
Abstract
The major reservoir of human multipotent mesenchymal stem/stromal cells (MSCs) is the bone marrow (BM) with the capability to control hematopoietic stem cell development. The regenerative potential of MSCs is associated with enhanced endogenous repair and healing mechanisms that modulate inflammatory responses. Our previous results revealed that MSC-like (MSCl) cells derived from pluripotent human embryonic stem cells resemble BM-derived MSCs in morphology, phenotype, and differentiating potential. In this study, we investigated the effects of MSCl cells on the phenotype and functions of dendritic cells (DCs). To assess how antiviral immune responses could be regulated by intracellular pattern recognition receptors of DCs in the presence of MSCl cells, we activated DCs with the specific ligands of retinoic acid-inducible gene-I (RIG-I) helicases and found that activated DCs cocultured with MSCl cells exhibited reduced expression of CD1a and CD83 cell surface molecules serving as phenotypic indicators of DC differentiation and activation, respectively. However, RIG-I-mediated stimulation of DCs through specific ligands in the presence of MSCl cells resulted in significantly higher expression of the costimulatory molecules, CD80 and CD86, than in the presence of BM-MSCs. In line with these results, the concentration of IL-6, IL-10, and CXCL8 was increased in the supernatant of the DC-MSCl cocultures, while the secretion of TNF-α, CXCL10, IL-12, and IFNγ was reduced. Furthermore, the concerted action of mechanisms involved in the regulation of DC migration resulted in the blockade of cell migration, indicating altered DC functionality mediated by MSCl cell-derived signals and mechanisms resulting in a suppressive microenvironment.
Introduction
M
MSCs are known to modulate immune responses through acting on multiple cell types, such as DCs, natural killer cells, and T and B lymphocytes [8,20], and were shown to suppress the differentiation, activation, migration, and antigen-presenting functions of conventional DCs [20], a cell population essential for linking innate and adaptive immune mechanisms through presenting antigenic peptides to naive T lymphocytes. This series of events can induce cell expansion and polarization of effector T lymphocytes to Th1, Th2, Th17, and Treg directions [21]. It has also been demonstrated that BM-derived MSCs can shift the differentiation of naive CD4+ T cells to an anti-inflammatory Th2 direction [22], underpinning the relevance of this type of regulation for utilization in clinical settings exemplified by BM transplantation, contact allergy, and autoimmune disorders [23]. Conventional DCs continuously binding and internalizing antigens use a wide spectrum of pattern recognition receptors (PRRs). Upon sensing endogenous and exogenous danger signals, DCs become activated and migrate into the draining lymph nodes where they present their antigenic cargo to naïve T lymphocytes. Retinoic acid-inducible gene-I (RIG-I) and melanoma differentiation-associated protein 5 (MDA5) are cytoplasmic PRRs expressed by a wide array of cell types and play essential roles in recognizing and eliminating viruses. DCs activated by specific ligands (ssRNA, dsRNA, or DNA) of the RIG-I receptors results in the secretion of proinflammatory cytokines and chemokines [24]. RIG-I is also expressed and is functional in MSCs and has an important role in supporting cell survival, indicating its possible contribution to the regulation of MSC functional activities [25]. Human MSCs express several TLRs, which are involved in the regulation of cell migration and the production of immunomodulatory factors. The functional properties of human MSCs were suggested to depend on the type of TLR as TLR4-primed MSCs gave rise to inflammatory MSC1, while TLR3-induced MSCs to suppressive MSC2 cell populations [26].
Encounter of conventional DCs with naïve T lymphocytes depends on the timely migration of activated DCs to the draining lymph nodes [27,28] driven by the CCR7 chemokine receptor expressed by stimulated DCs as well as by the expression of the CCR7 chemokine receptor ligands, CCL19 and/or CCL21 [28]. This receptor–ligand interaction upregulates the expression of matrix metalloproteinases (MMPs) responsible for the degradation and remodeling of the extracellular matrix (ECM) upon cell migration [29], while the proteolytic activity of MMPs is regulated by the tissue inhibitors of MMP (TIMP) enzymes. Maintaining the balance between these opposing activities MMPs and TIMPs are crucial for preventing uncontrolled enzymatic degradation of the ECM known to contribute to the initiation of inflammation, autoimmune disorders, and cancer metastasis [30].
In a previous work, we have characterized the expression patterns of MMP enzymes and TIMP inhibitors in monocytes and monocyte-derived DC subpopulations [31] and discovered that the chemokine-driven migration of activated DCs is regulated by the expression of the voltage-gated sodium channel, Nav1.7 [32]. Based on these findings, in this study, we aimed to investigate the effects of MSCl cells on DC activation triggered by ligand-specific RIG-I stimulation. Our results demonstrated for the first time that MSCl cells are able to modify the activation status of the DC, the secretion levels of cytokines and chemokines, as well as the outcome of DC-mediated T-cell polarization, suggesting their profound impact on the outcome of DC functionality. We also propose that MSCl cells could be used as a feasible model for human in vitro studies by offering means for developing novel MSC-based strategies to design directed immunosuppressive cellular therapies.
Materials and Methods
Generation of monocyte-derived DCs
Monocytes were separated from peripheral blood mononuclear cells (PBMCs) of healthy blood donors drawn at the Regional Blood Center of the Hungarian National Blood Transfusion Service, in accordance with the written approval of the Director of the National Blood Transfusion Service according to the directives of the European Union. PBMCs were separated by Ficoll Paque Plus (GE Healthcare Bio-Sciences AB, Uppsala, Sweden) gradient centrifugation, followed by positive selection of CD14+ monocytes by using anti-CD14-coated magnetic beads (Miltenyi Biotec, Bergisch Gladbach, Germany). Monocytes were plated at a 2×106 cell/mL concentration in RPMI (Hyclone, South Logan, UT) supplemented by 10% fetal calf serum (FCS; Gibco, Paisley, Scotland) and 1% antibiotic/antimycotic solution (Hyclone) in the presence of 100 ng/mL IL-4 and 75 ng/mL GM-CSF (Peprotech EC, London, United Kingdom) added on days 0 and 2.
