
Abstract
Oct4 pseudogenes and isoforms seriously confuse the detection of the pluripotency-associated Oct4A expression in somatic cells, which in many cases was not accurately determined. This confusion has recently been discussed, but the wrong conclusions have continuously been made. Most studies failed to detect the expression of Oct4 pseudogenes and isoforms in somatic cells but detected only Oct4A, for which the detection signals incorrectly came from its pseudogenes and isoforms. Some studies detected the expression of only Oct4 pseudogenes in somatic cells but failed to detect Oct4A. The other studies failed to detect the expression of any Oct4 genes. Oct4A is more homologous to its pseudogenes than its isoforms, and it is much more difficult to distinguish Oct4A from its pseudogenes, so this study focused on them. In this study, the strict experimental procedures were followed. Three pairs of Oct4A-specific polymerase chain reaction (PCR) primers were carefully designed and tested by sequencing reverse transcription-polymerase chain reaction (RT-PCR) clones, which showed that only one of them was truly specific to Oct4A. RT-PCR was also performed with the primers amplifying both Oct4A and its pseudogenes, and several hundreds of PCR clones from each cell type were sequenced to reliably distinguish the low-abundant Oct4A from its high-abundant pseudogenes. Western blot, immunocytochemistry, and flow cytometric analyses were performed with three Oct4 antibodies to confirm the results of Oct4 mRNA expression. This study undoubtedly made the correct conclusions about Oct4 expression in human somatic cells and showed that all the tested human somatic cells expressed both Oct4A and its pseudogenes but expressed Oct4A at much lower levels compared with its pseudogenes.
Introduction
H
There are six Oct4 pseudogenes (abbreviated as Oct4 pg1–6) (Supplementary Fig. S1B and Supplementary Table S1) [9 –11]. Oct4 pg1, pg3, and pg4 can encode proteins that are highly homologous to Oct4A, and Oct4 pg2, pg5, and pg6 cannot synthesize mRNAs and proteins [9,11]. Some commercially available Oct4 antibodies were described to specifically detect Oct4A, but they can actually bind to the N-termini of Oct4A and Oct4 pg1, pg3, and pg4 proteins. So far, there is no Oct4A-specific antibody commercially available.
It is clearly shown that Oct4A gene is expressed at high levels in ESCs, induced pluripotent stem cells (iPSCs), embryonic carcinoma cells, and germ cell tumors [12,13]. Up to now, the expression of Oct4A in somatic cells including mesenchymal stem cells (MSCs) [14,15], tumor cells, and differentiated cells is still questionable. The wrong conclusions have continuously been made about the expression of Oct4A in somatic cells and include the following three kinds of confusions. (i) Most studies failed to show the expression of Oct4 pseudogenes and isoforms in somatic cells but detected only Oct4A, for which the detection signals incorrectly came from its pseudogenes and isoforms, and the used polymerase chain reaction (PCR) primers and Oct4 antibodies could not actually distinguish Oct4A from its pseudogenes and isoforms (Supplementary Tables S2 and S3) [16 –39]. (ii) Some studies detected only Oct4 pseudogenes in somatic cells but failed to detect Oct4A due to the low sensitivity of the used detection methods (Supplementary Table S4) [11,40 –43]. (iii) The other studies failed to detect the expression of any Oct4 genes including Oct4A and its pseudogenes and isoforms in human somatic cells (Supplementary Tables S4 and S5) [44 –50]. Though these confusions have recently been realized, these wrong conclusions have continuously been made. Because Oct4A is a key transcription factor to regulate self-renewal and pluripotency in ESCs [2 –4] and also has important functions in somatic cells as described in the Discussion section below, and these wrong conclusions have seriously prevented the advancement of this important research area, it is urgent to replace the previous protocols, make the correct conclusions, and facilitate the advancement of this important research. In this study, the strict procedures were followed to undoubtedly detect the expression of Oct4A and its pseudogenes in human somatic cells.
Materials and Methods
Isolation of UC-MSCs and BM-MSCs
Human umbilical cords (UCs) were aseptically collected from full-term caesarean section patients, and human bone marrow (BM) aspirates were obtained from the iliac crest of donors (5–36 years old). These samples were obtained at the First Affiliated Hospital of Jinan University, Guangzhou, China after informed consent and approval from the local Ethical Review Board at Jinan University. The isolation and culture of UC-MSCs and BM-MSCs were performed as previously described [51].
Cell culture
The following cells were obtained from American Type Culture Collection (ATCC, Rockefeller, MD). MCF-7 and 293T were grown in high glucose Dulbecco's modified Eagle's medium (Invitrogen, Carlsbad, CA) containing 10% fetal bovine serum (FBS). DU145, PC3, and MRC-5 were grown in 1640 medium (Invitrogen) containing 10% FBS.
