferation (Fig. 2), indicating that the difficulties of ASCs to inhibit proliferation were not related to high proliferation rate of lymphocytes at the time of coculture. This delay between the time of stimulation and detection of proliferation has been previously reported by others and seems to be related to the dynamics of the sequential phases of lymphocyte activation, proliferation being the last one [26,27].
Importantly, preactivation of ASCs for 48 h with IFN-γ and, to a lesser extent, with Poly I:C (but not with TNF-α, IL-1β, TGF-β, SDF-1α, IL-17, or LPS) allowed ASCs to inhibit the proliferation of 48 h-prestimulated lymphocytes if the coculture was done in the medium conditioned by preactivated ASCs, indicating that conditioning of the medium was necessary. Remarkably, among all the stimuli tested, only IFN-γ and Poly I:C induced IDO activity (Fig. 5).
Indeed, a correlation between the level of IDO expression and Trp depletion at 48 h in the conditioned supernatants of preactivated ASCs and the level of inhibition of T cell proliferation was observed. Thus, IFN-γ, which showed to be a stronger enhancer of the inhibitory capacity of ASCs than Poly I:C, fully depleted Trp in the conditioned supernatants at 48 h, while Poly I:C did it only partially. Interestingly, the results obtained with IFN-γ mimic the kinetics of Trp degradation when PBMCs and ASCs are cocultured at the time of T lymphocyte activation in which Trp is fully consumed at 48 h, before lymphocytes have even started to proliferate.
The role of IDO and Trp depletion was further confirmed by using three approaches: (1) supplementation of Trp to the conditioned supernatant before PBMC:ASC coculture; (2) by using ASCs deficient in IDO expression (ASC-IDOsi); and (3) by using ASCs constitutively expressing IDO (ASC-IDO+). These results show that both IFN-γ and Poly I:C recuperate the capacity of ASCs to inhibit proliferation of prestimulated lymphocytes through a mechanism that requires the induction of IDO activity. Poly I:C also increased the production by ASCs of the immunomodulatory molecule PGE-2, which in our experimental settings seems to play, if any, a secondary role compared to IDO.
The results we report in this study, on the enhancer effect on the immunomodulatory properties of ASCs by IFN-γ preactivation, are in agreement with Polchert et al. and Duijvestein et al. that reported superior efficacy of IFN-γ-pretreated MSCs in animal models of graft versus host disease and colitis, respectively [28,29]. The authors and others have reported that the licensing effects of IFN-γ are mediated through both IDO-dependent and IDO-independent mechanisms [17,20,21,30,31].
In agreement to our results, the effects of IFN-γ in improving the capacity of MSCs to inhibit lymphocyte proliferation are IDO dependent [17,30,31], while the effects on the capacity of MSCs to inhibit effector function (eg, cytokine production) by lymphocytes is IDO independent [31]. It should be noted that in contrast to the results reported here, in these studies lymphocytes were not stimulated before the coculture with MSCs, but at the same time.
Poly I:C activation has been reported to drive MSCs toward an anti-inflammatory phenotype [32]. However, inconsistent results regarding the effect of Poly I:C on the capacity of ASCs to inhibit lymphocyte proliferation have been reported in recent years, and either none [33 –35], increasing [36] or decreasing [12,37] effects, has been reported. Differences in the experimental conditions likely explain the observed inconsistencies in vitro. Interestingly, preactivation with Poly I:C has been reported to enhance the therapeutic effect of MSCs in mouse models of cardiac repair, peripheral neuropathy, or sepsis [38 –40] suggesting that pretreatment of MSCs with Poly I:C may, indeed, enhance their therapeutic effects.
Additionally, synergistic/combinatorial effects between cytokines should also be considered as IL-17 that did not enhance the inhibitory effects of ASCs in our experiments, has been recently reported to enhance the immunosuppressive effect of MSCs activated by IFN-γ +TNF-α [41].
Our results show that there is a time window in which ASCs can inhibit proliferation of prestimulated lymphocytes and suggest that the kinetics of Trp depletion by IDO drives the efficient inhibition of T cell proliferation by ASCs, which requires a timely coordination between Trp depletion and initiation of T lymphocyte proliferation.
Indeed, the level of inhibitory effect seemed to be related to the degree of Trp depletion at 48 h. Thus, in conditions where IDO activity/expression was high and Trp was fully depleted at 48 h (ASCs cocultured with PBMCs at the time of lymphocyte stimulation or 48 h IFNγ preactivated ASCs cocultured with 48 h-prestimulated PBMCs) the inhibitory effect was very strong. However, in conditions where IDO activity/expression was lower due to delayed or suboptimal ASC activation (ASCs cocultured with 24 h-prestimulated PBMCs or 48 h Poly I:C preactivated ASCs cocultured with 48 h-prestimulated PBMCs) only a partial inhibitory effect was found.
In summary, our results indicate that preactivation of ASCs with IFN-γ or Poly I:C improves the capacity of ASCs to efficiently inhibit the proliferation of prestimulated lymphocytes. Such priming with the appropriate stimuli might also occur in inflammatory environments in vivo. Further investigation will be needed to confirm that preactivation of MSCs could be an alternative approach to enhance their therapeutic effects in clinical settings.
Footnotes
Acknowledgments
The authors thank Debjani Roy (TiGenix NV) for critical reading of the article. This project has received funding from the Spanish Ministerio de Economía y Competitividad (MINECO) and Comunidad Autónoma de Madrid (CAM) through Madrid Network to TiGenix, and the European Union's Seventh Program for research, technological development, and demonstration under grant agreement no 279174 to TiGenix and Erasmus Medical Center.
Author Disclosure Statement
P.M.-C., O.D.R., C.R., B.d.R., R.M., W.D., and E.L. are full-time employees of TiGenix.