
Abstract
Cardiomyocytes from human pluripotent stem cells (hPSC-CMs) are increasingly used to model cardiac disease, test drug efficacy and for safety pharmacology. Nevertheless, a major hurdle to more extensive use is their immaturity and similarity to fetal rather than adult cardiomyocytes. Here, we provide an overview of the strategies currently being used to increase maturation in culture, which include prolongation of time in culture, exposure to electrical stimulation, application of mechanical strain, growth in three-dimensional tissue configuration, addition of non-cardiomyocytes, use of hormones and small molecules, and alteration of the extracellular environment. By comparing the outcomes of these studies, we identify the approaches most likely to improve functional maturation of hPSC-CMs in terms of their electrophysiology and excitation–contraction coupling.
Introduction
H
hPSC-CMs, however, remain immature in culture and resemble heart cells of mid-gestation human foetuses. They show disorganized sarcomeres, small forces of contraction, and small action potentials (APs) compared with adult working cardiomyocytes. This limits their use in studying human cardiac physiology and pathology in the laboratory, while in the context of tissue repair, their immature properties such as spontaneous activity and slow conduction may cause lethal arrhythmias after transplantation in the heart [8]. Inducing hPSC-CM to a more adult state would significantly increase their value. Various strategies for this have already been described [9].
Here, we provide an overview of these studies that have included (1) increasing time in culture [10 –14]; (2) applying electrical stimulation to cause continuous contractile and electrical activity [15 –19]; (3) adding chemicals or small molecules [20 –22]; (4) applying mechanical stretch [23,24]; (5) co-culture with non-cardiac cells [25]; (6) growth as three-dimensional (3D) tissues [18,22,26]; and (7) adjusting composition, stiffness, and topography of extracellular environment [27 –29]. These strategies mainly mimic cardiac physiology; however, artificial methods in which the expression of key genes is modified have been applied as well [15,30,31]. The discussed approaches have often been identified in isolated and cultured rodent cardiomyocytes, in which the reversion from a mature to an immature state is a common phenomenon [32]. Where relevant, results of these studies are discussed as well. Before considering maturation strategies, however, we first describe the distinguishing features of mature (adult) and immature (fetal) cardiomyocytes that will allow benchmarking of maturation state of the derivative hPSC-CMs.
Distinguishing Features of Immature and Mature Cardiomyocytes
Morphology
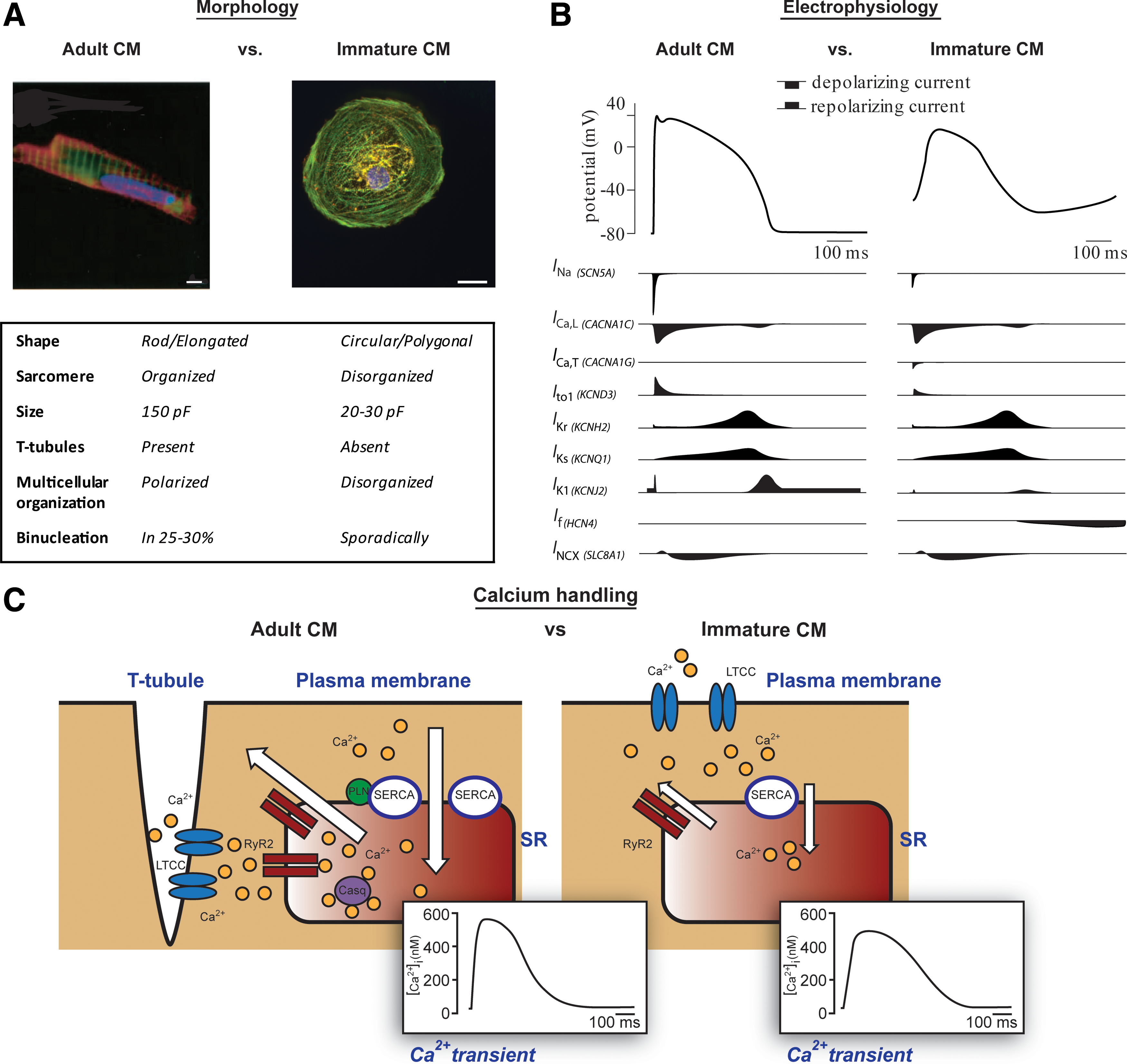
Adult cardiomyocytes are elongated and rod shaped with an aspect ratio (length to width) of 5:1, while fetal cardiomyocytes are typically round- or polygonal shaped [33,34]. As human cardiomyocytes mature, they increase in size due to physiological hypertrophy [35]. Early hPSC-CMs (10–15 days after onset of differentiation) are mostly round and small, although rod-shaped cells have been described [36]. Later (>50 days in culture), hPSC-CMs become more elongated although they remain smaller than adult cardiomyocytes [36]. In addition, adult cardiomyocytes in the heart are longitudinally aligned, facilitating fast electrical conduction and efficient muscle contraction via connecting intercalated discs [37]. However, fetal cardiomyocytes and monolayers of hPSC-CMs are chaotically organized. The number of nuclei per cell also distinguishes adult and fetal cardiomyocytes: 25%–30% of adult cardiomyocytes in the heart are binucleated independent of age, while fetal cardiomyocytes are almost exclusively mononucleated [38 –40]. Ploidy increases during maturation [39], and may be a better marker of maturity. hPCS-CMs are largely mononucleated, showing binucleation only sporadically [36].