Adipogenic and osteogenic differentiation of UC-MSCs
Adipogenic and osteogenic differentiation of UC-MSCs were performed as previously described [51]. After adipogenic induction for 21 days, the generation of neutral lipid droplets was detected by staining with oil red O (Sigma, St Louis, MO). After osteogenic induction for 21 days, osteogenic differentiation was examined by alkaline phosphatase and von Kossa staining (Sigma).
Real-time PCR
Total RNA was extracted from cells using Total RNA Kit I (Omega, Norcross, GA) and treated with DNase I (Promega, Madison, WI). One microgram of total RNA was reverse-transcribed using Reverse Transcription System (Promega). Real-Time PCR was performed using SYBR Premix Ex Taq™ (Takara Bio Ltd., Shiga, Japan) and the three pairs of Oct4 primers, including Oct4 intron 1a [49], Oct4 intron 1b, and Oct4A (Table 1). PCR was performed by initial denaturation at 95°C for 30 s, followed by 40 cycles of denaturation at 95°C for 5 s, annealing at 60°C (Oct4A primers) or 65°C (Oct4 intron 1a and 1b primers) for 20 s and extension at 72°C for 20 s. PCR amplicons were further examined by agarose gel electrophoresis.
PCR, polymerase chain reaction.
RT-PCR and sequencing of PCR amplicons
RT-PCR was performed with the four pairs of Oct4 primers, including Oct4 intron 1a, Oct4 intron 1b, Oct4A, and Oct4u (Table 1). PCR was performed by initial denaturation at 95°C for 30 s, followed by 40 cycles of denaturation at 95°C for 5 s, annealing at 60°C (Oct4A primers) or 65°C (Oct4 intron 1a, Oct4 intron 1b, and Oct4u primers) for 20 s, and extension at 72°C for 30 s. PCR amplicons were cloned in pGEM-T Easy Vector System (Promega). Clones were verified by PCR and sequenced. DNA sequences were analyzed by BLAST software for homology against public databases.
Western blot analysis
Cells were lysed in cell lysis buffer (Beyotime, Haimen, Jiangsu, China). Forty micrograms of cell lysate was separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), and proteins were electro-transferred to polyvinylidene fluoride (PVDF) membrane (Pall Corp., Port Washington, NY). The membranes were blocked for 1 h with 5% nonfat milk in Tris-buffered saline containing Tween-20 (TBST) and hybridized with anti-Oct4 antibodies including sc-5279 (Santa Cruz Biotechnology, Santa Cruz, CA), ab109187 (Abcam, Cambridge, MA), and 2840S (Cell Signaling Technology, Danvers, MA), and anti-β-actin antibody (Beyotime) was used as a housekeeping gene control. Membranes were incubated for 1 h with horseradish peroxidase-conjugated secondary antibody (Cell Signaling Technology), and signal was detected by an ECL detection kit (Thermo Fisher Scientific, Waltham, MA).
Immunocytochemistry
Cells were fixed for 15 min in 4% paraformaldehyde and blocked for 30 min with phosphate-buffered saline (PBS) containing 5% goat serum (Invitrogen) and 0.1% Triton X-100 (Sigma). Cells were incubated at 4°C overnight with anti-Oct4 antibodies sc-5279 (1:50), ab109187 (1:100), or 2840S (1:100) containing 5% goat serum. Normal mouse IgG2 (1:100) and rabbit IgG (1:100) (Santa Cruz Biotechnology) were used as antibody isotype controls respectively for sc-5279, ab109187, or 2840s. Cells were washed five times in PBST and incubated for 1 h at room temperature with phycoerythrin (PE)-labeled secondary antibodies including goat anti-mouse IgG and goat anti-rabbit IgG (Santa Cruz Biotechnology). Cells were counterstained with DAPI for 5 min and washed five times with PBST. Cells were examined under a fluorescence microscope (IX71; Olympus, Tokyo, Japan).
Flow cytometric analysis
Cells were fixed for 30 min with 4% paraformaldehyde at room temperature. They were permeabilized and blocked for 30 min at room temperature with PBS containing 0.1% Triton-X and 10% goat serum. Cells were incubated for 1 h with anti-Oct4 antibodies (sc-5279, ab109187 or 2840s) and then, for 1 h with PE-conjugated goat anti-mouse or goat anti-rabbit antibody. Normal mouse or rabbit IgG was used as an antibody isotype control. Cells were analyzed with a FACSCalibur flow cytometer (Becton Dickinson, Franklin Lakes, NJ).
