
Abstract
Small-sized adult bone marrow cells isolated by counterflow centrifugal elutriation and depleted of lineage markers (Fr25lin−) have the capacity to differentiate into insulin-producing cells and stabilize glycemic control. This study assessed competitive migration of syngeneic stem cells to the bone marrow and islets in a murine model of chemical diabetes. VLA-4 is expressed in ∼25% of these cells, whereas CXCR4 is not detected, however, it is transcriptionally upregulated (6-fold). The possibility to enrich stem cells by a bone marrow homing (BM-H) functional assay was assessed in sequential transplants. Fr25lin− cells labeled with PKH26 were grafted into primary myeloablated recipients, and mitotically quiescent Fr25lin−PKHbright cells were sorted from the bone marrow after 2 days. The contribution of bone marrow-homed stem cells was remarkably higher in secondary recipients compared to freshly elutriated cells. The therapeutic efficacy was further increased by omission of irradiation in the secondary recipients, showing a 25-fold enrichment of islet-reconstituting cells by the bone marrow homing assay. Donor cells identified by the green fluorescent protein (GFP) and a genomic marker in sex-mismatched transplants upregulated PDX-1 and produced proinsulin, affirming the capacity of BM-H cells to convert in the injured islets. There was no evidence of transcriptional priming of freshly elutriated subsets to express PDX-1, insulin, and other markers of endocrine progenitors, indicating that the bone marrow harbors stem cells with versatile differentiation capacity. Affinity to the bone marrow can be used to enrich stem cells for pancreatic regeneration, and reciprocally, conditioning reduces the competitive incorporation in the injured islets.
Introduction
C
Various subsets of the adult BMC participate in islet recovery through distinct mechanisms [16]. The small-sized stem cells convert to express specific pancreatic endocrine markers and stabilize blood glucose levels through neogenesis of insulin-producing cells [7,10]. In variance, large-sized hematopoietic progenitors support islet remodeling through neovascularization, immunomodulation, stabilization of the stroma, and provision of cytokines [10]. In this study, we assessed competitive distribution of Fr25lin− cells to the irradiated bone marrow and injured islets, following the observation that systemic conditioning reduces cell incorporation in the injured retina [8]. Therefore, we assessed whether irradiation impairs regeneration of β-cells by freeing space and attracting the cells to the injured marrow space. We found that preconditioning for stem cell transplantation compromises the maximal regenerative capacity of these cells in the islets due to competitive homing to the bone marrow. Nevertheless, initial homing to the bone marrow substantially increases the frequency of stem cells within the Fr25lin− subset endowed with the capacity to convert and produce insulin.
Materials and Methods
Animal preparation, diabetes, and transplantation
Mice used in this study were C57Bl/6J (B6, H2Kb, CD45.2), B6.SJL-Ptprca Pepcb/BoyJ (H2Kb, CD45.1), and C57BL/6-TgN(ACTbEGFP)1Osb (GFP, H2kb, CD45.2) purchased from Jackson Laboratories (Bar Harbor, ME) and housed in a barrier facility. The Institutional Animal Care Committee of the Schneider Medical Center has approved all procedures. Diabetes was induced in female mice (aged 6–8 weeks) by five daily consecutive intraperitoneal injections of 60 μg/g streptozotocin (STZ; Calbiochem, Darmstadt, Germany) [7]. Blood glucose levels were monitored with a standard glucometer (Accu-Chek Sensor; Roche Diagnostics, Indianapolis, IN) in mice fed ad libitum at constant daytime hours (9–11 AM), considering hyperglycemia in two consecutive measurements exceeding 250 mg/dL. The glucose tolerance test was performed by intraperitoneal injection of 2 g glucose and blood level measurements after 60 and 120 min. In some experiments, the recipients were sublethally irradiated at 675 rad (total body irradiation) using an X-ray irradiator (RadSource 2000, Suwanee, GA). Cells were grafted intravenously (lateral tail vein) and suspended in 0.2 mL of phosphate-buffered saline (PBS).
Cell isolation
Whole bone marrow cells from male wild-type and green fluorescent protein (GFP) donors were harvested by flushing of medullar cavities of femur, tibia, and iliac bones with PBS. Cell suspensions were loaded into the chamber of a counterflow centrifuge rotor J6 (Beckman Instruments, Palo Alto, CA) operating at a constant speed of 3,000 rpm (867 g) for collection of the smallest subset of nucleated cells (Fr25) at an elutriation flow rate of 25 mL/min and large-sized progenitors (R/O) in the rotor off position [17]. Fr25 cells were lineage depleted by incubation with saturating amounts of rat anti-mouse monoclonal antibodies (mAb) against CD5 (clone 53-7.3), B220 (clone RA3-3A1/6.1), GR-1 (clone RB6-8C5), and Mac-1 (clone M1/70), extracted from hybridoma cell lines (ATCC, Manassas, VA) and purified TER119 (eBioscience, San Diego, CA). Antibody-bound cells were conjugated to magnetic beads using goat anti-rat antibodies (Dynal Biotech, Oslo, Norway) and were collected by exposure to a magnetic field. The pancreas was dissected into small pieces, incubated with 20 μg/mL of Collagenase P (Roche Diagnostics) for 30 min at 37°C, and passed through a 40-μM nylon mesh (SPL Life Science, Gyeongg, Korea) to prepare single-cell suspensions.
