
Abstract
Adipose-derived stem cells (ADSCs) have been used to control several autoimmune or inflammatory diseases due to immunosuppressive properties, but their role in obesity-associated inflammation remains unestablished. This study aims to evaluate the effects of ADSCs on obesity-induced white adipose tissue (WAT) inflammation and insulin resistance. We found that diet-induced obesity caused a remarkable reduction of ADSC fraction in mouse WAT. Delivery of lean mouse-derived ADSCs, which could successfully locate into WAT of obese mice, substantially improved insulin action and metabolic homeostasis of obese mice. ADSC treatment not only reduced adipocyte hypertrophy but also attenuated WAT inflammation by reducing crown-like structures of macrophages and tumor necrosis factor (TNF)-α secretion. Importantly, ADSC treatment remodeled the phenotypes of adipose-resident macrophages from proinflammatory M1 toward anti-inflammatory M2-like subtypes, as characterized by decreased MHC class II-expressing but increased interleukin (IL)-10-producing macrophages together with low expression of TNF-α and IL-12. Coculture of ADSCs through the transwell or conditional medium with induced M1 macrophages also reproduced the phenotypic switch toward M2-like macrophages, which was substantiated by elevated arginase 1, declined inducible nitric oxide synthase, inhibition of NF-κB activity, and activation of STAT3/STAT6. Taken together, our data support that ADSC supplement in obese mice could sustain IL-10-producing M2-like macrophages in WAT through paracrine action, thereby suggesting the crucial role of ADSCs in resolving WAT inflammation, maintaining adipose homeostasis, and proposing a potential ADSC-based approach for the treatment of obesity-related diseases.
Introduction
O
Macrophages, recently referred to as classically activated M1 and alternatively activated M2 subtypes, change their polarization status and function differently to facilitate immune function and maintain tissue homeostasis in various microenvironments [9 –12]. A report from Lumeng et al. has demonstrated that obesity induces a phenotypic switch of adipose tissue macrophages from an anti-inflammatory M2 polarization state producing interleukin (IL)-10 and arginase 1 to a proinflammatory M1 polarization state with a high expression of TNF-α and inducible nitric oxide synthase (iNOS), thus losing the protection from TNF-α-induced insulin resistance [13]. This finding was substantiated by growing evidences, thereby providing the possibility to resolve WAT inflammation and insulin resistance by reversing the phenotypic switch of macrophages in the obese state [9,10,12,14 –16].
Adipose-derived stem cells (ADSCs), also known as adipose-derived stromal cells, mainly reside in the stromal vascular cell fraction (SVF) of adipose tissues. Similar to bone marrow (BM)-derived mesenchymal stem cells (MSCs), ADSCs express CD90, CD105, CD44, CD73, and display multipotent stem potency [17,18]. Accumulating data have elucidated the beneficial effects of human ADSCs on tissue engineering due to their immunoregulatory and regenerative potency [19,20]. More recently, ADSCs have been used in several experimental models of autoimmune or inflammatory diseases such as arthritis and colitis [21 –23]. A study has also reported that allogeneic ADSC therapy efficiently ameliorates autoimmune diabetes pathogenesis in NOD mice by attenuating a Th1 immune response, indicating the immunoregulatory role of ADSCs in metabolism-related inflammatory diseases [24]. However, the possible role of ADSCs in obesity-associated inflammation remains largely unknown. There is emerging evidence that BM-MSCs suppress inflammation by modulating macrophage phenotypes [25 –27], thereby promoting us to explore the regulatory role of ADSCs in obesity-induced WAT inflammation and phenotypes of adipose-resident macrophages.
In the current study, we confirmed the shortage of ADSC fraction in high-fat diet (HFD)-fed obese mice and further evaluated the effects of ADSC treatment in these mice. We demonstrate that ADSC delivery efficiently attenuates WAT inflammation and insulin resistance accompanied by improved metabolic homeostasis in these mice. ADSCs reprogram the macrophage phenotypes in WAT from inflammatory M1 toward anti-inflammatory M2-like macrophages, which could be a primary contributor to the above desirable effects, thereby proposing ADSC supplement as a potential therapeutic approach for obesity-associated inflammation and insulin resistance.
Materials and Methods
Animals and diets
C57BL/6 male mice were purchased from Vital River Laboratory Animal Technology Co., Ltd. (Beijing, China). All animal feed were obtained from Trophic Animal Feed High-tech Co., Ltd. (Nantong, China). After 1 week of diet adaptation, mice were induced obesity by feeding with HFD (60% of total calories) at the age of 8 weeks. Mice fed on a normal diet (ND) (10% of total calories) were used as lean controls. Body weight was recorded every week. All animal studies were conducted in adherence to the Animal Care and Utilization Committee of Shandong University and all experimental procedures were in accordance with institutional guidelines.
Isolation and expansion of ADSCs
ADSCs used in the experiments were derived from ND-fed mice at the age of 8–10 weeks. SVF was isolated, and ADSCs were expanded as previously described [28]. Briefly, minced epididymal fat pads were digested in the Krebs-Ringer Bicarbonate Buffer containing 2 mg/mL collagenase Type 1 (Worthington, Lakewood, NJ). After a filtration through a 100-μm mesh, SVF was incubated overnight in Dulbecco's Modified Eagle's Medium (DMEM) supplemented with 10% fetal bovine serum (FBS) and 100 U/mL penicillin/streptomycin at 37°C, 5% CO2. After removal of nonadherent cells, the medium was changed into a fresh one containing 5 ng/mL basic fibroblast growth factor (FGF; Peprotech, Rocky Hill, NJ). The third to fifth passages of ADSCs were used for the experiments.
Tracking of ADSCs in HFD-fed mice
ADSCs from epididymal fat pads were labeled with 10 μM 5-Ethynyl-2′-deoxyuridine (EdU) in DMEM at 37°C, 5% CO2 for 24 h. The labeled cells were delivered into HFD-fed mice by intraperitoneal injection (1×106/mice). After 3 or 7 days, the epididymal fat pads were harvested for EdU detection using the Cell-Light™ EdU Apollo®567 Cell Tracking Kit (RiboBio, Guangzhou, China) according to the manufacturer's instruction.
ADSC treatment in HFD-fed mice
After 8 weeks of HFD feeding, mice were administrated with ADSCs by intraperitoneal injection, 1×106 ADSCs in 0.5 mL normal saline (NS) were delivered per mice once a week. Mice fed on HFD or ND in control groups were treated with NS. After 6 weeks of ADSC delivery, mice were sacrificed under anesthesia. Serum and epididymal fat pads were collected. More than three independent experiments were performed, and four to six mice per group were included in each experiment.