Statistical analysis
Data were expressed as mean±standard error of the mean. Statistical comparisons were performed using the Student's t-test. P-values <0.05 were considered statistically significant.
Results
All the tested human somatic cells express Oct4A transcript but at much lower levels compared with human ESCs
To specifically detect Oct4A transcript, three pairs of PCR primers were designed and tested by RT-PCR with total RNA isolated from UC-MSCs (P6) and sequencing the clones of the PCR amplicons. Although these three pairs of PCR primers have 100% identity only to Oct4A mRNA as analyzed by NCBI BLAST software against the NCBI databases, the analysis of cDNA sequences cloned from PCR amplicons showed that only Oct4A primers specifically detected Oct4A transcript, and the other two pairs of primers detected both Oct4A and its pseudogene transcripts (Tables 1 and 2). Then, Oct4A primers were used to examine the Oct4A mRNA expression in human somatic cells by RT-PCR, and the PCR amplicons were cloned and sequenced. BM-MSCs, UC-MSCs, cancer cell lines (MCF-7, DU145, PC3, and HeLa), human embryonic kidney cells (293T), and fetal fibroblasts (MRC-5) were examined. Human iPSCs (hiPSCs) and human ESCs (hESCs) were included as positive controls. The results further confirmed that Oct4A primers detected only Oct4A mRNA, and all the tested human somatic cells expressed Oct4A mRNA (Table 3).
RT-PCR was performed with total RNA isolated from UC-MSCs (P6). PCR products were cloned and sequenced, and cDNA sequences were examined for homology against NCBI databases by NCBI BLAST. Oct4 intron 1a and intron 1b primers can amplify Oct4A, and its pseudogenes, and Oct4A primers can amplify Oct4A only.
RT-PCR, reverse transcription-polymerase chain reaction; UC-MSCs, umbilical cord mesenchymal stem cells.
RT-PCR was performed with total RNA isolated from the human somatic cells. PCR products were cloned and sequenced, and cDNA sequences were examined for homology against NCBI databases by NCBI BLAST. Oct4u primers can amplify Oct4A, and Oct4 pg1, pg3, and pg4 only, and Oct4A primers can amplify Oct4A only.
BM, bone marrow; hESC, human embryonic stem cell; hiPSC, human induced pluripotent stem cell; P3, passage 3; P6, passage 6; P9, passage 9.
The three pairs of PCR primers were also used for real-time PCR and detected similar levels of Oct4 mRNAs in all the tested cells except for MRC-5 and HeLa (Fig. 1A). hiPSCs and hESCs were included as positive controls. Real-time PCR with Oct4A primers showed that the expression of Oct4A mRNA was 1,160- and 800-fold higher in hESCs compared with BM-MSCs and UC-MSCs (P3), respectively. The expression of Oct4A mRNA decreased at the later passages of UC-MSCs (P6 and P9) and after the adipogenic and osteogenic differentiation. Cancer cell lines (MCF-7, DU145, and PC3) also expressed Oct4A mRNA at very low levels. MRC-5 and HeLa had undetectable Oct4A mRNA. These results were also confirmed by agarose gel electrophoresis of PCR amplicons (Fig. 1B).