Sarcomere
The sarcomere is the contractile unit of cardiomyocytes that consists of proteins that form thick and thin filaments. By electron microscopy, adult cardiomyocytes show highly organized and aligned sarcomeres with different landmarks corresponding to different functional units: Z-discs mark sarcomere borders; I-, H-, and A-bands mark different areas of thick, thin, and overlapping filaments; and M-bands form the central line [41,42]. In the human fetal heart, this organization develops gradually during gestation, with Z-discs and I-bands forming first, followed by H-, A-, and M-bands [41]. This process of sarcomerogenesis also occurs in maturing hPSC-CMs; however, in most cells, only Z-discs and I-bands are formed [36]. Sarcomere length, the distance between two Z-discs, is another indicator of cardiomyocyte maturation: Adult cardiomyocytes contain considerably longer sarcomeres than hPSC-CMs and fetal cardiomyocytes [11,26,34,38].
Proteins assembled in the sarcomeres are also differentially expressed in immature and mature cardiomyocytes: Both myosin heavy chain (MHC) and myosin light chain (MLC) protein isoforms change during maturation. In the adult ventricle, it is mainly the β-MHC (encoded by MYH7) that is expressed, with very low levels of the atrial isoform α-MHC (encoded by MYH6) [43]. This abundance of β-MHC over α-MHC is already clear in early fetal stages and shows a further increase at later gestational ages [43]. Interestingly, the ratio of β-MHC/α-MHC in hPSC-CMs is different from fetal and adult ventricular cardiomyocytes and changes little over time. This may be because hPSC-CMs are often a mixture of atrial and ventricular cardiomyocytes [11,12,26,44]. The two predominant isoforms of MLC are MLC2a and MLC2v, encoded by the genes MYL7 and MYL2. Again, an isoform switch occurs during maturation. MLC2a, the predominant isoform in adult atria [45], is expressed in human fetal ventricle although it gradually decreases with gestational age [45], when MLC2v is the isoform almost exclusively expressed [46]. In hPSC-CMs, both isoforms are expressed [26]. Again, a mixture of cardiomyocyte subtypes in the population may be the reason that there is no predominance of one over the other. Titin, a sarcomeric protein with many spliced isoforms, anchors between the Z-discs and the M-lines of the sarcomere and plays a crucial role in sarcomeric elasticity [47]. In fetal cardiomyocytes and hPSC-CMs, it is mainly the N2BA isoform that is expressed [48,49], while in adult cardiomyocytes, N2B is the most abundant isoform [48].
Electrophysiology
The cardiac AP governs cardiomyocyte electrical behavior. AP characteristics are unique for each cardiomyocyte subtype (atrial, ventricular, pacemaker, Purkinje). The sum of the activity of various depolarizing and repolarizing currents through different ion channels is responsible for the cardiac AP (Fig. 1B) [50]. Adult and fetal cardiomyocytes differ in the availability of ion channels on their cells, which results in different AP profiles. The AP is initiated by the large, rapid influx of Na+ (I Na) through Na+ channels, resulting in fast depolarization of the membrane; this is called the AP upstroke or phase 0. Subsequently, there is a brief repolarizing phase (phase 1), resulting from efflux of K+ caused by activation of the transient outward potassium current (I To1). Next, inward flow of Ca2+ (I Ca,L) through L-type calcium channels leads to the plateau phase (phase 2). Finally, the membrane repolarizes to its original state due to activation of the rapid and slow delayed rectifier K+ channels (conducting the I Kr and I Ks, currents, respectively) in phase 3 of the AP. Adult ventricular and atrial cardiomyocytes also exhibit phase 4 in which the resting membrane potential (RMP) does not change. This is due to the rectifying K+ current (I K1), which stabilizes the membrane potential at the reversal potential of K+, that is, −85 mV [50]. In hPSC-CMs, I K1, if even detectable, is only a small fraction of that in adult cardiomyocytes [10,51,52], and KCNJ2 mRNA expression (encoding the α-subunit of the channel) is correspondingly low [53]. Accordingly, the RMP is less negative in hPSC-CMs compared with adult cardiomyocytes (−50 to −60 mV vs. −85 mV) [33,51,52,54,55]. Moreover, low or absent I K1, together with the pacemaker current (funny current, I f), which is very low in adult ventricular cardiomyocytes, causes gradual diastolic depolarization and consequent spontaneous contractile activity in hPSC-CMs [10,11,15,56]. Because of the more depolarized state of hPSC-CMs, functional availability of Na+ is reduced [50], resulting in a slow upstroke of the AP [51,52,54,55], although low expression of sodium channels (encoded by SCN5A) in early hPSC-CMs might contribute to this as well [53]. Similarly, I To1 is inactivated at more positive membrane potentials, leading to less pronounced phase 1 repolarization [57]. Although the latter demonstrates similar expression and current density levels in hPSC-CMs and adult cardiomyocytes [57], I To1 increases during development and is also considered a sign of maturation [10,58,59]. The other main repolarizing currents, I Kr and I Ks, are present in hPSC-CMs at similar levels to adult cardiomyocytes [52,60,61]. To date, current densities in human fetal cardiomyocytes have not been reported but data in other animals suggest that I Ks increases during development [62].
Qualitative comparison of morphological
Two types of calcium channels are present in the human heart: L-type and T-type. While the functional presence of the T-type calcium current (I Ca,T) is typical of fetal cardiomyocytes [63], atrial and ventricular cardiomyocytes of the adult heart do not exhibit I Ca,T; here, I Ca,T is restricted to the conduction system [64]. For hPSC-CMs, the presence of I Ca,T has been debated: I Ca,T was reported in a subset of hPSC-CMs in one study [12], while in another, the current was not detected [52]. By contrast, I Ca,L is observed at similar densities in both hPSC-CMs and adult cardiomyocytes, although exact values have varied between hPSC-CMs [10,51,52,65]. There are no reported data from human fetal cardiomyocytes.