Generation of mesenchymal stromal cell-like cells
Mesenchymal stromal cell-like cells derived from the human embryonic stem cell lines, HUES9 and HUES1, were kindly provided by Douglas Melton, HHMI. MSCl cells were used according to the ethical permission, 6681/2012/EHR. The cells were cultured on the mitotically inactivated mouse embryonic fibroblast (MEF) to form embryonic bodies, followed by trypsinization to obtain single cell cultures, which were further cultured on gelatin-covered 10-cm plates in Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal bovine serum [18]. Cells of the confluent cultures exhibited fibroblast-like morphology and were further characterized in a collaborative study with the laboratory of Balázs Sarkadi, Membrane Research Group of the Hungarian Academy of Sciences, Semmelweis University, and National Blood Service, Budapest, Hungary [17]. MSCl cell passages in optomechanical-treated polystyrene flasks (TPP, Trasadingen, Switzerland) provided coherent cell layers in the presence of L-glutamine, 10% FCS, and 1% antimycoticum/antibioticum solution (Hyclone) in low-glucose DMEM (Hyclone). The cultured MSCl cells were used after 10 passages.
Cocultures of DCs and MSCl cells
MSCl cells were collected by using 0.05%–0.02% Trypsin/EDTA solution in Dulbecco's PBS, washed, and cultured at a cell density of 4×105 cells in optomechanical-treated six-well plates for optimal cell growth on an 8.9 cm2/well area (TPP). To achieve confluent cell layers, the MSCl cells were cultured in 2.5 mL RPMI supplemented with 10% FCS containing 1% antimycotic/antibiotic solution (Hyclone) for a minimum of 6 h; 2×106 freshly isolated monocytes were placed directly on the top of the adherent MSCl cells, and the cocultured DCs were differentiated in the presence of GM-CSF and IL-4 at the same concentration used for DC differentiation.
Activation of DCs by inflammatory stimuli
On day 5 of in vitro DC differentiation, the cells were activated by different inflammatory stimuli in the presence or absence of MSCl cells. These involved an inflammatory cocktail containing 10 ng/mL TNF-α, 5 ng/mL IL-1β, 75 ng/mL GM-CSF, 20 ng/mL IL-6, and 1 μg/mL PGE2 (Peprotech EC), the RIG-I ligand poly(I:C) used at 25 μg/mL, or with 1 μg/mL 5′ppp-dsRNA introduced by the LyoVec transfection reagent (Invivogen, San Diego, CA). Twenty-four hours after activation, DCs expressing CD209/DC-SIGN on the cell surface were separated by magnetic beads (Miltenyi Biotec).
Phenotypic characterization of DCs differentiated in the presence or absence of MSCl cells
The phenotypic characterization of resting and activated DCs was performed by flow cytometry on day 6 of in vitro cell differentiation and compared with cells cocultured with MSCl cells (BD Biosciences, Franklin Lakes, NJ), and the data were analyzed by the FlowJo software (Tree Star, Ashland, OR). MSCl cells were identified by the expression of CD105, whereas DCs were captured by CD209/DC-SIGN (BioLegend, San Diego, CA). To measure changes in the expression levels of the CD80, CD83, CD86, and CD1a cell surface markers, fluorochrome-labeled antibodies were used along with the respective isotype-matched control antibodies (BD Pharmingen, San Diego, CA).
RNA isolation, cDNA synthesis, and quantitative RT-PCR
Total RNA was isolated by using TRIzol reagent (MRC, Cincinnati, OH). Two micrograms of RNA was reverse transcribed at 37°C for 120 min using the High-Capacity cDNA Archive Kit (Applied Biosystems, Foster City, CA) and oligo-(dT) primers (Promega, Madison, WI). Quantitative real-time PCR was performed by using gene-specific TaqMan assays (Applied Biosystems), DreamTaq DNA polymerase (Fermentas St. Leon-Rot, Germany) in a final volume of 12.5 μL, and in the ABI StepOnePlus real-time PCR instrument (Applied Biosystems). The housekeeping gene, h36B4, was used for data normalization. Cycle thresholds were determined by using the StepOne software v2.1 (Applied Biosystems).
Cytokine measurements
Concentration of the proinflammatory cytokines, IL-1β, TNF-α, and IL-6, the chemokines, CXCL8 and CXCL10, and that of the T-cell polarizing cytokines, IL-12, IL-10, and IFNγ, was measured in the supernatants of cell cultures by ELISA kits (BD Biosciences) following the manufacturer's instructions. Optical densities were determined by a microplate reader (BioTek, Winooski, VT).
ELISPOT assay
Activated DCs previously cocultured or not with MSCl cells were cultured with allogeneic T cells in serum-free RPMI medium for 3 days at 37°C. The number of IFNγ-secreting T cells was detected by the avidin-HRP system (NatuTec GmbH, Frankfurt, Germany) and the results were analyzed by the ImmunoScan plate reader (CTL, Shaker Heights, OH).
Migration assays
Migration of the differentiated DCs cocultured or not with MSCl cells was tested in a transwell system of 6.5 mm diameter and 5-μm pore size (Corning, Inc., Glendale, AZ). DCs were cocultured with MSCl cells for 6 days and were activated by PolyI:C or 5
Statistical analyses
Statistical analyses were performed by the one-way analysis of variance with Bonferroni's post hoc test using the GraphPad Prism v.6 software (GraphPad Software, Inc., La Jolla, CA). Differences were considered to be statistically significant at *P<0.05; **P<0.01; and ***P<0.005.