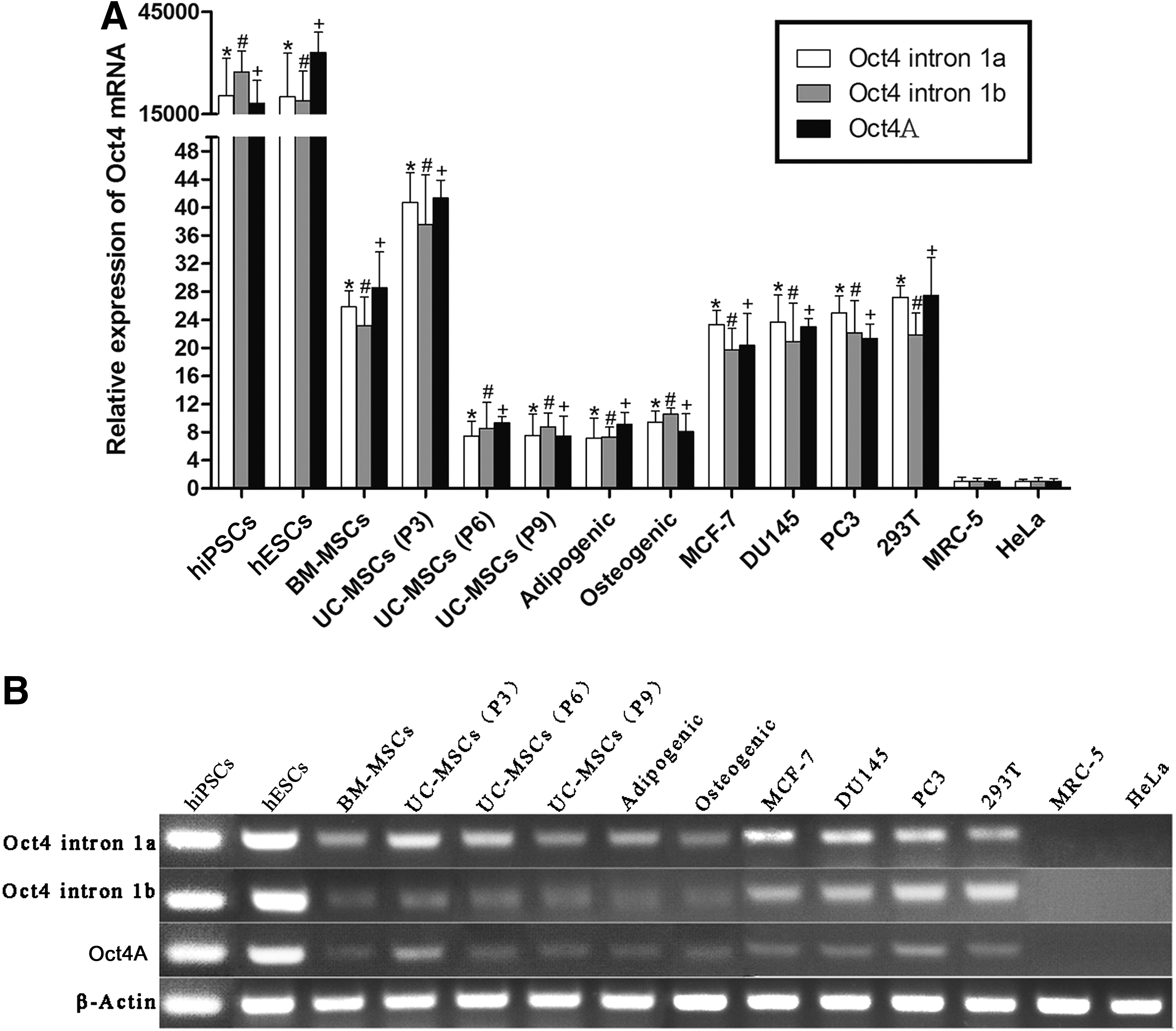
Human somatic cells express Oct4A transcript at much lower levels compared with human embryonic stem cells (hESCs).
Human somatic cells express Oct4A transcript at much lower levels compared with Oct4 pseudogene transcripts
Oct4A and Oct4 pg1, pg3, and pg4 are highly homologous in 5′ termini of their transcripts, which are absent or different from all the other Oct4 pseudogenes and isoforms (Supplementary Fig. S1A, B). To distinguish the expression of Oct4A from Oct4 pg1, pg3, and pg4, Oct4u primers were designed for RT-PCR to amplify only Oct4A and Oct4 pg1, pg3, and pg4 transcripts, and PCR amplicons were cloned and sequenced (Table 3). hiPSCs and hESCs expressed only Oct4A mRNA. In contrast, all the tested somatic cells expressed both Oct4A and its pseudogene mRNAs but expressed Oct4 pseudogenes at much higher levels compared with Oct4A. For example, the expression of Oct4 pg3 mRNA was 121- and 84-fold higher in BM-MSCs and UC-MSCs (P3) respectively compared with Oct4A. BM-MSCs, UC-MSCs of the three different passages, adipogenic cells differentiated from UC-MSCs, PC3, and MRC-5 cells predominantly expressed Oct4 pg3 mRNA. MCF-7 and osteogenic cells differentiated from UC-MSCs predominantly expressed Oct4 pg1 mRNA. DU145, 293T, and HeLa cells predominantly expressed Oct4 pg3 and pg4 mRNAs.