Conduction velocity
Given the reduced availability of sodium channels, propagation of the electrical signal, to which I Na is a major contributor [66], is relatively slow in monolayers of hPSC-CM (10–20 cm/s compared with 60 cm/s in adult human left ventricle) [67,68]. However, for a high conduction velocity, other contributors are of importance. Density and composition of gap junctions, for example, through which the electrical signal is conducted, is also critical. With respect to the gap junctions, fetal-, hPSC-CMs, and adult cardiomyocytes do not show clear differences in the expression of connexin 43 (GJA1), which forms gap junctions in the ventricle [11,69]. Conduction velocity is, however, also determined by the localization of gap junctions and sodium channels. In adult cardiomyocytes, gap junctions and sodium channels accumulate at the intercalated discs located at the short edges of two neighboring cells, resulting in a faster conduction in the longitudinal direction compared with transversal [70]. However, in fetal cardiomyocytes and hPSC-CMs, gap junctions are distributed around the circumference of the cells on all sides of the membrane [71], rather than at the ends. Finally, cell size, which is positively correlated with conduction velocity [72], may be an important factor contributing to slow conduction velocity in hPSC-CMs. Of note, other factors not related to cell maturation, such as fibrosis and non-cardiomyocyte cell populations, also impact conduction velocity [66]; therefore, this parameter should be taken into consideration when using this as a marker of maturation.
Calcium handing and excitation–contraction coupling
After an AP, influx of Ca2+ via L-type channels triggers the release of Ca2+ from the sarcoplasmic reticulum (SR) through ryanodine receptor (RyR) channels, a process known as Ca2+-induced- Ca2+ release [73]. For relaxation, Ca2+ is pumped back into the SR via sarco/endoplasmic reticulum Ca2+-ATPase (SERCA) and is extruded from the cell via the Na+-Ca2+ exchanger (NCX). This Ca2+ transient modulates cardiac contraction. Ca2+ release, and extrusion is efficient in adult cardiomyocytes [74]; however, in fetal cardiomyocytes and hPSC-CMs, kinetics are slow and amplitudes are small [75 –77] (Fig. 1C). In hPSC-CMs, Ca2+ intrusion may even be completely dependent on sarcolemmal influx (through L-type calcium channels) and not through release from the SR [78,79], although SR-mediated Ca2+ release has been described in most studies [75 –77,80,81]. Amplitudes of Ca2+ transients also decrease with pacing frequency in hPSC-CMs in contrast to adult cardiomyocytes, in which they increase [79,82]. Different components of the Ca2+ handling system are missing in fetal cardiomyocytes and hPSC-CMs. One of the most important are transverse tubules (T-tubules), invaginations in the membrane where L-type Ca2+ channels are concentrated near RyR channels. In adult cardiomyocytes, this compartmentalizes Ca2+, leading to fast excitation–contraction coupling (ECC) and synchronized contraction in multiple sarcomeres [83]. Because T-tubules are absent and expression of Ca2+ handling proteins is low in hPSC-CMs and fetal cardiomyocytes, ECC is slow. Although the expression of SERCA (encoded by ATP2A2) is generally high in hPSC-CMs and fetal cardiomyocytes (although lower than adult ventricular cardiomyocytes) [76,78], other important proteins involved in mediating uptake and release of Ca2+ from the SR, such as calsequestrin (CASQ2), ryanodine receptor type 2 (RyR2), and phospholamban (PLN), are expressed at low levels or even absent [53,84].
As for NCX channels, expression increases during human development, after which it decreases postnatally [85]. Expression of NCX in hPSC-CMs and adult cardiomyocytes is comparable, although current density increases over time in hPSC-CMs, as in fetal development [65].
Immaturity of hPSC-CM is also reflected in their small contraction forces. Active forces generated in 3D hPSC-CM tissue constructs are ∼0.1–0.5 mN/mm2 [24,86], compared with 10–50 mN/mm2 in adult ventricular cardiomyocytes [87,88]. Passive forces (resulting from myofibrillar compliance) also increase with maturation and in adult cardiomyocytes, they increase with increasing beating frequency (positive force-frequency relationship) [88]. Due to insufficiencies in Ca2+ release and uptake, there is a negative force–frequency relationship in fetal cardiomyocytes and hPSC-CMs [89].
Adrenergic stimulation
Fetal cardiomyocytes show chronotropic responses (increased beating frequency) in response to beta-adrenergic stimulation (norepinephrine) early in development [90]. This is also observed in hPSC-CMs [91]. The effects of this on contractile force (inotropy) and relaxation rate (lusitropy) are inconsistent, with some studies showing a response on beta-adrenergic stimulation and others none [91 –93]. If present, though, inotropic and lusitropic responses are not as robust as in adult cardiomyocytes [92].
Response to alpha adrenergic stimulation, which plays a role in physiological hypertrophy in the human heart [94], has been described for both human menbryonic stem cell (hESC)-CMs [95,96] and hiPSC-CMs [95]. Interestingly, a robust hypertrophic response on alpha-adrenergic stimulation is evoked in hESC-CMs [95,96], in contrast to hiPSC-CMs, where it does not appear to take place [95]. Compared with adult and fetal human cardiomyocytes, both hESC-CMs and hiPSC-CM lack expression of the main cardiac alpha-adrenergic receptor ADRA1A, suggesting that downstream signaling proteins are differentially functional in these cells.
Metabolism
Energy demand in the beating heart is high at rest compared with other tissues and increases dramatically during physical activity. Energy production pathways are therefore of utmost significance for working cardiomyocytes. Reflecting this, the mitochondrial-to-cell volume ratio in cardiomyocytes increases by 20%–30% during development [97]. From the increasing density and organization of mitochondria, substrate utilization changes from glucose and lactate in the fetal heart primarily to fatty acids in the adult heart [98,99]. Mitochondrial biogenesis also increases over time in hESC-CMs, mimicking that in heart development, primarily driven by the transcriptional co-activator PGC-1α [100]. In terms of their bioenergetic profile, it seems that even in immature hPSC-CMs oxidative phosphorylation accounts for the majority of ATP production. In terms of substrate utilization, one report has suggested that under baseline conditions, beta-oxidation is only a minor contributor to respiration but when energy demand is stimulated, it becomes more prominent [101]. Under the right physiological conditions, therefore, hPSC-CMs may well substantially utilize fatty acids. Maturation-related events such as the regulation of genes important for fatty acid uptake or oxidation may play an important role in determining this activity.