The elutriated subsets were evaluated for size (side scatter), expression of CD45, and lineage markers using a cocktail of labeled mAb corresponding to those used for lineage depletion (Supplementary Fig. S1A; Supplementary Data are available online at
Flow cytometry
Measurements and sorting were performed with a Vantage SE flow cytometer (Becton Dickinson, Franklin Lakes, NJ) on cells that underwent red cell lysis (Sigma-Aldrich, St. Louis, MO), as previously detailed [17]. Adhesion molecules were identified with anti-CD49d (integrin alpha 4, clone R2-1) for VLA-4, anti-CD184 (clone 2B11) for CXCR4 (eBioscience), and anti-CD62E (clone UZ4) for E-selectin (Millipore, Billerica, MA). Labeling was performed by incubation of 107 cells/mL of 2 μM PKH26 in diluent-C for 10 min at room temperature (Sigma-Aldrich). Cells were collected by centrifugation, resuspended in PBS supplemented with 10% fetal calf serum, and washed twice with PBS.
Determination of blood insulin
Serum from mice was collected by centrifugation and assessed in 96-well Microtiter Assay Plates (Millipore) using the Rat/Mouse Insulin ELISA Kit (R&D Systems, Abingdon, United Kingdom). Absorbance was determined using an ELISA PowerWave-10 in a plate reader (BioTeK, Winooski, VT), using insulin standards for calibration [7,10].
Tissue preparation
The procedures of tissue preparation, immunohistochemistry, and fluorescence in situ hybridization have been described in detail [18]. Mice sacrificed by CO2 asphyxiation were perfused with an ice-cold fixative containing 1.5% fresh paraformaldehyde (Bio-Lab, Jerusalem, Israel) and 0.1% glutaraldehyde (Sigma-Aldrich). Excised pancreata were placed in this medium for 2 h at 4°C for additional fixation, then were immersed in 30% sucrose overnight (Sigma-Aldrich). Samples were embedded in OCT (Sakura Finetek, Torrance, CA), frozen in isopentane (Sigma-Aldrich), suspended in liquid nitrogen, and sectioned (3–6 μm) with a Cryotome (Thermo Shandon, Cheshire, United Kingdom).
Immunofluorescence and FISH
Sections fixed in acetone were permeabilized with 0.2% saponin (Merck, Darmstadt, Germany), 1% bovine serum albumin (Sigma-Aldrich), and 0.1% triton-100 (Sigma-Aldrich), stained with primary antibodies for 1 h, washed and counterstained with respective secondary antibodies [18]. Nuclei were labeled with Hoechst-33342 (1:1,000; Molecular Probes, Eugene, OR) in air-dried sections mounted in the antifade medium (Dako, Glostrup, Denmark). Serial cryosections were immunostained with primary antibodies: polyclonal mouse anti-proinsulin (1:20; R&D Systems), polyclonal goat anti-PDX-1 mAb (1:5,000; Abcam, Cambridge, MA), and biotinylated rabbit anti-GFP mAb (1:100; Santa Cruz Biotechnology, Santa Cruz, CA). These primary antibodies were counterstained with labeled donkey anti-mouse (1:1,000; Jackson ImmunoResearch, West Grove, PA), rat anti-goat (1:200; Molecular Probes), and Cy3-conjugated streptavidin (1:400; Jackson ImmunoResearch).
Y chromosomes in male donor cells were visualized in immunostained slides after treatment with 0.025% pepsin (Sigma-Aldrich) as previously detailed [18]. Slides were rinsed in 2× saline sodium citrate (SSC) (Bio-Lab), dehydrated in ethanol, air-dried, then were fixed in 70% formamide (Sigma-Aldrich) and cold 70% ethanol, and were dehydrated again in graded ethanol solutions. Nuclear probes (Applied Spectral Imaging; Migdal Haemek, Israel) were denatured and hybridized overnight, then sequentially washed in SSC stringency solution with 0.1% NP4O detergent (Sigma-Aldrich). Washed slides were mounted with antifade containing DAPI and imaged with an Axioplan 2 (C. Zeiss, Göttingen, Germany) fluorescence microscope equipped with an Apotome. Images were pseudocolored and RGB reconstructed using Adobe Photoshop software.
Western blots and real-time polymerase chain reaction
Western blots used rabbit anti-PDX-1 polyclonal antibodies (1:500; Chemicon, Billerica, MA) and peroxidase-conjugated goat anti-rabbit IgG (1:10,000; Jackson ImmunoResearch) [10]. RT-PCR was used to study the expressions of insulin, PDX-1, VLA-4, and CXCR4 in nucleated whole BMC and the elutriated R/O and Fr25lin− subsets. Fractions were frozen in liquid nitrogen for total RNA extraction with TRIzol (Invitrogen, Grand Island, NY), followed by reverse transcription into cDNA using random hexamers (Amersham Biosciences, Cardiff, United Kingdom) and Moloney Murine Leukemia Virus Reverse Transcriptase (Promega, Madison, WI). cDNA was analyzed with a Sequence Detection System (ABI Prism 7900; Applied Biosystems, Foster City, CA) [19]. Gene expression was measured by normalization of cDNA input levels against mouse β-actin. Reactions were performed for forward and reverse primers in the Master Mix buffer (SYBR Green I; Applied Biosystems) according to the real time-polymerase chain reaction (RT-PCR) cycling conditions and quantified as previously described [10,20]. Primers are listed in Table 1.
Statistical analysis
Data are presented as means±standard deviations for each experimental protocol. Results in each experimental group were evaluated for reproducibility by linear regression of duplicate measurements. Differences between the experimental protocols were estimated with a post hoc Scheffe t-test, and significance was considered at P<0.05.