Results
MSCl cells inhibit the differentiation and activation of monocyte-derived DCs
Freshly isolated CD14+ monocytes were cultured in the presence of GM-CSF and IL-4 for 5 days to generate competent DCs used as control. Another fraction of DCs was differentiated at similar conditions together with MSCl cells added at a 5:1 monocyte-to-MSCl ratio present throughout the differentiation process. On day 5, both the control DCs and the DCs cocultured with MSCl cells were activated with poly(I:C), 5′pppRNA, or with an inflammatory cytokine cocktail containing GM-CSF, IL-1β, TNF-α, IL-6, and PGE2. On day 6, the activated DCs were separated from the MSCl cells by the positive selection of CD209/DC-SIGN-expressing cells and their proportion was measured in both the control and the MSCl-DC cocultures. This procedure resulted in cell populations with >90%±5% DCs in both the in vitro differentiated and the MSCl cocultured cells, indicating complete DC differentiation in both cases (data not shown). Cell surface expression of the CD1a (Fig. 1A) and CD83 (Fig. 1B) membrane proteins, used as indicators of DC activation, showed a significant decrease when the DCs were cocultured with MSCl cells irrespective of the mode of activation induced by poly(I:C), 5′pppRNA, or the inflammatory cocktail. Interestingly, stimulation of cells in the DC-MSCl cocultures by the specific RIG-I ligand, 5′pppRNA, resulted in significant upregulation of the costimulatory molecules, CD80 (Fig. 1C) and CD86 (Fig. 1D), on the DC surface compared with DCs cultured without MSCl cells, whereas the presence of MSCl cells did not affect the expression levels of these markers in DCs stimulated by the inflammatory cocktail or by poly(I:C) (Fig. 1C, D). These results indicated the potential of MSCl cells to modulate DC activation and consequently its other functional activities.
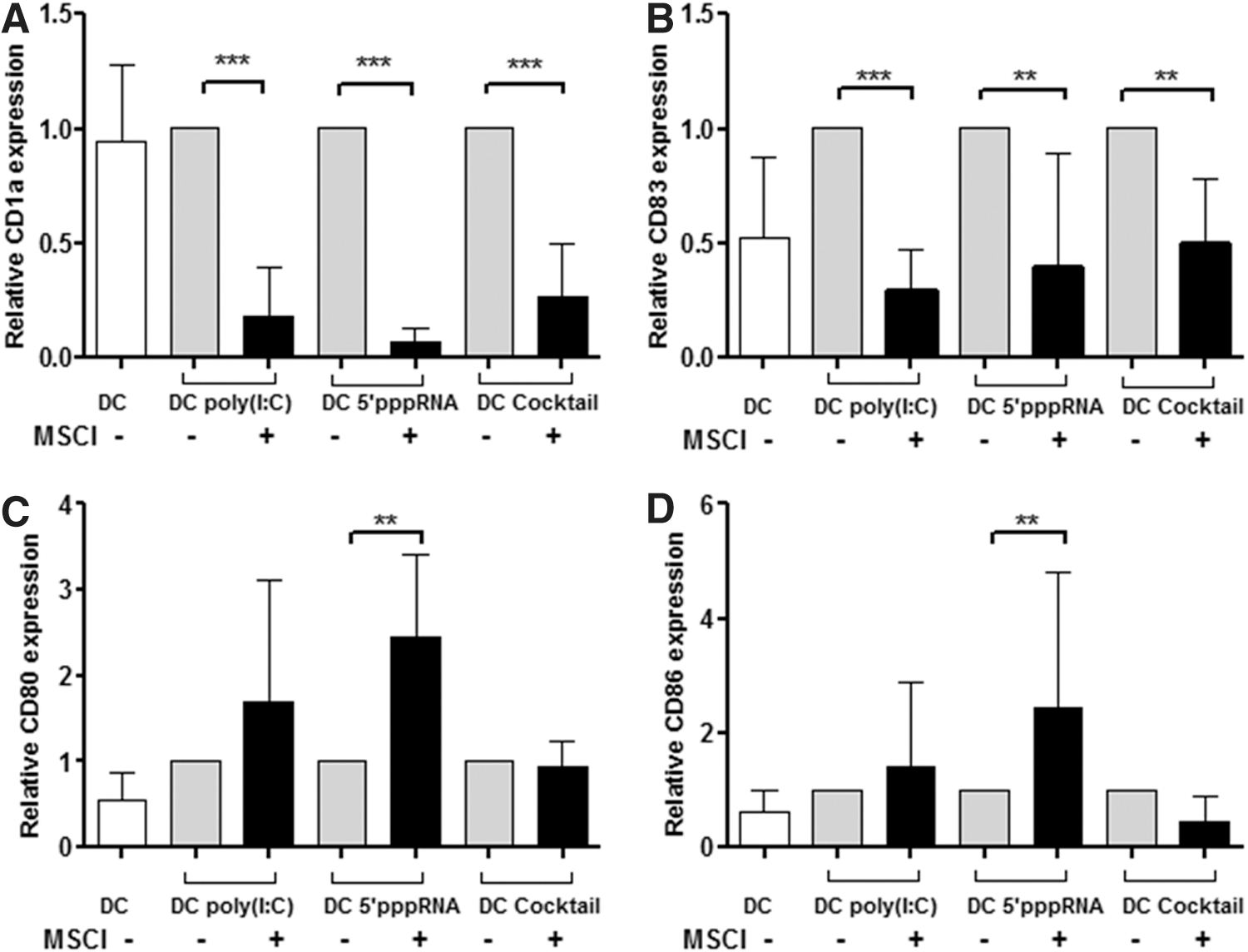
Relative expression of CD1a, CD83, CD80, and CD86 cell surface proteins by DCs activated in the absence or presence of MSCl cells. DCs were differentiated by GM-CSF+IL-4 in the absence or presence of MSCl cells at a DC:MSCl cell ratio of 5:1. On day 5 of DC differentiation, the cells were activated with poly(I:C), a synthetic ligand of RLRs, 5′pppRNA acting as a highly specific ligand of RIG-I, and by a proinflammatory cocktail containing GM-CSF, IL-6, TNF-α, IL-1β, and PGE2 for 24 h or were left untreated to serve as a negative control (resting DCs). On day 6, the activated DCs were separated by using CD209/DC-SIGN-specific antibody-coated magnetic beads. The relative expression levels of CD1a
MSCl cells modulate the cytokine secretion of activated DCs
To further analyze the functional cross talk of DCs and MSCl cells, the specific RIG-I ligands, poly(I:C) and 5′pppRNA, were added to the DC-MSCl cocultures and the secretion levels of the proinflammatory cytokines, IL-6 and TNF-α, the chemokines, CXCL8 and CXCL10, as well as the T-lymphocyte polarizing cytokines, IL-12 and IL-10, were measured in the supernatants of the activated DCs, MSCl cells, and DC-MSCl cocultures. We found that both RIG-I ligands could upregulate the secretion of IL-6 (Fig. 2A), CXCL8 (Fig. 2C), and IL-10 (Fig. 2E) in the DC-MSCl cocultures compared with activated DCs and MSCl cells cultured separately. Although IL-10 was present in the supernatants of both resting and activated DCs at low but measurable levels, we could not detect IL-10 in the supernatants of activated MSCl cells (Fig. 2E). However, the enhanced level of IL-10 in the supernatant of the cocultured and stimulated cells suggests that IL-10 could contribute to the anti-inflammatory effects of MSCl cells, presumably through inhibiting the secretion of proinflammatory cytokines.