Promoting Maturation of hPSC-CMs in Culture
Having considered the salient features of cardiomyocyte maturation, we now discuss how these might be enhanced in culture. A summary of the published functional and electrophysiological outcomes is given in Table 1. Where available, quantitative effects on AP characteristics are provided in Table 2.
Observed effects of each approach are in comparison to the control culture condition without the proposed strategy. Data are from hPSC-CMs, unless stated otherwise.
CV, conduction velocity; MDP, maximum diastolic potential; RMP, resting membrane potential; Vmax, maximum upstroke velocity; APD90%, action potential duration at 90% of repolarization; ↑, increase; ↓, decrease; hPSC-CMs, human pluripotent stem cell-derived cardiomyocytes.
The condition in which AP characteristics were improved followed by its control comparison is given for each reference with the asterisk, indicating statistical significance (P<0.05).
“∼” indicates an approximate value acquired from the mean representation (bar or other) from the original article.
iPSC, induced pluripotent stem cell; AP, action potential.
Time in culture
Intuitively, it was expected that maturity would increase with time in culture. Sartiani et al. [10] compared current densities and kinetics of several ion channels, and the expression of their corresponding genes in cardiomyocytes was kept for 15–30 days or for 55–110 days in culture. I K1 and I To1 densities clearly increased over time, whereas the kinetics of I f activation were slower in cardiomyocytes during prolonged culture. I Ca,L and I Kr densities were unchanged. By contrast, Ivashchenko et al. reported increased I Ca,L density and upregulation of the corresponding gene CACNA1C, after 37 days compared with 80 days [12]. Furthermore, increased I Na density, upregulation of GJA1 (connexin-43), SCN5A, and KCNJ2, and increased MYH6/MYH7 gene expression ratio were observed over time, although a decrease would have been expected for the MYH6/MYH7 ratio. Otsuji et al. [14] reported similar effects on I Ca,L and I Na densities in hESC-CMs at 28 days compared with 231 days. The expression of sodium channel genes, SCN5A and SCN1B (encoding the beta-subunit of the sodium channel) were clearly upregulated along with CACNA1C. The upstroke velocity of the AP was thus correspondingly increased. However, no increase in the I K1 density was observed and no upregulation of its alpha-subunit gene KCNJ2. Lundy et al. [11] also examined the effect of long-term culture and primarily focused on morphological maturation. hPSC-CMs were plated on two different substrates in succession and designated as early (20–40 days in culture) or late stage (80–100 days in culture). Multinucleation, sarcomere length, cell size, and elongated shape increased and MYH7, MYH6, GJA1, and SERCA2 gene expression was upregulated over time in culture. Although not expected, HCN4 (the gene primarily underlying the pacemaker current, I f) was also upregulated. AP upstroke velocity and amplitude were increased, and RMP became more negative in late versus early hPSC-CMs. Lastly, the Ca2+ handling of late-stage cardiomyocytes was characterized by higher upstroke and decay velocities, although the Ca2+ transient amplitude was unchanged. In another study focusing primarily on the ultrastructural phenotype, Kamakura et al. [13] reported gradual changes in sarcomere organization in hiPSC-CMs maintained over a 1 year in culture. Thirty day hiPSC-CMs showed only Z- and I- bands, while cardiomyocytes between 30 and 90 days developed sarcomeres that included Z-, I-, and A- bands. Impressively, 360-day hPSC-CMs also showed the presence of M-bands. However, there was considerable variability among the cells analysed.
In summary, some evidence suggests maturation of hPSC-CM over time in culture but even if prolonged, an adult cardiomyocyte phenotype is never acquired.
Electrical stimulation
hPSC-CMs usually exhibit spontaneous contractility. However, this electrical activity is usually irregular, and the frequency gradually decreases over time in culture [11]. In isolated and cultured rodent cells, electrical stimulation is known to improve cell-to-cell coupling and alignment [17]. Remodeling of these cells (“de-differentiation”) to an immature phenotype in culture is prevented (and partially reversed) by electrical stimulation. Several studies have shown beneficial effects of electrical stimulation on hPSC-CM maturation evidenced by increased I K1 and, consequently, lower RMP [15,18], improved Ca2+ handling (increase in amplitude, upstroke, and decay rate of the Ca2+ transient) [15,16], increased contractile forces [19], and a higher degree of sarcomeric organization [15,16,18,19]. Most studies used a physiological pulse rate of 1 Hz, but higher stimulus frequencies (1.5–2 Hz) generate a more robust response [19]. In one study, the pacing frequency was gradually increased [18], which led to a more robust response than stimulation at lower frequencies. In general, the mechanisms underlying increased maturation by electrical stimulation are indirect, as pacing generates cyclic mechanical stretch, leading to cardiomyocyte remodelling. However, the effects could also be direct: For example, electrical activity was shown to alter gene transcription to a more mature profile in the presence of the contraction inhibitor blebbistatin [102]. It has been suggested that intracellular generation of reactive oxygen species (ROS) on electrical stimulation may contribute to increased maturation [103]. High amounts of ROS are generally considered detrimental, but limited amounts of ROS can function as intracellular second messengers and determine cellular differentiation and maturation fate [104]. Although the production of ROS has been demonstrated [103], there are no studies describing direct effects on maturation or specific signaling pathways. Moreover, it is unclear whether the amount of ROS generated, especially in the case of continuous pacing, actually stimulates maturation or does the reverse. The latter might require antioxidants in the culture medium or continuous perfusion of the culture medium.
Mechanical strain
The heart continuously undergoes mechanical stress, the result of hemodynamic load (cyclic stretch), physical interaction with extracellular matrix (static stretch), and laminar blood flow (shear stress) [105]. It is believed that while undergoing mechanical stretch, cells are forced to change shape through transduction of the mechanical forces via the cytoskeleton and align in the direction of the applied traction, altering their gene expression as a result [105,106]. Structural and functional changes follow [107,108], including induced expression of vascular endothelial growth factor, tumor necrosis factor-α, and insulin-like growth factor (IGF-1) [109,110]. This occurs in cultured cells [111,112] and in the intact heart [113], making mechanical stress an obvious approach that may be beneficial for maturation of hPSC-CMs. To mimic cardiac hemodynamic load, cyclic stretch has been applied in several studies, demonstrating an increased rate of maturation in hPSC-CMs [22 –24,114,115]. In both hESC- [23,24] and hiPSC-derived CMs [24], cyclic stretch and strain at rates of 1–2 Hz promote maturation measured structurally (cell elongation, higher degree of sarcomeric organization, and higher density of gap junctions) and functionally (faster Ca2+ transients). The functional effects on Ca2+ handling are reflected in upregulation of RYR2 and SERCA2 [24]. Mihic et al. [23] also showed increased expression of ion channel genes, including KCNJ2, which could lead to increased I K1 and more negative membrane potentials, although this was not assessed. An alternative is to gradually increase static stretch [22]. This was reported to improve cellular and sarcomeric organization compared with cyclic stretch or increased passive and active forces. However, other functional parameters of maturation were not assessed in this study.