Results
Efficiency and mediators of migration to the injured islets
The efficiency of Fr25lin− cell homing to the bone marrow has been characterized in detail in myeloablated recipients, showing a superior affinity of Fr25lin− cells compared to R/O progenitors [11,14]. We have used models of sublethal irradiation in conjunction with STZ to induce chemical diabetes [7,10,18] and models of ischemic optic neuropathy [8]. In view of the superior incorporation of cells in the injured retina in the absence of irradiation, we were interested to quantify the efficiency of migration of the elutriated stem cell subsets to the bone marrow and pancreas under our experimental conditions. Irradiated diabetic recipients were grafted with 106 freshly elutriated Fr25lin− cells from donors that constitutively express GFP, and after 2 days, the bone marrow and pancreata were harvested and assessed for the presence of Fr25lin−GFP+ cells (Fig. 1A). Fractional incorporation of grafted cells in the pancreas was higher than in the bone marrow (Fig. 1B), demonstrating competitive distribution of Fr25lin− cells between the two sites of injury. In variance, the homing efficiency of R/O hematopoietic progenitors was substantially lower, with inverse fractional incorporation: 4.5-fold reduced homing to the bone marrow and 8-fold reduced migration to the pancreas compared to Fr25lin− cells.
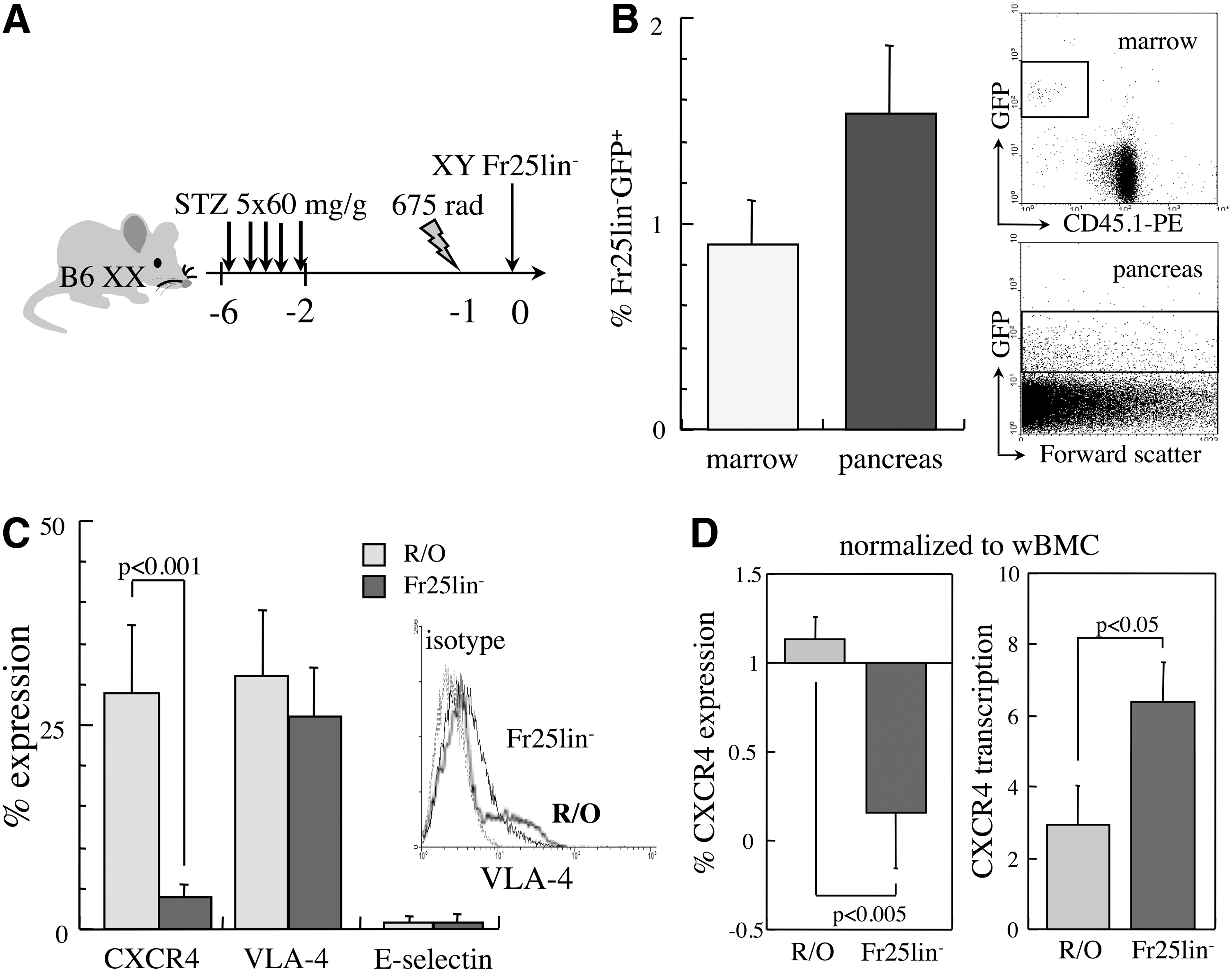
Molecular affinities of elutriated subsets to the bone marrow and injured islets.