Metabolic parameters measurement
Serum triglyceride (TG) and total cholesterol (TC) (n=7–12 mice/group) were measured with Triglyceride or Cholesterol Test Kits (BHKT clinical reagent, Beijing, China), respectively. Nonfasting plasma levels of glucose (n≥4 mice/group) were determined using OneTouch Ultra in vitro diagnosticum (LifeScan, Milpitas, CA). The glucose tolerance test (GTT) (n=4–5 mice/group) was performed in mice following an overnight fast, and blood glucose levels were determined before and 15, 30, 45, 60, 90, and 120 min after an intraperitoneal injection of
Histology and immunofluorescence
Mice adipose tissues were fixed in 4% paraformaldehyde and prepared into paraffin-embedded sections. Hematoxylin and eosin staining was performed according to a standard protocol. For adipose tissue sections, the average sizes of adipocytes (n=3–5 mice/group) were measured using Image-Pro Plus 6.0. To detect the infiltration of macrophages, adipose sections were stained with the rat anti-mouse CD68 antibody (Ab) (Bioss, Beijing, China) followed by Alexa 594-conjugated secondary Ab (Proteintech, Chicago, IL). Nuclei were stained by DAPI. The fluorescence signals were detected using the fluorescence microscope (Axio Imager A2; Carl Zeiss, Jena, Germany) (n≥3 mice/condition).
Adipose tissue explant culture
Adipose tissues from the epididymal fat pads were chopped into small pieces. The explants were dispersed in DMEM supplemented with 10% FBS at 0.3 g/mL and incubated at 37°C, 5% CO2 for 24 h. The supernatants from cultured adipose explants were collected for the inflammation assay.
Cytometric bead array immunoassay
Serum from individual mice and supernatants from adipose explants were collected for the inflammatory marker assay using the Cytometric Bead Array Mouse Inflammation Kit (BD biosciences, San Jose, CA) according to the manufacturer's instructions (n=3–4 mice/group). Data were collected with the FACS Calibur Flow Cytometer and analyzed by the FCAP Array v 1.0 (BD biosciences).
Coculture of macrophages with ADSCs
ADSCs were cultured in DMEM supplemented with 10% FBS and 100 U/mL penicillin/streptomycin in a humidified atmosphere at 37°C, 5% CO2, and the conditional medium was collected after 3 days of incubation. Peritoneal macrophages were isolated as previously described [29]. Briefly, murine macrophages were obtained from peritoneal lavages elicited by a sterile starch solution and planted in six-well plates at 1.5×106 per well in DMEM supplemented with 10% FBS and 100 U/mL penicillin/streptomycin. After overnight incubation, macrophages were induced to M1 polarization by 100 ng/mL lipopolysaccharide (LPS; Sigma-Aldrich) plus 20 ng/mL interferon-gamma (IFN-γ; Peprotech) in the presence or absence of ADSC conditional medium for different time periods. Untreated macrophages were used as control. In some experiments, macrophages were cocultured with ADSCs through transwell plates for different time periods.
Flow cytometry
To characterize the surface markers of ADSCs, expanded ADSCs were stained with fluorescein isothiocyanate (FITC)-conjugated monoclonal antibodies (mAbs) against mouse CD90, CD44, CD11c, CD34, phycoerythrin (PE)-conjugated mAb against CD105, or PE-cy5-conjugated mAb against MHC class II (MHC-II) (eBioscience, San Diego, CA). For SVF or macrophages, cells were stained with FITC-conjugated anti-CD90, PE-conjugated anti-CD105 mAbs or PE-conjugated anti-CD11b, PE-cy5-conjugated anti-MHC-II mAbs, followed by intracellular staining with FITC-conjugated anti-IL-10 mAb. Corresponding isotypic Abs were served as controls. Cells were acquired using the FACS Calibur Flow Cytometer (BD Biosciences) or Cytomics FC500 (Beckman Coulter, Pasadena, CA).
Quantitative reverse transcription PCR
Total RNA was extracted using the TRIzol reagent (Invitrogen, Carlsbad, CA) and reversely transcripted into cDNA using a ReverTra Ace qPCR RT Kit (TOYOBO Life Science, Shanghai, China) according to the manufacturer's protocol. Quantitative reverse transcription PCR was performed to examine the mRNA levels of target genes relative to 18s rRNA or GAPDH. Primers used are listed in Table 1.
Western blot assay
Cells were lysed in an RIPA buffer (Beyotime, Jiangsu, China) in the presence of protease inhibitor and phosphatase inhibitor. Equal amounts of proteins were resolved on sodium dodecyl sulfate–polyacrylamide gel electrophoresis gels and then transferred onto polyvinylidene fluoride membranes. After blocking with 5% bovine serum albumin in Tris-buffered saline containing 0.1% Tween 20, the membranes were probed overnight at 4°C with Abs against mouse arginase 1 (Santa Cruz Biotechnology, Dallas, TX), iNOS, phosphor (p)-p65, p-STAT3 (Cell Signaling Technology, Beverly, MA), p-STAT6 (Abcam, Cambridge, MA), GAPDH (Proteintech, Wuhan, China), respectively, followed by incubation with horseradish peroxidase-conjugated secondary Abs for 1 h at room temperature. Signals were visualized by the SuperSignal West Pico Chemiluminescent Substrate (Pierce Biotechnology, Rockford, IL).
Enzyme-linked immunosorbent assay
Culture supernatants from ADSCs or macrophages were harvested for analyzing the production of cytokines using commercial enzyme-linked immunosorbent assay kits (Dakewe Biotech, Beijing, China) following the manufacturer's instructions.
Statistical analysis
Student's t-test and one-way or two-way ANOVA were performed to determine statistical significance using GraphPad Prism 5. Values are presented as the mean±standard error of the mean. Differences were considered statistically significant at P values less than 0.05.
Results
HFD induces a decrease of ADSC fraction in SVF from WAT
To determine whether obesity influences ADSC fraction in SVF, stem cell-related markers CD90 and CD105 were examined in SVF from epididymal fat pads of HFD- and ND-fed mice. Compared with ND-fed mice, HFD-fed obese mice showed a marked decrease in the expression levels of these two markers, as reflected by reduced expression intensities in both CD90 and CD105, as well as decreased expression rates in CD105 (Fig. 1A, B), indicating that HFD feeding induced a decline of ADSC fraction in SVF from WAT, which could contribute to adipocyte dysfunction and WAT inflammation in obese mice.