Chemically induced maturation
Among the most promising reagents to date identified as inducing maturation of hPSC-CM are thyroid hormones, which are known to play a crucial role in cardiac development [116]. For example, reduction in levels of triiodothyronine (T3) during development induced by thyroidectomy of pregnant females results in fewer binucleated cardiomyocytes in the fetal heart and lower SERCA2 expression, which is rescued by T3 replacement [117]. Furthermore, T3 converts the sarcomeric protein titin from the fetal N2BA- to the adult N2B isoform in cultured embryonic rat cardiomyocytes [21]. Moreover, reduced expression of SERCA2 and restricted conversion of β-MHC to α-MHC is observed during development of hypothyroid mice [118]. Although α-MHC downregulation would usually be expected during maturation, increased α-MHC/β-MHC ratio and higher SERCA expression are observed in hPSC-CMs, where T3 also reduces expression of the fetal genes NPPB [encoding brain natriuretic peptide (BNP)] and HCN4 [12,119]. Structurally, T3 was reported to increase cardiomyocyte size, induce cell elongation, and increase sarcomere length [119]. In this study, Ca2+ handling and contractility also showed remarkable improvements, as evidenced by increased Ca2+ transient upstroke and decay velocities. With respect to ion channels, only mouse ESC-CMs have been studied to date. Here, addition of T3 resulted in more negative RMPs, accompanied by increased expression of KCNJ2. Moreover, much as in hPSC-CMs described earlier, Ca2+ handling improved, reflecting changes in expression of SERCA2 and RyR2 [120].
Ascorbic acid, which stimulates cardiomyocyte differentiation [121], has also been shown to enhance maturation in mouse iPSC-CMs. Treated cells showed improved calcium handling at baseline and increased responsiveness to β-adrenergic and muscarinic stimulation [122]. In hiPSC-CMs, ascorbic acid enhanced contraction forces and sarcomeric organization and led to increased intercalated disc formation, titin expression, collagen deposition, and metabolic activity [22]. Ascorbic acid promotes collagen synthesis through the MEK-ERK1/2-pathway, which stimulates proliferation of cardiac progenitor cells [122]. Both the composition and amount of extracellular matrix as well as the differentiation efficiency can influence the degree of maturation [123].
Neuregulin-1β (NRG-1β), which agonizes the Notch signaling pathway and plays a role in cardiomyocyte subtype specification, might also promote cardiomyocyte maturation. NRG-1β has been shown to promote hESC-CM differentiation toward ventricular-like cells [124], while its inhibition leads to a nodal-cell phenotype. In mouse iPS-CMs, NRG-1β increases AP upstroke velocities, reduces the RMP, and upregulates MYH6, RYR2, MYL3, and SERCA2A as well as genes involved in fatty acids metabolism (PDK4, CD36) [20]. Genes involved with glycolysis are downregulated (SLC2A1, SLC2A4).
Co-culture with non-cardiomyocytes
Kim et al. [25] provided among the earliest evidence that co-culture with non-cardiomyocytes improved electrophysiological characteristics and calcium handling properties of hESC-CMs. Using a puromycin resistance cassette in the promoter region of α-MHC to select pure cardiomyocyte populations from differentiating hESC followed by remixing, the cardiomyocytes grown as spheroids without other cell types present were more immature than those from spheroids containing non-cardiomyocytes: AP upstroke velocities and amplitudes were lower, and RMPs less negative. Overall though, the AP characteristics remained relatively immature compared with other studies. The study did not elucidate the cell specificity of these non-cardiomyocytes. Ou et al. [125], on the other hand, used rat embryonic cardiac fibroblasts co-cultured with mouse ESCs and found an increase both in the efficiency of the cardiomyocyte differentiation and in the organization of sarcomeric structures with clear gap junction patterns. The expression of the GJA1 gene was also increased in this co-culture system compared with mouse ESC-derived cardiomyocytes that were not co-cultured. Similar findings were described in a study by Blin et al. [126], in which cardiac progenitor cells derived from primate (Rhesus) ESCs were co-cultured with human atrial cardiomyocytes and cardiac fibroblasts. The resulting cardiomyocytes had increased sarcomere size and organization and expressed connexin 43 on their membrane as well as MLC2v (in 80% of the cells) and β-MHC (in 50% of the cells). Thavandiran et al. also investigated the effect of cardiac fibroblasts on hESC-CMs by mixing them in co-cultures in different ratios. They examined morphology, gene expression, sarcomere structure, and conduction in engineered heart tissue structures called “cardiac micro-wires” [127] and found that a 3:1 ratio of hESC-CMs to cardiac fibroblasts yielded cardiac micro-tissues with the highest atrial natriuretic factor (ANF), BNP, MYL7, MYL2, and MYH7/MH6 expression levels. Z-disks and H bands were present in these tissue structures and most impressively, they displayed conduction velocities comparable to those of an adult human heart.
3D culture
Cardiomyocytes tend to form 3D structures over time in culture, a phenomenon that is also observed in differentiating hPSC-CMs [128]. This has been the basis constructing cardiac tissues with a predetermined 3D structure using either primary cardiomyocytes [129 –131] or, more recently, hPSC-CMs [17 –19,23,24,26,86,127,132 –134]. The most common approach is to cast molds, which determine the 3D form. Hydrogels of collagen type I are placed in these moulds with cardiomyocytes and attached between two anchor points that can move cyclically back and forth to induce remodeling and alignment of the cardiomyocytes in myocardial conduits [22,24]. Other techniques include circular casting molds [19,134], porous sponges [17,132], and stacked cardiac sheets [133].
Although the original goal of this “tissue engineering” was to create tissue replacement for the heart, it soon became clear that growth in 3D conduits affects phenotype and is a better mimic of real myocardium than culture on plastic [135]. Moreover, 3D tissues allow easy measurement of contractile forces.
Most information on the effects of 3D growth has been derived using cardiomyocytes from chicken embryos [129] and rodents [130,131]. As stated earlier, primary cardiomyocytes dedifferentiate (remodel) in culture but loss of their sarcomeric organization and contractile force (basal and after beta-adrenergic stimulation) is attenuated in 3D [130]. Prolonged culture under these conditions almost caused terminal differentiation of neonatal rat ventricular myocytes [136].