To quantify the efficiency of homing to the bone marrow and migration to the pancreas, fractions were normalized against the total number of cells obtained from the two tissues at 2 days after grafting into irradiated recipients. The number of Fr25lin−GFP+ harvested from the hind limbs (femurs and tibia) was 44±0.5×103 Fr25lin−GFP+ donor cells within 4.6±0.5×106 total BMC. Considering that the marrow space of the hind limbs represents 15% of the total marrow space [21], 2.9±0.3×105 donor cells homed to the entire murine marrow correspond to a homing efficiency of 14% of the Fr25lin− graft. In parallel, 1.3±0.3×105 Fr25lin−GFP+ donor cells were found within 8.4±1.8×106 pancreatic cell suspensions, corresponding to a migration efficiency of 13%. Therefore, quantitative incorporation of Fr25lin− cells is evenly distributed between the bone marrow and pancreas in this model of sublethally irradiated diabetic mice.
Mediators of cell migration
CXCR4, VLA-4, and E-selectin were evaluated as major molecular mechanisms involved in hematopoietic cell homing, because their cognate ligands are abundant in the injured islets [22 –24] as well as the irradiated bone marrow [21,25]. Approximately 25%–30% of R/O progenitors expressed both VLA-4 and CXCR4, and VLA-4 was detected in a similar fraction of Fr25lin− cells along very low levels of CXCR4, whereas E-selectin was virtually absent in both subsets (Fig. 1C). To determine the impact of separation by elutriation on the prevalence of these adhesion molecules, the isolated fractions were normalized against whole nucleated BMC. R/O progenitors displayed increased transcription and translation of CXCR4, whereas the Fr25lin− subset showed markedly increased (6-fold) transcription in the presence of very low levels of the protein product (P<0.005, Fig. 1D). These data revealed that unlike hematopoietic progenitors, CXCR4 is transcriptionally upregulated but not translated in Fr25lin− stem cells.
Bone marrow homing enriches stem cells
Efficient homing to the bone marrow represents a significant characteristic of Fr25lin− cells endowed with versatile differentiation capacity [3,11 –14]. To determine whether homing to the bone marrow enriches stem cells within the Fr25lin− subset, this functional assay was applied to our model of chemical diabetes. Fr25lin− cells labeled with PKH26 membrane linkers were grafted intravenously (male→female) into lethally irradiated primary syngeneic recipients (Fig. 2A). BMC from the hind limbs were harvested after 2 days, and mitotically quiescent donor cells were sorted according to retention of bright PKH26 fluorescence (Fig. 2B). These BM-H FR25lin−PKHbright cells were subsequently grafted into secondary recipients following induction of diabetes with STZ and sublethal irradiation (Fig. 1A). A series of calibration experiments determined a threshold of 2.5×105 R/O progenitors to support short-term hematopoiesis and survival of the sublethally irradiated diabetic recipients. Grafting of female R/O progenitors and 4×104 male Fr25lin−PKHbright cells significantly reduced the fasting blood glucose levels (296±90 mg/dL, P<0.05 vs. untreated diabetic mice, Fig. 2C) and improved the responses to the glucose tolerance test (Fig. 2D). Superior glycemic control was accompanied by a consistent rise in insulin levels at 16 weeks post-transplantation of BM-H Fr25lin−PKHbright cells (P<0.01, Fig. 2E). Considering that R/O cells do not have a significant impact on insulin levels, glucose levels in this experiment were higher than the joint therapeutic efficacy of 106 freshly elutriated R/O and Fr25lin− cells in this model (190±35 mg/dL) [10].