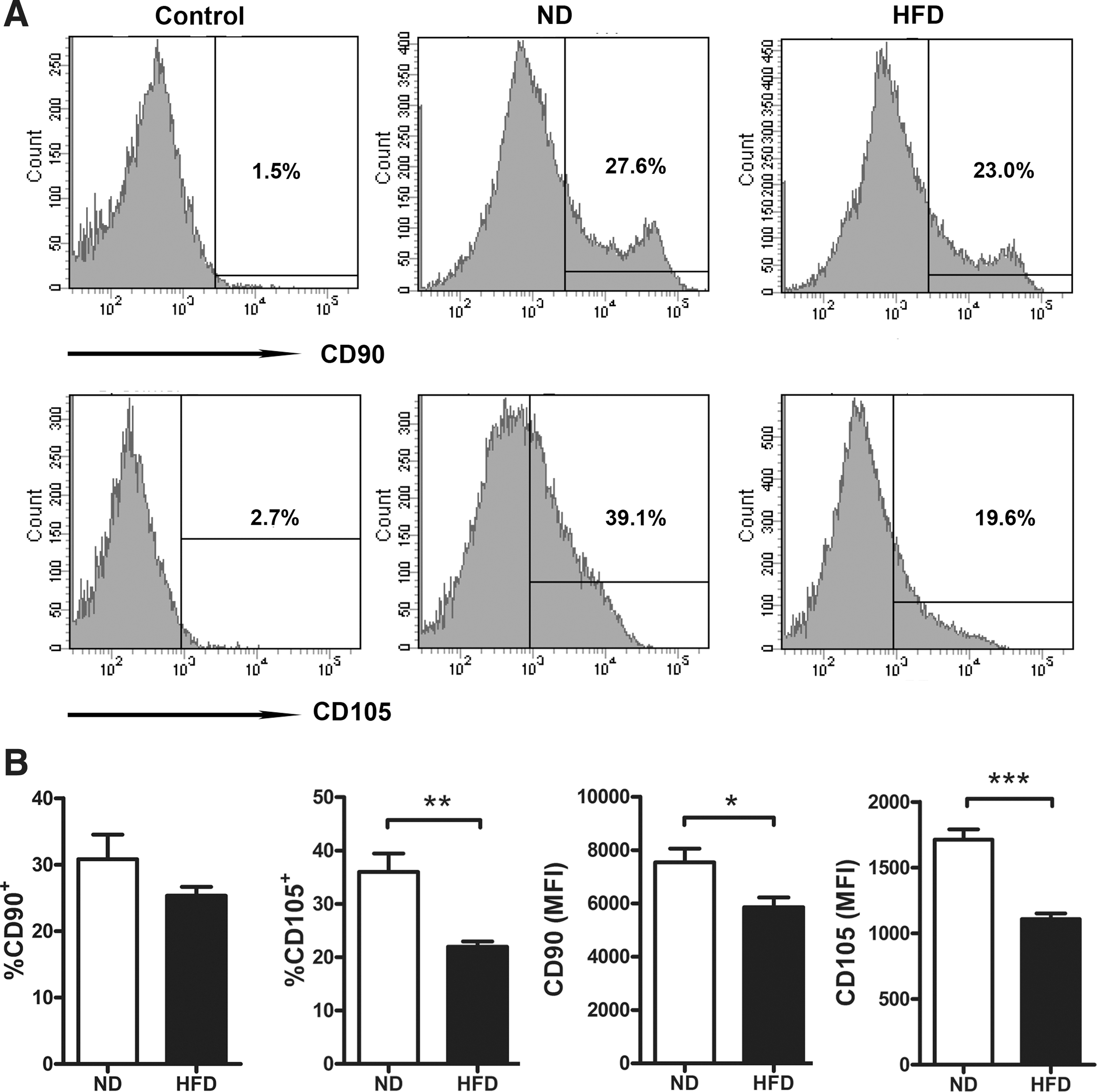
High-fat diet (HFD) reduces adipose-derived stem cell (ADSC) fraction in white adipose tissues (WAT). C57BL/6 male mice (n=8) were fed on normal diet (ND) or HFD for 14 weeks at the age of 8 weeks. Stromal vascular cell fraction (SVF) was isolated from epididymal fat pads, and expression of ADSC-related markers CD90 and CD105 was detected by flow cytometry. Representative
In vitro expansion and identification of ADSCs
Based on the immunoregulatory effects of ADSCs and their shortage in HFD-fed mice, ADSCs from ND-fed mice were isolated and expanded in vitro, for the purpose of supplementing in HFD-fed mice. In vitro cultured ADSCs exhibited classical fibroblast-like morphology and phenotypes similar to BM-MSCs, which were positive for CD90, CD105, and CD44, but negative or weakly positive for CD11c, MHC-II, and CD34 [18,30] (Supplementary Fig. S1A, B; Supplementary Data are available online at
Exogenous ADSCs successfully locate in WAT of HFD-fed mice
Next, we determined the localization of exogenous ADSCs by intraperitoneal delivery of EdU-labeled ADSCs into HFD-fed mice. Figure 2A shows that the majority of ADSCs (more than 90%) were successfully labeled with EdU after 24 h of incubation. After 3 days of delivery, fluorescence signals from EdU-labeled ADSCs were clearly detected on sections from epididymal adipose tissues, which supported the localization of exogenous ADSCs in WAT. After 7 days of delivery, more fluorescence signals were observed, probably caused by the proliferation or continuous recruitment of ADSCs (Fig. 2B). Thus, intraperitoneal delivery could successfully locate exogenous ADSCs into WAT of HFD-fed mice.

Exogenous ADSCs locate in WAT of HFD-fed mice.
ADSC treatment improves metabolic homeostasis in HFD-fed mice
After six treatments with ADSCs, plasma TG levels showed a dramatic decrease in HFD-fed mice compared with those in the NS-treated control group, whereas no detectable decline was observed in plasma TC levels (Fig. 3A, B). To clarify whether ADSC treatment also influences TG levels in ND-fed mice, we test the plasma TG in ADSC-treated mice fed on ND, while no significant change was observed compared to the NS-treated group, indicating that the TG-lowering effect of ADSCs is only elicited by HFD stimulus (Supplementary Fig. S2). Different from the slight fluctuation of glucose levels in HFD-fed mice, ADSC delivery led to a decreasing trend in nonfasting glucose levels (Fig. 3C). Of note, after four to five treatments with ADSCs, HFD-fed mice showed much more glucose tolerance and insulin sensitivity, as indicated by a significant decrease of glucose levels 15, 60 min after glucose injection (till the end of GTT) as well as 15, 45 min after insulin injection (till the end of ITT) (Fig. 3D, E). These results suggest that ADSC treatment improves lipid homeostasis and insulin sensitivity in HFD-fed mice.