3D tissue engineering of hPSC-CMs has been reported [18,19,22 –24,26,86,132 –134], although not all of these studies made the direct comparison with two-dimensional (2D) models in terms of maturation rate. Moreover, 3D cultures are often combined with other strategies that promote maturation, such as mechanical stretch or electrical stimulation. Where directly compared with 2D monolayers, results have been unambiguous. One recent study showed gene expression of more mature cells in 3D tissues, including downregulation of fetal genes NPPA, NPPB, and MYH6, upregulation of KCNJ2, increased I K1 density, more negative RMPs, higher AP upstroke velocities, and less automaticity [18]. Moreover, 3D tissue hPSC-CMs had larger surface areas and lower proliferation rates, again indicating increased maturity. A second study demonstrated increased sarcomere length, faster conduction velocities, increased expression of the Ca2+ handling genes SERCA2 and CASQ2, and increased β-MHC/α-MHC and MLC2v/MLC2a ratios [26]. Of note, although the addition of isoproterenol had inotropic effects, relaxation rate was not increased, suggesting that Ca2+ handling was not functionally mature.
The benefits of 3D culture on cardiomyocytes could partially result from increased mechanical stretch. In most formats, the tissue is attached to two static rods. As the cardiomyocytes are spontaneously active, the static rods provide the mechanical load on each contraction and thereby force the cells to align properly. As it is clear that extracellular architecture (both on macro- and on micro-scale) affects cellular function and fate robustly, it is not possible to attribute the observed effect exclusively to mechanical load. As discussed next, adjustments of the extracellular microstructure in culture is also considered a promising approach to increase maturity of hPSC-CMs.
Extracellular substrates
Cell culture substrates are not inert. Maturity of hPSC-CMs can be induced through the substrate by (1) coating with extracellular matrix proteins that can act directly as messenger molecules, by either biochemical or mechanical signaling [137 –139]; (2) changing/modulating the intrinsic elasticity or stiffness of the substrate, providing mechanical load not present when cells are cultured on rigid glass slides [28,140,141]; and (3) changing/modulating surface topography, at both the nano- and micro-scale, providing mechanical cues and forcing cardiomyocytes to align and elongate [27,29,142].
The effect of extracellular matrix in signaling was demonstrated in a study in which Matrigel, a mixture of laminin, collagen IV, and proteoglycan, was added on top of differentiating hPSC-CMs [137]. This extracellular matrix “sandwich” not only resulted in higher efficiencies of cardiac differentiation but also promoted cardiomyocyte maturation. Electrophysiologically, the resulting cardiomyocytes were comparable to those from late-stage aggregate cultures (embryoid bodies), suggesting their increased maturity. The rationale behind this approach is that cardiomyocytes could acquire a more realistic phenotype in an extracellular environment that closely resembled the native human heart. Here, the extracellular matrix consists of laminin, fibronectin, collagen, and proteoglycans synthesized by cardiac fibroblasts [143]. This mixture (also known as “cardiogel”) can be artificially generated in culture [143]. Although beneficial effects are observed in cultured ventricular rat cells [144,145], similar effects have not been demonstrated in hPSC-CMs. Only one study evaluated the effects of cardiogel on mouse ESC-CMs by a direct comparison with Matrigel [123], but no clear benefit on maturation was observed.
In the heart, the extracellular matrix is a dynamic, compliant structure, entirely different from rigid glass or plastic surfaces used in standard tissue culture. By adjusting culture substrate hardness (and not only the chemical composition of the extracellular matrix per se) such that it resembles that in real heart tissue, it was predicted that maturation would be induced. This has been explored in different cell types, usually by means of polyacrylamide hydrogels of varying stiffness. In most studies using cardiomyocytes other than hPSC-CMs, substrate stiffness in the physiological range (Young's modulus 10–20 kPa) resulted in well-organized myofibrillar structures [140,141], high twitch power, calcium amplitudes [146], and morphologically elongated cells [147], although one study using adult rat ventricular myocytes demonstrated optimal sarcomeric structure and calcium handling in soft (7 kPa) or very stiff (255 kPa) substrates [148]. For hPSC-CMs, however, results are less clear. In one study, increasing stiffness to 100 kPa led to increased contraction forces in hiPSC-CM, in line with the Frank–Starling principle. Cell morphology and sarcomeric organization, on the other hand, clearly affected by substrate stiffness in other cell types, remained immature as under standard conditions [28]. In this study, however, functional characterization by electrophysiology and Ca2+ handling was not carried out. Another study showed that morphology of hESC-CMs was affected by increased substrate stiffness [149], with greater cell spreading and the formation of myofibrillar stress fibers. These stress fibers are also observed in neonatal rat cardiomyocytes [140] at stiffness above physiological values and are detrimental for sarcomeric function.
By forcing cells to grow on a topographically predetermined substrate, for example, micro-grooved patterns with widths of several micrometers, cardiomyocytes align in that pattern and adjust their shape to adopt typical length-to-width ratios observed in mature ventricular cardiomyocytes [29]. The resulting cell–cell and cell–matrix adhesions, which also physiologically resemble those of native heart tissue, influence functional phenotype, as evidenced, for example, by neonatal rat cardiomyocytes; here aligned cells show polarized distribution of the proteins N-cadherin and connexin43 in intercalated discs [150]. In hPSC-CMs, this also improves maturation as evidenced by faster calcium kinetics [27,142] and increased active force [151].
Ectopic expression or direct regulation of genes associated with cardiomyocyte maturation
Another option to induce maturation is to express ectopically key genes normally present in adult cardiomyocytes to see whether they actually drive functional maturation in culture. One such gene is KCNJ2 that encodes the alpha subunit of Kir2.1 channel (I K1) and determines the RMP. Since I K1 is the main current that determines the RMP of the cardiac AP, Lieu et al. [15] used adenovirus to transfer Kir2.1 to embryonic bodies of hESC during cardiac differentiation. The resulting cardiomyocytes showed decreased spontaneous activity and hyperpolarized RMP values (−80 mV vs. −60 mV in controls). However, Ca2+ handling was not improved (slow kinetics, small calcium transient amplitudes) and the expression levels of key sarcomeric proteins were even significantly decreased compared with controls. In a similar approach, Liu et al. [30] forcibly expressed calsequestrin in hESC-CMs. The Ca2+ transient amplitudes were significantly increased, and kinetics improved (higher upstroke and higher decay velocities). These outcomes were linked to neither increased I Ca,L nor cell hypertrophy, leading to the speculation that if the expression of other proteins involved in Ca2+ handling such as junctin, triadin, RyR2, and SERCA2 were increased, the cells might mature even further. Other strategies might also include the use of micro-RNAs. Fu et al. [152] observed that lentiviral-mediated transduction of miR-1 caused the RMP of hESC-CMs to become significantly more negative compared with controls. In addition, there was an improvement in the Ca2+ transient amplitude and kinetics and upregulation in the expression of other ion channel genes with the exception of HCN4. However, upstroke velocities remained as controls.