Concentration of cytokines and chemokines produced by resting and activated DCs in the presence or absence of MSCl cells. DCs were differentiated in the presence or absence of MSCl cells as described in the Materials and Methods section. On day 5 of DC differentiation, the cells were activated by poly(I:C) or 5′pppRNA for 24 h, and the concentration of the secreted cytokines and chemokines was measured in the supernatants of resting DCs (empty bars) and DCs activated at different conditions (gray bars). MSCl cells treated with poly(I:C) and 5′pppRNA correspond to dashed bars, while the supernatants of the cocultured cells containing both DCs and MSCl cells are indicated by black bars. The concentration of the proinflammatory cytokines, IL-6
Our results also revealed that the concomitant secretion of TNF-α (Fig. 2B) and CXCL10 (Fig. 2D) did not induce a synergistic inflammatory effect, but could efficiently be inhibited by MSCl cells, indicating their potent anti-inflammatory activity. In this experimental system, the production of IL-12, a key Th1-polarizing cytokine, was also dramatically decreased in the presence of MSCl cells (Fig. 2F), likely owing to its potential to induce clonal expansion and differentiation of IFNγ-producing CD4+ T lymphocytes. Moreover, IL-12 was able to stimulate the production of other inflammatory cytokines, such as TNF-α, IFNγ, and the interferon-induced chemokine, CXCL10, to multiply the inflammatory response.
In line with these results, the production of IL-6 (Fig. 2A) and CXCL8 (Fig. 2C), both acting as important mediators of tissue repair and angiogenesis, was detected in the supernatants of DCs cocultured with MSCl cells at increased levels, suggesting that they may contribute to create a local anti-inflammatory milieu in response to inflammatory signals. These observations also imply that beside direct cell-to-cell contacts, cytokines and chemokines could also contribute to the regulatory functions of MSCl cells.
MSCl cells interfere with DC-mediated allogeneic T-lymphocyte activation and polarization