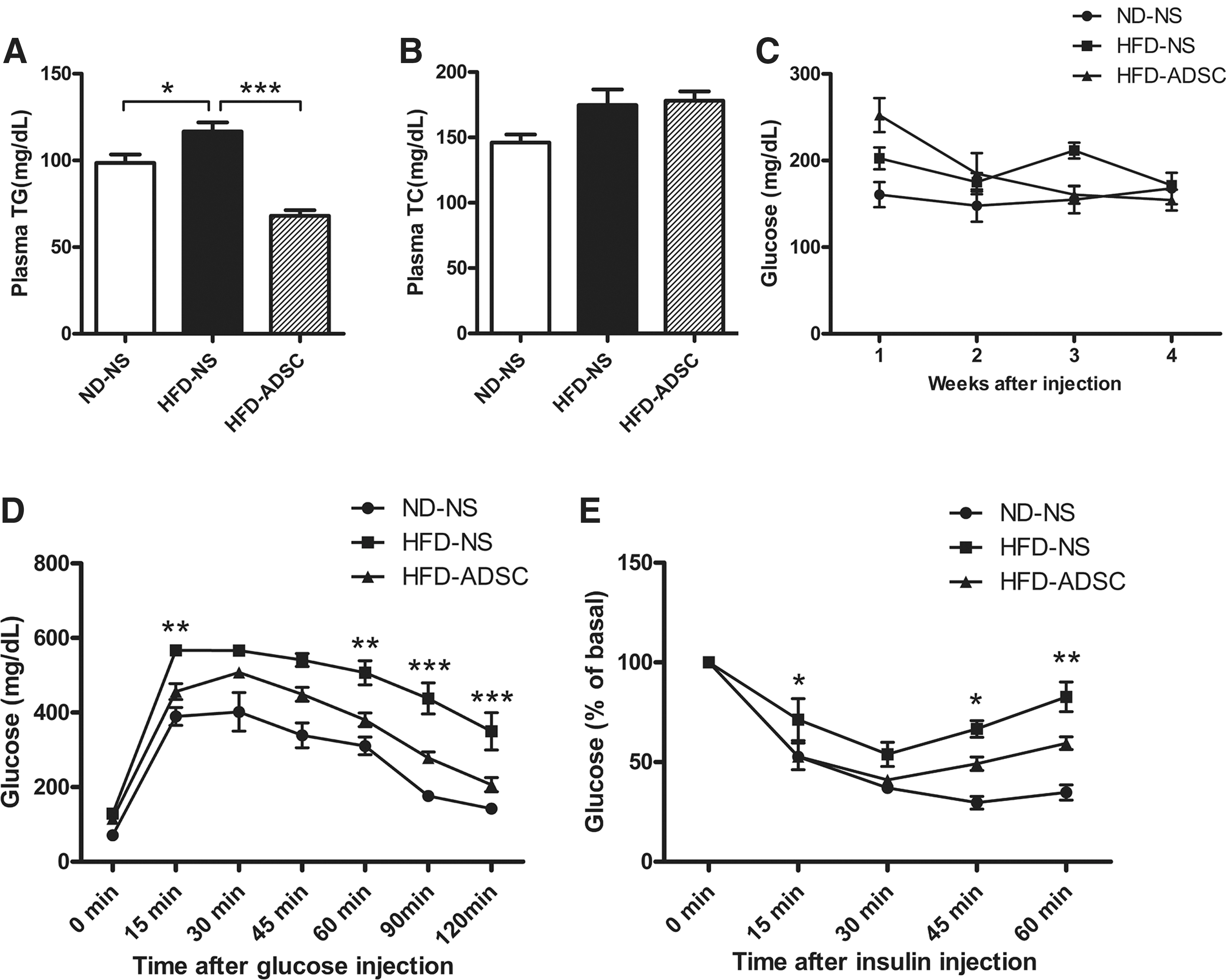
ADSC treatment improves metabolic homeostasis in HFD-fed mice. C57BL/6 male mice were fed on ND or HFD for 14 weeks, ADSC intervention was performed after 8 weeks of diet. Metabolic parameters were analyzed.
ADSC treatment attenuates adipocyte hypertrophy in HFD-fed mice
To determine whether ADSC treatment influences adiposity in mice fed on HFD, the body weight and epididymal fat mass were examined during or after ADSC delivery. Compared to ND-fed mice, HFD-fed mice showed obviously increased body weight that begins with 3 weeks of HFD intervention, whereas no significant change in body weight was observed when ADSCs were administrated to these HFD-fed mice (Fig. 4A, B). Accordingly, no significant differences were detected in the epididymal fat mass between ADSC-treated and NS-treated HFD-fed mice (Fig. 4C).