Transcriptional activity is, in part, determined by epigenetic state; this encompasses histone modifications and DNA methylation and is highly dynamic during cardiac differentiation [153]. One study demonstrated that by temporarily adding the histone deacetylase inhibitor valproic acid to hESC-CMs, the epigenetic state of the promoters of Ca2+ handling and ion channel encoding genes changed, resulting in increased gene expression, as well as increased cell size, resembling hypertrophic growth [31]. Since epigenetic regulation is finely tuned, both repressive and stimulatory effects may result from histone modification. It is, therefore, doubtful whether it would be possible to alter transcriptional activity specifically by histone deacetylase inhibitors to affect only those genes involved in maturation.
Interfering with I K1 function to drive the RMP of hPSC-CMs to more negative values has proved more challenging than expected. An alternative in silico approach using dynamic patch clamp has recently been developed and utilized to address this issue. Dynamic clamp integrates a computer-modeled current recording to live cells on patch clamp and allows the impact of this particular current on the overall AP output of the cell to be assessed in real time. Using a variation of this approach, Bett et al. [154] artificially injected I K1 current to paced hiPSC-CMs and observed tremendous hyperpolarization of the RMP as well as an increase in the amplitude and upstroke velocity of the AP, resembling values highly similar to adult ventricular cardiomyocytes.
Concluding Remarks: Past, Present, and Future
Despite their limited maturation, hPSC-CMs are already proving useful for safety pharmacology, as models of cardiac disease and drug screening. The US Food and Drug Administration (FDA) proposed in a directive in 2013 that it intended within 2 years to require new drugs to be tested for their effects on all ion channels in human cardiomyocytes. hiPSC-CMs are the cell type of choice for this; despite their immaturity, the FDA considered they were already near appropriate for this purpose. hiPSC lines have been generated from patients affected by cardiac diseases that until recently could not be studied appropriately in existing animal models because of species differences in cardiac physiology [155]. Efforts are now focused on enhancing the functional characteristics of hPSC-CMs to improve their accuracy and robustness as models. To date, inducing maturation (and mimicking aging) of hPSC-CMs is still the most challenging aspect in the field. The approaches described in this review that address this issue have a common underlying rational: mimic the biochemical cues that drive heart development in vivo and simulate the cardiac microenvironment. Certain features of the adult cardiomyocyte phenotype have, however, never been reproduced in culture, such as the presence of T-tubules, key structures of the ECC mechanism in mature cardiomyocytes that are important for normal Ca2+ handling. Furthermore, although the organization of sarcomeric structure in single-hPSC-CMs may increase over time in culture, it is still considered poor compared with an adult cardiomyocyte. M-bands have been noted by Kamakura et al. but these were not ubiquitous (only in 10% of cells) [13] and they only appeared after very long-term culture (360 days). Long differentiation and maturation protocols are unlikely to be practical. At the genomic level, expression of key sarcomeric genes may be ectopically upregulated, but levels are still considerably lower than in the adult heart. Heterogeneous maturation, reflected by different degrees of sarcomeric organization and variability of electrophysiological parameters, is another issue that has not been solved in studies to date.
Despite the limitations, studies on hPSC-CM maturation have revealed that there are conditions under which hPSC-CMs lower their spontaneous beating rate, hyperpolarize the RMP, and increase the force of contraction. hPSC-CMs that no longer spontaneously contract and have adopted prominent ventricular- or atrial-like features may have a more mature electrophysiological state. However, the absence of triggered electrical activity in these cells in culture may lead to the downregulation of sarcomeric proteins and proteins related to the contractile machinery, which could be counterproductive for maturation in the long run [13]. In addition to the absence of continuous electrical stimulation, other factors might contribute to cell immaturity. These include not only the absence of haemodynamic workload (mechanical strain) but also the lack of adjacent non-cardiomyocytes that could act via paracrine and humoral signals in vivo. Since each of the features of maturity may be independently regulated, strategies focusing on one particular aspect of the structural, electrophysiological, or functional phenotype alone may be insufficient to improve overall cardiomyocyte maturity. Combined approaches that impact multiple parameters at different levels simultaneously could be more effective in achieving this goal (Fig. 2). For example, in the study of Nunes et al. [18], a 3D culture system with adaptable mechanical properties was used in combination with electrical stimulation and in the presence of non-cardiomyocyte cells. The resulting cardiomyocytes had considerably improved structural, functional, and electrophysiological properties.