Human somatic cells express Oct4A and its pseudogene proteins
Some commercially available Oct4 antibodies were described to specifically detect Oct4A, but they can actually bind to the N-termini of Oct4A and Oct4 pg1, pg3, and pg4 proteins. So far, there is no Oct4A-specific antibody commercially available. The expression of Oct4 proteins was investigated with the three different Oct4 antibodies by western blot analysis to confirm the results of Oct4 mRNA expression. These three Oct4 antibodies bind to Oct4 N-termini and detect Oct4A in addition to Oct4 pg1, pg3, and pg4, but not the other Oct4 pseudogenes and isoforms. The 45 kDa band was detected in all the tested cells and represents only Oct4A and Oct4 gp1 according to the predicted protein sizes, and all the three antibodies gave similar results (Fig. 2A). hiPSCs and NTERA-2 (a human embryonic carcinoma cell line derived from a teratocarcinoma) were included as positive controls. All the tested somatic cells expressed Oct4 proteins at much lower levels compared with hiPSCs and NTERA-2. Oct4 protein expression decreased in UC-MSCs at the later passages (P6 and P9) and after the adipogenic and osteogenic differentiation. Four tumor cells (MCF-7, DU145, PC3, and HeLa) also expressed Oct4 proteins.

Human somatic cells express Oct4A and its pseudogene proteins.
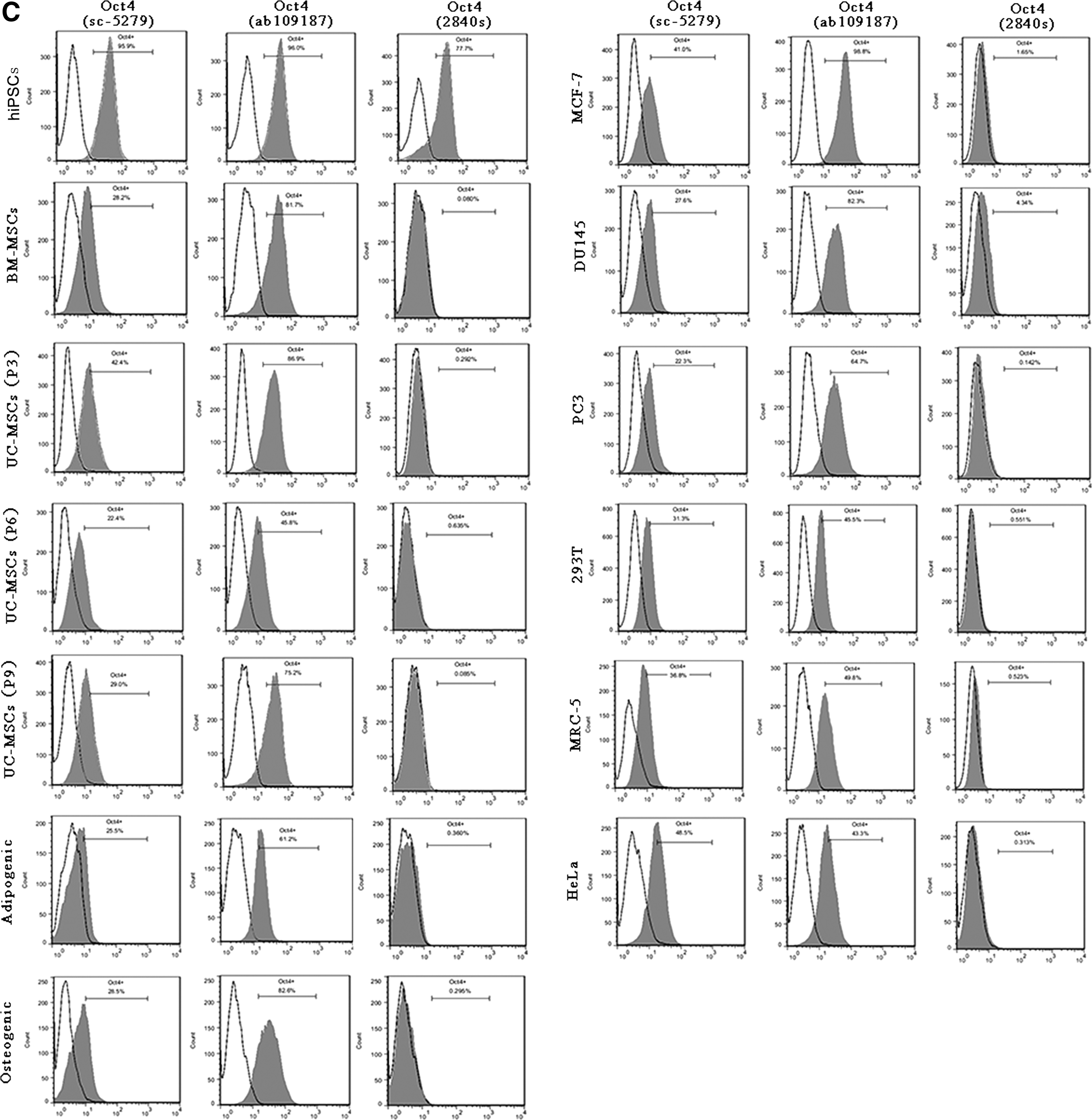
The Oct4 protein expression was also examined with the same three Oct4 antibodies by immunocytochemistry (Fig. 2B and Table 4). The antibody ab109187 gave stronger signal, and the antibody 2840s gave weaker signal. hiPSCs were included as a positive control. Somatic cells expressed Oct4 proteins at much lower levels compared with hiPSCs. Oct4 protein expression decreased in UC-MSCs at passage 9 (P9) and after the adipogenic and osteogenic differentiation. Four tumor cells (MCF-7, DU145, PC3, and HeLa) also expressed Oct4 proteins.