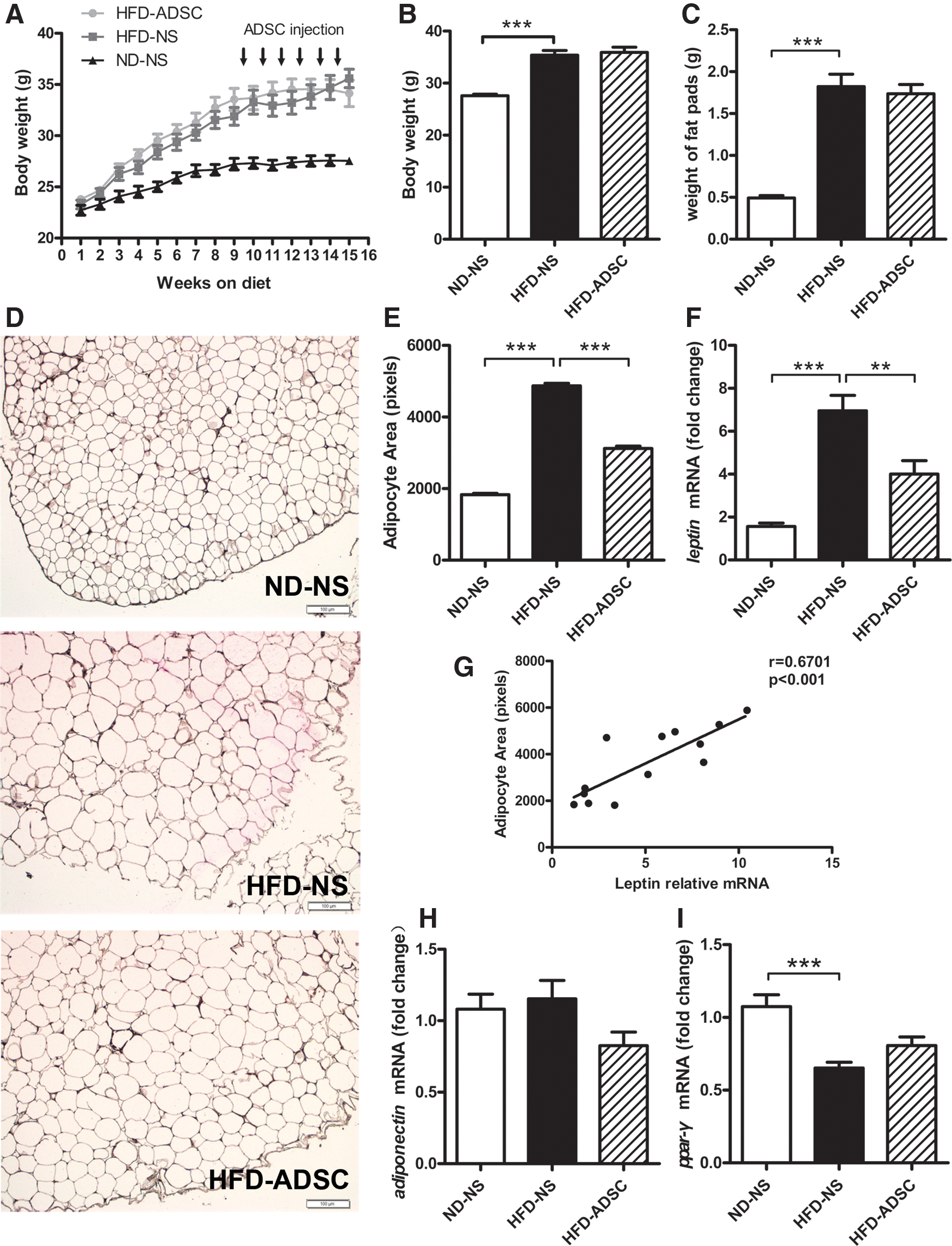
Effects of ADSC treatment on adiposity in HFD-fed mice. C57BL/6 male mice were fed on ND or HFD for 14 weeks, and ADSC intervention was performed after 8 weeks of diet.
Interestingly, ADSC treatment led to a marked reduction of adipocyte hypertrophy in WAT from HFD-fed mice, as evidenced by a clearly visible decrease in sizes of adipocytes from epididymal adipose tissues (Fig. 4D, E). Although adipocyte hypertrophy with varying degrees still remained in ADSC-treated HFD-fed mice, much smaller adipocytes were found in ADSC-treated other than NS-treated HFD-fed mice (Fig. 4D). Consistently, ADSC treatment for HFD-fed mice caused a dramatic decrease in mRNA levels of leptin, which had a closely positive correlation with the adipocyte sizes (Fig. 4F, G). ADSC treatment also led to an increase, but not significantly, in mRNA levels of peroxisome proliferator-activated receptor gamma (PPAR-γ). No significant change was found in mRNA levels of adiponectin (Fig. 4H, I). Moreover, ADSC treatment in HFD-fed mice could not increase the mRNA levels of key enzymes for lipolysis, including adipose triglyceride lipase and hormone-sensitive lipase, indicating that lipolysis may not be a direct contributor to the reduced adipocyte hypertrophy. Conversely, some degree of decline in these two genes was observed after ADSC treatment (Supplementary Fig. S3), possibly due to the relatively small adipocyte size and enrichment of ADSCs in adipose tissues, which were partially consistent with previous studies [31,32]. These data indicate that ADSC treatment attenuates adipocyte hypertrophy and helps to maintain adipocyte homeostasis in HFD-fed mice.
ADSC treatment ameliorates WAT inflammation in HFD-fed mice
Since WAT inflammation contributes to metabolic disorders in the obese state, we further evaluated WAT inflammation by examination of macrophage infiltration and inflammatory cytokine release from epididymal adipose tissues. As shown in Fig. 5A, numerous CD68+ macrophages displaying classical crown-like structures, which were observed in WAT from HFD-fed obese mice, were dramatically reduced by ADSC delivery. Importantly, ADSC treatment for HFD-fed mice remarkably reduced the secretion of proinflammatory cytokine TNF-α, but enhanced that of anti-inflammatory cytokine IL-10 from cultured adipose explants. A significant elevation, but relatively low levels, of IFN-γ was also found in ADSC-treated HFD-fed mice. No detectable differences were obtained in the levels of IL-6, IL-12, and monocyte chemoattractant protein-1 (MCP-1) (Fig. 5B). In addition, ADSC treatment in HFD-fed mice showed decreasing trends in serum inflammatory cytokines, including IL-6, TNF-α, IFN-γ, IL-12, and MCP-1, whereas no significant differences were observed (Supplementary Fig. S4). These data demonstrate that ADSC treatment ameliorates WAT inflammation in HFD-fed mice.