To further analyze the outcome of DC-mediated functional activities in the presence of MSCl cells, we sought to measure the secretion of IFNγ directly by ELISA and also as a result of DC-mediated allogeneic T-lymphocyte polarization by using ELISPOT assays. Our results demonstrated that IFNγ secretion of the DC could be induced by both poly(I:C) and 5′pppRNA; however, the presence of MSCl cells decreased its secretion significantly (Fig. 3A). The potential of poly(I:C) or 5′pppRNA-stimulated DCs to drive allogeneic T-lymphocyte polarization confirmed that MSCl cells are able to interfere with this translational event through exerting potent anti-inflammatory effects on DC activation (Fig. 3B). These results altogether demonstrate that DCs, upon interacting with MSCl cells, can create an anti-inflammatory local environment.
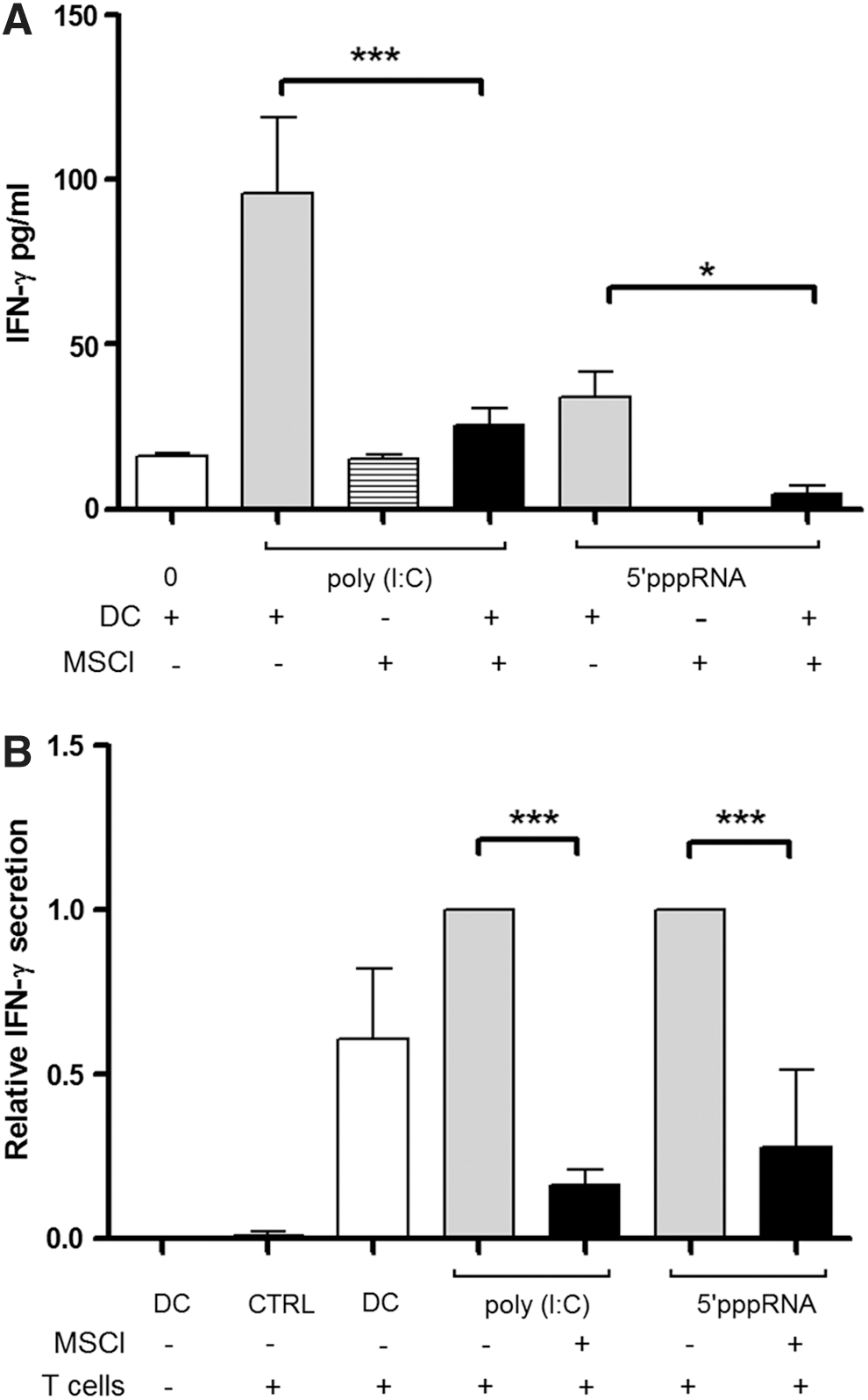
Production of IFNγ by activated DCs and by DC-induced allogeneic T lymphocytes.
The effect of MSCl cells on DC migration
Previous results demonstrated that efficient presentation of peptide antigens to naive T lymphocytes critically depends on the migration of tissue-resident DCs to the draining lymph nodes [33 –35]. As a novel finding, our previous studies demonstrated that the low expression level of the Nav1.7 ion channel, in combination with the high expression of CCR7, is the prerequisite of DC migration from the site of inflammation to the lymph nodes [32]. Based on this information, we used a transwell system to study the effects of MSCl cells on the migratory potential of DCs in the presence or absence of MSCl cells. In this experimental setting, the upper chamber contained resting DCs or DCs stimulated either with poly(I:C) or the inflammatory cytokine cocktail in the presence or absence of MSCl cells. The lower chamber was filled with fresh medium containing 0.5% bovine serum albumin and the chemokine, CCL19/MIP-3β, a strong inducer of activated DC migration guided by the membrane-bound chemokine receptor, CCR7. The upper and lower chambers were connected by a membrane of 5-μm pore size to allow chemokine-driven migration of DCs toward CCL19 gradients. The migrated cells were collected from the lower chamber, and after 24 h, the number of migrating cells was counted by flow cytometry. In correlation with the level of CCR7 chemokine expression in the presence of MSCl cells, a statistically significant decrease in the number of migratory cells was observed (Fig. 4A, B).
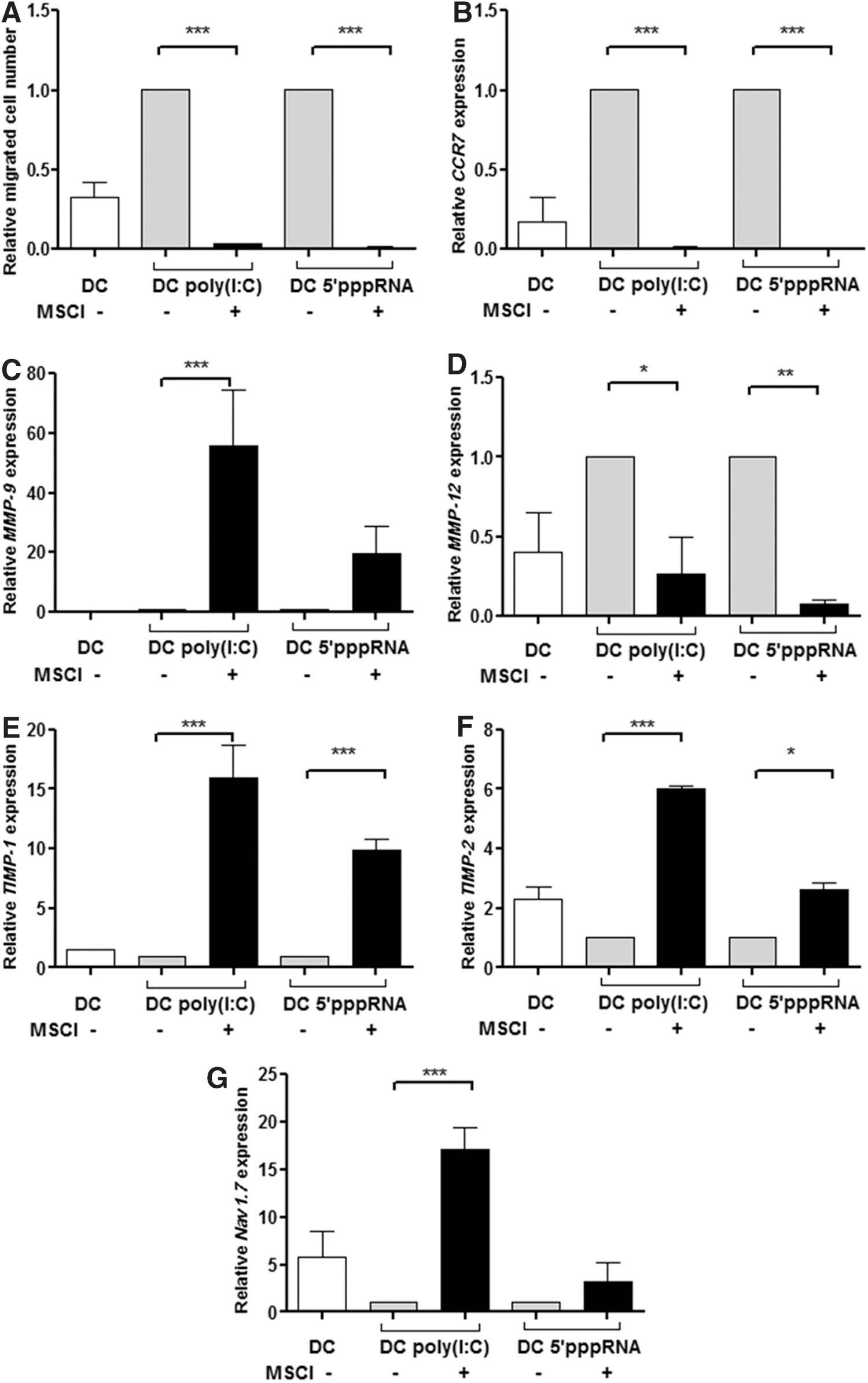
Migratory potential of DCs generated in the presence of MSCl cells.