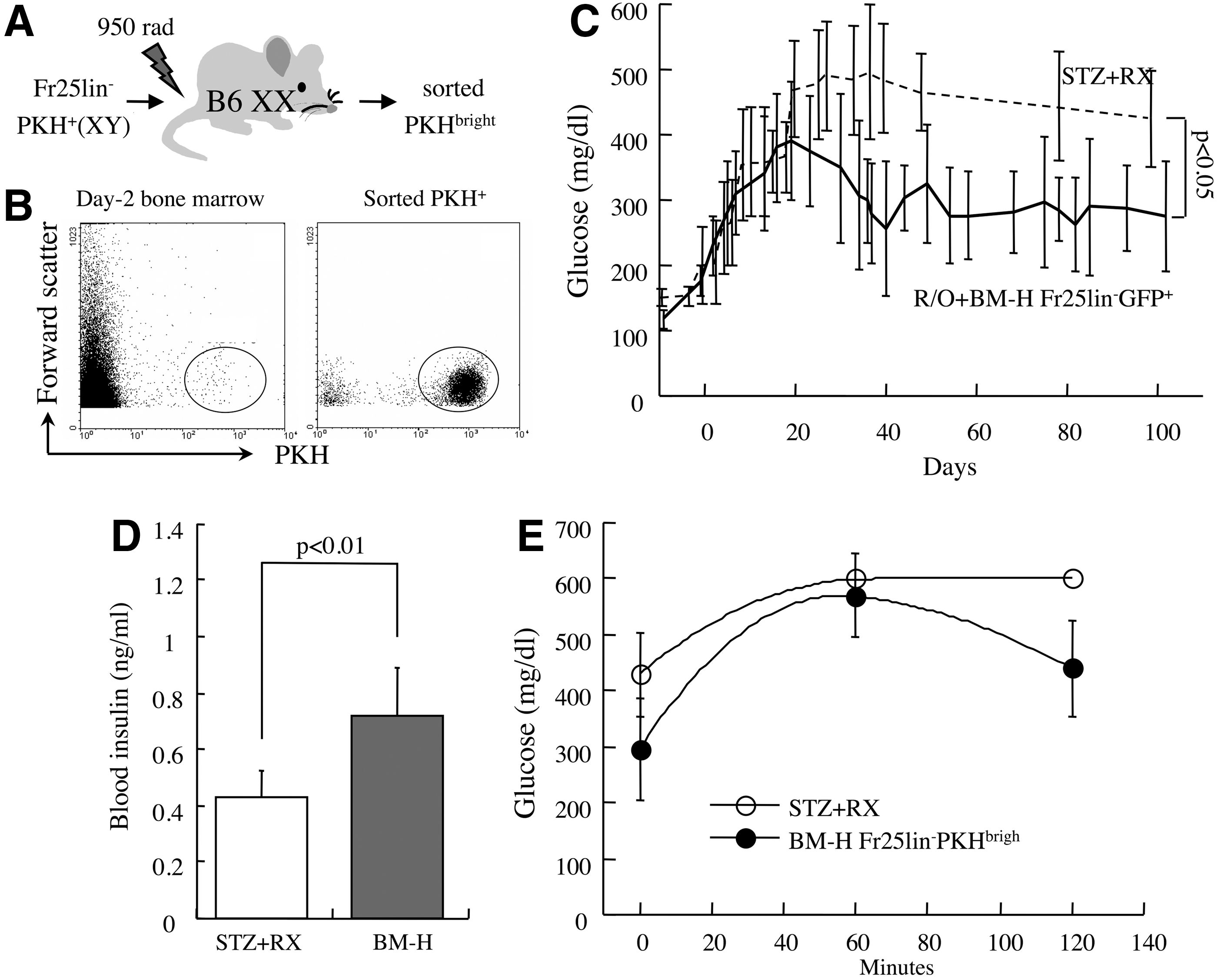
Bone marrow-homed Fr25lin− cells ameliorate chemical diabetes.
The most significant contribution of Fr25lin− cells to islet recovery is mediated by neogenesis of insulin-producing cells that upregulate PDX-1 and downregulate GFP [7,10], whereas R/O progenitors do not display such conversion [10]. To validate that day-2 bone BM-H Fr25lin−GFP+PKH26bright cells convert to produce insulin, recipient pancreata were assessed at 6 weeks after transplantation in a sex-mismatched (male→female) setting. GFPdim cells incorporated in the injured islets stained positive for proinsulin, whereas cells located at the islet perimeter retained bright GFP fluorescence (Fig. 3A). Donor origin of the differentiating cells was validated by detection of a genomic marker (Y chromosome) in cells positively identified to convert by nuclear PDX-1 expression in these sex-mismatched transplants (Fig. 3B). Consequently, proinsulin was coexpressed with dim GFP in the cytosol of donor cells displaying nuclear PDX-1 (Fig. 3C). Upregulation of PDX-1 and production of proinsulin in diabetic mice displaying improved glycemic control and elevated insulin levels affirmed that day-2 bone marrow-homed Fr25lin− stem cells retained endocrine pancreatic differentiation capacity. Notably, the vast majority of donor cells incorporated in the pancreas retained bright GFP fluorescence at extra-islet locations (surrounding islets, ducts, vasculature).

Bone marrow-homed Fr25lin− cells convert to upregulate PDX-1 and produce proinsulin. Day-2 bone marrow-homed Fr25lin−GFP+PKHbright cells from male green fluorescent protein (GFP) donors were grafted into diabetic female recipients.
Conditioning decreases stem cell migration to the injured islets
Competitive affinity of Fr25lin− cells to the bone marrow and pancreas (Fig. 1B) and superior outcome of recipients of bone marrow-homed cells (Fig. 2C–E) suggest that irradiation of the diabetic mice reduces the therapeutic efficacy. The same experimental model (Fig. 1A) was used to evaluate the efficiency incorporation of bone marrow-homed stem cells in nonirradiated secondary diabetic recipients, which evidently do not require transplantation of supporting R/O progenitors. Glycemic control was markedly improved by transplantation of 4×104 day-2 BM-H Fr25lin−PKHbright cells (210±48 mg/dL at 16 weeks) compared to untreated mice (P<0.005, Fig. 4A). Before sacrifice for histological analysis, blood insulin levels (Fig. 4B) and the glucose tolerance test (Fig. 4C) showed marked improvement following stem cell grafting compared to untreated diabetic mice. Thus, omission of irradiation resulted in a superior outcome compared to grafting the same number of BM-H Fr25lin−PKHbright stem cells in conjunction with supporting R/O progenitors into sublethally irradiated recipients (P<0.05, Fig. 2C) and was comparable to transplantation of 106 freshly elutriated Fr25lin− cells in sublethally irradiated diabetic recipients (205±48 mg/dL) [10]. The longest follow-up period showed sustained glycemic control at 26 weeks in two mice. These data correspond to an approximate 25-fold enrichment of stem cells within the Fr25lin− subset attained by the following: (a) a priori bone marrow homing assay and (b) grafting into nonconditioned diabetic mice.