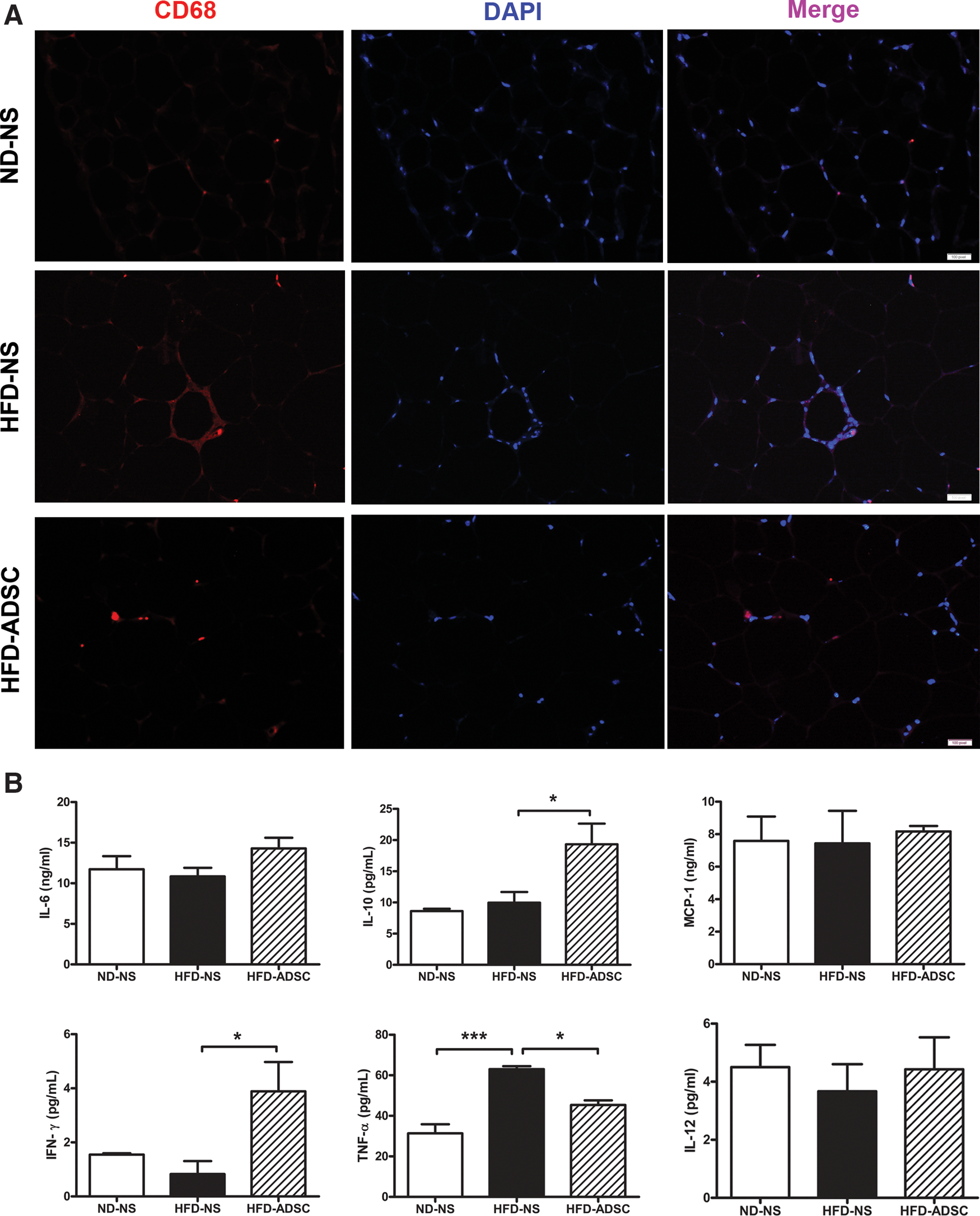
ADSC treatment attenuates WAT inflammation in HFD-fed mice. C57BL/6 male mice were fed on ND or HFD for 14 weeks, and ADSC intervention was performed after 8 weeks of diet. The macrophage infiltration in epididymal adipose tissues and inflammatory cytokine secretion by epididymal adipose tissue explants were detected.
ADSC treatment remodels the macrophage phenotypes in WAT from HFD-fed mice
Considering that macrophage polarization contributes to WAT homeostasis or inflammation, we further evaluated M1- or M2-related markers on SVF to verify the roles of macrophages. After 6 weeks of ADSC treatment, the mRNA levels of proinflammatory TNF-α and IL-12, which were higher in HFD-fed mice than those in ND-fed mice, decreased significantly; in contrast, the mRNA levels of anti-inflammatory IL-10 substantially increased (Fig. 6A). Accordingly, the elevation of the iNOS/arginase 1 ratio caused by HFD feeding was abolished by ADSC administration (Fig. 6B). Finally, Flow cytometry data substantiated that ADSC delivery effectively triggered the production of IL-10 from adipose macrophages and, surprisingly, also dramatically inhibited the upregulation of MHC-II-expressing macrophages in HFD-fed mice (Fig. 6C–E). Thus, ADSC treatment for HFD-fed mice reprograms the phenotypes of adipose macrophages from proinflammatory M1 to an anti-inflammatory M2-like subtype.
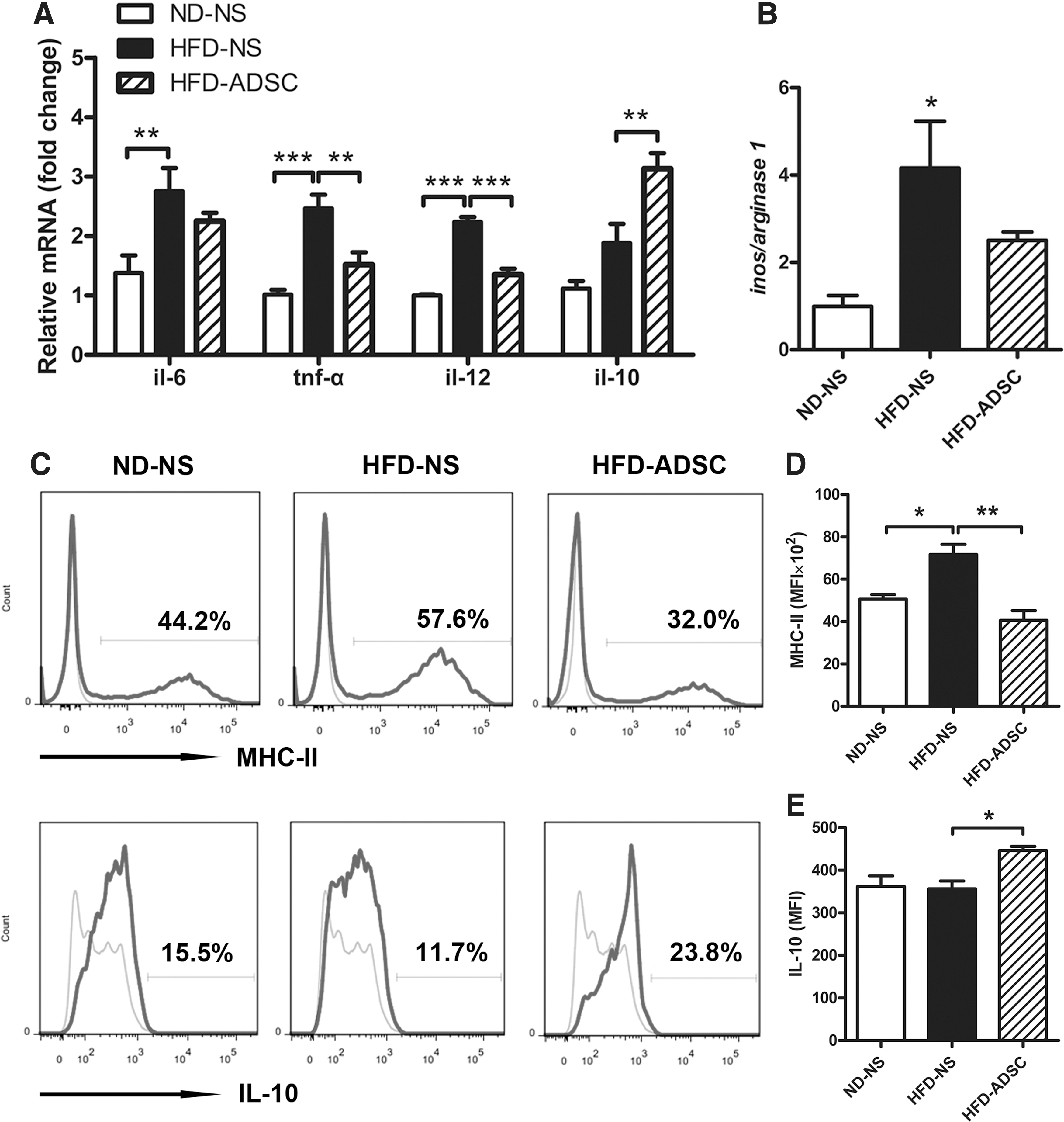
ADSC treatment remodels the macrophage phenotypes in WAT from HFD-fed mice. C57BL/6 male mice were fed ND or HFD for 14 weeks, and ADSCs were delivered after 8 weeks of diet (n=4–5 mice/group). SVF was isolated from epididymal fat pads, and the expression of macrophage phenotype-related genes was examined.