Considering that the expression of MMP enzymes and their specific inhibitors (TIMP) plays important roles in regulating DC migration, we also assessed the contribution of these enzymes to the regulation of MSCl cell-mediated inhibitory functions. Expression of the MMP9, MMP12, TIMP1, and TIMP2 genes, which could be induced by the interaction of the CCR7 receptor with its specific ligand, CCL19, was monitored in both resting and activated DCs in the presence or absence of MSCl cells. We showed that the expression of MMP9 was upregulated (Fig. 4C), whereas that of MMP12 was downregulated (Fig. 4D). The relative gene expression levels of TIMP1 and TIMP2 were higher in CD1a- DC than in its inflammatory CD1a+ counterpart [32] and could be associated with a DC phenotype similar to cells cocultured with MSCl cells (Fig. 4E, F).
Another regulatory mechanism that might be involved in regulating MSCl cell-related functions could be associated with the activity of the Nav1.7 ion channel. We have previously shown that functionally active Nav1.7 is predominantly expressed in resting DCs, but its expression is decreased in parallel with DC activation, thus providing a sensitive tool for correlating the state of DC activation with Nav1.7 channel activity [32]. When the expression of Nav1.7 was measured in activated DCs without MSCl cells, the level of Nav1.7 expression decreased, even when the cells were stimulated by the inflammatory cocktail or by poly (I:C), but it was increased in DCs suppressed by MSCl cells (Fig. 4G). These changes are in good accordance with the low/undetectable numbers of inflammatory CD1a+ DCs detected in these cultures compared with the tolerogenic CD1a- DC subset [36,37].
MSCl cells inhibit the expression of RIG-like receptor family members
Our results summarized in Figs. 1 and 2 show that MSCl cells are potent inhibitors of DC functionality induced by the specific ligands, poly(I:C) and 5′pppRNA, of the cytosolic RIG-like receptors, RIG-I, MDA5, and LGP-2, recognizing dsRNA, ssRNA, or DNA [24]. Taking a step further, we hypothesized that coculturing DCs with MSCl cells might have a direct inhibitory effect on the expression of intracellular RIG-I receptors. Indeed, our Q-PCR results revealed that mRNA expression of the RIG-I, MDA5, and LGP-2 receptors belonging to the RLR family of cytosolic PRRs is upregulated upon stimulation by specific ligands, but in the presence of MSCl cells, this induction does not occur likely due to the inhibited expression of these receptors (Fig. 5A–C) accompanied by the decreased production of IFNβ (Fig. 5D). Based on these data, we conclude that in the presence of MSCl cells, the specific ligands of RIG-I could not be recognized by DCs due to the downmodulation of RLR receptor expression by MSCl cells.
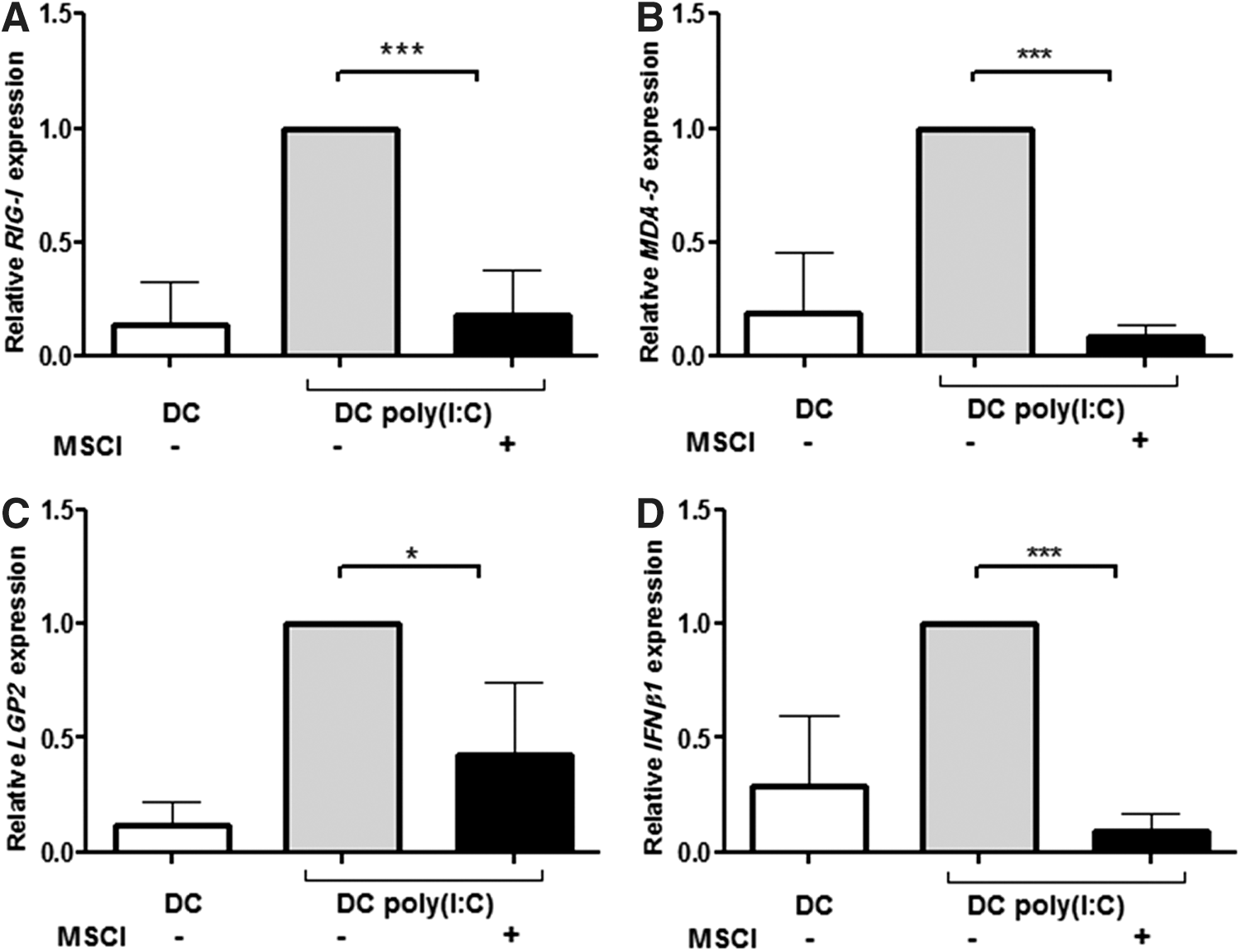
MSCl cells modulate gene expression levels of RLR and IFN-β1 in DCs. The mRNA expression levels of RIG-I
Discussion
The unique capability of MSCs of different origin is to modulate the outcome of allogeneic BM transplantation that has been pioneered by Le Blanc and her group in humans and confirmed in several mice models [9,38]. These results also revealed the potential of MSCs to exert multiple effects on other cell types through regulatory, anti-inflammatory, and bystander effects and through targeting injured tissues as described in several diseases, such as grade IV severe acute GVHD of the gut and liver [6] and steroid-resistant severe acute GVHD [39]. The utility of MSCs also involves the prevention of transplanted skin graft rejection [40], treatment of osteogenesis imperfecta by mismatched allogeneic liver-derived MSCs in an immunocompetent fetus [41], autoimmune encephalomyelitis [42], diabetes mellitus [43], and collagen-induced arthritis [44]. Further studies also demonstrated