+, low level of Oct4 expression; ++++, high level of Oct4 expression; —, no Oct4 expression.
Oct4 protein expression was also tested with the same three Oct4 antibodies by flow cytometry (Fig. 2C). The antibodies sc-5279 and ab109187 detected Oct4 proteins in all the tested cells while the antibody 2840s could not detect Oct4 proteins in all the tested somatic cells except for hiPSCs as a positive control. Since the three Oct4 antibodies cannot distinguish Oct4A from Oct4 pg1, pg3, and pg4, the positive signal detected by immunocytochemistry and flow cytometry can come from Oct4A or Oct4 pg1, pg3, or pg4 proteins.
Discussion
Up to now, Oct4A expression in somatic cells is still questionable. Supplementary Table S4 lists some references that failed to detect Oct4A expression or detected only its pseudogenes in human somatic cells and discusses the possible reasons why Oct4A expression was not detected. It is due to the low sensitivity of the used methods, the lack of specificity of the used PCR primers and antibodies or only small number of PCR amplicon clones being sequenced.
PCR primers for the detection of Oct4A expression should be carefully designed to be identical to only Oct4A mRNA. The difference between Oct4A and Oct4 pg1, pg3, and pg4 mainly lies in the first 40 nucleotides of Oct4A mRNA, which are absent in the other Oct4 isoforms and pseudogenes so that the forward primer should be designed in this region to specifically detect Oct4A mRNA. However, Oct4 primers used in many studies are actually homologous to Oct4A and its isoforms and pseudogenes (Supplementary Table S3). After Oct4A-specific primers are designed, they must be further tested by real-time PCR or RT-PCR. In this study, three pairs of primers (Oct4A and Oct4 intron 1a and 1b) were designed to be identical to only Oct4A mRNA and tested by RT-PCR (Fig. 3). Oct4 intron 1a forward primer has a single nucleotide at 3′ end different from Oct4 pg1 and still detected Oct4 pg1 transcript in UC-MSCs (Fig. 3 and Table 2). Likewise, each of Oct4 intron 1b forward and reverse primers has a single nucleotide at 3′ end different from Oct4 pg3 and still detected Oct4 pg3 transcript in UC-MSCs (Fig. 3 and Table 2). These results demonstrate that a primer containing only a single nucleotide at its 3′ end, which is different in two transcripts, is not enough to distinguish them. If Taq polymerase has 3′ to 5′ exonuclease activity, it can excise the mismatched nucleotide at a primer's 3′ end and make extension using the template. If Taq polymerase lacks 3′ to 5′ exonuclease activity, it can make an extension by low reliability from the mismatched nucleotide at a primer's 3′ end. In addition, Oct4 intron 1a forward primer has a single nucleotide inside the primer different from Oct4 pg4 and still detected Oct4 pg4 transcript (Fig. 3 and Table 2). Oct4 intron 1a reverse primer has a single nucleotide inside the primer different from Oct4 pg3 and still detected Oct4 pg3 transcript (Fig. 3 and Table 2). Oct4A forward primer nucleotides are much different from Oct4 pg1 and absent in Oct4 pg3 and pg4 and specifically detected only Oct4A transcript (Fig. 3 and Table 2).

Nucleotide sequence alignment of three pairs of Oct4 primers against Oct4, and Oct4 pg1, pg3, and pg4 cDNAs.