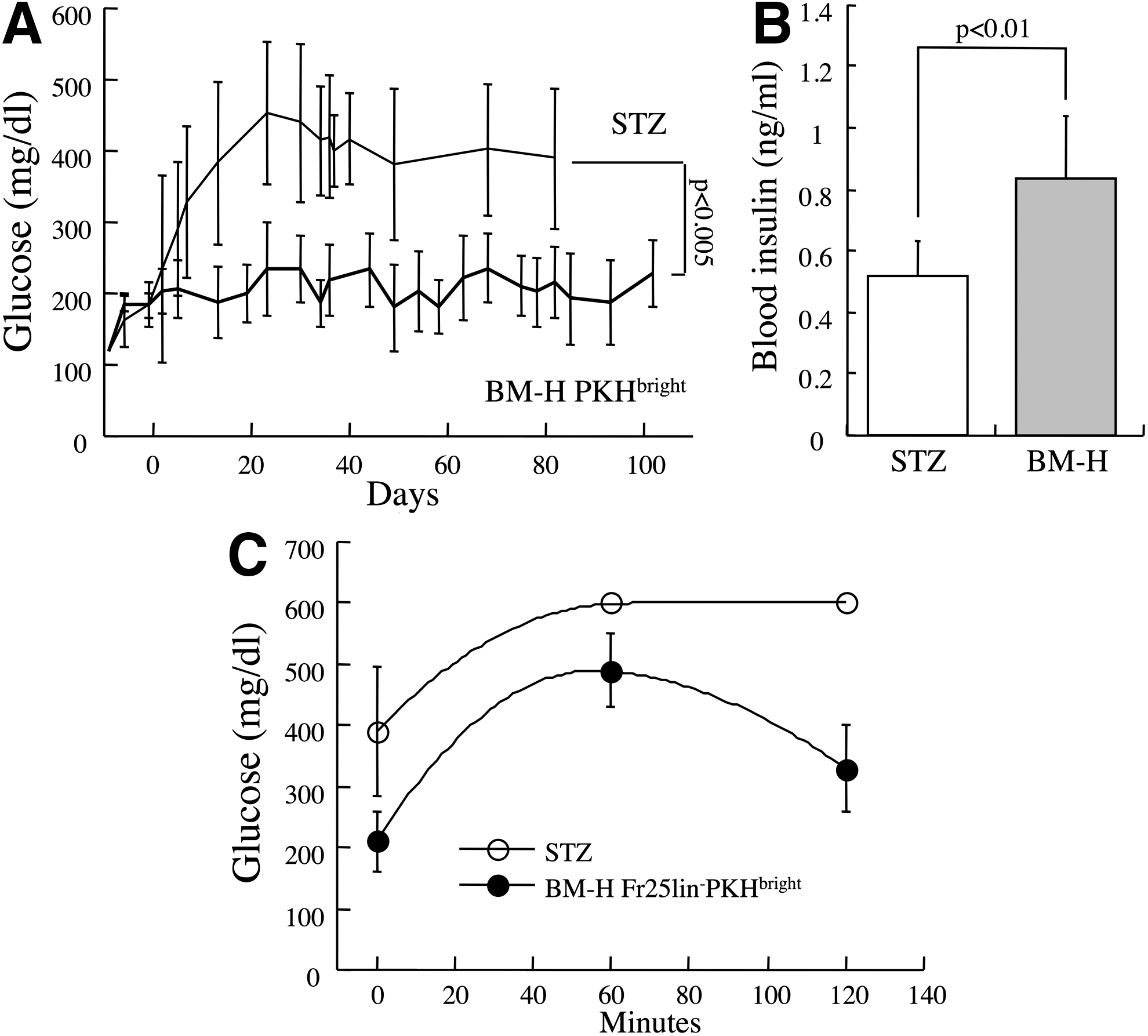
Superior therapeutic efficacy of bone marrow-homed Fr25lin− cells in the absence of radiation. Primary myeloablated (950 rad) primary female recipients were injected with male Fr25lin−PKH26+ cells, after 2 days the grafted PKHbright cells were sorted from the bone marrow and grafted into secondary diabetic recipients without radiation.
The bone marrow does not contain progenitors of the endocrine pancreas
An ongoing debate relates to the possible sequestration of progenitors for various tissues within the bone marrow compartment [26,27]. To determine whether pancreatic endocrine progenitors in the bone marrow are extracted by the elutriation procedure, we assessed transcription of two hallmarks of the endocrine pancreas. Whole BMC and Fr25lin− cells display transcription of 2%–3% PDX-1 (Fig. 5A) and insulin (Fig. 5B) compared to neonate pancreas and, consistently, the corresponding protein products were not detected. These findings are inconsistent with the possibility that the bone marrow harbors progenitors of the endocrine pancreas.