ADSCs screw the polarization of macrophages in vitro
To clarify the direct role of ADSCs in shifting the M1/M2 balance, macrophages induced by LPS and IFN-γ were cocultured with ADSCs through transwell or with the ADSC conditional medium. As expected from in vivo experiments, the conditional medium from ADSCs evidently inhibited M1 polarization by decreasing mRNA levels of TNF-α and IL-12, but increasing that of IL-10 (Fig. 7A). In particular, enhanced IL-10 production together with reduced MHC-II expression was also reproduced in ADSC-educated macrophages (Fig. 7B), substantiating the capacity of ADSCs for screwing polarized macrophages toward the M2-like subtype. The increase of arginase 1 and decrease of iNOS, which confirmed the phenotypic switch from M1 to M2 subtypes, were verified in ADSC-educated macrophages. Importantly, these macrophages showed remarkably enhanced p-STAT3 and p-STAT6, accompanied by reduced p-p65, indicating that this switch is mediated by activation of STAT3/STAT6 and inhibition of the NF-κB pathway (Fig. 7C and Supplementary Fig. S5).
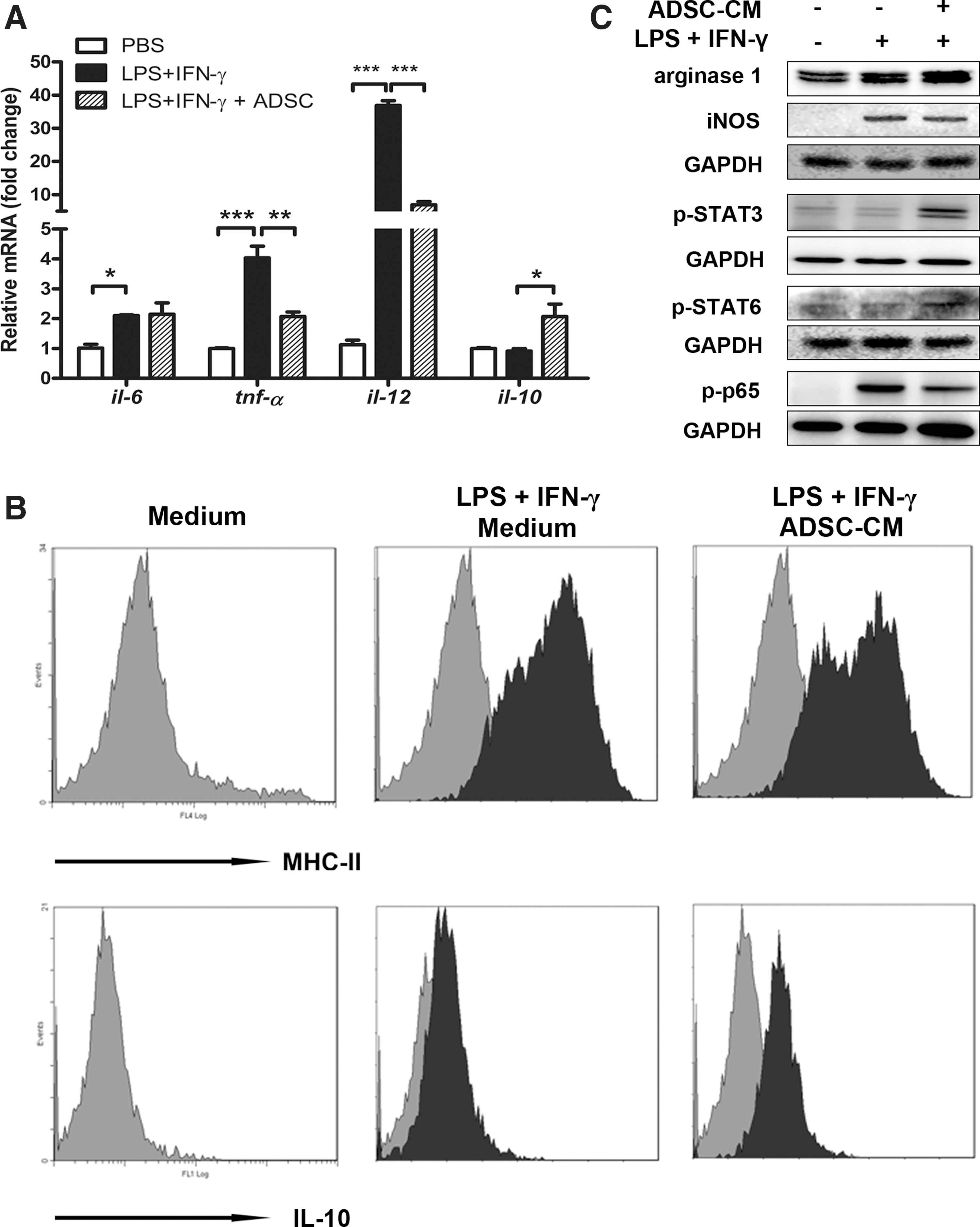
ADSCs reprogram macrophage polarization in vitro. Peritoneal macrophages from C57BL/6 male mice were cocultured through transwell or ADSC-conditioned medium (ADSC-CM) in the presence of 20 ng/mL interferon-gamma (IFN-γ) and 100 ng/mL lipopolysaccharide (LPS), using unstimulated macrophages as control. Cells were collected after 6 h (for western blot assay of p-STAT3), 24 h (for detection of mRNA levels or western blot assay), or 48 h of incubation (for flow cytometry).