that the anti-inflammatory cytokine environment created by the transplanted MSCs at the site of inflammation was able to improve the outcome of acute renal, neural, and lung injuries [42,45,46]. Thus, in the past years, MSCs became clinically important cell types due to their regenerative potential that can be utilized in cell therapies aimed to treat inflammatory and autoimmune disorders or apply them for tissue engineering. This approach is also supported by the unique capability of MSCs to bypass MHC compatibility for inducing immunosuppression [47]. It is well established that MSCl cells can be stimulated by TLRs [26] and, upon stimulation, they migrate to damaged tissues driven by cytokines, chemokines, and secreted ECM proteins [48]. Beside the secretion of soluble factors, the direct contact of MSCs with immune cells seems to be essential for creating a supportive local milieu [49]. However, application of these cells for therapeutic interventions is limited due to the low number of MSCs available. To overcome this restriction, several groups explored new strategies for identifying cell lines with immunosuppressive properties [50]. In a previous collaborative study, we described the phenotypic, some functional, as well as the differentiating capability of a cell line of embryonic origin with the MSCl phenotype and compared its functional activities with BM-MSCs and a human foreskin fibroblast cell line [17]. This analysis demonstrated similar morphology, phenotype, and functional attributes resembling BM-derived MSCs and allowed us to exploit the further functional characterization of these cells in terms of their immunosuppressive potential. The present study provided evidence for the potent immunosuppressive nature of the MSCl cell line characterized by its profound effects on multiple functional properties of resting and activated monocyte-derived DCs. In a recent report, the immunosuppressive effects of human ESC-derived MA07 cells on CD83 expression and IL-12p70 secretion by IFNγ and LPS-activated DCs were observed to be associated with enhanced IL-2-induced expansion of regulatory T cells [16].
Our results performed with human primary monocyte-derived DCs also revealed that MSCl cells could affect the maturation, activation, and a wide array of functional activities in a similar manner as human BM-MSCs. As a consequence, the phenotype of DCs was modified in the presence of MSCl cells, resulting in decreased expression of the CD1a and CD83 activation markers on the surface of activated DCs induced by an inflammatory cytokine cocktail or by specific ligands of the RIG-I receptors. A similar effect was described in LPS-induced activation of mice in the presence of BM-MSCs [51] and TNF-α-induced stimulation of umbilical cord blood or BM-derived MSCs [52]. In contrast, the expression of the CD80 and CD86 costimulatory molecules was upregulated in the human DC-MSC cocultures, while in mice, the expression of these molecules was decreased in the presence of both murine and human BM-MSCs [53,54], indicating species-related differences in the inhibitory function of MSCs.
The secretion of CXCL8 and IL-6 by MSC1 and CXCL10 by MSC2 cells suggested the induction of different cytokine combinations induced by these MSC subsets [26] similar to our results showing increased secretion of IL-6, CXCL8, and CXC10 by MSCl cells activated by poly(I:C). However, no detectable changes could be observed when the RIG-I-specific 5′pppRNA ligand was used for DC activation. In contrast, the activation of cocultured DCs and MSCs resulted in dramatic changes in the secretion of cytokines and chemokines. Even though changes in cytokine secretion by DCs have been demonstrated in the presence of MSCs of various origins, the mechanistic background of this regulation has not been explored.