PCR amplicons should be cloned, and several hundreds of clones must be sequenced to assure of the detection of low-abundant Oct4A transcript and confirm the specificity of the primers (Table 3). In addition, total RNA used for real-time PCR or RT-PCR must be treated with DNase I to remove the genomic DNA contamination even if the primers are designed to span Oct4 introns. This is especially important since Oct4 pseudogenes do not contain introns and cannot be distinguished from Oct4A by the sizes of PCR amplicons. This study showed that all the three pairs of PCR primers detected the similar levels of Oct4 expression in the different somatic cells by real-time PCR (Fig. 1A, B). Though Oct4 intron 1a and 1b primers can amplify Oct4 pseudogene mRNAs, the detection efficiency should be low due to the nucleotides different between the primers and Oct4 pseudogene mRNAs (Fig. 3), the relative amount of Oct4A or its pseudogene mRNA different in different cells and the competition for the amplification of Oct4A or its pseudogene mRNAs. Real-time PCR showed that Oct4A mRNA expression was about 1,000-fold higher in hESCs and hiPSCs than the other cells (Fig. 1A). However, this is not consistent with Oct4 protein expression as determined by flow cytometry, which showed almost identical expression levels in the tested cells (Fig. 2C). The reasons for this could be that mRNA expression level may not be the same as protein expression level, and flow cytometry is not a sensitively quantitative method and analyzes protein expression at individually cellular level. In addition, Oct4A primers specifically amplify Oct4A while Oct4 antibodies can detect both Oct4A and its pseudogenes. Larger difference of Oct4A protein levels between hiPSCs and the other cells was detected by western blot analysis and immunocytochemistry compared with flow cytometry (Fig. 2). In addition, flow cytometry showed that Oct4 antibody 2840s detected Oct4 protein exclusively in hiPSCs but not in the other cells (Fig. 2C). This result does not prove that this antibody is more selective for Oct4A because western blot analysis and immunocytochemistry showed that it also detected Oct4 expression in the other cells (Fig. 2A, B and Table 4). It is unknown why this antibody did not detect Oct4 in the other cells by flow cytometry, and this result suggests that it may not be suitable for flow cytometry.
It is also important to choose appropriate Oct4 antibodies for the detection of Oct4 protein. The difference between Oct4A protein and its isoforms mainly lies in the N-termini, but Oct4A protein is highly homologous to Oct4 pg1, pg3, and pg4 so that so far there is no Oct4A-specific antibody commercially available. The antibodies used in many studies cannot actually distinguish Oct4A from Oct4 pg1, pg3, and pg4 [16 –27,29 –39]. For example, one study examined the expression of Oct4A by using the sc-5279 antibody against its amino acids 1–134, and this antibody cannot distinguish Oct4A from Oct4 pg1, pg3, and pg4 due to very high homology of N-termini of these proteins [28]. Especially, Oct4A and Oct4 pg1 proteins are almost identical, and Oct4A protein has only one more amino acid compared with Oct4 pg1 so that it is very difficult to distinguish them [9]. Another article also showed that Oct4A and Oct4 pg1 proteins have almost identical amino acid sequences, which can lead to the false-positive detection of Oct4A expression [11]. Oct4 pg3 protein is much smaller than Oct4A, and Oct4 Pg4 protein is also smaller than Oct4A. So, Oct4A protein can be distinguished from Oct4 pg3 and pg4 by western blot according to their different sizes. In this study, three Oct4 antibodies, which can bind to N-termini of Oct4A and Oct4 pg1, pg3, and pg4 proteins, were used to detect and compare Oct4 protein expression by western blot, immunocytochemistry, and flow cytometry. Different detection methods should be chosen, and different Oct4 antibodies should be used to detect and compare Oct4 protein expression. There is an urgent need for the identification of an antibody that detects only Oct4A and no pseudogenes, binds to N-terminal of Oct4A protein, and recognizes the amino acid different between Oct4A and Oct4 pg1 protein. This new Oct4A-specific antibody should be helpful for stem cell researchers.
Based on this study, we strongly suggest to strictly follow the eight important approaches below to exclusively detect Oct4A expression and distinguish the expression of Oct4A from its isoforms and pseudogenes in somatic cells. (i) PCR primers should be carefully designed and checked by BLAST software to be identical only to Oct4A. Since the main difference between Oct4A and its isoforms and pseudogene mRNAs lies in exon 1, the forward primer should be in exon 1; (ii) PCR primers must be tested by RT-PCR, and PCR amplicons must be cloned and sequenced to confirm that the primers are truly specific for Oct4A; (iii) total RNA must be treated with DNase I to remove the genomic DNA contamination. This is especially important since Oct4 pseudogenes do not contain introns. (iv) Since Oct4A is highly homologous to Oct4 pg1, pg3, and pg4, RT-PCR should also be performed with primers that can amplify transcripts of Oct4A and Oct4 pg1, pg3, and pg4. PCR amplicons should be cloned, and several hundreds of clones from each cell type must be sequenced to distinguish the low-abundant Oct4A mRNA from the high-abundant Oct4 pseudogenes; (v) so far, there is no Oct4A-specific antibody commercially available. Since the N-terminal of Oct4A protein is highly homologous to only Oct4 pg1, pg3, and pg4, Oct4 antibodies should be selected to specifically bind to the N-termini of Oct4 proteins, and at least three Oct4 antibodies should be used to compare positive signals; (vi) western blot should be performed to distinguish Oct4A and Oct4 pg1 proteins from the other Oct4 isoforms and pseudogenes because Oct4A protein has only one additional amino acid compared with Oct4 pg1; (vii) immunocytochemistry and flow cytometric analysis should also be performed to confirm Oct4 protein expression; (viii) appropriate controls are crucial for the exclusion of false positives, including positive control cells such as hESCs, no reverse transcriptase control and antibody isotype control.