Discussion
Transplantation of ADSCs as regenerative cells has been successful in some animal models, which has led to the ongoing clinical trials in patients, such as heart infarction, Crohn's fistulae, and lipodystrophies [33,34]. Recent studies on therapy for arthritis, colitis, and type 1 diabetes underlined the immunoregulatory nature of ADSCs [21,22], thus promoting us to explore their role in obesity-associated inflammation. We have confirmed the shortage of ADSCs in WAT from HFD-fed mice in the current study, as indicated by decreased expression of stem cell-related markers CD105 and CD90. The scarcity of preadipocytes accompanied by overload nutrient definitely drive the hypertrophy and lipolysis of adipocytes, which can induce severe WAT inflammation and insulin resistance. Therefore, we sought to resolve adipocyte hypertrophy and WAT inflammation through the ADSC supplement, which was supposed to combine the advantages of ADSCs in both regenerative and anti-inflammatory effects.
Few studies have delivered ADSCs into obese mice, probably because of the worry about exacerbated adiposity and inflammation. Previous study implanting 3T3-L1 preadipocytes into the visceral mesenteric site indeed led to the adverse metabolic effects in nude mice [35]. However, in the present study, we have shown the beneficial effects of ADSC treatment for HFD-fed mice, as suggested by improved insulin action and metabolic homeostasis. Despite no resistance to obesity, ADSC treatment significantly reduced adipocyte hypertrophy and leptin mRNA levels in WAT (Fig. 4D–F). This change is an important contributor to improved insulin sensitivity as previously reported [36]. Previous reports have suggested that adipocyte size per se may be the critical regulator of leptin mRNA expression [37 –39]. In the present study, we also showed strong correlation of leptin mRNA levels with adipocyte sizes in WAT (Fig. 4G), which meant that the decrease in leptin mRNA levels might be caused by reduced adipocyte sizes in ADSC-treated mice. As for adiponectin mRNA levels, no significant variation was observed after ADSC treatment in HFD-fed mice, even between ND- and HFD-fed mice (Fig. 4H). Since no significant difference was shown in adiponectin mRNA levels of adipocytes with different sizes, there seems no direct association between adiponectin mRNA expression and adipocyte hypertrophy [40]. While in the current experimental settings many regulating factors like PPAR-γ, TNF-α, IL-6, and HFD could produce comprehensive influence and bring uncertainty on the mRNA expression of adiponectin [41,42], the underlying complex mechanisms remain to be investigated. Nevertheless, ADSC treatment helps to maintain adipose homeostasis by inhibiting adipocyte hypertrophy, thereby favoring the beneficial metabolism in obese mice. In addition, we also tested the effects of single-round injection of ADSCs in HFD-fed obese mice. Unfortunately, no significant improvement was observed on insulin resistance, adiposity, adipocyte hypertrophy, and adipose inflammation (Supplementary Fig. S6A–H), indicating the insufficiency of single-round injection to reverse the deleterious effects induced by obesity.
We demonstrate that ADSC delivery into obese mice efficiently reduces WAT inflammation by decreasing TNF-α, but increasing IL-10 secretion from WAT explants. Several lines of evidences reveal the contribution of adipose-resident macrophages to this alteration in ADSC-treated obese mice. First, less crown-like structures of macrophages were detected in WAT of these mice compared with that of the control obese mice. Second, and more importantly, similar to WAT explants, decreased TNF-α together with increased IL-10 mRNA levels was observed in SVF from WAT, which can exclude the interference from adipocyte-derived TNF-α. Last, and most importantly, decreased MHC class II-expressing but increased IL-10-producing macrophages were detected in WAT, providing direct evidence that novel M2-like phenotypes were reprogrammed through ADSC delivery.
A very recent report emphasizes the essential role of MHC-II-expressing adipocytes in obesity-induced adipose inflammation [43]. In this study, our observations indicate the involvement of MHC-II-expressing macrophages in adipose inflammation of obese mice. More interestingly, ADSC delivery into obese mice dramatically restricted the elevation of MHC-II-expressing macrophages, while triggering the increase of IL-10-producing macrophages in WAT. Several studies have demonstrated that IL-10 can inhibit the deleterious effects of proinflammatory cytokines on insulin signaling, thus increasing insulin sensitivity [12,13,44 –46]. Adipose regulatory T or B cells have been identified to produce IL-10 in very recent studies, which plays a pivotal role in maintaining adipose tissue homeostasis [47,48]. Our findings add another cell source that produces IL-10, M2-like macrophages, on the list of adipose tissue homeostasis and insulin action.
Furthermore, we reveal the novel capacity of ADSCs in remodeling M2-like macrophages in obese mice, which is partially dependent on soluble factors from ADSCs and activation of STAT3/STAT6 in macrophages. The activation of both STAT3 and STAT6 by cytokines like IL-6, IL-10, IL-4, and IL-13, or growth factors like FGF and vascular endothelial growth factor (VEGF) is involved in the transcription of M2-related gene expression, including arginase 1 and IL-10 [9,49 –52]. Therefore, our data showing enhanced STAT3/STAT6 activation could be attributed to two possible mechanisms. First, ADSC-derived VEGF and/or FGF directly activate STAT6; second, abundant IL-6 released from ADSCs, just as shown in Supplementary Fig. S7, drives the activation of STAT3 and then primes the activation of STAT6, this action mode has been clearly described in recent reports [49,50]. In addition, we cannot rule out other factors and pathways that regulate M1/M2 switch; the cross talk of ADSC and macrophage remains to be further investigated.
Adipose eosinophils are recognized as the main cells expressing IL-4/IL-13 and sustaining the M2-like macrophage polarization state in WAT [12,53]. In the present study, we propose ADSCs as another independent contributor to the maintenance of M2-like macrophages in WAT of lean mice. Obesity induces the scarcity of ADSCs accompanied by M1 polarization, while delivery of ADSCs efficiently reverses this process. Therefore, ADSC delivery into obese mice effectively resolves WAT inflammation and insulin resistance, thus suggesting the critical role of ADSCs in maintaining adipose homeostasis and their promising potency for the treatment of obesity-related diseases.
Footnotes
Acknowledgments
This study was supported by the National Natural Science Foundation of China (81471065, 81270923) and the National “973” program of China (2011CB503906). Our fluorescence imaging work was performed at the Microscopy Characterization Facility, Shandong University. We acknowledge Huaizhu Wu, Department of Medicine, Baylor College of Medicine, USA, and Chunmei Liu, Department of Clinical Laboratory, Shandong Provincial Hospital Affiliated to Shandong University, China, for providing technical assistance for this study.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.