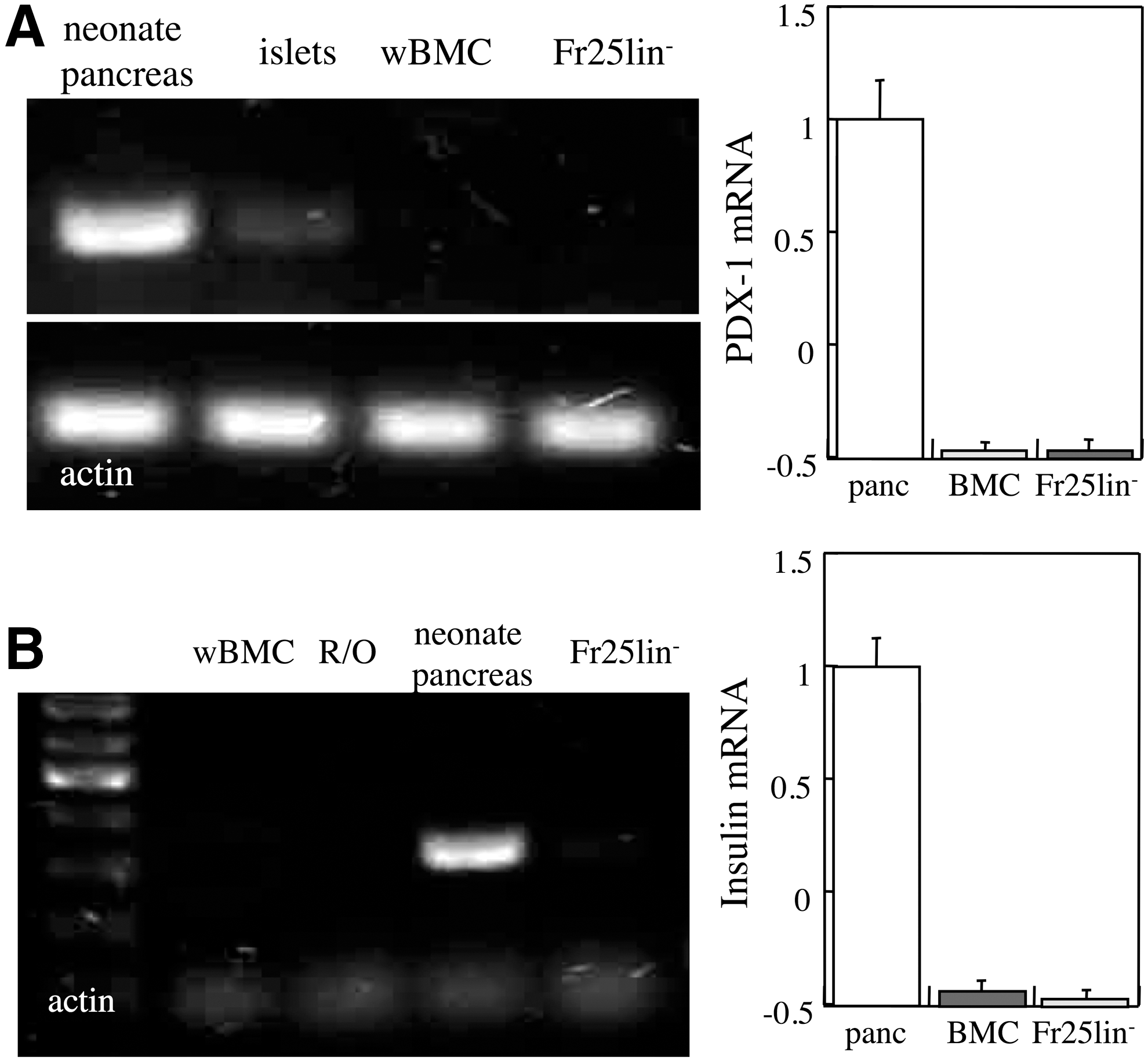
Absence of transcription of β-cell markers in the elutriated bone marrow subsets. PDX-1
Discussion
The adult bone marrow contains a subset of small-sized stem cells endowed with the capacity to adopt versatile differentiation traits [5 –8], including neogenesis of insulin-producing cells in injured islets [10,18]. Isolation of this subset by elutriation is best characterized by mitotic quiescence [11 –13], absence of lineage and progenitor markers [5 –10], and high affinity to the bone marrow [11,13,14]. The outstanding homing efficiency of stem cells to the bone marrow has dual implications in regenerative approaches to pancreatic islets. On one hand, preconditioning of the bone marrow reduces competitive migration of the cells to the injured islets, as also seen in a model of ischemic retinal injury [19,20]. Glycemic control attained by BM-H Fr25lin−PKHbright cells was markedly improved in the absence of irradiation, even without supporting R/O progenitors that have a rather modest impact on islet remodeling [10]. On the other hand, the biological affinity of these primitive precursors harvested from the adult bone marrow to their physiological site of residence can be used as a functional assay for enrichment of candidate stem cells for islet regeneration. Sequential homing to the bone marrow in primary recipients and participation in islet regeneration in secondary recipients indicate that the same cells migrate to both sites and retain the capacity to convert and produce insulin. Cells were harvested from the bone marrow 2 days after grafting into primary myeloablated recipients, a time point determined by prior studies of the kinetics of homing [11,14], but not necessarily the optimal time for use in tissue regeneration.
The vast majority of grafted hematopoietic cells are trapped in the liver, lungs [28,29], and other tissues, as emphasized by the presence of GFP+ hematopoietic cells at the islet perimeter, vasculature, and ducts [7,18]. Although more primitive hematopoietic progenitors are characterized by higher affinity for the bone marrow [21,28], the process of homing to and seeding in the bone marrow persists for several hours following intravenous transplantation, with dynamic entry and egress from the marrow space [30]. Conditioning increases substantially bone marrow retention of hematopoietic cells and engraftment, although significant numbers seed even in the absence of preparative conditioning without yielding effective hematopoiesis [28]. We cannot exclude the possibility that the cells were transcriptionally primed [31] in the bone marrow initially and then migrated to the islets, considering that hematopoietic precursors display dynamic entry into and egress from the marrow space in the early post-transplant period [21,30].
Interestingly, durable multilineage hematopoietic reconstitution [9,11,17] is the last among the versatile differentiation traits adopted by Fr25lin− cells [5 –8,10,18 –20]. This is likely because residents of the bone marrow, a compartment committed to hematopoiesis, should be restricted from engaging in hematopoietic differentiation to prevent extinction and preserve their primitive state. Although stem cells within the Fr25lin− subset are mitotically quiescent and can be, therefore, harvested from the marrow after 2 days by high fluorescence intensity [12 –14], they are responsive to short-lived inductive factors. For example, vascular endothelial growth factor (VEGF) signaling through VEGF-R1/Flt-1 and brain-derived and ciliary neurotrophic factors modulate the quantitative and qualitative contribution of Fr25lin− cells to retinal injury during the documented period of mitotic quiescence [19,20].