It is well known that Oct4A is a key transcription factor to maintain the self-renewal and pluripotency in ESCs. Though somatic cells are not pluripotent, many articles also showed that Oct4A has functions in different somatic cells, and some articles were described below as examples. One study showed that Oct4A promotes Dnmt1 expression in human BM-MSCs, which represses the expression of p16 and p21 and genes associated with development and lineage differentiation, and the knockdown of Oct4A and Nanog expression in human BM-MSCs decreases proliferation and differentiation potential and enhances spontaneous differentiation [24]. Another study reported that Oct4A can upregulate the expression of CD49f, which increases the proliferation and multipotency of human umbilical cord blood MSCs through the prolonged activation of phosphatidylinositol 3-kinase (PI3K)/AKT and suppressed expression of p53 [26]. It was shown that the forced expression of Oct4A can activate the expression of other stemness genes, such as Sox2, Nanog, c-Myc, FoxD3, KLF4, and β-Catenin in human BM-MSCs and fibroblasts, leading to the activation of the pluripotent network [52]. It was found that the suppression of Oct4A expression in human umbilical cord blood-derived MSCs leads to downregulation of the expression of CCNA2, SOX2, REX1, and c-MYC [3]. These results and the other studies [53 –57] showed that Oct4A can maintain the self-renewal and multipotency in MSCs. Oct4A also has functions in cancer cells. One article showed that the coexpression of Oct4A and Nanog enhances malignancy in human lung adenocarcinoma by inducing cancer stem cell-like properties and epithelial–mesenchymal transition (EMT), and double knockdown of Oct4A and Nanog reverses the EMT process and blocks the tumorigenic and metastatic ability [58]. It was reported that Oct4A can promote tumorigenesis and inhibit apoptosis of cervical cancer cells by directly inducing expression of miR-125b, which inhibits its direct target BAK1 [59]. Another study showed that Oct4A can upregulate BIRC5 and CCND1 expression, which promotes the proliferation of hepatocellular carcinoma cells [60]. It has been shown that Oct4A overexpression can upregulate the expression of IL-8 and IL-32, which can regulate the cancer stem cell-like properties that promote tumorigenesis of colorectal cancer cells [61]. It was found that the reduction in Oct4A expression in human breast cancer cells MCF7 induces cancer stem cell-like cell apoptosis through the Oct4A/Tcl1/Akt1 pathway [62]. These results and the other articles [63 –71] demonstrated that Oct4A can promote tumorigenesis of different cancers. In addition to its functions in MSCs and cancers, Oct4A also has functions in other somatic cells. For example, overexpression of Oct4A protein in primary human endometriotic stromal cells and human RL95-2 and HEC1A endometrial carcinoma cell lines results in the decreased expression of E-cadherin; the increased expression of the Vimentin, Twist, and Slug proteins; and an increase in the migration activity of endometrial cells [72]. After human skin keratinocytes are transiently transfected with Oct4A, these cells display the expression of endogenous embryonic genes and show reduced genomic methylation. More importantly, these cells can be specifically converted into neuronal and contractile mesenchymal cell types [73]. Oct4A downregulation accelerates neuronal differentiation of progenitor cells while its sustained expression prevents neuronal differentiation, and transplantation of neurospheres into the adult rat brain shows that downregulation of Oct4A promotes differentiation of neurosphere cells in vivo [74]. Oct4A in cooperation with Nanog strongly suppresses the myogenic differentiation program and promotes pluripotency in myoblasts [75].
Footnotes
Acknowledgments
This work was supported by National Natural Science Foundation of China (grant no. 31171304 and grant no. 30870650 to X.W.), Guangdong Science and Technology [no. 20130901 (no. 61) to X.W.], Guangzhou Science and Technology (no. 1563000464 to X.W.), and Research Foundation for Doctoral Discipline of Higher Education (grant no. 20114401110007 to X.W.).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.