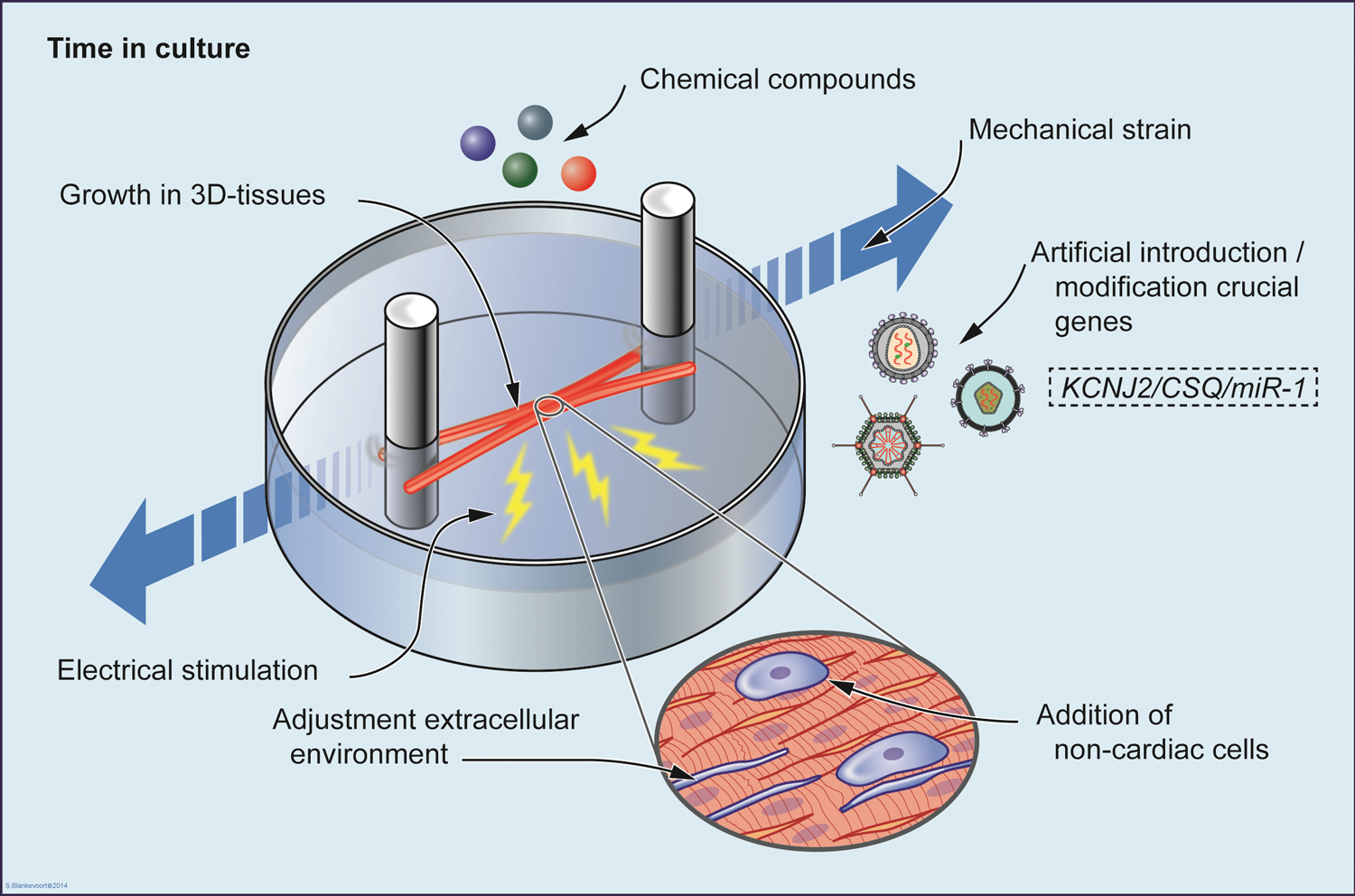
Illustrative representation of all suggested strategies applied to favor maturation of cardiomyocytes from human pluripotent stem cells, that is: prolonged time in culture, electrical stimulation, addition of chemical compounds, provision of mechanical strain, addition of non-cardiac cells, growth in a three-dimensional structure, adjustment of extracellular environment, and artificial introduction or modification of key genes involved with cardiac maturation. Color images available online at
Another important observation is that, although the expression of key markers of maturation (such as sarcomeric proteins or ion channels) may be considerably upregulated using a particular method, this does not always coincide with functional maturation. Gene expression may be used as an additional readout, but, ultimately, it is the functional/electrophysiological phenotype of hPSC-CMs that defines a successful maturation method.
It should be noted that much of the discussion here does not take into account heterogeneity among the hPSC-CMs resulting in the differentiation process. While electrophysiology measurements are often on single cells with specific AP morphology, such as nodal-, atrial-, or ventricular-like, gene expression is often determined on whole populations that include a mixture of subtypes of cardiomyocytes. It could thus be difficult to draw conclusions from gene expression profiles of each approach toward cardiomyocyte maturation. For example, HCN4 expression may be downregulated [10] or upregulated [11] in long-term cultures of hPSC-CMs corresponding to a population shift toward pacemaker cells. The MHC transition (β-MHC/α-MHC) might also reflect changes in the composition of cardiac population rather than maturation per se (α-MHC is more abundant in atrial than in ventricular cells). The same applies for T-type calcium currents, which are expressed by more cardiomyocytes over time in culture [12], perhaps reflecting a population shift toward nodal-like cells within the culture. Therefore, discrepancies in the expression of particular genes or functional properties among studies need to be evaluated carefully since they may be attributed to differences in cardiomyocyte subpopulation ratios rather than the maturation state per se. Methods to distinguish different hPSC-CM subtypes and attribute maturation to these specifically would greatly benefit research [156,157].
In determining whether hPSC-CM maturation is complete, it is important that morphological, functional, and electrophysiological characteristics are all assessed. Cell metabolism, for instance, has been measured in hPSC-CMs [100,158,159] but is rarely included in maturation studies. There remain other unresolved issues: The stiffness of the substrate on which cardiomyocytes are cultured seems to be important but it is not yet clear whether rigid substrates (such as glass) promote maturity [28,146] compared with softer substrates or vice versa; the best time for electrical stimulation, frequency, pulse duration, and the like also need optimization; and much remains to be learnt about the effects of chemical compounds, miRNAs, and growth hormones (including IGF-1 [160]). Ultimately, determining the exact molecular cues for hPSC-CM maturation would greatly benefit the attempts to induce an adult cardiomyocyte phenotype. Differences in materials and methods however, including details as apparently trivial as the presence or absence of serum in culture medium, confound clear conclusions at present. Moreover, hPSC-CMs show some line-to-line variability with respect to electrophysiological/functional output [161]. This has been attributed to various factors such as the reprogramming conditions or genetic background; it not only limits their use in modeling disease but should also be taken into account when evaluating the functional outcome of maturation protocols. It is essential that any method claiming cardiomyocyte maturation be confirmed in at least three other hiPSC or hESC lines to ensure that it is not a line specific effect.
To date, no conditions have been described under which hPSC-CMs become functionally identical to adult cardiomyocytes. It is still unclear whether this is actually achievable since human cardiomyocytes continue to develop even postnatally, with proliferation being replaced by hypertrophic growth and t-tubule formation only beginning to take place [162]. However, the present literature already demonstrates that a fully mature state might not be strictly necessary for hPSC-CMs to serve as useful human heart (disease) models. Nevertheless, with the development of new maturation strategies along the lines we have described, it may be expected that hPSC-CMs will achieve neonatal states and some features, such as APs, will resemble adult cardiomyocytes. Until then, those using hPSC-CMs as a model need to bear their present shortcomings in mind.
Footnotes
Acknowledgments
This work was supported by grants from the Rembrandt Institute of Cardiovascular Science (to C.L.M., for G.K. and C.C.V.), the Netherlands Institute of Regenerative Medicine (to C.L.M., for S.C. and M.B.), European Research Council ERC-AdG-STEMCARDIOVASC (C.L.M.), EU Marie Curie FP7-people-2011-IEF program, and HPSCLQT 29999 (M.B.), and the authors would like to thank Bas Blankevoort for his help with the figures and Matthew J. Birket for his contribution to this article.
Author Disclosure Statement
No competing financial interests exist.