It was also shown that MSCs, isolated from MEFs, could induce the generation of IL-10-dependent regulatory DCs through SOCS3 activation, leading to increased secretion of anti-inflammatory IL-10 [55]. The cytokines, IL-6 and CXCL8, have been considered as important mediators of tissue repair and angiogenesis. In LPS-stimulated DCs, the presence of MSCs increased the secretion of IL-6 and CXCL8 [56,57], and in response to poly(I:C) or 5′pppRNA, the increased secretion of IL-6, IL-10, and CXCL8 was also detected, opposing the significantly decreased levels of IL-12 and TNF-α in the supernatant of the DC-MSCl cocultures. These results suggest that the contact and/or the communication of cells and soluble factors may trigger anti-inflammatory and/or healing mechanisms that shift the balance of the immune response to tissue regeneration and changes in cytokine and chemokine secretion.
The CCL19 chemokine is expressed in the thymus and lymph nodes and its secretion is essential for the migration of CCR7high DCs to the draining lymph nodes where they interact with naive T lymphocytes [33]. However, the expression of CCR7 and the migration of DCs derived from the DC-MSCl cocultures exhibited significantly decreased migration compared with DCs cultured in the absence of MSCl cells. One of the mechanisms involved in this complex regulation was discovered recently [32], showing the contribution of the voltage-gated membrane channel, Nav1.7, which regulates intracellular Ca2+ concentration in DCs and also acts as a master regulator of the cell cycle. High intracellular Ca2+ concentration was shown to be essential for DC activation and was associated with downregulated Nav1.7 expression. As anticipated, the expression of the voltage-gated sodium channel, Nav1.7, was also dramatically downmodulated and could be used as a sensitive indicator of the DC activation state [32]. On the contrary, Nav1.7 expression was maintained in the presence of MSCl cells, which may indicate either dysregulation of the Ca2+ signal accompanying DC activation or an alternative pathway for the regulation of Nav1.7 expression. Either way, MSCl cells interfere with DC activation, thereby identifying this sodium channel as a novel marker of suppressed DCs. Overall, these data indicate that in the presence of MSCl cells, the generation of DCs results in semiactivated Nav1.7high and CCR7low cells, which express high amounts of active MMP9 enzyme and also TIMP with inhibitory potential to induce dramatically reduced migratory potential of DCs.
Upregulation of CXCL8 chemokine was shown to support the expression of MMP-2 and MMP-9 in trophoblast cells [58]. The balance of MMP and TIMP is also known to have an impact on cell migration, homeostasis, survival, and other functional activities of immune cells through regulating the cleavage of ECM components to generate soluble cytokines, chemokines, and growth factors together with their matching receptors [59]. Considering that the regulation of the MMP-TIMP axis is also involved in the triggering of signal transduction pathways connected to DC functions, we provoked DC migration with an inflammatory cocktail or with poly(I:C) and could increase the expression levels of MMP-9 and MMP-12 in DCs accompanied by decreased TIMP-1 and TIMP-2 expression showing a tight control of the inflammatory response [31]. However, in the presence of MSCl cells, the expression level of TIMP was dramatically upregulated and resulted in inhibited DC migration through blocking ECM degradation. More importantly, these effects could also modify the regulatory capacity of MMPs by the secretion and proteolytic cleavage of cytokines and chemokines. Based on these results, we conclude that MSCl cells are able to modulate the expression of key molecules involved in DC migration. Moreover, the presence of MSCl cells may also have an impact on the level and activity of secreted cytokines and chemokines and can affect the expression of their receptors for fine-tuning DC activities in the actual inflammatory environment.
In vivo studies have revealed the inhibition of antigen-specific effector T-cell functions in the course of mouse allograft rejection [40], in GVHD [60], autoimmune encephalomyelitis [42], and collagen-induced arthritis [44]. In a murine model, DCs conditioned by allogeneic MSCs could inhibit MHC class I and II expression and antigen presentation by DCs, while decreasing CD69 expression on CD8+ T cells. In a human system, the presence of activated DCs cocultured with BM-MSCs was shown to decrease the secretion of IFNγ by T cells [61]. In the present study, the inhibited secretion of IFNγ could be detected by a sensitive ELISPOT assay. In this experimental setting, human allogeneic T lymphocytes, primed by activated DCs and cocultured with MSCl cells, were able to downregulate T-cell polarization to the Th1 direction to a similar extent as DCs cocultured with BM-MSCs. In our human in vitro studies, MSCs also could alter DC functions significantly and together with our previous studies confirmed the potential of MSCl cells to exert inhibitory signals on antigen-specific T-cell responses.
In this context, however, the expression of RIG-like receptors has not been investigated. We hypothesized that the functional changes induced by the RLR ligand, poly(I:C), are the consequence of decreased RLR expression. Our results indeed demonstrated that in the presence of MSCl cells, the expression of RIG-I, MDA5, and LGP-2 in DCs was dramatically impaired. Referring to the previously described partially activated DCs with tolerogenic and immunosuppressive properties [62], MSCl cells may assist in the maintenance of DCs in a semimature suppressive state [63]. Our results obtained in a human system suggest that MSCl cells with unlimited proliferating capacity could be harnessed as a reliable model for inhibiting DC activation by MSC-mediated immunosuppression.
Footnotes
Acknowledgments
This research work was supported by the Hungarian Scientific Research Grant, OTKA NK 101538, the European Union, and the State of Hungary, cofinanced by the European Social Fund in the framework of the TÁMOP 4.2.4.A/2-11-1-2012-0001 National Excellence Program and the TÁMOP 4.2.2.A-11/1/KONV-2012-0023 DEFENSE-NET project.
Author Disclosure Statement
No competing financial interests exist.