Approximately 25% of both elutriated fractions of Fr25lin− stem cells and R/O progenitors express VLA-4, a receptor binding multiple bone marrow stromal ligands [21], which are also abundant in chemically injured islets [22]. In the absence of this receptor, other adhesion mechanisms mediate bone marrow homing [32]; however, neither E-selectin nor CXCR4 was expressed at significant levels in Fr25lin− cells. Remarkable CXCR4 transcription suggests possible participation in stem cell migration to the bone marrow and islets expressing the cognate ligand stromal cell-derived factor-1 (SDF-1) [23 –25]. CXCR4 may be acutely expressed in transcriptionally primed stem cells [21] and may mediate migration to sites of injury expressing the cognate ligand [23 –25]. This possibility is consistent with demonstration that pretransplant expression of adhesion molecules does not correlate to the efficiency of homing [32 –34], and the demonstration that VLA-4 is acutely upregulated in the process of hematopoietic progenitor homing to and seeding in the bone marrow [21]. Additional receptors may be involved in common navigation to the bone marrow and islets, including VEGF-R1/Flt-1, which is expressed in Fr25lin− cells, modulates their incorporation and fate in the retina [20], and is involved in traffic to and from the bone marrow [35]. Fr25lin− cells are also transcriptionally primed to produce insulin-like growth factor-1 (Igf-1), which modulates cell migration both through enhanced motility [36] and induces SDF-1 expression in the injured tissues [37]. Both VEGF and IGF-1 modulate progenitor migration to and incorporation in hematopoietic and parenchymal compartments [38,39].
We found no evidence that the bone marrow serves a reservoir for sequestration of multiple tissue progenitors such as the endocrine pancreas [26,27]. Fr25lin− stem cells displayed competitive and sequential affinity to the marrow and injured islets, and acquisition of specific morphology and phenotype characteristic of selective retinal layers [8,19,20], argue in favor of in situ differentiation of multipotent stem cells. The absence of the pancreatic endocrine markers PDX-1 and insulin at the transcriptional and translational levels reinforces the contention that the bone marrow contains a subset of small-sized cells endowed with versatile differentiation capacity. Likewise, neither putative markers of pancreatic progenitors, such as Thy-1 [40], nor common markers characteristic of endocrine and neural differentiation, such as nestin, NeuroD, and Isl-1 [24,41,42], were detected in either one of the elutriated fractions at both transcriptional and translational levels [8]. Altogether, we could not detect specific markers of commitment to pancreatic endocrine differentiation and nonspecific markers of neural and pancreatic progenitors in BMC and Fr25lin− cells. A similar subset of very small embryonic-like cells (VSEL) [43] corresponding to a phenotype CXCR4+SCA-1+lin−CD45− [44] has been shown to perform diverse differentiation tasks in response to injury [45 –48]. The common characteristics of Fr25lin− and VSEL subsets include small size, absence of lineage markers, and low or absent CD45 expression; however, we could not document consistent expression of SCA-1 and c-kit associated with conversion to produce insulin [10]. Remarkable differences include positive identification of CXCR4 and SCA-1 in VSEL cells, but not in the Fr25lin− subset; adhesion molecules such as VLA-4 and activation markers such as CD34 and SCA-1 are rather unreliable in prospective isolation of progenitors and are acutely upregulated throughout the process of homing [2,21,32]. Therefore, the possibility to enrich by the affinity to the bone marrow compensates for the absence of phenotypic markers that may be used for prospective isolation [2,49,50]. Endogenous bone marrow definitely contributes to islet remodeling after injury by provision of various hematopoietic lineages participating in inflammation, clearance of debris, revascularization, and possibly units of β-cell regeneration, although largely insufficient to restore euglycemia [10]. In addition, various mesenchymal stromal cell types contribute directly and indirectly to islet remodulation after injury and regeneration through induction of endogenous precursors or direct differentiation [51 –54].
Following grafting of Fr25lin− cells into sublethally irradiated diabetic mice, 13% homing efficacy to the injured islets corresponds to incorporation of 1.3×105 in murine pancreas. Within the precision of these estimates, this frequency of incorporation is close to ∼105 differentiated stem cells required to reinstitute glycemic control, considering regeneration through neogenesis of 8% insulin-producing cells [7] in mice with ∼1.2×106 β-cells [55,56]. However, these figures exceed the total number of 4×104 grafted bone marrow-homed stem cells sufficient to restore glycemic control in the absence of radiation, indicating effective proliferation of the units of regeneration. Although we could not directly measure proliferation, it has been demonstrated in elegant single-cell experiments that Fr25lin− stem cells have the capacity to self-renew and regenerate multiple epithelial tissues [5].
In summary, we demonstrate that homing to the irradiated bone marrow is a characteristic of cells that convert to produce insulin and can be used as a functional assay to enrich stem cells, and reciprocally, irradiation reduces stem cell participation in islet regeneration by shifting competitive migration toward the bone marrow.
Footnotes
Acknowledgments
We thank Dr. Saul Sharkis and Dr. Michael Collector for the outstanding support, discussion, and conceptual contribution to this study. Funding was provided by generous grants from the Leah and Edward M. Frankel Trust for bone marrow transplantation, the Krieger Fund, Israel Academy of Sciences and Humanities (1371/08 and 1189/12), the Eldor-Metzner Clinician Scientist Award, and the Israel Ministry of Health (3-3741).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.